
Long-Term Patterns of Amphibian Diversity, Abundance and Nutrient Export from Small, Isolated Wetlands


Abstract
:1. Introduction
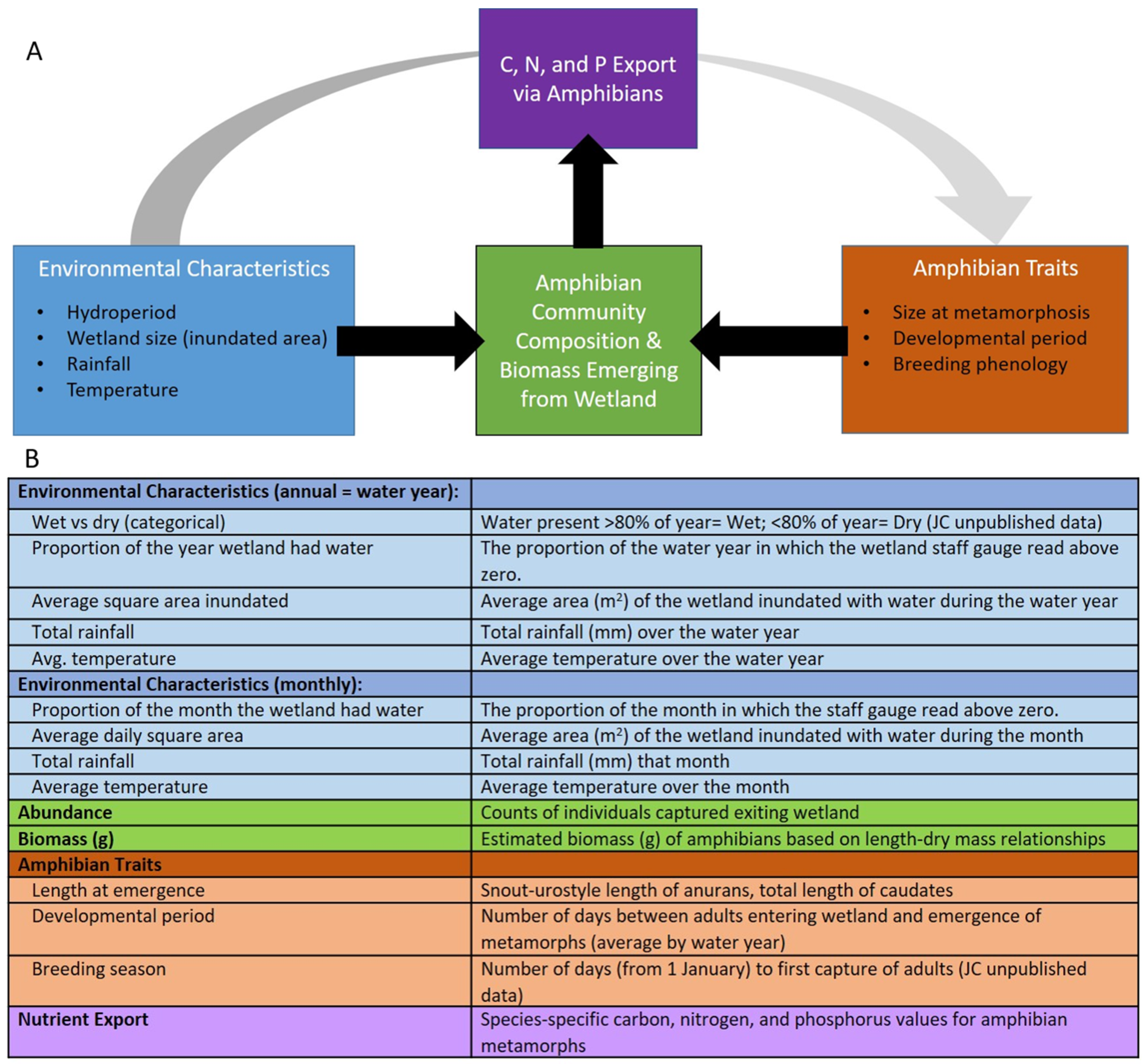
2. Materials and Methods
2.1. Study Site
2.2. Temperature, Rainfall and Wetland Hydrology
2.3. Amphibian Sampling Methods
2.4. Biomass, Nutrient Flux and Species Traits
3. Data Analyses
4. Results
4.1. Environmental Conditions
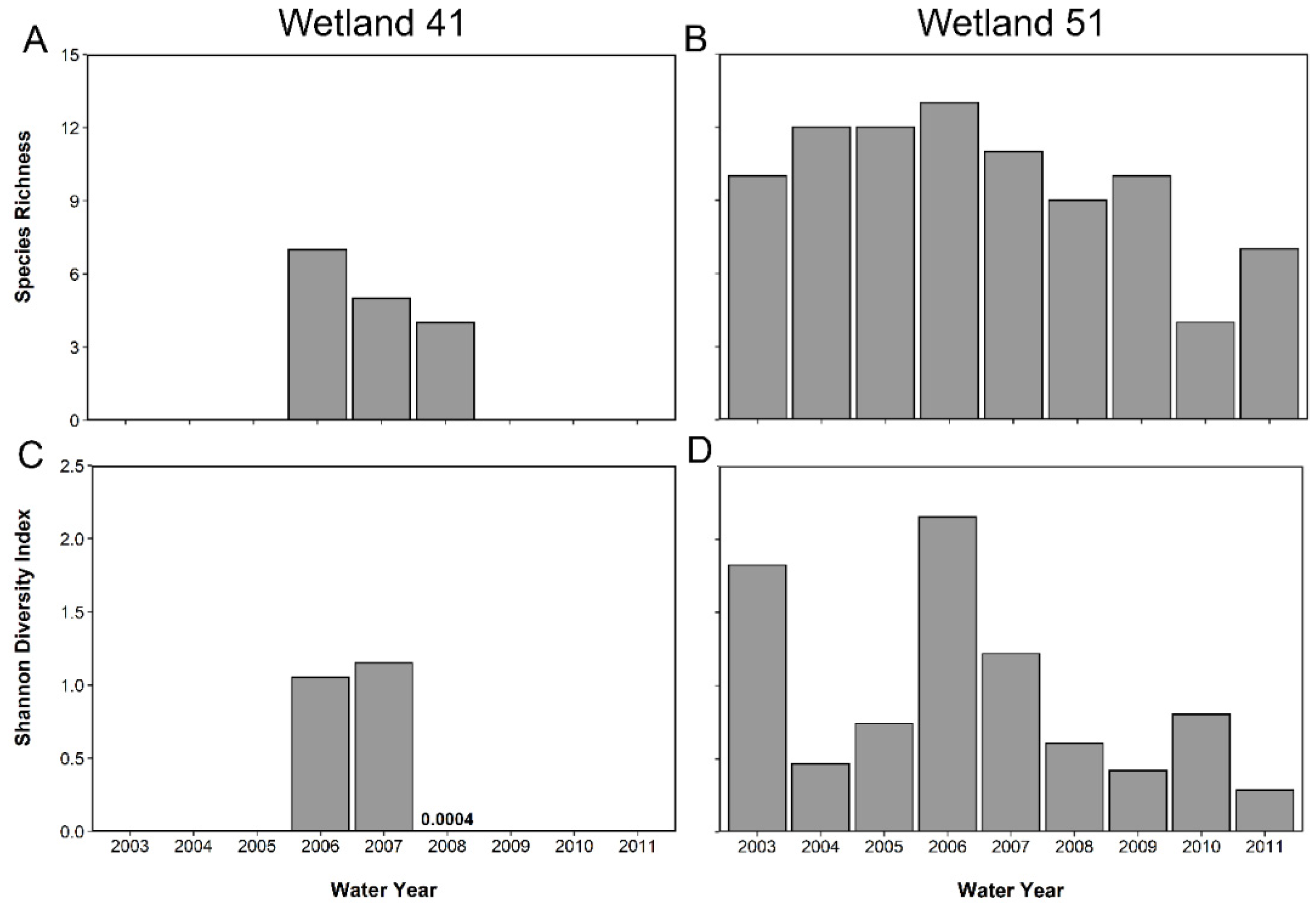
4.2. Amphibian Diversity and Abundance
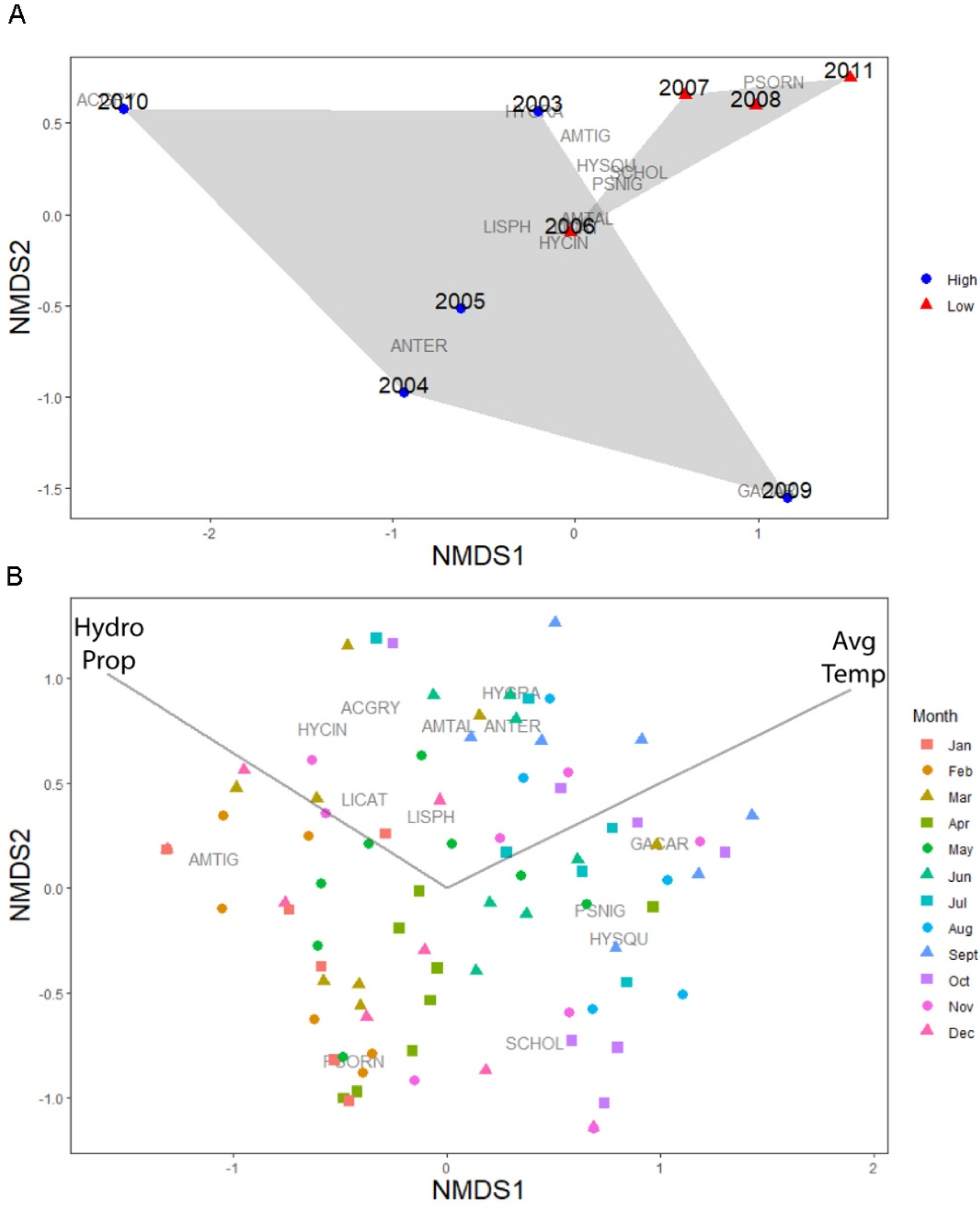
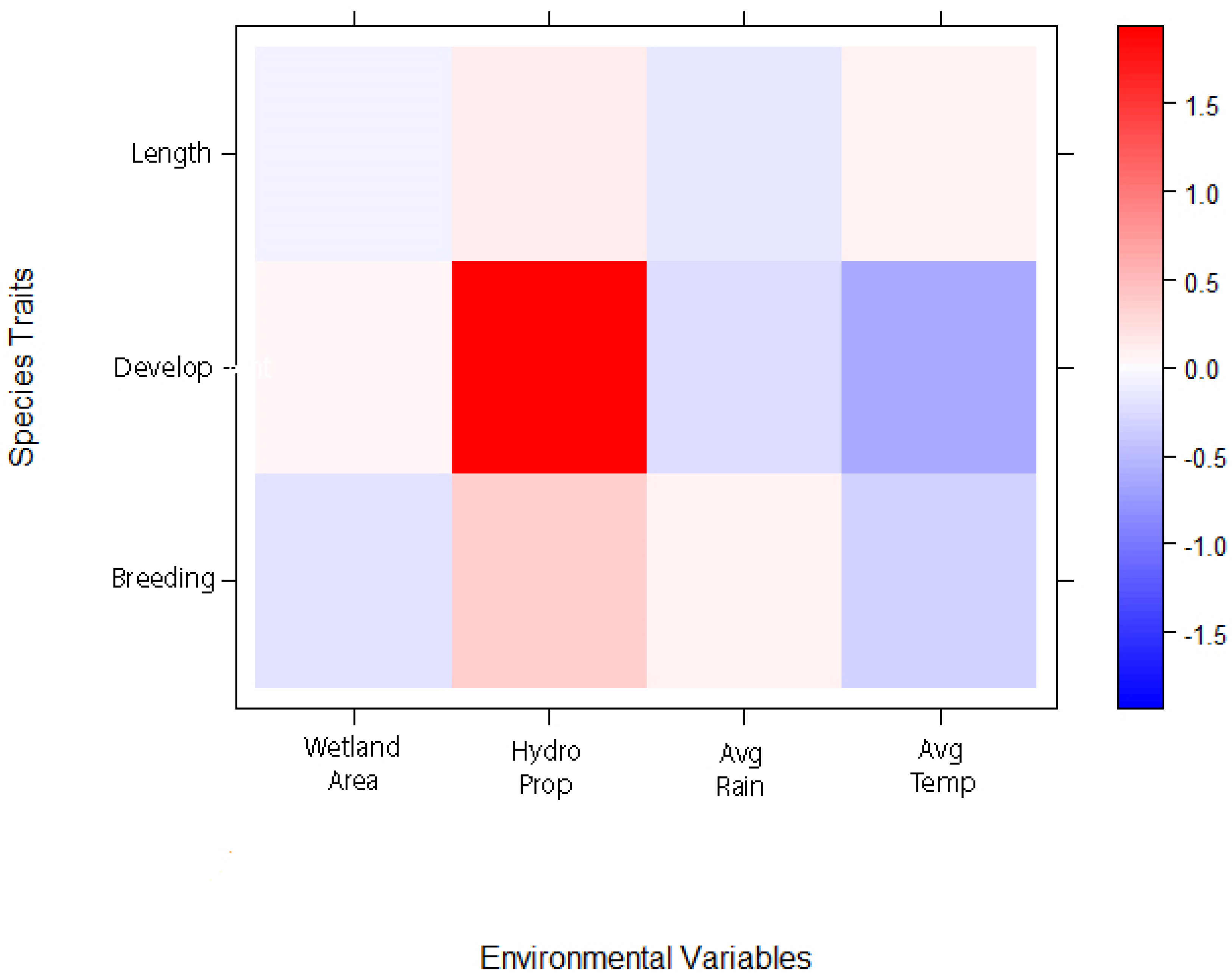
4.3. Environmental Drivers of Amphibian Community Composition at W51
5. Discussion
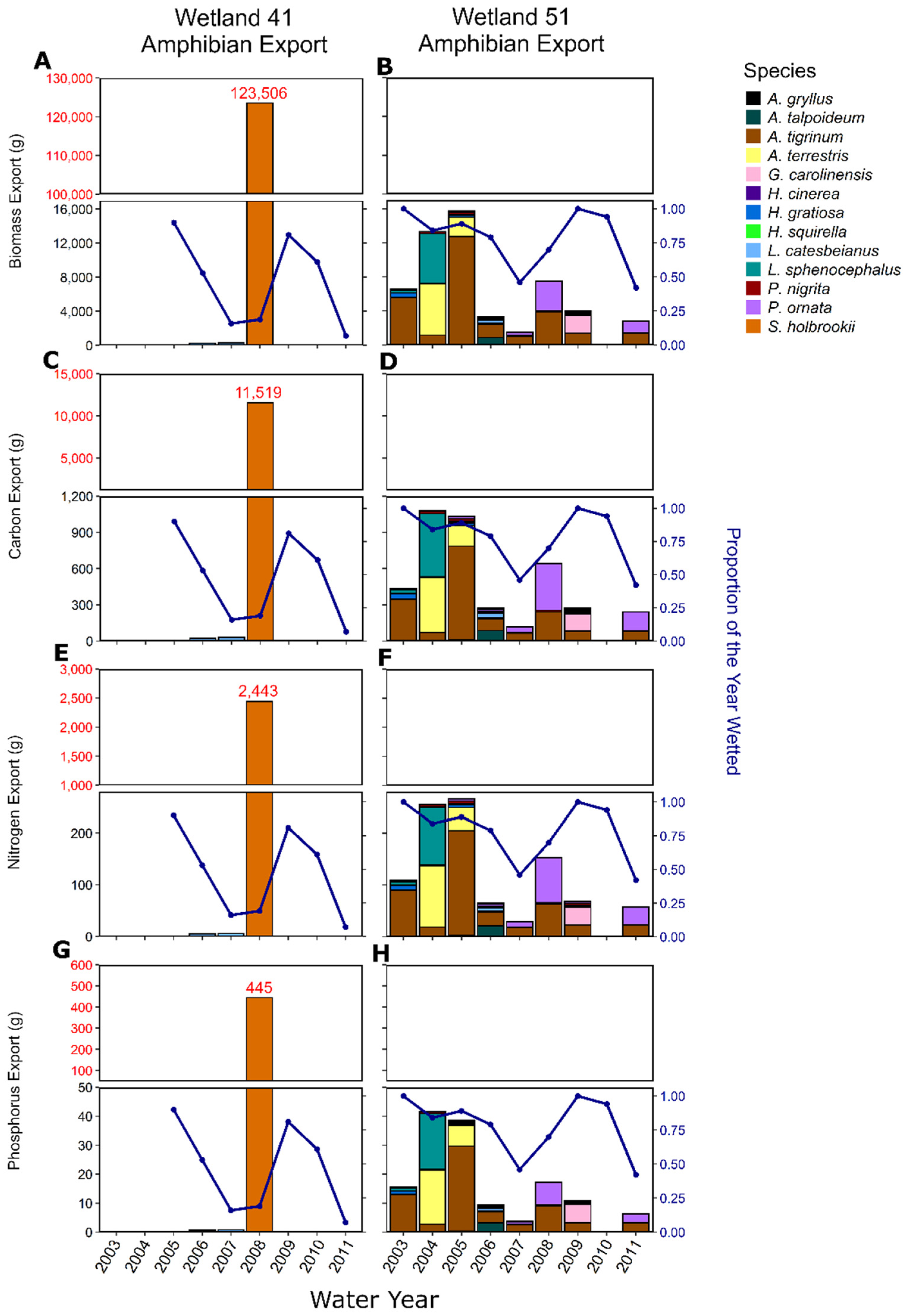
5.1. Amphibian Species Diversity and Biomass Export
5.2. Amphibian Nutrient Export
5.3. Hydroperiod and Temperature as Environmental Controls on Amphibian Emergence
5.4. Future Changes in Land Use and Climate—Consequences for Amphibian Diversity and Export
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Leibowitz, S.G. Isolated wetlands and their functions: An ecological perspective. Wetlands 2003, 23, 517–531. [Google Scholar] [CrossRef]
- Kirkman, L.K.; Smith, L.L.; Quintana-Ascencio, P.F.; Kaeser, M.J.; Golladay, S.W.; Farmer, A.L. Is species richness congruent among taxa? Surrogacy, complementarity, and environmental correlates among three disparate taxa in geographically isolated wetlands. Ecol. Indic. 2012, 18, 131–139. [Google Scholar] [CrossRef]
- EPA. Connectivity of Streams and Wetlands to Downstream Waters: A Review and Synthesis of the Scientific Evidence; EPA/600/R-14/475F; US Environmental Protection Agency: Washington, DC, USA, 2015.
- Cohen, M.J.; Creed, I.F.; Alexander, L.; Basu, N.B.; Calhoun, A.J.; Craft, C.; D’Amico, E.; DeKeyser, E.; Fowler, L.; Golden, H.E. Do geographically isolated wetlands influence landscape functions? Proc. Natl. Acad. Sci. USA 2016, 113, 1978–1986. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L.; Subalusky, A.L.; Atkinson, C.L.; Kirkman, L.K. Ecology and Restoration of the Longleaf Pine Ecosystem. In Geographically Isolated Wetlands: Embedded Habitats in Longleaf Pine Forests; Kirkman, L.K., Jack, S.B., Eds.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar]
- Snodgrass, J.W.; Komoroski, M.J.; Bryan, A.L.; Burger, J. Relationships among isolated wetland size, hydroperiod, and amphibian species richness: Implications for wetland regulations. Conserv. Biol. 2000, 14, 414–419. [Google Scholar] [CrossRef]
- Griffiths, R.A. Temporary ponds as amphibian habitats. Aquat. Conserv. Mar. Freshw. Ecosyst. 1997, 7, 119–126. [Google Scholar] [CrossRef]
- Battle, J.; Golladay, S.W. Water quality and macroinvertebrate assemblages in three types of seasonally inundated limesink wetlands in southwest Georgia. J. Freshw. Ecol. 2001, 16, 189–207. [Google Scholar] [CrossRef] [Green Version]
- Leeper, D.A.; Taylor, B.E. Abundance, biomass and production of aquatic invertebrates in Rainbow Bay, a temporary wetland in South Carolina, USA. Archiv Hydrobiol. 1998, 143, 335–362. [Google Scholar] [CrossRef]
- Stubbington, R.; Datry, T. The macroinvertebrate seedbank promotes community persistence in temporary rivers across climate zones. Freshw. Biol. 2013, 58, 1202–1220. [Google Scholar] [CrossRef] [Green Version]
- Subalusky, A.L.; Fitzgerald, L.A.; Smith, L.L. Ontogenetic niche shifts in the American Alligator establish functional connectivity between aquatic systems. Biol. Conserv. 2009, 142, 1507–1514. [Google Scholar] [CrossRef]
- Williams, D.D.; Hynes, H.N. The ecology of temporary streams I. The faunas of two Canadian streams. Int. Rev. Der Gesamten Hydrobiol. Hydrogr. 1976, 61, 761–787. [Google Scholar] [CrossRef] [Green Version]
- Schofield, K.A.; Alexander, L.C.; Ridley, C.E.; Vanderhoof, M.K.; Fritz, K.M.; Autrey, B.C.; DeMeester, J.E.; Kepner, W.G.; Lane, C.R.; Leibowitz, S.G.; et al. Biota Connect Aquatic Habitats Throughout Freshwater Ecosystem Mosaics. JAWRA J. Am. Water Resour. Assoc. 2018, 54, 372–399. [Google Scholar] [CrossRef]
- Fritz, K.A.; Whiles, M.R. Amphibian-mediated nutrient fluxes across aquatic–terrestrial boundaries of temporary wetlands. Freshw. Biol. 2018, 63, 1250–1259. [Google Scholar] [CrossRef]
- Fritz, K.A.; Whiles, M.R. Reciprocal subsidies between temporary ponds and riparian forests. Limnol. Oceanogr. 2021, 66, 3149–3161. [Google Scholar] [CrossRef]
- Hannan, L.B.; Roth, J.D.; Ehrhart, L.M.; Weishampel, J.F. Dune vegetation fertilization by nesting sea turtles. Ecology 2007, 88, 1053–1058. [Google Scholar] [CrossRef]
- Holtgrieve, G.W.; Schindler, D.E. Marine-derived nutrients, bioturbation, and ecosystem metabolism: Reconsidering the role of salmon in streams. Ecology 2011, 92, 373–385. [Google Scholar] [CrossRef]
- Booth, M.T.; Hairston Jr, N.G.; Flecker, A.S. Consumer movement dynamics as hidden drivers of stream habitat structure: Suckers as ecosystem engineers on the night shift. Oikos 2020, 129, 194–208. [Google Scholar] [CrossRef]
- Seale, D.B. Influence of amphibian larvae on primary production, nutrient flux, and competition in a pond ecosystem. Ecology 1980, 61, 1531–1550. [Google Scholar] [CrossRef]
- Moore, J.W.; Schindler, D.E.; Carter, J.L.; Fox, J.; Griffiths, J.; Holtgrieve, G.W. Biotic control of stream fluxes: Spawning salmon drive nutrient and matter export. Ecology 2007, 88, 1278–1291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regester, K.J.; Lips, K.R.; Whiles, M.R. Energy flow and subsidies associated with the complex life cycle of ambystomatid salamanders in ponds and adjacent forest in southern Illinois. Oecologia 2006, 147, 303–314. [Google Scholar] [CrossRef] [PubMed]
- Capps, K.A.; Berven, K.A.; Tiegs, S.D. Modelling nutrient transport and transformation by pool-breeding amphibians in forested landscapes using a 21-year dataset. Freshw. Biol. 2015, 60, 500–511. [Google Scholar] [CrossRef]
- Sitters, J.; Atkinson, C.L.; Guelzow, N.; Kelly, P.; Sullivan, L.L. Spatial stoichiometry: Cross-ecosystem material flows and their impact on recipient ecosystems and organisms. Oikos 2015, 124, 920–930. [Google Scholar] [CrossRef]
- Polis, G.A.; Anderson, W.B.; Holt, R.D. Toward an integration of landscape and food web ecology: The dynamics of spatially subsidized food webs. Annu. Rev. Ecol. Syst. 1997, 28, 289–316. [Google Scholar] [CrossRef] [Green Version]
- Leroux, S.J.; Loreau, M. Subsidy hypothesis and strength of trophic cascades across ecosystems. Ecol. Lett. 2008, 11, 1147–1156. [Google Scholar] [CrossRef] [PubMed]
- Gibbons, J.W.; Winne, C.T.; Scott, D.E.; Willson, J.D.; Glaudas, X.; Andrews, K.M.; Todd, B.D.; Fedewa, L.A.; Wilkinson, L.; Tsaliagos, R.N.; et al. Remarkable amphibian biomass and abundance in an isolated wetland: Implications for wetland conservation. Conserv. Biol. 2006, 20, 1457–1465. [Google Scholar] [CrossRef]
- Lannoo, M. Amphibian Declines; University of California Press: Berkeley, CA, USA, 2005; p. 2040. [Google Scholar]
- Pechmann, J.H.; Scott, D.E.; Gibbons, J.W.; Semlitsch, R.D. Influence of wetland hydroperiod on diversity and abundance of metamorphosing juvenile amphibians. Wetl. Ecol. Manag. 1989, 1, 3–11. [Google Scholar] [CrossRef]
- Altig, R.; McDiarmid, R.W. Handbook of Larval Amphibians of the United States and Canada; Cornell University Press: Ithaca, NY, USA, 2015. [Google Scholar]
- Knapp, D.D.; Smith, L.L.; Atkinson, C.L. Larval anurans follow predictions of stoichiometric theory: Implications for nutrient storage in wetlands. Ecosphere 2021, 12, e03466. [Google Scholar] [CrossRef]
- Golladay, S.; Clayton, B.; Brantley, S.; Smith, C.; Qi, J.; Hicks, D. Forest restoration increases isolated wetland hydroperiod: A long-term case study. Ecosphere 2021, 12, e03495. [Google Scholar] [CrossRef]
- Greenberg, C.H.; Neary, D.G.; Harris, L.D. A comparison of herpetofaunal sampling effectiveness of pitfall, single-ended, and double-ended funnel traps used with drift fences. J. Herpetol. 1994, 28, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Gibbons, J.W.; Semlitsch, R.D. Survivorship and longevity of a long-lived vertebrate species: How long do turtles live? J. Anim. Ecol. 1982, 51, 523–527. [Google Scholar] [CrossRef]
- Jensen, J.B. Amphibians and Reptiles of Georgia; University of Georgia Press: Athens, GA, USA, 2008. [Google Scholar]
- Wright, A.H. Life-Histories of the Frogs of Okefinokee Swamp, Georgia: North American Salientia (Anura) No. 2.; Macmillan Company: New York, NY, USA, 1932. [Google Scholar]
- Castaneda, L.E.; Sabat, P.; Gonzalez, S.P.; Nespolo, R.F. Digestive plasticity in tadpoles of the Chilean giant frog (Caudiverbera caudiverbera): Factorial effects of diet and temperature. Physiol. Biochem. Zool. 2006, 79, 919–926. [Google Scholar] [CrossRef] [Green Version]
- Luhring, T.M.; DeLong, J.P.; Semlitsch, R.D. Stoichiometry and life-history interact to determine the magnitude of cross-ecosystem element and biomass fluxes. Front. Microbiol. 2017, 8, 814. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Brown, A.M.; Warton, D.I.; Andrew, N.R.; Binns, M.; Cassis, G.; Gibb, H. The fourth-corner solution–using predictive models to understand how species traits interact with the environment. Methods Ecol. Evol. 2014, 5, 344–352. [Google Scholar] [CrossRef]
- Legendre, P.; Galzin, R.; Harmelin-Vivien, M.L. Relating behavior to habitat: Solutions to the fourth-corner problem. Ecology 1997, 78, 547–562. [Google Scholar] [CrossRef]
- Wang, Y.; Naumann, U.; Wright, S.T.; Warton, D.I. mvabund–an R package for model-based analysis of multivariate abundance data. Methods Ecol. Evol. 2012, 3, 471–474. [Google Scholar] [CrossRef]
- De Steven, D.; Toner, M.M. Vegetation of upper coastal plain depression wetlands: Environmental templates and wetland dynamics within a landscape framework. Wetlands 2004, 24, 23–42. [Google Scholar] [CrossRef] [Green Version]
- Kirkman, L.K.; Goebel, P.C.; West, L.; Drew, M.B.; Palik, B.J. Depressional wetland vegetation types: A question of plant community development. Wetlands 2000, 20, 373–385. [Google Scholar] [CrossRef]
- Martin, G.I.; Kirkman, L.K.; Hepinstall-Cymerman, J. Mapping geographically isolated wetlands in the Dougherty Plain, Georgia, USA. Wetlands 2012, 32, 149–160. [Google Scholar] [CrossRef]
- Burton, T.M.; Likens, G.E. Salamander populations and biomass in the Hubbard Brook experimental forest, New Hampshire. Copeia 1975, 1975, 541–546. [Google Scholar] [CrossRef]
- Davic, R.D.; Welsh, H.H. On the Ecological Roles of Salamanders. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 405–434. [Google Scholar] [CrossRef] [Green Version]
- Watt, K.M.; Golladay, S.W. Organic matter dynamics in seasonally inundated, forested wetlands of the gulf coastal plain. Wetlands 1999, 19, 139–148. [Google Scholar] [CrossRef]
- Atkinson, C.L.; Capps, K.A.; Rugenski, A.T.; Vanni, M.J. Consumer-driven nutrient dynamics in freshwater ecosystems: From individuals to ecosystems. Biol. Rev. 2017, 92, 2003–2023. [Google Scholar] [CrossRef] [PubMed]
- Balik, J.A.; Jameson, E.E.; Wissinger, S.A.; Whiteman, H.H.; Taylor, B.W. Animal-Driven Nutrient Supply Declines Relative to Ecosystem Nutrient Demand Along a Pond Hydroperiod Gradient. Ecosystems 2021. [Google Scholar] [CrossRef]
- Matthews, K.R.; Knapp, R.A.; Pope, K.L. Garter snake distributions in high-elevation aquatic ecosystems: Is there a link with declining amphibian populations and nonnative trout introductions? J. Herpetol. 2002, 36, 16–22. [Google Scholar] [CrossRef]
- Gray, L.J. Response of insectivorous birds to emerging aquatic insects in riparian habitats of a tallgrass prairie stream. Am. Midl. Nat. 1993, 129, 288–300. [Google Scholar] [CrossRef]
- Miller, D.A.W.; Grant, E.H.C.; Muths, E.; Amburgey, S.M.; Adams, M.J.; Joseph, M.B.; Waddle, J.H.; Johnson, P.T.J.; Ryan, M.E.; Schmidt, B.R.; et al. Quantifying climate sensitivity and climate-driven change in North American amphibian communities. Nat. Commun. 2018, 9, 3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semlitsch, R.D. Principles for management of aquatic-breeding amphibians. J. Wildl. Manag. 2000, 64, 615–631. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. MECHANISMS CREATING COMMUNITY STRUCTURE ACROSS A FRESHWATER HABITAT GRADIENT. Annu. Rev. Ecol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef] [Green Version]
- Holomuzki, J.R.; Collins, J.P.; Brunkow, P.E. Trophic control of fishless ponds by tiger salamander larvae. Oikos 1994, 55–64. [Google Scholar] [CrossRef]
- Semlitsch, R.D.; Bodie, J.R. Are small, isolated wetlands expendable? Conserv. Biol. 1998, 12, 1129–1133. [Google Scholar] [CrossRef] [Green Version]
- Napton, D.E.; Auch, R.F.; Headley, R.; Taylor, J.L. Land changes and their driving forces in the Southeastern United States. Reg. Environ. Chang. 2010, 10, 37–53. [Google Scholar] [CrossRef]
- Martin, G.I.; Hepinstall-Cymerman, J.; Kirkman, L.K. Six decades (1948–2007) of landscape change in the Dougherty Plain of Southwest Georgia, USA. Southeast. Geogr. 2013, 53, 28–49. [Google Scholar] [CrossRef]
- Atkinson, C.L.; Golladay, S.W.; First, M.R. Water quality and planktonic microbial assemblages of isolated wetlands in an agricultural landscape. Wetlands 2011, 31, 885–894. [Google Scholar] [CrossRef]
- Scheele, B.C.; Skerratt, L.F.; Grogan, L.F.; Hunter, D.A.; Clemann, N.; McFadden, M.; Newell, D.; Hoskin, C.J.; Gillespie, G.R.; Heard, G.W. After the epidemic: Ongoing declines, stabilizations and recoveries in amphibians afflicted by chytridiomycosis. Biol. Conserv. 2017, 206, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Smith, L.L.; Subalusky, A.L.; Atkinson, C.L.; Earl, J.E.; Mushet, D.M.; Scott, D.E.; Lance, S.L.; Johnson, S.A. Biological Connectivity of Seasonally Ponded Wetlands across Spatial and Temporal Scales. JAWRA J. Am. Water Resour. Assoc. 2019, 55, 334–353. [Google Scholar] [CrossRef]
- Blaustein, A.R.; Walls, S.C.; Bancroft, B.A.; Lawler, J.J.; Searle, C.L.; Gervasi, S.S. Direct and indirect effects of climate change on amphibian populations. Diversity 2010, 2, 281–313. [Google Scholar] [CrossRef]
- Lips, K.R.; Brem, F.; Brenes, R.; Reeve, J.D.; Alford, R.A.; Voyles, J.; Carey, C.; Livo, L.; Pessier, A.P.; Collins, J.P. Emerging infectious disease and the loss of biodiversity in a Neotropical amphibian community. Proc. Natl. Acad. Sci. USA 2006, 103, 3165–3170. [Google Scholar] [CrossRef] [Green Version]
- Hof, C.; Araújo, M.B.; Jetz, W.; Rahbek, C. Additive threats from pathogens, climate and land-use change for global amphibian diversity. Nature 2011, 480, 516–519. [Google Scholar] [CrossRef]
- Whiles, M.R.; Hall, R.O.; Dodds, W.K.; Verburg, P.; Huryn, A.D.; Pringle, C.M.; Lips, K.R.; Kilham, S.S.; Colon-Gaud, C.; Rugenski, A.T.; et al. Disease-driven amphibian declines alter ecosystem processes in a tropical stream. Ecosystems 2013, 16, 146–157. [Google Scholar] [CrossRef]
- Rantala, H.M.; Nelson, A.M.; Fulgoni, J.N.; Whiles, M.R.; Hall Jr, R.O.; Dodds, W.K.; Verburg, P.; Huryn, A.D.; Pringle, C.M.; Kilham, S.S. Long-term changes in structure and function of a tropical headwater stream following a disease-driven amphibian decline. Freshw. Biol. 2015, 60, 575–589. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water Year | Average Temperature (°C) | Total Rainfall (mm) | W51 Average Wetted Square Area (m2) | W51 Proportion of the Year Wetted | W51 Proportion Year Full |
|---|---|---|---|---|---|
| 2003 | 18.06 | 1521.99 | 7691.53 | 1 | 0.79 |
| 2004 | 18.19 | 1107.22 | 5131.68 | 0.84 | 0.63 |
| 2005 | 18.6 | 1513.31 | 6344.92 | 0.89 | 0.72 |
| 2006 | 18.86 | 1205.74 | 2028.96 | 0.79 | 0.31 |
| 2007 | 18.84 | 973.08 | 1351.12 | 0.46 | 0.21 |
| 2008 | 18.91 | 1282.19 | 2195.3 | 0.70 | 0.30 |
| 2009 | 18.62 | 1546.3 | 5114.34 | 1 | 0.63 |
| 2010 | 18.51 | 1303.45 | 5064.35 | 0.94 | 0.6 |
| 2011 | 18.6 | 920.26 | 1812.5 | 0.42 | 0.27 |
| Wetland | Species | Species Code | Total Count | DM (g) | C (g) | N (g) | P (g) |
|---|---|---|---|---|---|---|---|
| 41 | Ambystoma tigrinum | AMTIG | 1 | 2.0809 | 0.9552 | 0.2494 | 0.0361 |
| Anaxyrus terrestris | ANTER | 60 | 2.6869 | 1.0025 | 0.2614 | 0.0414 | |
| Gastrophryne carolinensis | GACAR | 249 | 6.7277 | 2.7425 | 0.6887 | 0.1243 | |
| Hyla cinerea | HYCIN | 3 | 0.7417 | 0.3455 | 0.0802 | 0.0092 | |
| Hyla squirella | HYSQU | 8 | 0.9189 | 0.4095 | 0.1015 | 0.0141 | |
| Lithobates catesbeianus | LICAT | 32 | 96.4978 | 48.5005 | 9.2246 | 1.1576 | |
| Lithobates sphenocephalus | LISPH | 8 | 4.0203 | 1.8747 | 0.4008 | 0.0694 | |
| Pseudacris nigrita | PSNIG | 1 | 0.0579 | 0.0269 | 0.0061 | 0.0007 | |
| Pseudacris ornata | PSORN | 1 | 0.1474 | 0.0703 | 0.0158 | 0.0014 | |
| Scaphiopus holbrookii | SCHOL | 310,443 | 26,110.8979 | 11,520.0421 | 2443.0225 | 445.4137 | |
| Totals | 310,806 | 26,224.78 | 11,575.97 | 2454.05 | 446.87 | ||
| 51 | Acris gryllus | ACGRY | 305 | 11.1323 | 5.1339 | 1.1346 | 0.1358 |
| Ambystoma talpoideum | AMTAL | 161 | 222.3275 | 105.2983 | 26.3022 | 3.9663 | |
| Ambystoma tigrinum | AMTIG | 1829 | 3805.9662 | 1747.0118 | 456.2059 | 66.0520 | |
| Anaxyrus terrestris | ANTER | 37,824 | 1693.7992 | 631.9565 | 164.8067 | 26.0879 | |
| Gastrophryne carolinensis | GACAR | 13,146 | 355.1911 | 144.7885 | 36.3590 | 6.5647 | |
| Hyla cinerea | HYCIN | 3 | 0.7417 | 0.3455 | 0.0802 | 0.0092 | |
| Hyla gratiosa | HYGRA | 503 | 169.8054 | 81.8459 | 16.2930 | 1.9173 | |
| Hyla squirella | HYSQU | 42 | 4.8242 | 2.1499 | 0.5329 | 0.0741 | |
| Lithobates catesbeianus | LICAT | 26 | 78.4045 | 39.4067 | 7.4950 | 0.9405 | |
| Lithobates sphenocephalus | LISPH | 2520 | 1266.3803 | 590.5454 | 126.2644 | 21.8464 | |
| Pseudacris nigrita | PSNIG | 3216 | 186.1234 | 86.4802 | 19.4608 | 2.1156 | |
| Pseudacris ornata | PSORN | 9034 | 1331.8570 | 635.4914 | 143.0331 | 12.5425 | |
| Scaphiopus holbrookii | SCHOL | 819 | 68.8849 | 30.3918 | 6.4451 | 1.1751 | |
| Totals | 69,428 | 9195.44 | 4100.85 | 1004.41 | 143.43 |
| Wetland | Water Year | Count (ind m−2 y −1) | DM (g m−2 y−1) | Carbon (mg C m−2 y−1) | Nitrogen (mg N m−2 y−1) | Phosphorus (mg P m−2 y−1) |
|---|---|---|---|---|---|---|
| 41 | 2006 | 0.30 | 0.05 | 25.13 | 5.11 | 0.715 |
| 2007 | 0.08 | 0.06 | 31.74 | 6.11 | 0.777 | |
| 2008 | 310.42 | 26.11 | 11519.10 | 2442.83 | 445.376 | |
| Avg | 103.60 | 8.74 | 3858.66 | 818.02 | 148.960 | |
| StDev | 179.11 | 15.04 | 6634.14 | 1407.13 | 256.710 | |
| 51 | 2003 | 0.16 | 0.13 | 436.07 | 108.97 | 15.572 |
| 2004 | 4.18 | 0.35 | 1077.66 | 255.23 | 41.498 | |
| 2005 | 1.71 | 0.32 | 1032.92 | 266.01 | 38.699 | |
| 2006 | 0.24 | 0.08 | 272.59 | 65.69 | 9.441 | |
| 2007 | 0.14 | 0.04 | 119.73 | 29.11 | 3.689 | |
| 2008 | 0.87 | 0.19 | 642.28 | 153.19 | 17.189 | |
| 2009 | 1.88 | 0.09 | 274.94 | 68.21 | 10.902 | |
| 2010 | 0.01 | 0.00 | 1.91 | 0.42 | 0.060 | |
| 2011 | 0.32 | 0.07 | 242.75 | 57.58 | 6.378 | |
| Avg | 1.06 | 0.14 | 455.65 | 111.60 | 15.940 | |
| StDev | 1.36 | 0.12 | 384.96 | 95.05 | 14.720 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Atkinson, C.L.; Knapp, D.D.; Smith, L.L. Long-Term Patterns of Amphibian Diversity, Abundance and Nutrient Export from Small, Isolated Wetlands. Diversity 2021, 13, 598. https://doi.org/10.3390/d13110598
Atkinson CL, Knapp DD, Smith LL. Long-Term Patterns of Amphibian Diversity, Abundance and Nutrient Export from Small, Isolated Wetlands. Diversity. 2021; 13(11):598. https://doi.org/10.3390/d13110598
Chicago/Turabian StyleAtkinson, Carla L., Daniel D. Knapp, and Lora L. Smith. 2021. "Long-Term Patterns of Amphibian Diversity, Abundance and Nutrient Export from Small, Isolated Wetlands" Diversity 13, no. 11: 598. https://doi.org/10.3390/d13110598
APA StyleAtkinson, C. L., Knapp, D. D., & Smith, L. L. (2021). Long-Term Patterns of Amphibian Diversity, Abundance and Nutrient Export from Small, Isolated Wetlands. Diversity, 13(11), 598. https://doi.org/10.3390/d13110598