
Biological Interaction as a Possible Ultimate Driver in the Local Extinction of Cedrus atlantica in the Iberian Peninsula

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
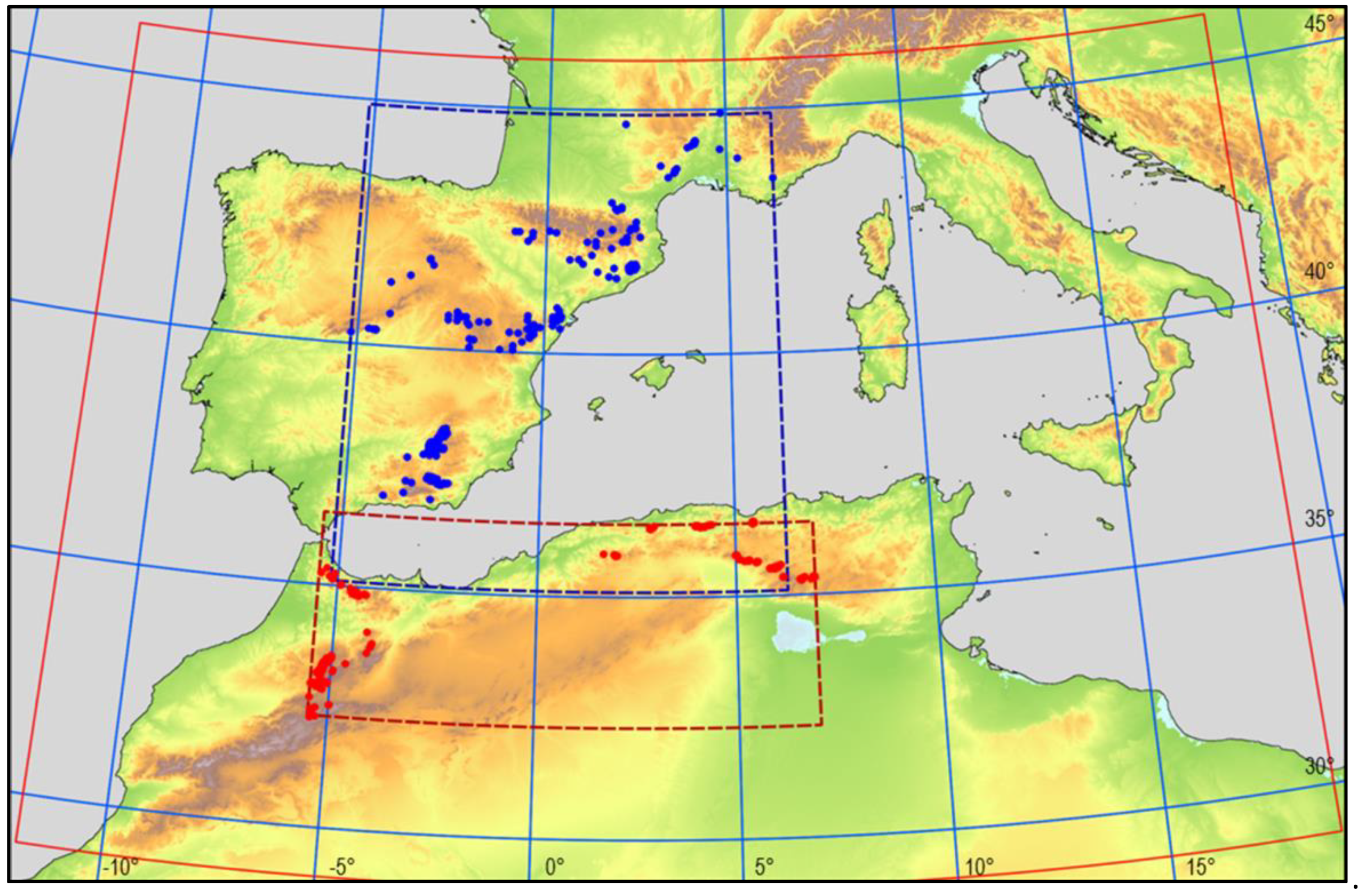
2.1. Study Area
2.2. Species
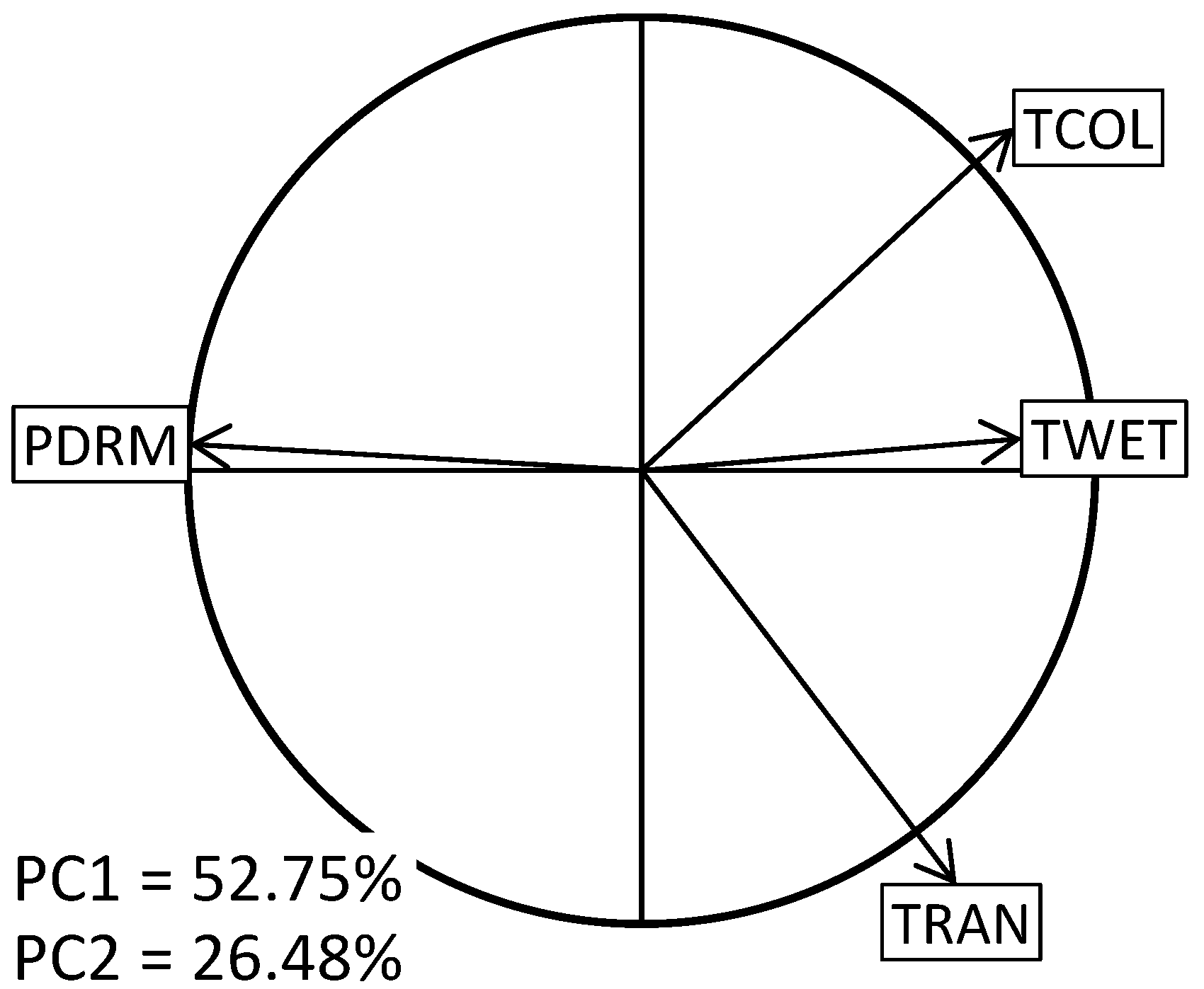
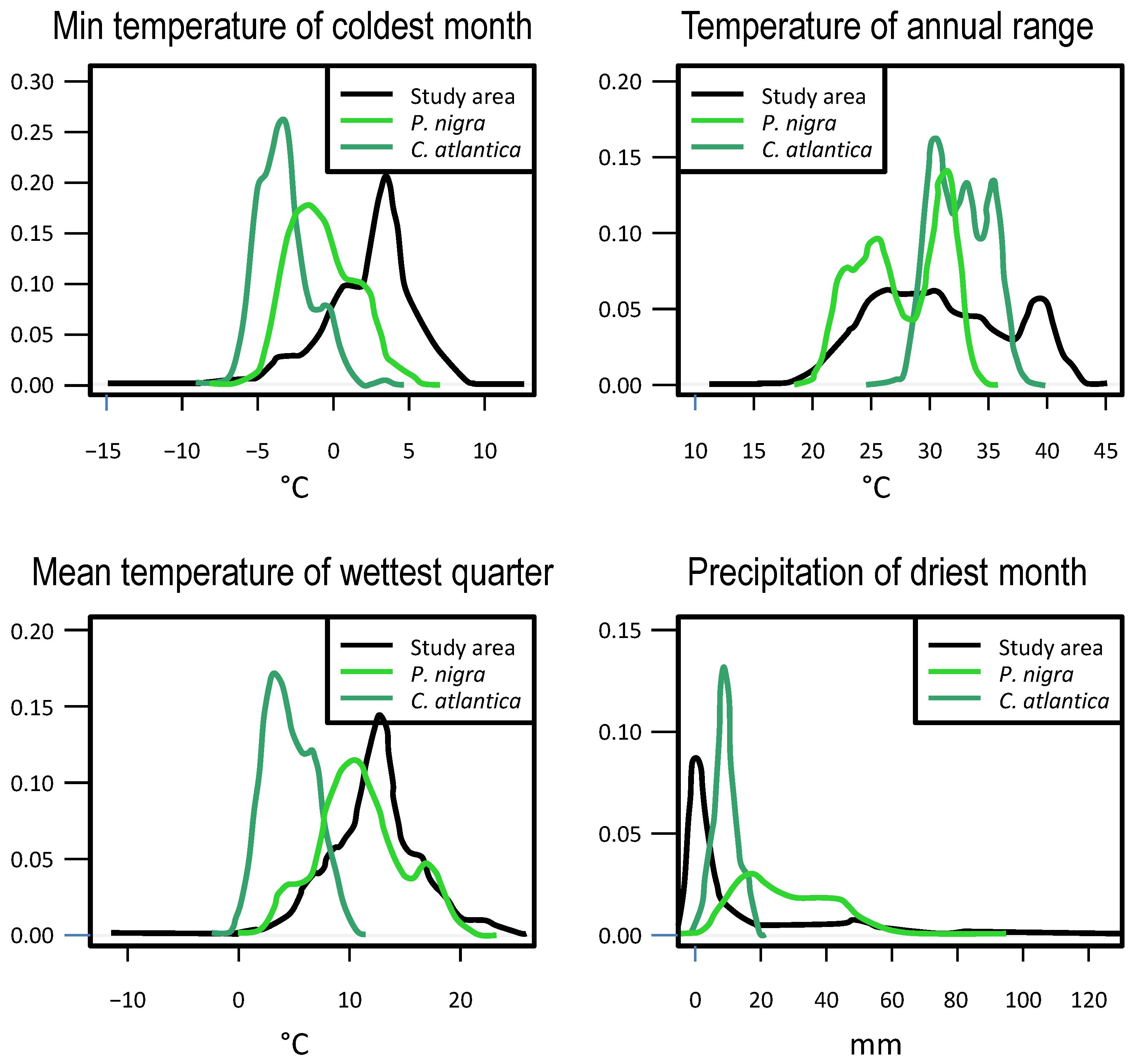
2.3. Variables
- Minimum temperature of the coldest month (bio6);
- Temperature annual range (bio7);
- Mean temperature of the wettest quarter (bio8);
- Precipitation of the driest month (bio14).
2.4. Analysis
3. Results
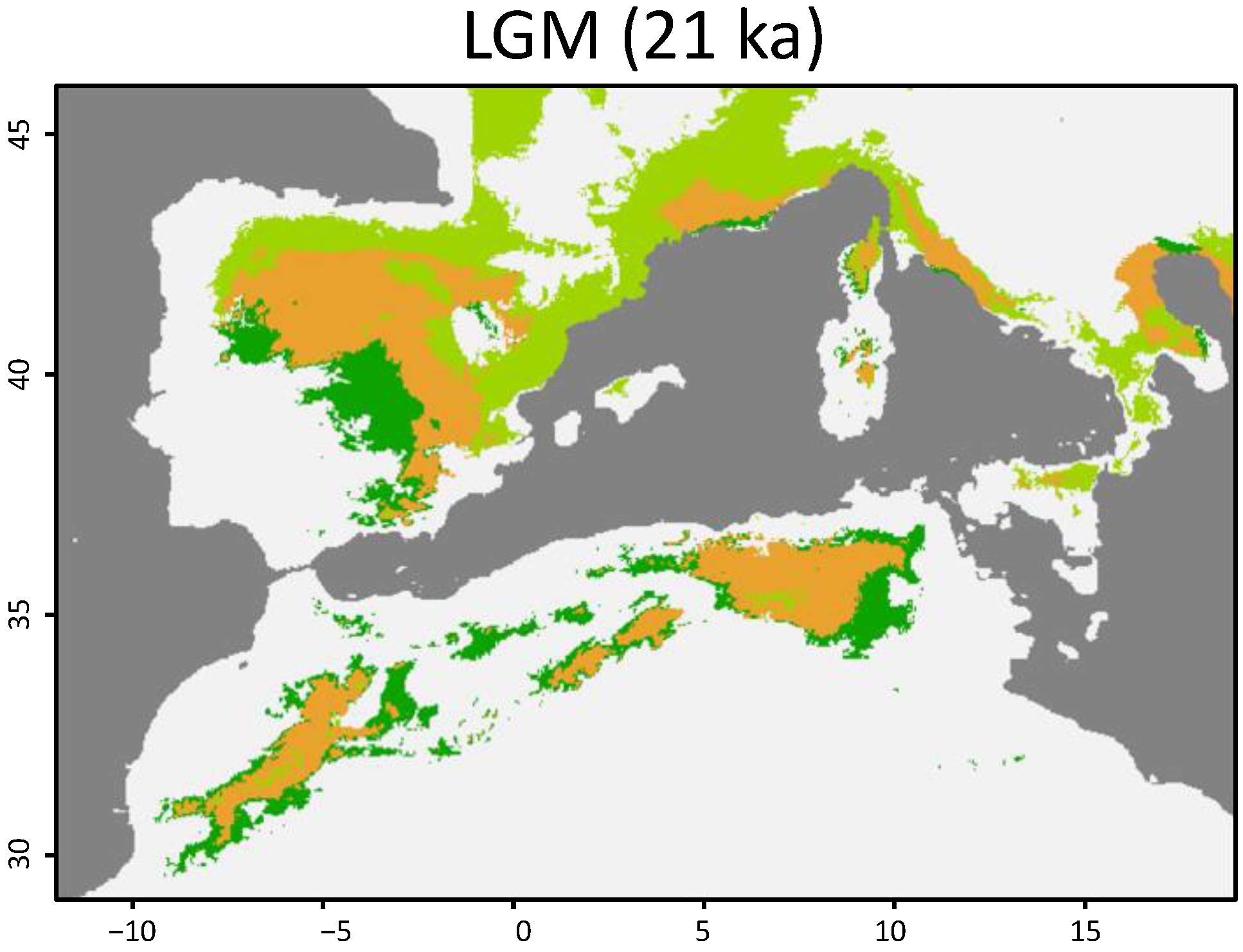
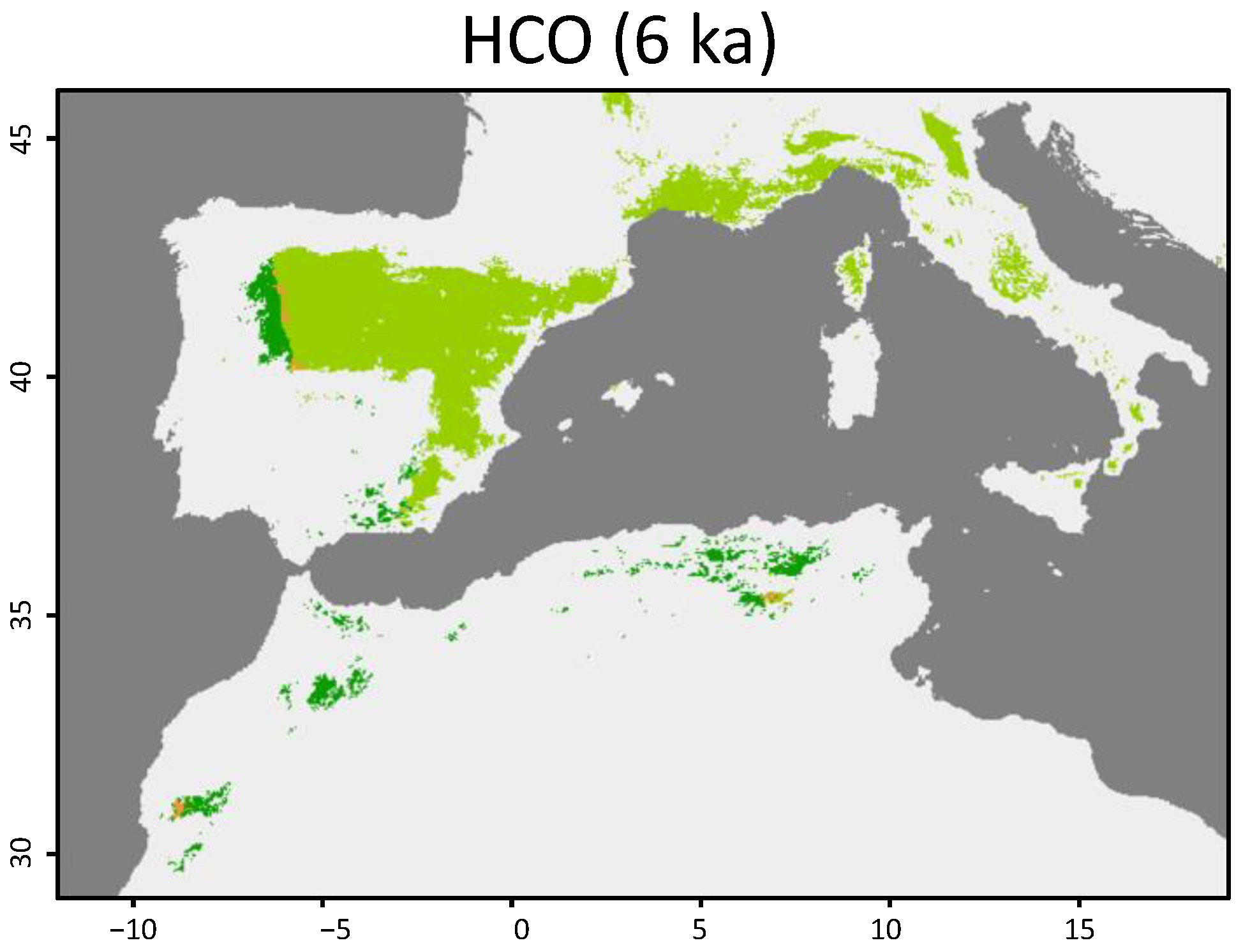
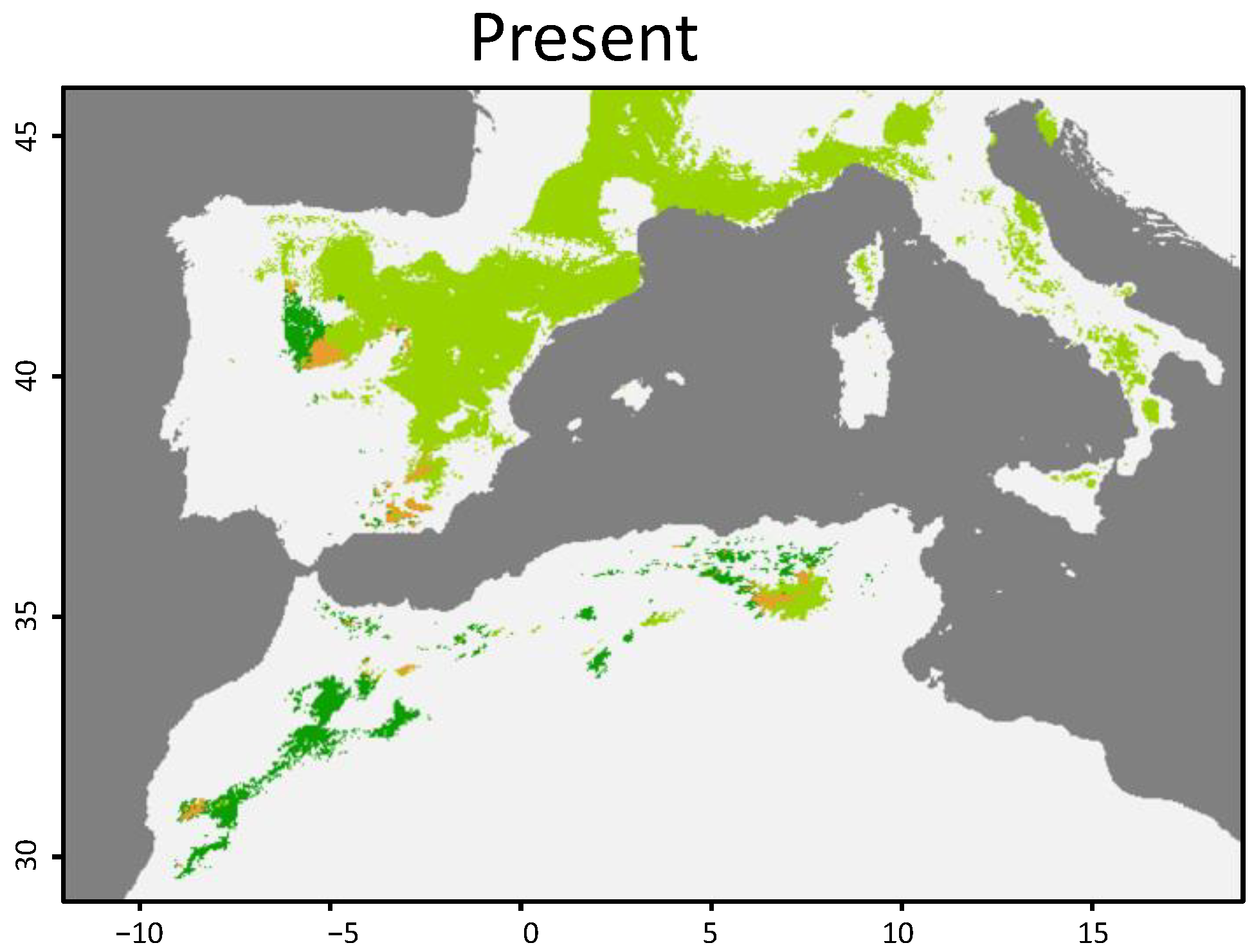
3.1. Geographical Distribution
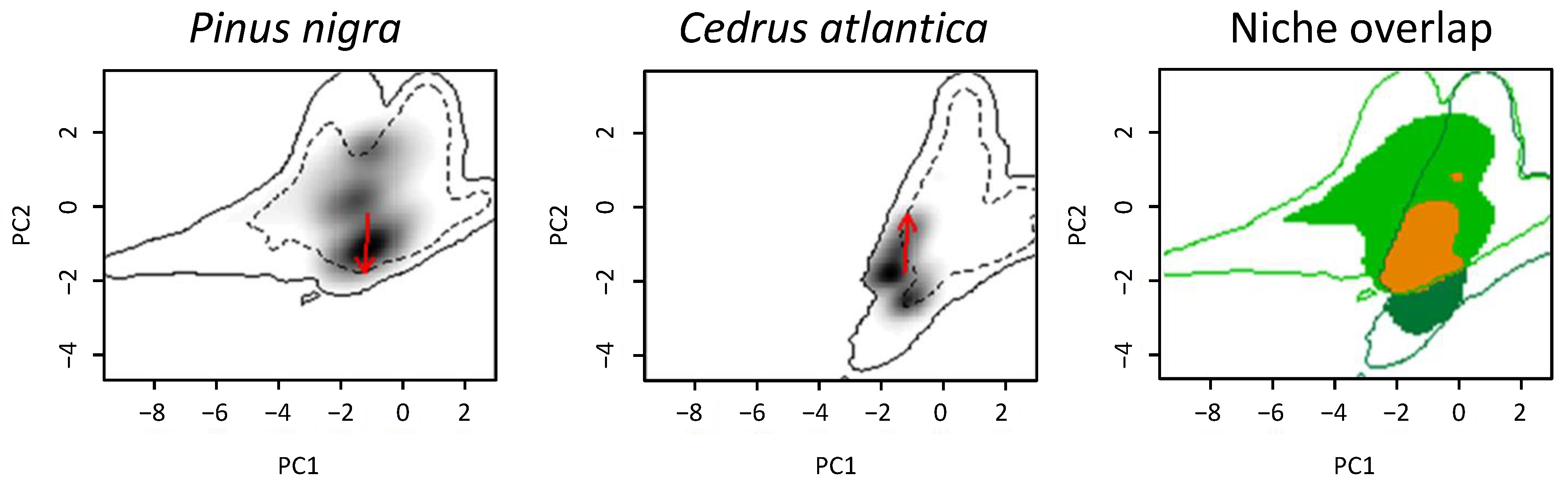
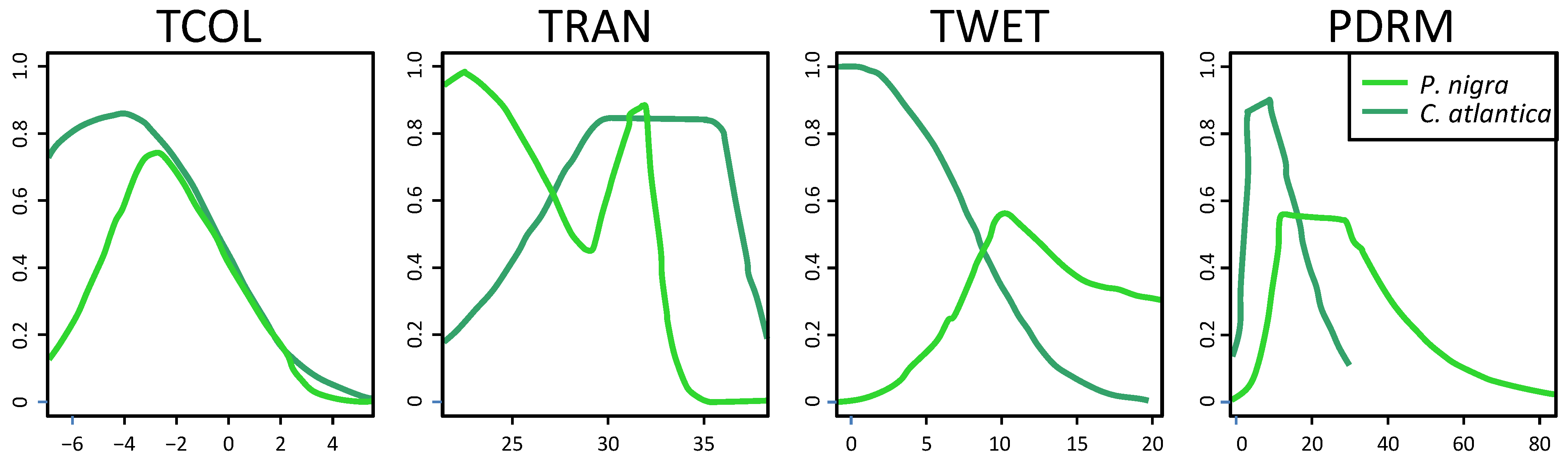
3.2. Niche Overlap
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TCOL | TRAN | TWET | PDRM | |
|---|---|---|---|---|
| TCOL | 1 | |||
| TRAN | 0.055 | 1 | ||
| TWET | 0.464 | 0.434 | 1 | |
| PDRM | −0.687 | −0.504 | −0.436 | 1 |
| HCO | LGM | |||
|---|---|---|---|---|
| CCSM4 | MIROC-ESM | CCSM4 | MIROC-ESM | |
| Obs. AUC | 0.7396 | 0.7830 | 0.5011 | 0.5425 |
| Null AUC | 0.7584 | 0.7398 | 0.7616 | 0.7769 |




References
- Abel-Schaad, D.; Iriarte, E.; López-Sáez, J.A.; Pérez-Díaz, S.; Sabariego Ruiz, S.; Cheddadi, R.; Alba-Sánchez, F. Are Cedrus atlantica forests in the Rif Mountains of Morocco heading towards local extinction? Holocene 2018, 28, 1023–1037. [Google Scholar] [CrossRef] [Green Version]
- Magri, D.; Di Rita, F.; Aranbarri, J.; Fletcher, W.; González-Sampériz, P. Quaternary disappearance of tree taxa from Southern Europe: Timing and trends. Quat. Sci. Rev. 2017, 163, 23–55. [Google Scholar] [CrossRef] [Green Version]
- Postigo-Mijarra, J.M.; Morla, C.; Barrón, E.; Morales-Molino, C.; García, S. Patterns of extinction and persistence of Arctotertiary flora in Iberia during the Quaternary. Rev. Palaeobot. Palynol. 2010, 162, 416–426. [Google Scholar] [CrossRef]
- Postigo-Mijarra, J.M.; Barrón, E.; Gómez Manzaneque, F.; Morla, C. Floristic changes in the Iberian Peninsula and Balearic Islands (South-West Europe) during the Cenozoic. J. Biogeogr. 2009, 36, 2025–2043. [Google Scholar] [CrossRef] [Green Version]
- Magri, D. Quaternary History of Cedrus in Southern Europe. Ann. Bot. 2012, 2, 57–66. [Google Scholar] [CrossRef]
- Boyd, A. Relict Conifers from the Mid-Pleistocene of Rhodes, Greece. Hist. Biol. 2009, 21, 1–15. [Google Scholar] [CrossRef]
- Jiménez-Moreno, G.; Anderson, R.S.; Ramos-Román, M.J.; Camuera, J.; Mesa-Fernández, J.M.; García-Alix, A.; Jiménez-Espejo, F.J.; Carrión, J.S.; López-Avilés, A. The Holocene Cedrus pollen record from Sierra Nevada (S Spain), a proxy for climate change in N Africa. Quat. Sci. Rev. 2020, 242, 106468. [Google Scholar] [CrossRef]
- Magri, D.; Parra, I. Late Quaternary Western Mediterranean pollen records and African winds. Earth Planet. Sci. Lett. 2002, 200, 401–408. [Google Scholar] [CrossRef]
- Aranbarri, J.; González-Sampériz, P.; Valero-Garcés, B.; Moreno, A.; Gil-Romera, G.; Sevilla-Callejo, M.; García-Prieto, E.; Di Rita, F.; Mata, M.P.; Morellón, M.; et al. Rapid climatic changes and resilient vegetation during the Lateglacial and Holocene in a continental region of South-Western Europe. Glob. Planet. Chang. 2014, 114, 50–65. [Google Scholar] [CrossRef] [Green Version]
- González-Sampériz, P.; Leroy, S.A.G.; Carrión, J.S.; Fernández, S.; García-Antón, M.; Gil-García, M.J.; Uzquiano, P.; Valero-Garcés, B.; Figueiral, I. Steppes, savannahs, forests and phytodiversity reservoirs during the Pleistocene in the Iberian Peninsula. Rev. Palaeobot. Palynol. 2010, 162, 427–457. [Google Scholar] [CrossRef] [Green Version]
- González-Sampériz, P.; Gil-Romera, G.; García-Prieto, E.; Aranbarri, J.; Moreno, A.; Morellón, M.; Sevilla-Callejo, M.; Leunda, M.; Santos, L.; Franco-Múgica, F.; et al. Strong continentality and effective moisture drove unforeseen vegetation dynamics since the Last Interglacial at inland Mediterranean areas: The Villarquemado sequence in NE Iberia. Quat. Sci. Rev. 2020, 242, 106425. [Google Scholar] [CrossRef]
- Manzano, S.; Carrión, J.S.; López-Merino, L.; Ochando, J.; Munuera, M.; Fernández, S.; González-Sampériz, P. Early to Mid-Holocene spatiotemporal vegetation changes and tsunami impact in a paradigmatic coastal transitional system (Doñana National Park, Southwestern Europe). Glob. Planet. Chang. 2018, 161, 66–81. [Google Scholar] [CrossRef] [Green Version]
- Bell, B.A.; Fletcher, W.J. Modern Surface Pollen assemblages from the Middle and High Atlas, Morocco: Insights into pollen representation and transport. Grana 2016, 55, 286–301. [Google Scholar] [CrossRef] [Green Version]
- Cheddadi, R.; Henrot, A.-J.; François, L.; Boyer, F.; Bush, M.; Carré, M.; Coissac, E.; De Oliveira, P.E.; Ficetola, F.; Hambuckers, A.; et al. Microrefugia, climate change, and conservation of Cedrus atlantica in the Rif Mountains, Morocco. Front. Ecol. Evol. 2017, 5, 114. [Google Scholar] [CrossRef] [Green Version]
- Alba-Sánchez, F.; Abel-Schaad, D.; López-Sáez, J.A.; Sabariego Ruiz, S.; Pérez-Díaz, S.; González-Hernández, A. Paleobiogeografía de Abies spp. y Cedrus atlantica en el Mediterráneo Occidental (Península Ibérica y Marruecos). Rev. Ecosist. 2018, 27, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Alba-Sánchez, F.; Abel-Schaad, D.; López-Sáez, J.A.; Sabariego-Ruiz, S.; Pérez-Díaz, S.; Luelmo-Lautenschlaeger, R.; Garrido-García, J.A. Early anthropogenic change in Western Mediterranean mountains (Sierra Nevada, SE Spain). Anthropocene 2021, 33, 100278. [Google Scholar] [CrossRef]
- Cheddadi, R.; Fady, B.; François, L.; Hajar, L.; Suc, J.-P.; Huang, K.; Demarteau, M.; Vendramin, G.G.; Ortu, E. Putative glacial refugia of Cedrus Atlantica deduced from Quaternary pollen records and modern genetic diversity. J. Biogeogr. 2009, 36, 1361–1371. [Google Scholar] [CrossRef]
- Linares, J.C.; Tíscar, P.A.; Camarero, J.J.; Taïqui, L.; Viñegla, B.; Seco, J.I.; Merino, J.; Carreira, J.A. Tree growth decline on relict Western-Mediterranean mountain forest: Causes and impacts. In Forest Decline: Causes and Impacts; Environmental Science, Engineering and Technology; Jenkins, J.A., Ed.; Nova Science Publishers: Hauppauge, NY, USA, 2011; pp. 91–110. ISBN 978-1-61470-002-9. [Google Scholar]
- Carrión, J.S.; Sánchez-Gómez, P.; Mota, J.F.; Yll, R.; Chaín, C. Holocene vegetation dynamics, fire and grazing in the Sierra de Gádor, Southern Spain. Holocene 2003, 13, 839–849. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S.; Fuentes, N.; González-Sampériz, P.; Sánchez Quirante, L.; Finlayson, J.C.; Fernández, S.; Andrade, A. Holocene environmental change in a montane region of Southern Europe with a long history of human settlement. Quat. Sci. Rev. 2007, 26, 1455–1475. [Google Scholar] [CrossRef]
- Campbell, J.F.E.; Fletcher, W.J.; Joannin, S.; Hughes, P.D.; Rhanem, M.; Zielhofer, C. Environmental drivers of Holocene forest development in the Middle Atlas, Morocco. Front. Ecol. Evol. 2017, 5, 113. [Google Scholar] [CrossRef] [Green Version]
- Carrión, J.S. Paleoflora y Paleovegetación de la Península Ibérica e Islas Baleares; Universidad de Murcia, Secretariado PU: Murcia, Spain, 2012; ISBN 978-84-615-9026-1. [Google Scholar]
- Muller, S.D.; Rhazi, L.; Andrieux, B.; Bottollier-Curtet, M.; Fauquette, S.; Saber, E.-R.; Rifai, N.; Daoud-Bouattour, A. Vegetation History of the Western Rif Mountains (NW Morocco): Origin, Late-Holocene Dynamics and Human Impact. Veg. Hist. Archaeobotany 2015, 24, 487–501. [Google Scholar] [CrossRef]
- Reille, M. Analyse pollinique de sédiments postglaciaires dans le Moyen Atlas et le Haut Atlas Marocains: Premiers résultats. Ecol. Mediterr. 1976, 2, 153–170. [Google Scholar] [CrossRef]
- Reille, M. Contribution pollenanalytique à l’histoire holocène de la végétation des montagnes du Rif (Maroc septentrional). Suppl. Bull. AFEQ 1977, 50, 53–76. [Google Scholar]
- Stambouli-Essassi, S.; Roche, E.; Bouzid, S. Evolution de la végétation et du climat dans le Nord-ouest de la Tunisie au cours des 40 derniers millénaires. Geo-EcoTrop 2007, 31, 171–214. [Google Scholar]
- Blois, J.L.; Gotelli, N.J.; Behrensmeyer, A.K.; Faith, J.T.; Lyons, S.K.; Williams, J.W.; Amatangelo, K.L.; Bercovici, A.; Du, A.; Eronen, J.T.; et al. A Framework for evaluating the influence of climate, dispersal limitation, and biotic interactions using fossil pollen associations across the Late Quaternary. Ecography 2014, 37, 1095–1108. [Google Scholar] [CrossRef]
- D’Amen, M.; Mod, H.K.; Gotelli, N.J.; Guisan, A. Disentangling Biotic interactions, environmental filters, and dispersal limitation as drivers of species co-occurrence. Ecography 2018, 41, 1233–1244. [Google Scholar] [CrossRef] [Green Version]
- Broennimann, O.; Fitzpatrick, M.C.; Pearman, P.B.; Petitpierre, B.; Pellissier, L.; Yoccoz, N.G.; Thuiller, W.; Fortin, M.-J.; Randin, C.; Zimmermann, N.E.; et al. Measuring ecological niche overlap from occurrence and spatial environmental data. Glob. Ecol. Biogeogr. 2012, 21, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Warren, D.L.; Glor, R.E.; Turelli, M. Environmental niche equivalency versus conservatism: Quantitative approaches to niche evolution. Evolution 2008, 62, 2868–2883. [Google Scholar] [CrossRef] [PubMed]
- Benito Garzón, M.; Sánchez de Dios, R.; Sáinz Ollero, H. Predictive modelling of tree species distributions on the Iberian Peninsula during the Last Glacial Maximum and Mid-Holocene. Ecography 2007, 30, 120–134. [Google Scholar] [CrossRef]
- López-Tirado, J.; Hidalgo, P.J. A high resolution predictive model for relict trees in the Mediterranean-mountain forests (Pinus sylvestris L., P. nigra Arnold and Abies pinsapo Boiss.) from the South of Spain: A Reliable management tool for reforestation. For. Ecol. Manag. 2014, 330, 105–114. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Manzanedo, R.D.; Rodriguez-Vallejo, C.; Gazol, A.; Palacios-Rodríguez, G.; Camarero, J.J. Competition modulates the response of growth to climate in pure and mixed Abies pinsapo subsp. maroccana forests in Northern Morocco. For. Ecol. Manag. 2020, 459, 117847. [Google Scholar] [CrossRef]
- Valladares, A. Abetales de Abies pinsapo Boiss. In Bases Ecológicas Preliminares para la Conservación de los Tipos de Hábitat de Interés Comunitario en España; Ministerio Medio Ambiente y Medio Rural y Marino: Madrid, Spain, 2009; Volume 9520, ISBN 978-84-491-0911-9. [Google Scholar]
- Alba-Sánchez, F.; López-Sáez, J.A.; Benito de Pando, B.M.; Linares, J.C.; Nieto-Lugilde, D.; López-Merino, L. Past and present potential distribution of the Iberian Abies species: A phytogeographic approach using fossil pollen data and species distribution models. Divers. Distrib. 2010, 16, 214–228. [Google Scholar] [CrossRef] [Green Version]
- Blois, J.L. Recent advances in using species distributional models to understand past distributions. Front. Biogeogr. 2012, 3, 123–124. [Google Scholar] [CrossRef] [Green Version]
- Brewer, S.; Jackson, S.T.; Williams, J.W. Paleoecoinformatics: Applying geohistorical data to ecological questions. Trends Ecol. Evol. 2012, 27, 104–112. [Google Scholar] [CrossRef]
- Svenning, J.-C.; Fløjgaard, C.; Marske, K.A.; Nógues-Bravo, D.; Normand, S. Applications of species distribution modeling to paleobiology. Quat. Sci. Rev. 2011, 30, 2930–2947. [Google Scholar] [CrossRef]
- Varela, S.; Lobo, J.M.; Hortal, J. Using species distribution models in paleobiogeography: A matter of data, predictors and concepts. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2011, 310, 451–463. [Google Scholar] [CrossRef]
- Bouahmed, A.; Vessella, F.; Schirone, B.; Krouchi, F.; Derridj, A. Modeling Cedrus atlantica potential distribution in North Africa across time: New putative glacial refugia and future range shifts under climate change. Reg. Environ. Chang. 2019, 19, 1667–1682. [Google Scholar] [CrossRef]
- Demarteau, M.; Francois, L.; Cheddadi, R.; Roche, E. Réponses de Cedrus atlantica aux changements climatiques passés et futurs. Geo-Eco-Trop 2007, 31, 105–146. [Google Scholar]
- Benito Garzón, M.; Sánchez de Dios, R.; Sáinz Ollero, H. The evolution of the Pinus sylvestris L. area in the Iberian Peninsula from the Last Glacial Maximum to 2100 under climate change. Holocene 2008, 18, 705–714. [Google Scholar] [CrossRef]
- Di Pasquale, G.; Saracino, A.; Bosso, L.; Russo, D.; Moroni, A.; Bonanomi, G.; Allevato, E. Coastal pine-oak glacial refugia in the Mediterranean Basin: A biogeographic approach based on charcoal analysis and spatial modelling. Forests 2020, 11, 673. [Google Scholar] [CrossRef]
- Terrab, A.; Hampe, A.; Lepais, O.; Talavera, S.; Vela, E.; Stuessy, T.F. Phylogeography of North African Atlas cedar (Cedrus atlantica, Pinaceae): Combined molecular and fossil data reveal a complex Quaternary history. Am. J. Bot. 2008, 95, 1262–1269. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P. Cedrus atlantica. The IUCN Red List of Threatened Species. e.T42303A2970716. 2013. Available online: https://www.iucnredlist.org/species/42303/2970716 (accessed on 27 November 2014).
- Farjon, A. Pinus nigra. The IUCN Red List of Threatened Species. e.T42386A2976817. 2013. Available online: https://www.iucnredlist.org/species/42386/2976817 (accessed on 13 January 2020).
- Phillips, S.J.; Dudık, M.; Shapire, R. Maxent Software for Species Habitat Modeling. 2013. Available online: http://biodiversityinformatics.amnh.org/open_source/maxent/ (accessed on 25 January 2013).
- Hijmans, R.J.; Phillips, S.J.; Leathwick, J.; Elith, J. Package ‘Dismo’. Species Distribution Modeling. R Package Version 0.7-17. 2014. Available online: https://rspatial.org/raster/sdm/ (accessed on 10 September 2014).
- Phillips, S.J. A Brief Tutorial on Maxent 2017. Available online: https://pdf4pro.com/view/a-brief-tutorial-on-maxent-american-museum-of-natural-5a321d.html (accessed on 2 April 2021).
- Hijmans, R.J.; Cameron, S.E.; Parra, J.L.; Jones, P.G.; Jarvis, A. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol. 2005, 25, 1965–1978. [Google Scholar] [CrossRef]
- EPD European Pollen Database. Available online: http://www.europeanpollendatabase.net/data/ (accessed on 26 April 2012).
- Booth, T.H.; Nix, H.A.; Busby, J.R.; Hutchinson, M.F. Bioclim: The first species distribution modelling package, its early applications and relevance to most current MaxEnt studies. Divers. Distrib. 2014, 20, 1–9. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.J.; Hastie, T.; Dudík, M.; Chee, Y.E.; Yates, C.J. A statistical explanation of MaxEnt for ecologists. Divers. Distrib. 2011, 17, 43–57. [Google Scholar] [CrossRef]
- Merow, C.; Smith, M.J.; Silander, J.A. A practical guide to MaxEnt for modeling species’ distributions: What it does, and why inputs and settings matter. Ecography 2013, 36, 1058–1069. [Google Scholar] [CrossRef]
- Rodríguez-Sánchez, F.; Hampe, A.; Jordano, P.; Arroyo, J. Past tree range dynamics in the Iberian Peninsula inferred through phylogeography and palaeodistribution modelling: A review. Rev. Palaeobot. Palynol. 2010, 162, 507–521. [Google Scholar] [CrossRef]
- Raes, N.; ter Steege, H. A null-model for significance testing of presence-only species distribution models. Ecography 2007, 30, 727–736. [Google Scholar] [CrossRef]
- Di Cola, V.; Broennimann, O.; Petitpierre, B.; Breiner, F.T.; D’Amen, M.; Randin, C.; Engler, R.; Pottier, J.; Pio, D.; Dubuis, A.; et al. Ecospat: An R package to support spatial analyses and modeling of species niches and distributions. Ecography 2017, 40, 774–787. [Google Scholar] [CrossRef]
- Camuera, J.; Jiménez-Moreno, G.; Ramos-Román, M.J.; García-Alix, A.; Toney, J.L.; Anderson, R.S.; Jiménez-Espejo, F.; Bright, J.; Webster, C.; Yanes, Y.; et al. Vegetation and climate changes during the last two glacial-interglacial cycles in the Western Mediterranean: A new long pollen record from Padul (Southern Iberian Peninsula). Quat. Sci. Rev. 2019, 205, 86–105. [Google Scholar] [CrossRef]
- Pons, A.; Reille, M. The Holocene- and Upper Pleistocene pollen record from Padul (Granada, Spain): A new study. Palaeogeogr. Palaeoclimatol. Palaeoecol. 1988, 66, 243–263. [Google Scholar] [CrossRef]






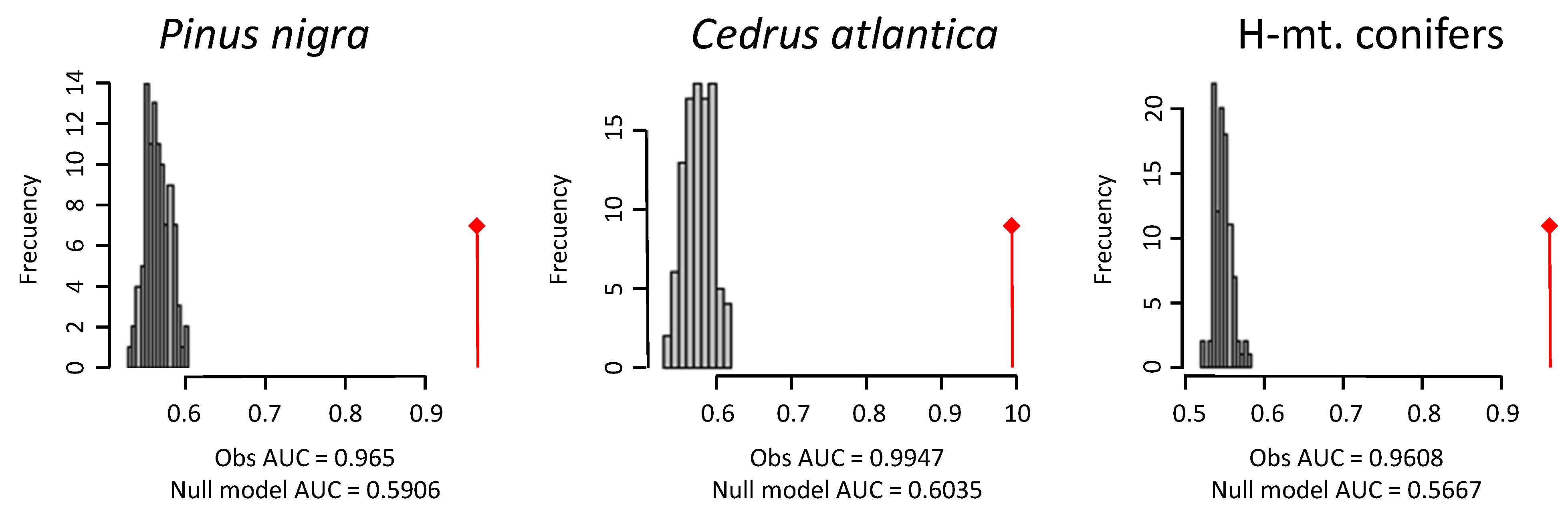
| SPECIES | n | AUC | Null Model AUC |
|---|---|---|---|
| Pinus nigra | 257 | 0.965 | 0.591 |
| Cedrus atlantica | 211 | 0.995 | 0.604 |
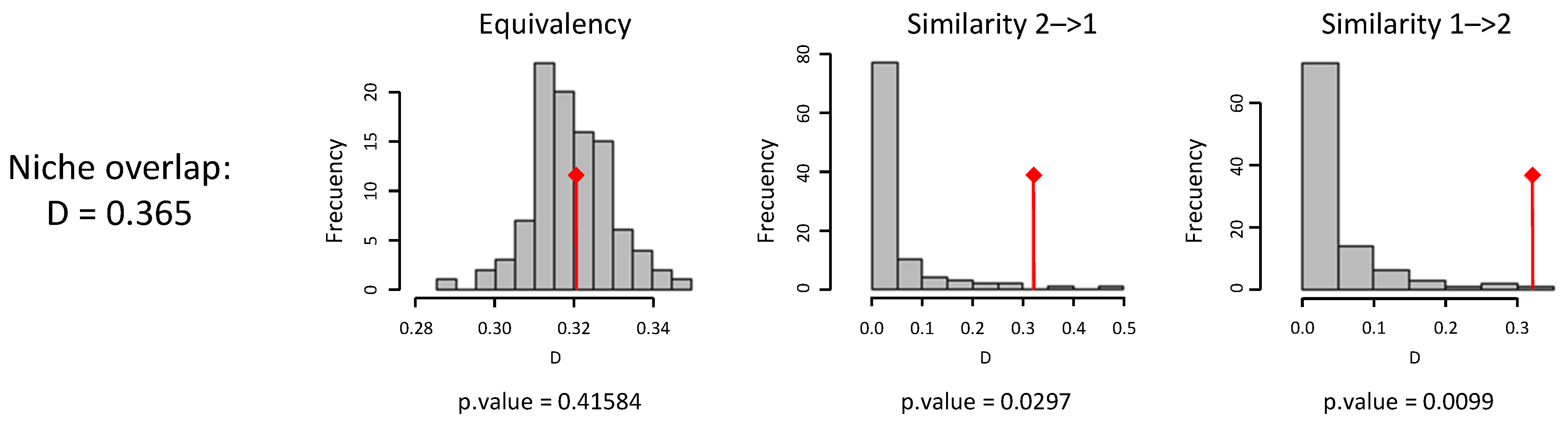
| SPECIES | Overlap (D) | Equivalence | Similarity 1→2 | Similarity 2→1 |
|---|---|---|---|---|
| P. nigra–C. atlantica | 0.365 | ns | Similar * | Similar ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Hernández, A.; Nieto-Lugilde, D.; Alba-Sánchez, F.; Peñas, J. Biological Interaction as a Possible Ultimate Driver in the Local Extinction of Cedrus atlantica in the Iberian Peninsula. Diversity 2022, 14, 136. https://doi.org/10.3390/d14020136
González-Hernández A, Nieto-Lugilde D, Alba-Sánchez F, Peñas J. Biological Interaction as a Possible Ultimate Driver in the Local Extinction of Cedrus atlantica in the Iberian Peninsula. Diversity. 2022; 14(2):136. https://doi.org/10.3390/d14020136
Chicago/Turabian StyleGonzález-Hernández, Antonio, Diego Nieto-Lugilde, Francisca Alba-Sánchez, and Julio Peñas. 2022. "Biological Interaction as a Possible Ultimate Driver in the Local Extinction of Cedrus atlantica in the Iberian Peninsula" Diversity 14, no. 2: 136. https://doi.org/10.3390/d14020136
APA StyleGonzález-Hernández, A., Nieto-Lugilde, D., Alba-Sánchez, F., & Peñas, J. (2022). Biological Interaction as a Possible Ultimate Driver in the Local Extinction of Cedrus atlantica in the Iberian Peninsula. Diversity, 14(2), 136. https://doi.org/10.3390/d14020136