
Small Mammal Diversity in Response to Land Transformation and Seasonal Variation in South Africa



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Umetsu, F.; Pardini, R. Small mammals in a mosaic of forest remnants and anthropogenic habitats—evaluating matrix quality in an Atlantic forest landscape. Landsc. Urban Plan. 2007, 22, 517–530. [Google Scholar] [CrossRef]
- Irwin, M.; Junge, R.; Raharison, J.; Samonds, K.E. Variation in physiological health of diademed sifakas across intact and fragmented forest at Tsinjoarivo, eastern Madagascar. Am. J. Primatol. 2010, 72, 1013–1025. [Google Scholar] [CrossRef] [PubMed]
- Waters, C.N.; Zalasiewicz, J.; Summerhayes, C.; Barnosky, A.D.; Poirier, C.; Gauszka, A.; Cearreta, A.; Edgeworth, M.; Ellis, E.C.; Ellis, M.; et al. The Anthropocene is functionally and stratigraphically distinct from the Holocene. Science 2016, 351, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Christie, A.P.; Amano, T.; Martin, P.A.; Shackelford, G.E.; Simmons, B.I.; Sutherland, W.J. Simple study designs in ecology produce inaccurate estimates of biodiversity responses. J. Appl. Ecol. 2019, 56, 2742–2754. [Google Scholar] [CrossRef] [Green Version]
- WWF. Living Planet Report 2016. Risk and Resilience in a New Era; WWF International: Gland, Switzerland, 2016. [Google Scholar]
- Marris, E. Pre-emptive strike: Outwitting extinction. Nat. Clim. Change 2008, 1, 140–141. [Google Scholar] [CrossRef]
- Cadotte, M.W.; Arnillas, C.A.; Livingstone, S.W.; Yasui, S.L.E. Faunal indicator taxa selection for monitoring ecosystem health. Funct. Ecol. 2016, 92, 185–197. [Google Scholar]
- Wilson, J.R.U.; Faulkner, K.T.; Rahlao, S.J.; Richardson, D.M.; Zengeya, T.A.; van Wilgen, B.W. Indicators for monitoring biological invasions at a national level. J. Appl. Ecol. 2018, 55, 2612–2620. [Google Scholar] [CrossRef]
- Heroldová, M.; Bryja, J.; Zejda, J.; Tkadlec, E. Structure and diversity of small mammal communities in agriculture landscape. Agric. Ecosyst. Environ. 2007, 120, 206–210. [Google Scholar] [CrossRef]
- Foord, S.H.; Swanepoel, L.H.; Evans, S.W.; Schoeman, S.; Erasmus, B.F.N.; Schoeman, M.C.; Keith, M.; Smith, A.; Mauda, E.V.; Maree, N.; et al. Animal taxa contrast in their scale-dependent responses to land use change in rural Africa. PLoS ONE 2018, 13, e0194336. [Google Scholar] [CrossRef] [Green Version]
- Monadjem, A.; Perrin, M. Population fluctuations and community structure of small mammals in a Swaziland grassland over a three-year period. Afr. Zool. 2003, 38, 127–137. [Google Scholar] [CrossRef]
- Simelane, F.N.; Mahlaba, T.A.M.; Shapiro, J.T.; MacFadyen, D.; Monadjem, A. Habitat associations of small mammals in the foothills of the Drakensberg Mountains, South Africa. Mammalia 2017, 82, 144–152. [Google Scholar] [CrossRef] [Green Version]
- Magurran, A.E. Measuring Biological Diversity; Blackwell Publishing: Victoria, Australia, 2004. [Google Scholar]
- Ramahlo, M.; Chimimba, C.; Pirk, C.; Ganswindt, A. Non-invasive monitoring of adrenocortical activity in free-ranging Namaqua rock mice Micaelamys namaquensis from South Africa in response to anthropogenic land use and season. Wildl. Biol. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Cincotta, R.P.; Wisnewski, J.; Engelman, R. Human population in the biodiversity hotspots. Nature 2000, 404, 990–992. [Google Scholar] [CrossRef] [PubMed]
- Scott, D.M.; Brown, D.; Mahood, S.; Denton, B.; Silburn, A.; Rakotondraparany, F. The impacts of forest clearance on lizard, small mammal and bird communities in the arid spiny forest, southern Madagascar. Biol. Conserv. 2006, 127, 72–87. [Google Scholar] [CrossRef]
- Colles, A.; Liow, L.H.; Prinzing, A. Are specialists at risk under environmental change? Neoecological, paleoecological and phylogenetic approaches. Ecol. Lett. 2009, 12, 849–863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, J.K.; Sciulli, P.W.; Fooce, C.D.; Waite, T.A. Forecasting global biodiversity threats associated with human population growth. Biol. Conserv. 2004, 115, 161–164. [Google Scholar] [CrossRef]
- Watson, J.E.M.; Whittaker, R.J.; Dawson, T.P. Habitat structure and proximity to forest edge affect the abundance and distribution of forest-dependent birds in tropical coastal forests of southeastern Madagascar. Biol. Conserv. 2004, 120, 315–331. [Google Scholar] [CrossRef]
- Galetti, M.; Giacomini, H.C.; Bueno, R.S.; Bernardo, C.S.S.; Marques, R.M.; Bovendorp, R.S.; Steffler, C.E.; Rubim, P.; Gobbo, S.K.; Donatti, C.I.; et al. Priority areas for the conservation of Atlantic forest large mammals. Biol. Conserv. 2009, 142, 1229–1241. [Google Scholar] [CrossRef]
- Merritt, J.F. The Biology of Small Mammals; Johns Hopkins University Press: Baltimore, MD, USA, 2010. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-1. Available online: https://www.iucnredlist.org (accessed on 8 January 2021).
- Mills, M.G.L.; Hes, L. The Complete Book of Southern African Mammals; Struik Publishers: Cape Town, South Africa, 1997. [Google Scholar]
- Skinner, J.; Chimimba, C. The Mammals of the Southern African Subregion, 3rd ed.; Cambridge University Press: Cape Town, South Africa, 2005. [Google Scholar]
- Skinner, J.; Smithers, R. The Mammals of the Southern African Subregion, 2nd ed.; University of Pretoria: Pretoria, South Africa, 1990. [Google Scholar]
- Monadjem, A.; Taylor, P.J.; Denys, C.; Cotterill, F.P.D. Rodents of Sub-Saharan Africa: A Biogeographic and Taxonomic Synthesis; De Gruyter: Berlin, Germany, 2015. [Google Scholar]
- Rymer, T.L.; Pillay, N.; Schradin, C. Extinction or survival? Behavioral flexibility in response to environmental change in the African striped mouse Rhabdomys. Sustainability 2013, 5, 163–186. [Google Scholar] [CrossRef] [Green Version]
- Spear, D.; Foxcroft, L.C.; Bezuidenhout, H.; McGeoch, M.A. Human population density explains alien species richness in protected areas. Biol. Conserv. 2013, 159, 137–147. [Google Scholar] [CrossRef]
- Ellis, E.C.; Kaplan, J.O.; Fuller, D.Q.; Vavrus, S.; Klein-Goldewijk, K.; Verburg, P.H. Used planet: A global history. Proc. Natl. Acad. Sci. USA 2013, 110, 7978–7985. [Google Scholar] [CrossRef] [Green Version]
- Rutherford, M.C.; Mucina, L. Biomes and bioregions of southern Africa: The vegetation of South Africa, Lesotho and Swaziland. Strelitzia 2006, 19, 31–51. [Google Scholar]
- Louw, J.; Business Connexions, De Wildt, North West Province, South Africa; Louw, E.; Business Connexions, De Wildt, North West Province, South Africa. Personal communication, 2020. Rainfall statistics for De Wildt, North West province.
- Newbery, C.H. A key to the Soricidae, Macroscelididae, Gliridae and Muridae of Gauteng, North West province, Mpumalanga and the Northern province, South Africa. Koedoe 1999, 42, 51–55. [Google Scholar] [CrossRef] [Green Version]
- Aplin, K.P.; Brown, P.R.; Jacob, J.; Krebs, C.J.; Singleton, G.R. Field Methods for Rodent Studies in Asia and the Indo-Pacific. Australian Centre for International Agricultural Research Monograph; CSIRO Publishing: Canberra, Australia, 2003. [Google Scholar]
- Fleckman, P.; Jaeger, K.; Silva, K.A.; Sundberg, J.P. Comparative anatomy of mouse and human nails. Anat. Rec. 2013, 296, 521–532. [Google Scholar] [CrossRef] [Green Version]
- Meheretu, Y.; Welegerima, K.; Sluydts, V.; Bauer, H.; Gebrehiwot, K.; Deckers, J.; Makundi, R.; Leirs, H. Reproduction and survival of rodents in crop fields: The effects of rainfall, crop stage and stone-bund density. Wildl. Res. 2015, 42, 158. [Google Scholar] [CrossRef]
- Anderson, M.J.; Crist, T.O.; Chase, J.M.; Vellend, M.; Inouye, B.D.; Freestone, A.L.; Sanders, N.J.; Cornell, H.V.; Comita, L.S.; Davies, K.F.; et al. Navigating the multiple meanings of β diversity: A roadmap for the practicing ecologist. Ecol. Lett. 2011, 14, 19–28. [Google Scholar] [CrossRef]
- Bastos, A.D.; Nair, D.; Taylor, P.J.; Brettschneider, H.; Kirsten, F.; Mostert, E.; von Maltitz, E.; Lamb, J.M.; van Hooft, P.; Belmain, S.R.; et al. Genetic monitoring detects an overlooked cryptic species and reveals the diversity and distribution of three invasive Rattus congeners in South Africa. BMC Genet. 2011, 12, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.G.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Baselga, A. The relationship between species replacement, dissimilarity derived from nestedness. Glob. Ecol. Biogeogr. 2008, 21, 1223–1232. [Google Scholar] [CrossRef]
- Baselga, A. Partitioning the turnover and nestedness components of beta diversity. Glob. Ecol. Biogeogr. 2009, 19, 134–143. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package, R Package; Version 2.5-7; R Core Team: Vienna, Austria, 2020; Available online: https://CRAN.R-project.org/package=vegan (accessed on 25 October 2021).
- Saetnan, E.R.; Skarpe, C. The effect of ungulate grazing on a small mammal community in southeastern Botswana. Afr. Zool. 2006, 41, 9–16. [Google Scholar] [CrossRef]
- Blaum, N.; Rossmanith, E.; Jeltsch, F. Land use affects rodent communities in Kalahari savannah rangelands. Afr. J. Ecol. 2007, 45, 189–195. [Google Scholar] [CrossRef]
- Mason, G.; Littin, K. The humaneness of rodent pest control. Anim. Welf. 2003, 12, 1–38. [Google Scholar]
- Welegerima, K.; Meheretu, Y.; Haileselassie, T.H.; Gebre, B.; Kidane, D.; Massawe, A.W.; Mbije, N.E.; Makundi, R.H. Abundance and microhabitat use of rodent species in crop fields and bushland in Ethiopia. J. Vertebr. Biol. 2020, 69, 1–6. [Google Scholar] [CrossRef]
- Van den Heuvel, I.M.; Midgley, J.J. Towards an isotope ecology of Cape Fynbos small mammals. Afr. Zool. 2014, 49, 195–202. [Google Scholar] [CrossRef]
- Muteka, S.P.; Chimimba, C.T.; Bennett, N.C. Reproductive seasonality in Aethomys namaquensis (Rodentia: Muridae) from southern Africa. J. Mammal 2006, 87, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Leirs, H.; Verhagen, R.; Verheyen, W. The basis of reproductive seasonality in Mastomys rats (Rodentia: Muridae) in Tanzania. J. Trop. Ecol. 1994, 10, 55–66. [Google Scholar] [CrossRef]
- Flynn, D.F.B.; Gogol-Prokurat, M.; Nogeire, T.; Molinari, N.; Richers, B.T.; Lin, B.B.; Simpson, N.; Mayfield, M.M.; DeClerck, F. Loss of functional diversity under land use intensification across multiple taxa. Ecol. Lett. 2009, 12, 22–33. [Google Scholar] [CrossRef]
- Horváth, A.; March, I.J.; Wolf, J.H.D. Rodent diversity and land use in Montebello, Chiapas, Mexico. Stud. Neotrop. Fauna Environ. 2001, 36, 169–176. [Google Scholar] [CrossRef]
- Schmidt, N.M.; Olsen, H.; Bildsøe, M.; Sluydts, V.; Leirs, H. Effects of grazing intensity on small mammal population ecology in wet meadows. Basic Appl. Ecol. 2005, 6, 57–66. [Google Scholar] [CrossRef]
- Chia, E.K.; Bassett, M.; Leonard, S.W.J.; Holland, G.J.; Ritchie, E.G.; Clarke, M.F.; Bennett, A.F. Effects of the fire regime on mammal occurrence after wildfire: Site effects vs. landscape context in fire-prone forests. For. Ecol. Manag. 2016, 363, 130–139. [Google Scholar] [CrossRef]
- Ferreira, S.; van Aarde, R. Maintaining diversity through intermediate disturbances: Evidence from rodents colonizing rehabilitating coastal dunes. Afr. J. Ecol. 2000, 38, 286–294. [Google Scholar] [CrossRef] [Green Version]
- Wright, N.I. Ecological Impacts of Highveld Gerbils (Tatera brantsii) on a Rehabilitated Ash Disposal Site 2006. Doctoral Dissertation, North West University, Potchefstroom, South Africa.
- Monadjem, A.; Perrin, M. Population dynamics of Lemniscomys rosalia (Muridae: Rodentia) in a Swaziland grassland: Effects of food and fire. Afr. Zool. 1997, 32, 129–135. [Google Scholar]
- Field, A.C. Seasonal changes in reproduction, diet and body composition of two equatorial rodents. Afr. J. Ecol. 1975, 13, 221–235. [Google Scholar] [CrossRef]
- Richardson, E.J.; Perrin, M.R. Seasonal changes in body mass, torpidity, and reproductive activity of captive fat mice, Steatomys pratensis. Isr. J. Ecol. Evol. 1992, 38, 315–322. [Google Scholar]
- Sánchez-Cordero, V. Elevation gradients of diversity for rodents and bats in Oaxaca, Mexico. Ecology 2001, 10, 63–76. [Google Scholar] [CrossRef]
- Magurran, A.E.; Henderson, P.A. More than the sum of the parts: Annual partitioning within spatial guilds underpins community regulation. Proc. R. Soc. B 2008, 285, 1–6. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Study Site | Land Use | Abbreviation | GPS Location | Primary Vegetation | Topography |
|---|---|---|---|---|---|
| 1 | Animal breeding and rehabilitation Ecotourism | ABR | 25°40′25.2″ S 27°55′17.4″ E | Thorny, semi-open thickets | Rocky, slight incline near the base of the mountain |
| 2 | Agricultural Residential | ARF | 25°39′31.3″ S 27°55′08.0″ E | Grassy herbaceous layer | Flat at the base of the mountain |
| 3 | Residential | RFS | 25°39′20.3″ S 27°55′15.7″ E | Herbaceous and semi-open thickets | Flat at the base of the mountain |
| 4 | Industrial Agricultural Residential | IAR | 25°38′43.3″ S 27°55′48.8″ E | Herbaceous and semi-open thickets | Flat at the base of the mountain |
| 5 | Conservation Residential | NCR | 25°40′39.6″ S 27°57′48.9″ E | Herbaceous and semi-open thickets | Rocky, steep incline along the mountain |
| Species Name | ABR | ARF | RFS | IAR | NCR | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Dry | Wet | Dry | Wet | Dry | Wet | |
| Macroscelidea | ||||||||||
| Elephantulus brachyrhynchus/myurus [Elephant shrew] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Rodentia | ||||||||||
| Aethomys ineptus [Tete veld rat] | 3 | 21 | 0 | 0 | 0 | 0 | 0 | 3 | 10 | 17 |
| Dendromus mystacalis [Climbing mouse] | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 |
| Gerbilliscus brantsii/leucogaster [Bushveld/Highveld gerbil] | 0 | 0 | 0 | 0 | 0 | 0 | 20 | 2 | 0 | 2 |
| Lemniscomys rosalia [Single-striped mouse] | 1 | 8 | 3 | 7 | 15 | 14 | 11 | 10 | 9 | 0 |
| Mastomys coucha [Multimammate mouse] | 4 | 19 | 20 | 120 | 45 | 70 | 12 | 29 | 14 | 2 |
| Micaelamys namaquensis [Namaqua rock mouse] | 2 | 16 | 0 | 18 | 0 | 12 | 0 | 21 | 23 | 46 |
| Mus minutoides [Pygmy mouse] | 2 | 0 | 6 | 1 | 10 | 0 | 8 | 1 | 2 | 0 |
| Otomys irroratus [Vlei rat] | 0 | 0 | 0 | 0 | 2 | 3 | 0 | 2 | 3 | 0 |
| Rhabdomys pumilio [Three-striped mouse] | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | 0 |
| Saccostomus campestris [Pouched mouse] | 0 | 0 | 0 | 3 | 0 | 1 | 2 | 6 | 0 | 0 |
| Steatomys pratensis [Fat mouse] | 0 | 0 | 0 | 1 | 0 | 15 | 2 | 2 | 0 | 0 |
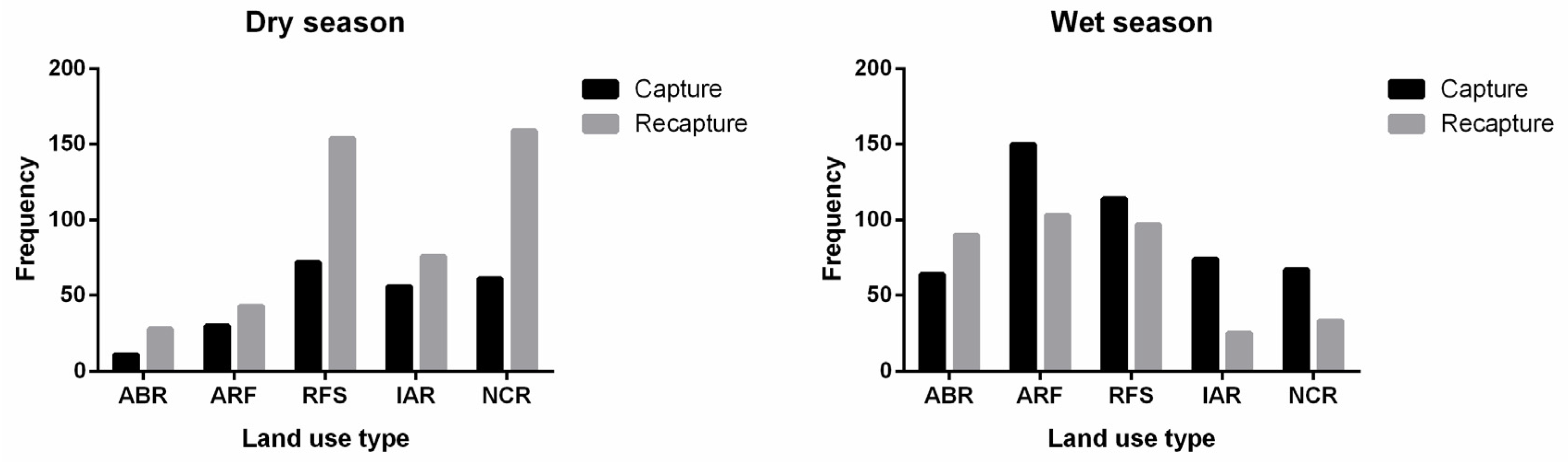
| Captures (N) * | 12 | 64 | 29 | 150 | 72 | 115 | 57 | 76 | 63 | 67 |
| Species richness | 5 | 4 | 3 | 6 | 4 | 6 | 7 | 9 | 8 | 4 |
| Shannon diversity index (H) * | 1.52 | 1.33 | 0.82 | 0.72 | 0.99 | 1.20 | 1.64 | 1.66 | 1.66 | 0.82 |
| Simpson diversity index (D) | 0.78 | 0.73 | 0.47 | 0.34 | 0.55 | 0.59 | 0.82 | 0.75 | 0.76 | 0.46 |
| Pielou’s evenness index (J) | 0.94 | 0.96 | 0.74 | 0.40 | 0.71 | 0.67 | 0.84 | 0.76 | 0.80 | 0.59 |
| ABR | ARF | RFS | IAR | |||||
|---|---|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Dry | Wet | Dry | Wet | |
| ARF | 0.25 | 0.40 | - | - | - | - | - | - |
| RFS | 0.33 | 0.40 | 0.14 | 0.17 | - | - | - | - |
| IAR | 0.50 | 0.38 | 0.40 | 0.20 | 0.45 | 0.20 | - | - |
| NCR | 0.23 | 0.25 | 0.45 | 0.60 | 0.33 | 0.60 | 0.60 | 0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ramahlo, M.; Somers, M.J.; Hart, D.W.; Ganswindt, A. Small Mammal Diversity in Response to Land Transformation and Seasonal Variation in South Africa. Diversity 2022, 14, 138. https://doi.org/10.3390/d14020138
Ramahlo M, Somers MJ, Hart DW, Ganswindt A. Small Mammal Diversity in Response to Land Transformation and Seasonal Variation in South Africa. Diversity. 2022; 14(2):138. https://doi.org/10.3390/d14020138
Chicago/Turabian StyleRamahlo, Mmatsawela, Michael John Somers, Daniel William Hart, and Andre Ganswindt. 2022. "Small Mammal Diversity in Response to Land Transformation and Seasonal Variation in South Africa" Diversity 14, no. 2: 138. https://doi.org/10.3390/d14020138
APA StyleRamahlo, M., Somers, M. J., Hart, D. W., & Ganswindt, A. (2022). Small Mammal Diversity in Response to Land Transformation and Seasonal Variation in South Africa. Diversity, 14(2), 138. https://doi.org/10.3390/d14020138