
Diversity and Molecular Phylogeny of Pagurid Hermit Crabs (Anomura: Paguridae: Pagurus)


Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling
2.2. Retrieval of Molecular Data
2.3. Characteristics of Analysed Sequences Data
2.4. Phylogenetic Analyses
2.5. Estimating Diversification Rates from Phylogenetic Information
2.6. Morphological Characters Analyses
3. Results
3.1. Species Composition
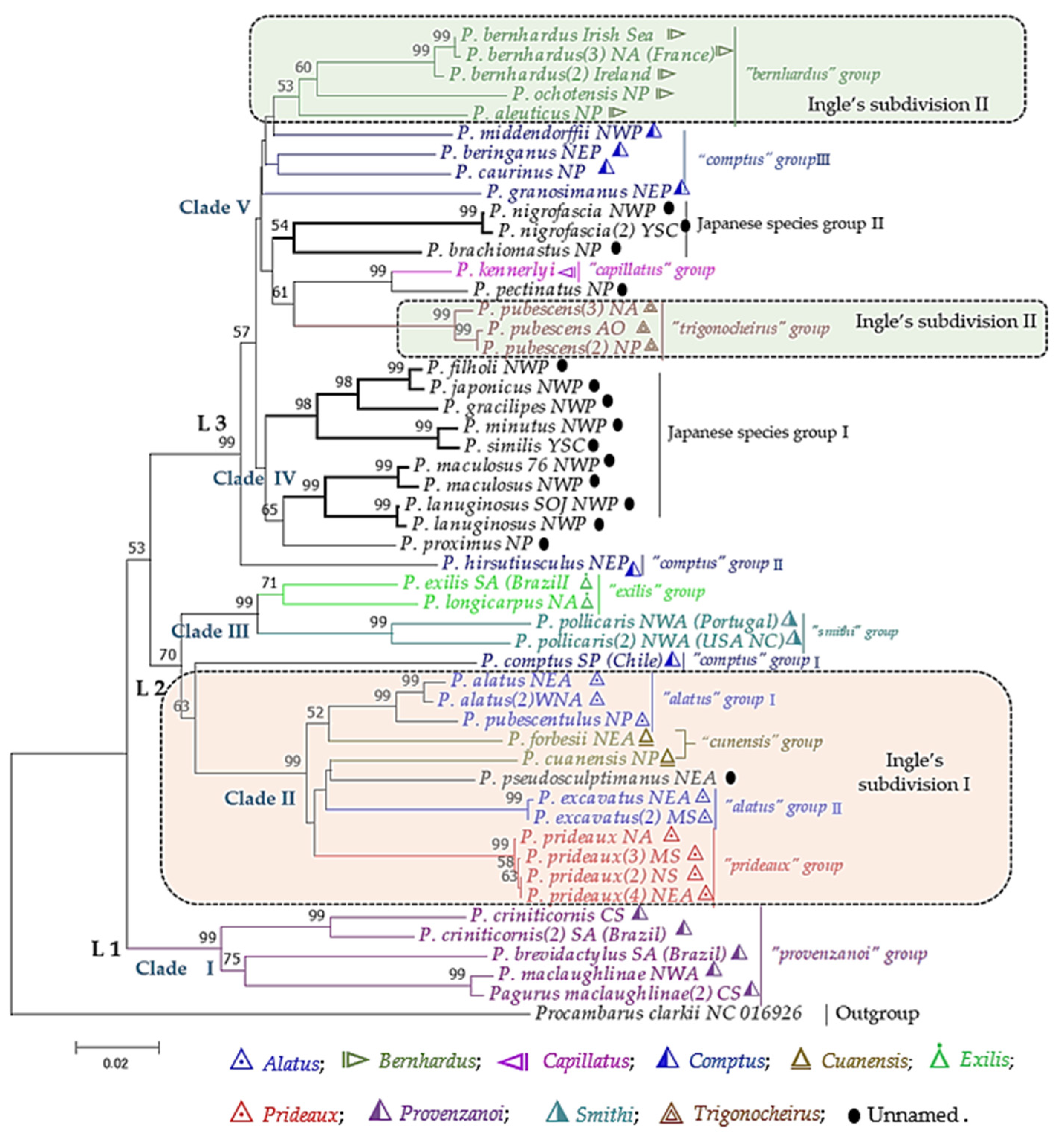
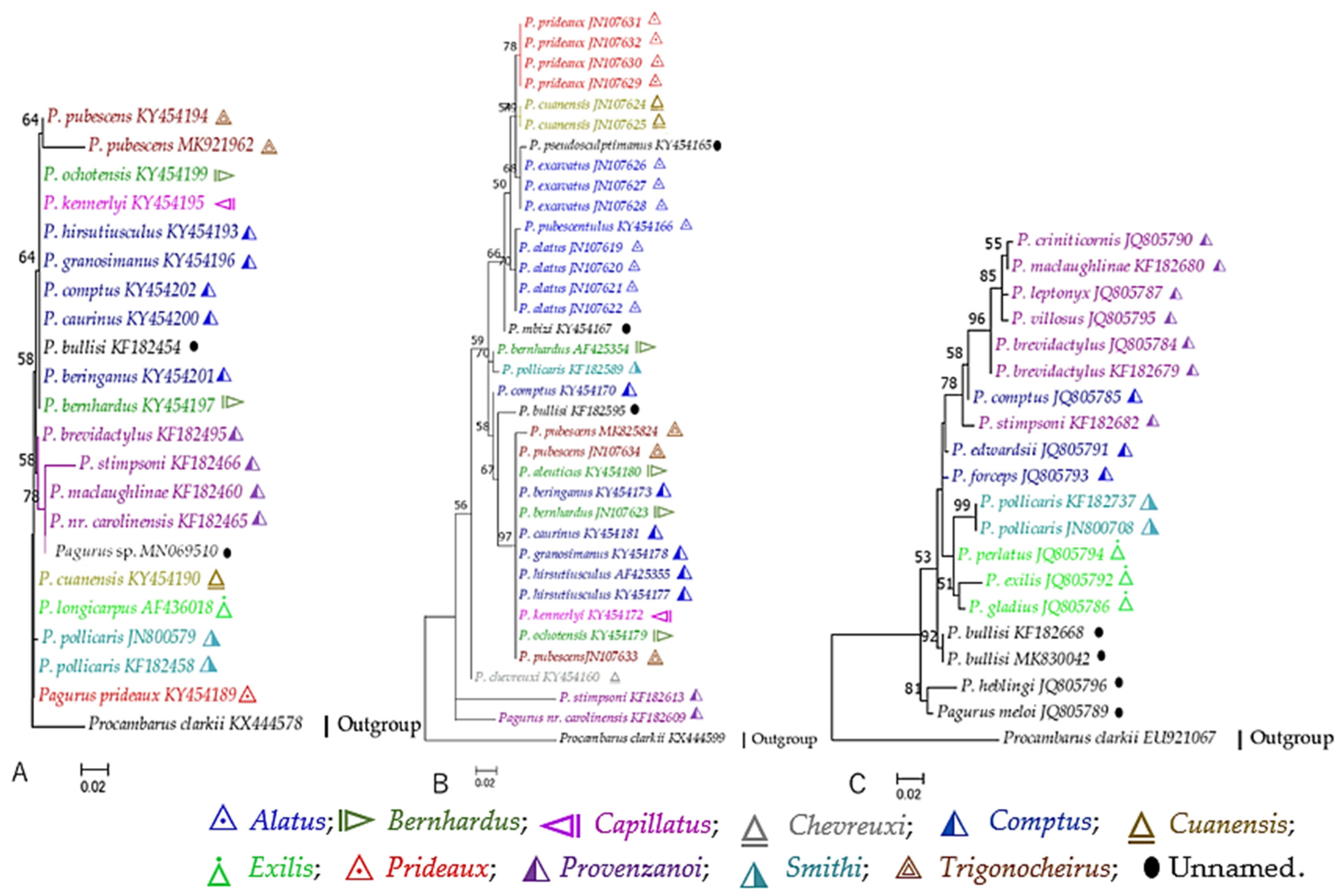
3.2. Phylogenetic Aspects
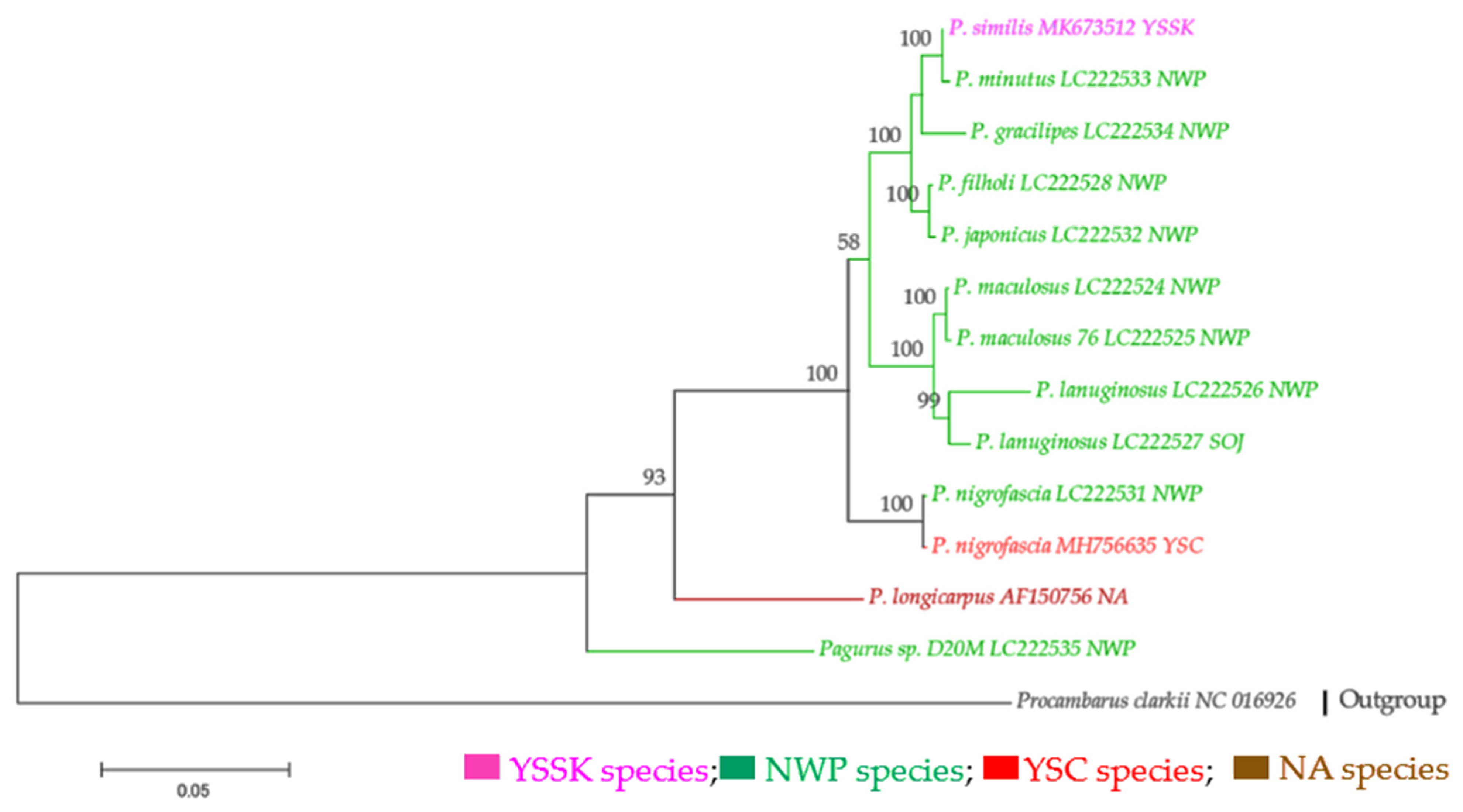
3.2.1. Complete mt Genome Sequences
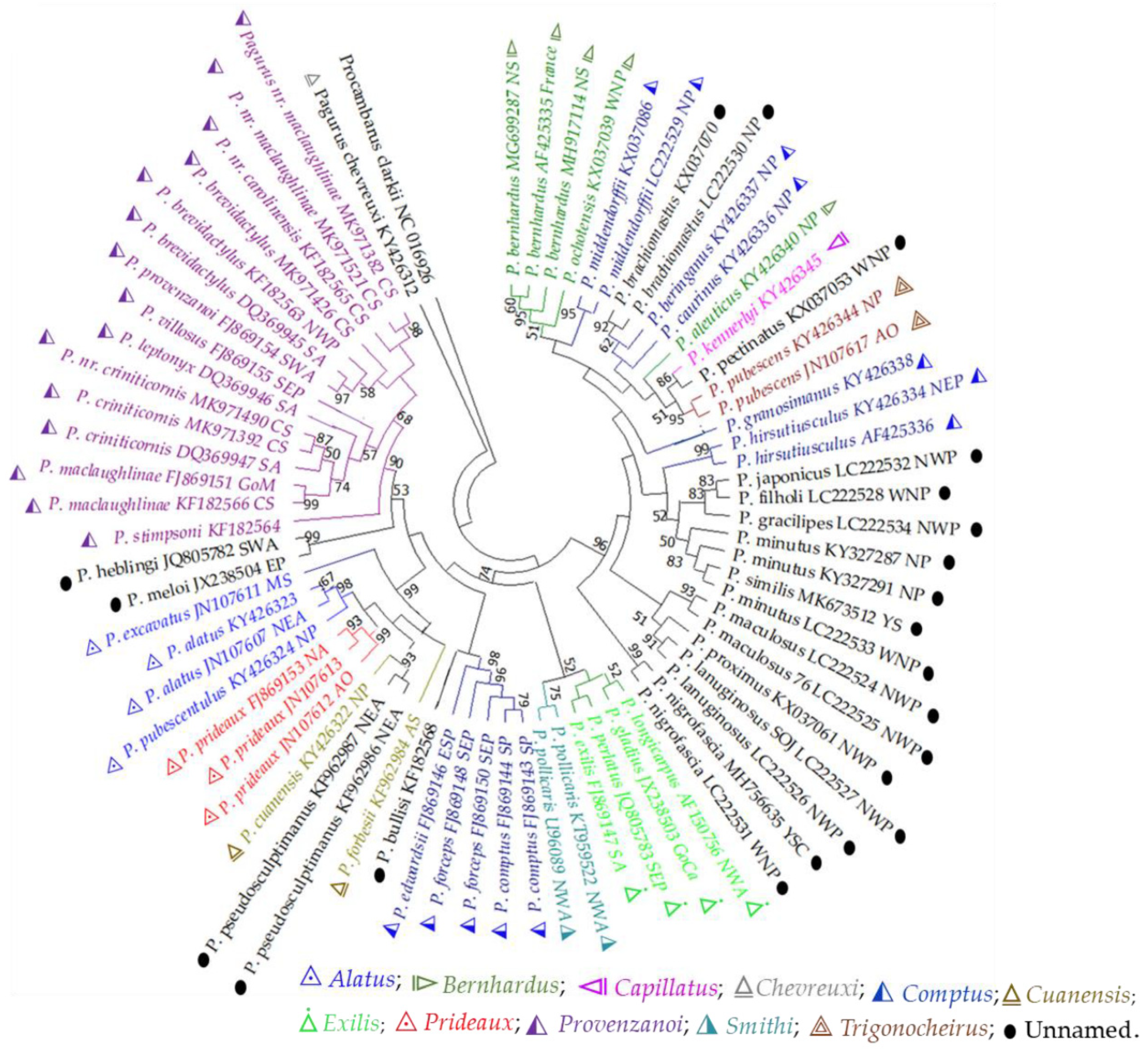
3.2.2. Partial mtDNA Markers
3.2.3. Partial Nuclear DNA Markers
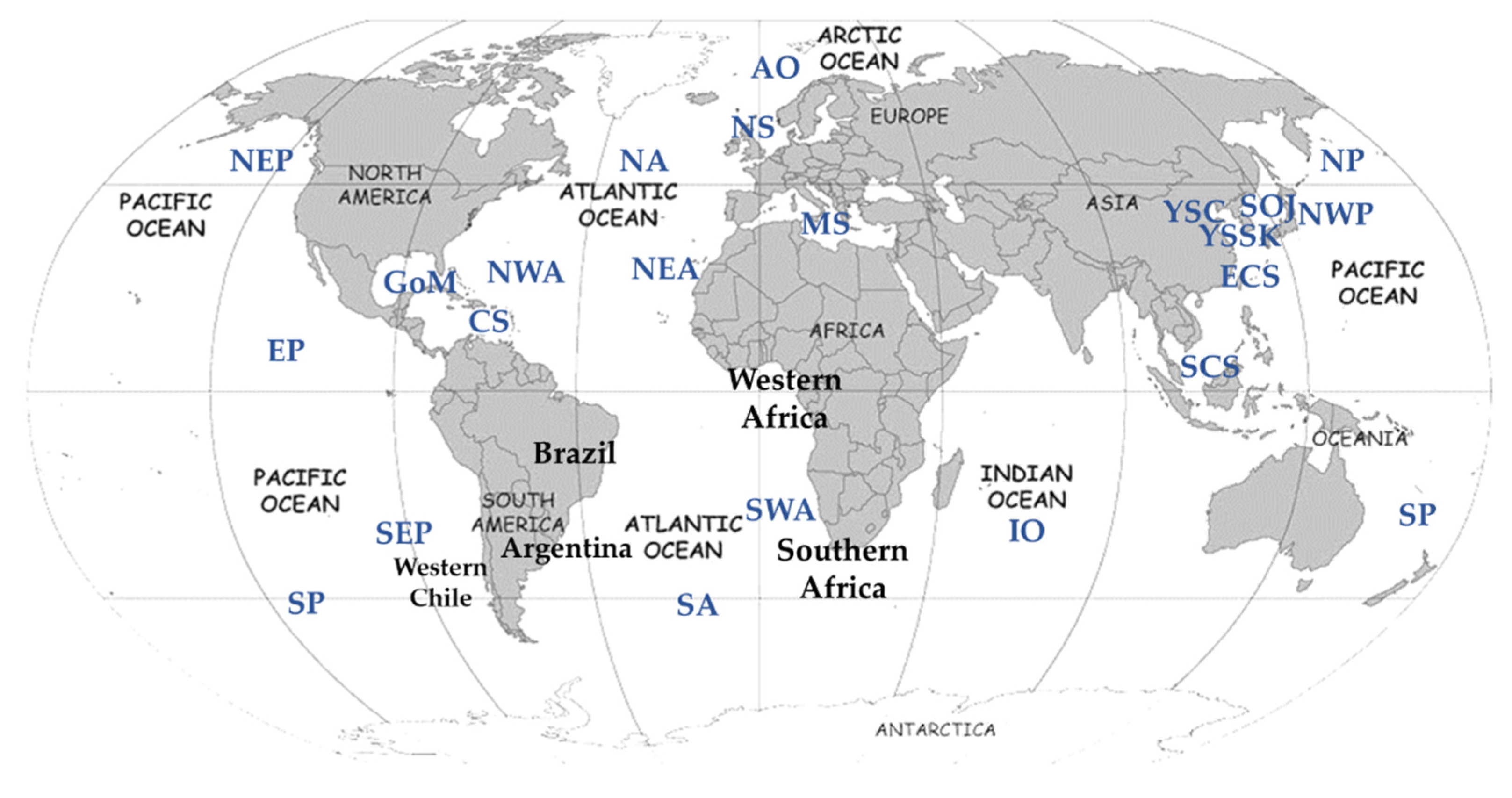
3.3. Genetic Variability among Geographical Regions
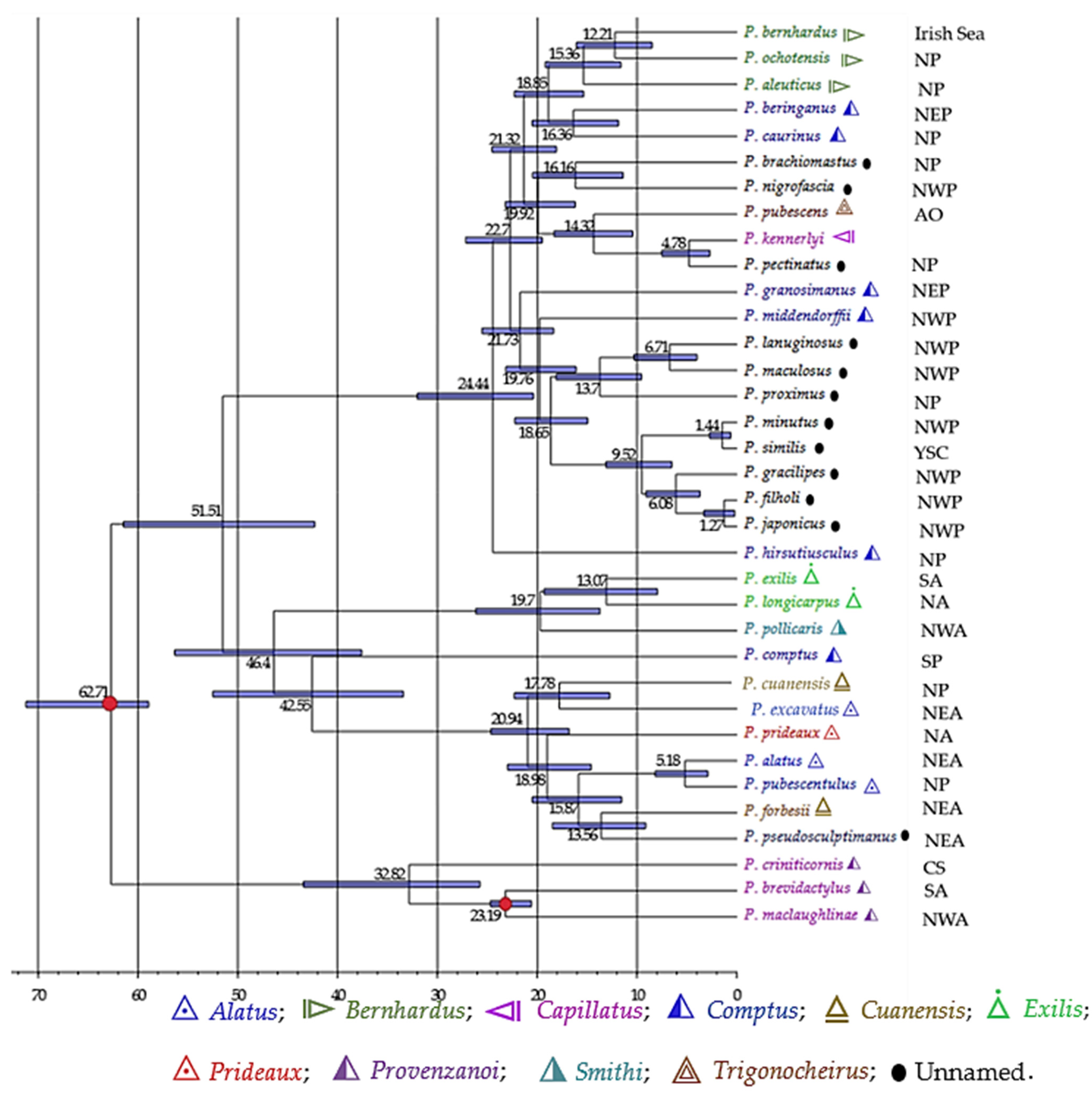
3.4. Geographical Diversification
3.5. Morphological Characteristics within Pagurus Species
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, S. Criteria for recognizing marine hermit crabs in the fossil record using gastropod shells. J. Paleontol. 1992, 66, 535–558. [Google Scholar] [CrossRef]
- Tsang, L.M.; Chan, T.Y.; Ahyong, S.T.; Chu, K.H. Hermit to king, or hermit to all: Multiple transitions to crab-like forms from hermit crab ancestors. Syst. Biol. 2011, 60, 616–629. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.W.; Blackstone, N.W.; Buss, L.W. Evolution of king crab from hermit crab ancestors. Nature 1992, 355, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, J.M.; Luque, J.; Bracken-Grissom, H.D. How to become a crab: Phenotypic constraints on a recurring body plan. BioEssays 2021, 43, 2100020. [Google Scholar] [CrossRef]
- McLaughlin, P.A. A review of the phylogenetic position of the Lomidae (Crustacea, Decapoda, Anomala). J. Crust. Biol. 1983, 3, 431–437. [Google Scholar] [CrossRef]
- McLaughlin, P.A.; Lemaitre, R. Carcinization in the Anomura—Fact or fiction? I. Evidence from adult morphology. Contrib. Zool. 1997, 67, 79–123. [Google Scholar] [CrossRef] [Green Version]
- Tudge, C.C. Spermatophore morphology in the hermit crab families Paguridae and Parapaguridae (Paguroidea, Anomura, Decapoda). Invertebr. Reprod. Dev. 1999, 35, 203–214. [Google Scholar] [CrossRef]
- McLaughlin, P.A.; Lemaitre, R.; Tudge, C.C. Carcinization in the Anomura—Fact or fiction? Part II. Evidence from larval, megalopal and early juvenile morphology. Contrib. Zool. 2004, 73, 165–205. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, P.A.; Lemaitre, R.; Sorhannus, U. Hermit crab phylogeny: A reappraisal and its “fall-out”. J. Crust. Biol. 2007, 27, 97–115. [Google Scholar] [CrossRef] [Green Version]
- Tsang, L.M.; Ma, K.Y.; Ahyong, S.T.; Chan, T.Y.; Chu, K.H. Phylogeny of Decapoda using two nuclear protein-coding genes: Origin and evolution of the Reptantia. Mol. Phylogenet. Evol. 2008, 48, 359–368. [Google Scholar] [CrossRef]
- Lemaitre, R.; McLaughlin, P.A. Recent advances and conflicts in concepts of anomuran phylogeny (Crustacea: Malacostraca). Arthropod Syst. Phylogeny 2009, 67, 119–135. [Google Scholar]
- Lemaitre, R.; McLaughlin, P.A.; Sorhannus, U. Phylogenetic relationships within the Pylochelidae (Decapoda: Anomura: Paguroidea): A cladistics analysis based on morphological characters. Zootaxa 2009, 2022, 1–14. [Google Scholar] [CrossRef]
- Scelzo, M.A.; Fantucci, M.Z.; Mantelatto, F.L. Spermatophore and gonopore morphology of the Southwestern-Atlantic hermit crab Pagurus exilis (Benedict, 1892) (Anomura, Paguridae). Zool. Stud. 2010, 49, 421–433. [Google Scholar]
- Tirelli, T.; Gamba, M.; Pessani, D.; Tudge, C. Spermatophore and spermatozoal ultrastructure of the Mediterranean hermit crab Pagurus excavatus (Paguridae: Anomura: Decapoda). JMBA 2013, 93, 1363–1371. [Google Scholar] [CrossRef]
- Bracken-Grissom, H.D.; Cannon, M.E.; Cabezas, P.; Feldmann, R.M.; Schweitzer, C.E.; Ahyong, S.T.; Felder, D.L.; Lemaitre, R.; Crandall, K.A. A comprehensive and integrative reconstruction of evolutionary history for Anomura (Crustacea: Decapoda). BMC Evol. Biol. 2013, 13, 128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, M.H.; Gan, H.M.; Lee, Y.P.; Linton, S.; Grandjean, F.; Bartholomei-Santos, M.L.; Miller, A.D.; Austin, C.M. ORDER within the chaos: Insights into phylogenetic relationships within the Anomura (Crustacea: Decapoda) from mitochondrial sequences and gene order rearrangements. Mol. Phylogenet. Evol. 2018, 127, 320–331. [Google Scholar] [CrossRef]
- Fabricius, J.C. Systema entomologiae, sistens insectorum classes, ordines, genera, species, adiectis synonymis, locis, descriptionibus, observationibus. Flensbg. Lipsiae Kortii 1775, 832, 1745–1808. [Google Scholar]
- Susana, M.P.; Fran, R. Hermit crabs (Decapoda: Crustacea) from deep Mauritanian waters (NW Africa) with the description of a new species. Zootaxa 2015, 3926, 151–190. [Google Scholar]
- De Grave, S.; Pentcheff, N.D.; Ahyong, S.T.; Chan, T.-Y.; Crandall, K.A.; Dworschak, P.C.; Felder, D.L.; Feldmann, R.M.; Fransen, C.H.J.M.; Goulding, L.Y.D.; et al. A classification of living and fossil genera of decapods crustaceans. Raffles Bull. Zool. 2009, 21, 1–109. [Google Scholar]
- Oba, T.; Goshima, S. Temporal and spatial settlement patterns of sympatric hermit crabs and the influence of shell resource availability. Mar. Biol. 2004, 144, 871–879. [Google Scholar] [CrossRef]
- Gherardi, F.; Nardone, F. The question of coexistence in hermit crabs: Population ecology of a tropical intertidal assemblage. Crustaceana 1997, 70, 608–629. [Google Scholar]
- Biagi, R.; Meireles, A.L.; Mantelatto, F.L. Bio-ecological aspects of the hermit crab Paguristes calliopsis (Crustacea, Diogenidae) from Anchieta Island, Brazil. An. Acad. Bras. Ciênc. 2006, 78, 451–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouvier, E.-L. Décapodes Marcheurs Faune de France, 37; Paul Lechevalier: Paris, France, 1940; 404p. [Google Scholar]
- MacDonald, J.D.; Pike, R.B.; Williamson, D.I. Larvae of the British species of Diogenes, Pagurus, Anapagurus and Lithodes (Crustacea, Decapoda). Proc. Zool. Soc. 1957, 128, 209–257. [Google Scholar] [CrossRef]
- Pike, R.B.; Williamson, D.I. Crustacea, Decapoda: Larvae XI. Paguridea, Coenobitidea, Dromiidea and Homolidea; Zooplankton Identification Sheet; Conseil International pour l’Exploration de la Mer ICES: Copenhagen, Danmai, 1959; Volume 81, pp. 1–9. [Google Scholar]
- McLaughlin, P.A. The hermit crabs (Crustacea, Decapoda, Paguridae) of northwestern North America. Zool. Verh. 1974, 130, 1–396. [Google Scholar]
- McLaughlin, P.A. On the identity of Pagurus brevidactylus (Stimpson) (Decapoda: Paguridae), with the description of a new species of Pagurus from the western Atlantic. Bull. Mar. Sci. 1975, 25, 359–376. [Google Scholar]
- Ingle, R.W. Northeastern Atlantic and Mediterranean hermit crabs (Crustacea: Anomura: Paguroidea: Paguridae). I. The genus Pagurus Fabricius, 1775. J. Nat. Hist. 1985, 19, 745–769. [Google Scholar] [CrossRef]
- McLaughlin, P.A.; Gore, R.H.; Crain, J.A. Studies on the Provenzanoi and other pagurid groups: II. A reexamination of the larval stages of Pagurus hirsutiusculus (Dana) (Decapoda: Anomura: Paguridae) reared in the laboratory. J. Crust. Biol. 1988, 8, 430–450. [Google Scholar] [CrossRef]
- Lemaitre, R.; Castaño, N.C. A new species of Pagurus Fabricius, 1775 from the Pacific coast of Colombia, with a checklist of the eastern Pacific species of the genus. Nauplius 2004, 12, 71–82. [Google Scholar]
- Scotland, R.W.; Olmstead, R.G.; Bennett, J.R. Phylogeny reconstruction: The role of morphology. Syst. Biol. 2003, 52, 539–548. [Google Scholar] [CrossRef]
- Machordom, A.; Macpherson, E. Rapid radiation and cryptic speciation in squat lobsters of the genus Munida (Crustacea, Decapoda) and related genera in the south west Pacific: Molecular and morphological evidence. Mol. Phylogenet. Evol. 2004, 33, 259–279. [Google Scholar] [CrossRef] [PubMed]
- Negri, M.; Pileggi, L.G.; Mantelatto, F.L. Molecular barcode and morphological analyses reveal the taxonomic and biogeographical status of the striped-legged hermit crab species Clibanarius sclopetarius (Herbst, 1796) and Clibanarius vittatus (Bosc, 1802) (Decapoda: Diogenidae). Invert. Syst. 2012, 26, 561–571. [Google Scholar] [CrossRef]
- Hultgren, K.M.; Stachowicz, J.J. Alternative camouflage strategies mediate predation risk among closely related co-occurring kelp crabs. Oecologia 2008, 155, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Bickford, D.; Lohman, D.J.; Sodhi, N.S.; Ng, P.K.L.; Meier, R.; Winker, K.; Ingram, K.K.; Das, I. Cryptic species as a window on diversity and conservation. Trends Ecol. Evol. 2007, 22, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Piffaretti, J.; Vanlerberghe-Masutti, F.; Tayeh, A.; Clamens, A.L.; D’Acier, A.C.; Jousselin, E. Molecular phylogeny reveals the existence of two sibling species in the aphid pest Brachycaudus helichrysi (Hemiptera: Aphididae). Zool. Scr. 2012, 41, 266–280. [Google Scholar] [CrossRef]
- Mantelatto, F.L.; Pardo, L.M.; Pileggi, L.G.; Felder, D.L. Taxonomic re-examination of the hermit crab species Pagurus forceps and Pagurus comptus (Decapoda: Paguridae) by molecular analysis. Zootaxa 2009, 2133, 20–32. [Google Scholar] [CrossRef] [Green Version]
- da Silva, M.J.; dos Santos, A.; Cunha, M.R.; Costa, F.O.; Creer, S.; Carvalho, G.R. Multigene molecular systematics confirm species status of morphologically convergent Pagurus hermit crabs. PLoS ONE 2011, 6, e28233. [Google Scholar]
- Olguín, N.; Mantelatto, F.L. Molecular analysis validates of some informal morphological groups of Pagurus (Fabricius, 1775) (Anomura: Paguridae) from South America. Zootaxa 2013, 3666, 436–448. [Google Scholar] [CrossRef]
- Lemaitre, R.; Tavares, M. New taxonomic and distributional information on hermit crabs (Crustacea: Anomura: Paguroidea) from the Gulf of Mexico, Caribbean Sea, and Atlantic coast of South America. Zootaxa 2015, 3994, 451–506. [Google Scholar] [CrossRef] [Green Version]
- Sultana, Z.; Asakura, A.; Kinjo, S.; Nozawa, M.; Nakano, T.; Ikeo, K. Molecular phylogeny of ten intertidal hermit crabs of the genus Pagurus inferred from multiple mitochondrial genes, with special emphasis on the evolutionary relationship of Pagurus lanuginosus and Pagurus maculosus. Genetica 2018, 146, 369–381. [Google Scholar] [CrossRef]
- Hwang, J.-Y.; Haque, N.; Lee, D.-H.; Kim, B.-M.; Rhee, J.-S. Complete mitochondrial genome of the intertidal hermit crab, Pagurus similis (Crustacea, Anomura). Mitochondrial DNA B 2019, 4, 1861–1862. [Google Scholar] [CrossRef]
- Gong, L.; Jiang, H.; Zhu, K.; Lu, X.; Liu, L.; Liu, B.; Jiang, L.; Ye, Y.; Lü, Z. Large-scale mitochondrial genere arrangements in the hermit crab Pagurus nigrofascia and phylogenetic analysis of the Anomura. Gene 2019, 695, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Landschoff, J.; Gouws, G. DNA barcoding as a tool to facilitate the taxonomy of hermit crabs (Decapoda: Anomura: Paguroidea). J. Crust. Biol. 2018, 38, 780–793. [Google Scholar] [CrossRef] [Green Version]
- Takezaki, N.; Gojobori, T. Correct and incorrect vertebrate phylogenies obtained by the entire mitochondrial DNA sequences. Mol. Biol. Evol. 1999, 15, 727–737. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–442. [Google Scholar]
- Kimura, M.A. simple method for estimating evolutionary rate of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Felsenstein, J. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 1985, 39, 783–791. [Google Scholar] [CrossRef]
- Hedges, S.B.; Kumar, S. The Timetree of Life; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Yang, J.-S.; Lu, B.; Chen, D.-F.; Yu, Y.-Q.; Yang, F.; Nagasawa, H.; Tsuchida, S.; Fujiwara, Y.; Yang, W.-J. When did decapods invade hydrothermal vents? Clues from the western Pacific and Indian oceans. Mol. Biol. Evol. 2013, 30, 305–309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Z. PAML 4: A program package for phylogenetic analysis by maximum likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, J.G.; Miya, M.; Lam, K.; Tay, B.H.; Danks, J.A.; Bell, J.; Walker, T.I.; Venkatesh, B. Evolutionary origin and phylogeny of the modern holocephalans (Chondrichthyes: Chimaeriformes): A mitogenomic perspective. Mol. Biol. Evol. 2010, 27, 2576–2586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babarinde, I.A.; Saitou, N. The dynamics, causes, and impacts of mammalian evolutionary rates revealed by the analyses of Capybara draft genome sequences. Genome Biol. Evol. 2020, 12, 1444–1458. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, J.E.; Cuesta, J.A.; Raso, J.E. Taxonomic study of the Pagurus forbesii “complex” (Crustacea: Decapoda: Paguridae). Description of Pagurus pseudosculptimanus sp. nov. from Alborán Sea (Southern Spain, Western Mediterranean Sea). Zootaxa 2014, 3753, 25–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komai, T. The identity of Pagurus brachiomastus and descriptions of two new species of Pagurus (Crustacea: Decapoda: Anomura: Paguridae) from the Northwestern Pacific. Species Divers. 2000, 5, 229–265. [Google Scholar] [CrossRef] [Green Version]
- Komai, T.; Imafuku, M. Redescription of Pagurus lanuginosus with the establishment of a neotype, and description of a new closely related species (Decapoda: Anomura: Paguridae). J. Crust. Biol. 1996, 16, 782–796. [Google Scholar] [CrossRef] [Green Version]
- Komai, T. Identities of Pagurus japonicus (Stimpson, 1858) and P. similis (Ortmann, 1892), with description of a new species of Pagurus. Zoosystema 2003, 25, 377–411. [Google Scholar]
- Noever, C.; Glenner, H. The origin of king crabs: Hermit crab ancestry under the magnifying glass. Zool. J. Linn. Soc. 2017, 182, 300–318. [Google Scholar] [CrossRef]
- Landschoff, J.; Komai, T.; du Plessis, A.; Gouws, G.; Griffiths, C.L. MicroCT imaging applied to description of a new species of Pagurus Fabricius, 1775 (Crustacea: Decapoda: Anomura: Paguridae), with selection of three-dimensional type data. PLoS ONE 2018, 13, e0203107. [Google Scholar]
- Trivedi, J.N.; Vachhrajani, K.D. An annotated checklist of hermit crabs (Crustacea, Decapoda, Anomura) of Indian waters with three new records. J. Asia Pac. Biodivers. 2017, 10, 175–182. [Google Scholar] [CrossRef]
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luiz, O.J.; Madin, J.S.; Robertson, D.R.; Rocha, L.; Wirtz, P.; Floeter, S.R. Ecological traits influencing range expansion across large oceanic dispersal barriers: Insights from tropical Atlantic reef fishes. Proc. Biol. Sci. 2012, 279, 1033–1040. [Google Scholar] [CrossRef] [PubMed]
- Hellberg, M.E. Gene flow and isolation among populations of marine animals. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 291–310. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| References | Ingle (1985) [28] | McLaughlin et al. (1988) [29] | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species groups of the genus Pagurus | Subdivision I | Subdivision II | ||||||||||||||||
| Group 1 “Alatus” | Group 2 “Prideaux” | Group 3 “Cuanensis” | Group 1 (Group V) “Bernhardus” | Group 2 (Group VIII) “Trigonocheirus” | Group 3 (Group XI) “Anachoretus” | Group 4 “Chevreuxi” | Group 5 “Carneus” | Group I “Provenzanoi” | Group II “Exilis” | Group III “Comptus” | Group IV “Gaudichaudii” | Group VI “Capillatus” | Group VII “Confragosus” | Group X “Smithi” | Group IX “Unnamed” | Group XII “Unnamed” | ||
| Species assigned in the groups | P. alatus | P. prideaux | P. cuanensis | P. aleuticus | P. brandti | P. anachoretoides | P. chevreuxi | P. carneus | P. annexus* | P. albus* | P. beringianus | P. gaudichaudii | P. arcuatus | P. confragosus | P. impressus | P. delsolari | P. alatus | |
| P. excavatus | P. pycnacanthus | P. forbesii | P. armatus | P. dalli | P. anachoretus | P. arenisaxatilis* | P. gladius | P. caurinus | P. capillatus | P. cornutus | P. pollicaris | P. imarpe | P. cuanensis | |||||
| P. pubescentulus | P. bernhardus | P. pubescens | P. gordonae | P. benedicti | P. limatulus | P. comptus | P. kennerlyi | P. tanneri | P. smithi | P. ruber | ||||||||
| P. pulchellus | P. ochotensis | P. stevensae | P. laurentae | P. brevidactylus | P. longicarpus | P. edwardsii | P. setosus | P. spilocarpus | P. sculptimanus | |||||||||
| P. trigonocheirus | P. souriei | P. carolinensis | P. mertensii | P. forceps | P. pubescentulus | |||||||||||||
| P. undosus | P. triangularis | P. criniticornis | P. perlatus* | P. granosimanus | P. prideaux | |||||||||||||
| P. galapagensis | P. hemphilli | P. variailis | ||||||||||||||||
| P. gymnodactylus | P. hirsutiusculus | |||||||||||||||||
| P. lepidus | P. middendorffii | |||||||||||||||||
| P. leptonyx | P. samuelis | |||||||||||||||||
| P. maclaughlinae | P. quaylei | |||||||||||||||||
| P. nanodes* | ||||||||||||||||||
| P. nesiotes* | ||||||||||||||||||
| P. protuberocarpus | ||||||||||||||||||
| P. provenzanoi | ||||||||||||||||||
| P. redondoensis | ||||||||||||||||||
| P. retrorsimanus* | ||||||||||||||||||
| P. rhabdotus* | ||||||||||||||||||
| P. spighti* | ||||||||||||||||||
| P. stimpsoni | ||||||||||||||||||
| P. trichocerus | ||||||||||||||||||
| P. vetaultae* | ||||||||||||||||||
| P. villosus | ||||||||||||||||||
| P. virgulatus* | ||||||||||||||||||
| Compiled species groups of the genus Pagurus | Group 1 “Alatus” | Group 2 “Prideaux” | Group 3 “Cuanensis” | Group 4 “Bernhardus” | Group 5 “Trigonocheirus” | Group 6 “Anachoretus” | Group 7 “Chevreuxi” | Group 8 “Carneus” | Group 9 “Provenzanoi” | Group 10 “Exilis” | Group 11 “Comptus” | Group 12 “Gaudichaudii” | Group 13 “Capillatus” | Group 14 “Confragosus” | Group 15 “Smithi” | Group 16 “Unnamed” (combined of Group IX and XII) | Ungrouped Species | |
| Species assigned in the groups | P. alatus | P. prideaux | P. cuanensis | P. aleuticus | P. brandti | P. anachoretoides | P. chevreuxi | P. carneus | P. annexus* | P. albus* | P. beringianus | P. gaudichaudii | P. arcuatus | P. confragosus | P. impressus | P. delsolari | Pagurus acadianus | |
| P. excavatus | P. pycnacanthus | P. forbesii | P. armatus | P. dalli | P. anachoretus | P. arenisaxatilis* | P. gladius | P. caurinus | P. capillatus | P. cornutus | P. pollicaris | P. hartae* | Pagurus boriaustraliensis | |||||
| P. pubescentulus | P. bernhardus | P. pubescens | P. gordonae | P. benedicti | P. limatulus | P. comptus | P. kennerlyi | P. tanneri | P. smithi | P. imarpe | Pagurus brachiomastus | |||||||
| P. pulchellus | P. ochotensis | P. stevensae | P. laurentae | P. brevidactylus | P. longicarpus | P. edwardsii | P. setosus | P. spilocarpus | P. meloi* | Pagurus bullisi | ||||||||
| P. trigonocheirus | P. souriei | P. carolinensis | P. mertensii | P. forceps | P. rathbuni* | Pagurus constans | ||||||||||||
| P. undosus | P. triangularis | P. criniticornis | P. perlatus* | P. granosimanus | P. retrorsimanus* | Pagurus emmersoni | ||||||||||||
| P. galapagensis | P. hemphilli | P. ruber | Pagurus filholi | |||||||||||||||
| P. gymnodactylus | P. hirsutiusculus | P. sculptimanus | Pagurus fraserorum | |||||||||||||||
| P. lepidus | P. middendorffii | P. townsendi* | Pagurus gracilipes | |||||||||||||||
| P. leptonyx | P. samuelis | P. variailis | Pagurus heblingi | |||||||||||||||
| P. maclaughlinae | P. quaylei | P. venturensis* | Pagurus japonicus | |||||||||||||||
| P. nanodes* | Pagurus kulkarni | |||||||||||||||||
| P. nesiotes* | Pagurus lanuginosus | |||||||||||||||||
| P. protuberocarpus | Pagurus liochele | |||||||||||||||||
| P. provenzanoi | Pagurus maculosus | |||||||||||||||||
| P. redondoensis | Pagurus mbizi | |||||||||||||||||
| P. retrorsimanus* | Pagurus minutus | |||||||||||||||||
| P. rhabdotus* | Pagurus nigrivittatus | |||||||||||||||||
| P. spighti* | Pagurus nigrofascia | |||||||||||||||||
| P. stimpsoni | Pagurus pectinatus | |||||||||||||||||
| P. trichocerus | Pagurus pitagsaleei | |||||||||||||||||
| P. vetaultae* | Pagurus proximus | |||||||||||||||||
| P. villosus* | Pagurus pseudosculptimanus | |||||||||||||||||
| P. virgulatu | Pagurus quinquelineatus | |||||||||||||||||
| Pagurus rectidactylus | ||||||||||||||||||
| Pagurus rubrior | ||||||||||||||||||
| Pagurus similis | ||||||||||||||||||
| Pagurus simulans | ||||||||||||||||||
| Pagurus spina | ||||||||||||||||||
| NEA | NWA | NP | NEP | NA | SA | SP | CS | MS | NWP | NWA | YSC | NS | AO | |
| NEA | ||||||||||||||
| NWA | 0.06 | |||||||||||||
| NP | 0.14 | 0.12 | ||||||||||||
| NEP | 0.16 | 0.15 | 0.11 | |||||||||||
| NA | 0.14 | 0.13 | 0.11 | 0.12 | ||||||||||
| SA | 0.15 | 0.14 | 0.15 | 0.15 | 0.15 | |||||||||
| SP | 0.14 | 0.13 | 0.15 | 0.15 | 0.15 | 0.15 | ||||||||
| CS | 0.17 | 0.16 | 0.16 | 0.17 | 0.16 | 0.12 | 0.16 | |||||||
| MS | 0.08 | 0.09 | 0.15 | 0.17 | 0.14 | 0.16 | 0.14 | 0.17 | ||||||
| NWP | 0.16 | 0.14 | 0.12 | 0.11 | 0.12 | 0.15 | 0.16 | 0.17 | 0.16 | |||||
| NWA | 0.16 | 0.16 | 0.16 | 0.16 | 0.16 | 0.14 | 0.15 | 0.14 | 0.16 | 0.17 | ||||
| YSC | 0.17 | 0.15 | 0.11 | 0.10 | 0.12 | 0.15 | 0.15 | 0.16 | 0.17 | 0.10 | 0.16 | |||
| NS | 0.08 | 0.09 | 0.15 | 0.17 | 0.13 | 0.16 | 0.15 | 0.18 | 0.05 | 0.16 | 0.16 | 0.17 | ||
| AO | 0.17 | 0.15 | 0.10 | 0.10 | 0.10 | 0.15 | 0.17 | 0.17 | 0.18 | 0.11 | 0.17 | 0.10 | 0.17 |
| Common features | Morpho-traits | Ingle (1985) Subdivision I | Ingle (1985) Subdivision II | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Maxillule (endopod outer distal margin) | rounded or obtusely produced | produced as a conspicuous lobe or papilla, sometimes recurved, rerely segmented | ||||||||||||||
| Pereiopod 3 (anterior lobe of sternite) | triangular to sub-triangular, and setose with acute to subacute spines | broadly subquadrate to suboval, and setos, smooth | ||||||||||||||
| Telson (posterior margin) | armed with teeth that extend on to lateral margin | armed with teeth but that not extend on to lateral margin | ||||||||||||||
| Male’s pleopod (number) | 0-4 unpaired | 3 unpaired | ||||||||||||||
| Species groups | Group 1 “Alatus” | Group 2 “Prideaux” | Group 3 “Cuanensis” | Group 4 “Bernhardus” | Group 5 “Trigonocheirus” | Group 6 “Anachoretus” | Group 7 “Chevreuxi” | Group 8 “Carneus” | Group 9 “Provenzanoi” | Group 10 “Exilis” | Group 11 “Comptus” | Group 12 “Gaudichaudii” | Group 13 “Capillatus” | Group 14 “Confragosus” | Group 15 “Smithi” | |
| Diagnostic features | ocular peduncles | stout, noticeably subcylindrical, | stout, noticeably subcylindrical, | long and cylindrical | short, moderatelystout | subcylindrical | / | / | / | subcylindrical | strong | narrow, subcylindrical | / | cylindrical | moderately short, stout | / |
| ocular acicles | spatulate to subtriangular, dorsal surface concave | spatulate, dorsal surface relatively flat | spatulate to subtriangular, dorsal surface slightly concave | spatulate, dorsal surface concave | subtriangular, dorsal surface flat to slightly concave | subtriangular, dorsal surface flat | subtriangular, dorsal surface flat | narrowly subtriangular, dorsal surface slightly concave | broad, denticulate terminally | triangular, dorsal surface concave | triangular, with subterminal spine inserted ventrally | / | acutely or ovately triangular | ovately triangular | / | |
| Maxilliped 1 (peduncle of exopod) | moderately broadened proximally | slightly broadened proximally | broadened proximally | broadened proximally | moderately broadened proximally | slightly broadened proximally | / | broadened proximally | very narrow | strongly inflated proximally | / | / | slender | moderately inflated | / | |
| Maxilliped 3 | outer distal margin of corpus and merus with a spine, crista dentata with many acute teeth | outer distal margin of corpus and merus with a spine; crista dentata with moderate number of acute teeth | outer distal margin of corpus and merus without spines; crista dentata with many teeth | endopod, inner margin and outer distal margin of merus each with a spine; crista dentata with moderate number of teeth | endopod, inner margin and outer distal margin of merus each with a spine; crista dentata with many teeth | endopod, outer distal margin of merus without or with a spine; crista dentata with moderate number of teeth | endopod, inner and outer distal margin of merus with a spine; crista dentata with numerous small teeth | outer margin of merus with a spine; crista dentata with numerous small teeth | outer margin of merus unarmed; crista dentata with one tooth | outer margin of merus with small spines; crista dentata with few teeth | / | / | outer margin of merus with spines; crista dentata with 1 or 2 teeth | merus and carpus each with spin, crista dentata with 1 tooth | / | |
| Left cheliped (propodal outer surface) | medially elevated and with prominent longitudinal row of acute tubercles | slightly elevated and, with scattered tubercles | medially elevated and, with acute to subacute tubercles | gently convex and tuberculate | medially elevated with spiniform tubercles | convex and relatively smooth | relatively flat | a longitudinal median elevation bearing tubercles | with spiny tubercles and numerus bristles | compressed laterally. | / | / | oblique, not produced into prominent ridge | strongly oblique, concave, elevated into prominent ridge | / | |
| pereiopods 2, 3 (dactyls) | slightly obliquely compressed distally | obliquely compressed distally | slightly compressed distally not obliquely | obliquely compressed distally | obliquely compressed distally | not obliquely compressed distally | not obliquely compressed distally | slightly obliquely compressed distally | terminating in sharp, corneous claw | long, arched, ventral margin with spiniform bristles | short, ornamented by strong, spiniform bristles on ventral margin | / | straight or twisted, with setos | moderately long, slender, usually twisted | / | |
| Telson (posterior margin ) | biconcave, many teeth extending on to lateral margin | concave, many teeth extending on to lateral margin | biconcave, and small to medium size teeth extending on to lateral margin | concave, with a few small teeth | biconcave, with a few large, recurved teeth | biconcave, with many or a few small teeth | biconcave, with numerous small teeth | concave, with large subacute teeth | lest uropod bordered with long, ribbon-like bristles | difurcated lobes, spinous, without bristles | separated by fine median incision, denticulated | / | oblique, with 2-8 strong spines | subtriangular, oblique, denticulate or with small spinules | / | |
| Male’s pleopod (number) | 3 | 4 | 4 | 3 | 3 | / | / | / | 3 | 3 | / | / | 3 | 3 | / | |
| Reference Literature | Ingle (1985) [28] | Mclaughlin (1974) [26] | ||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sultana, Z.; Babarinde, I.A.; Asakura, A. Diversity and Molecular Phylogeny of Pagurid Hermit Crabs (Anomura: Paguridae: Pagurus). Diversity 2022, 14, 141. https://doi.org/10.3390/d14020141
Sultana Z, Babarinde IA, Asakura A. Diversity and Molecular Phylogeny of Pagurid Hermit Crabs (Anomura: Paguridae: Pagurus). Diversity. 2022; 14(2):141. https://doi.org/10.3390/d14020141
Chicago/Turabian StyleSultana, Zakea, Isaac Adeyemi Babarinde, and Akira Asakura. 2022. "Diversity and Molecular Phylogeny of Pagurid Hermit Crabs (Anomura: Paguridae: Pagurus)" Diversity 14, no. 2: 141. https://doi.org/10.3390/d14020141
APA StyleSultana, Z., Babarinde, I. A., & Asakura, A. (2022). Diversity and Molecular Phylogeny of Pagurid Hermit Crabs (Anomura: Paguridae: Pagurus). Diversity, 14(2), 141. https://doi.org/10.3390/d14020141