
Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review




Abstract
:1. Introduction
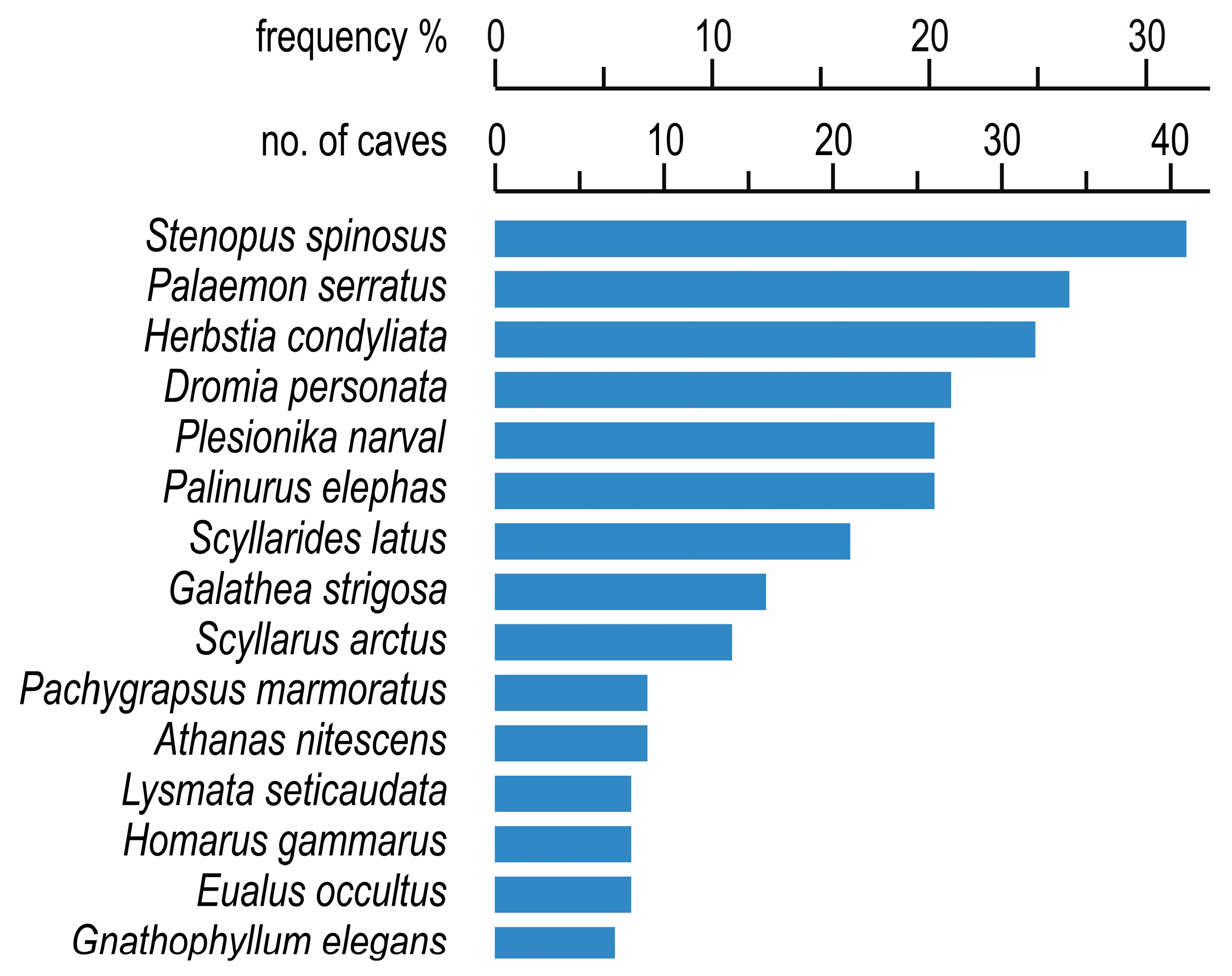
2. Species Inventory and Frequency
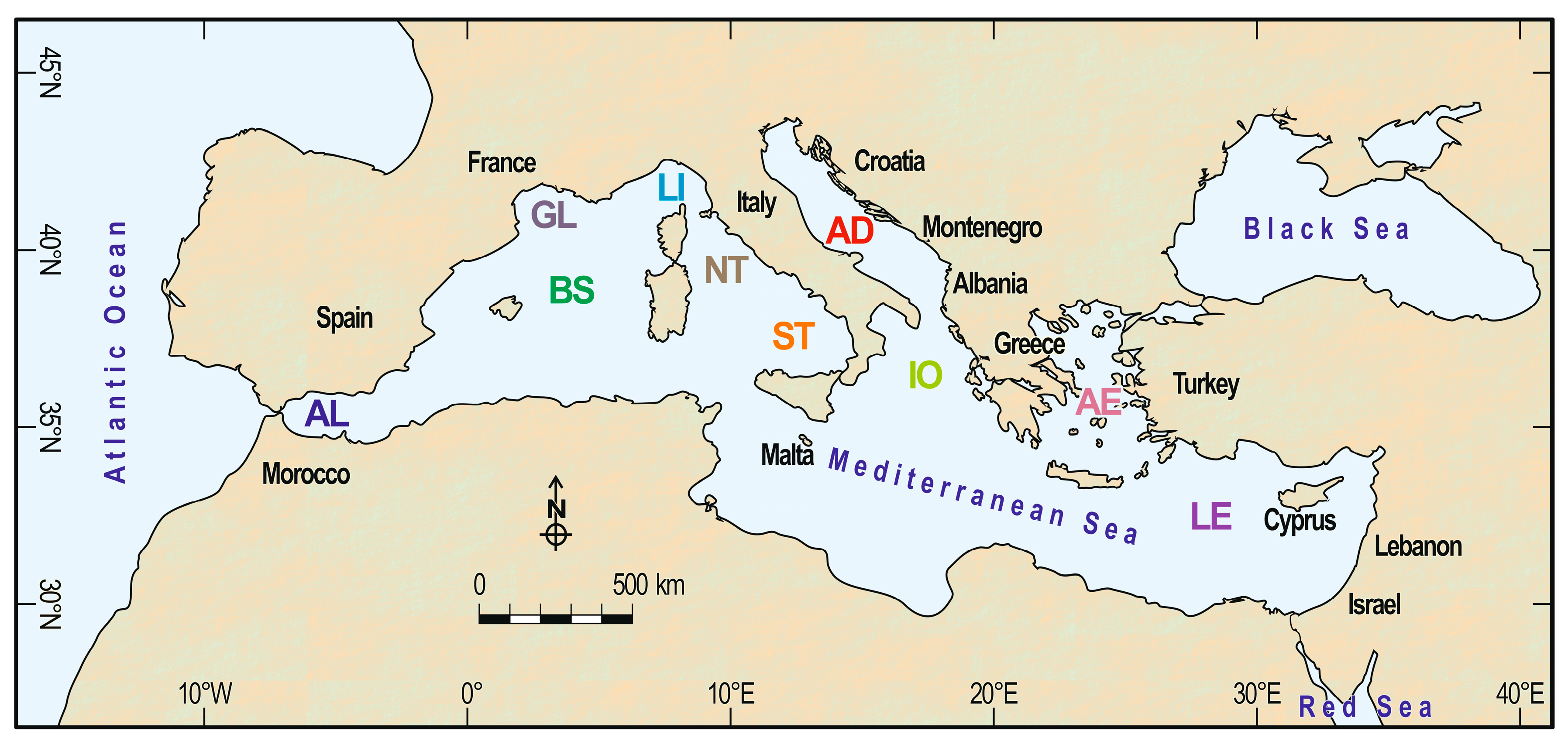
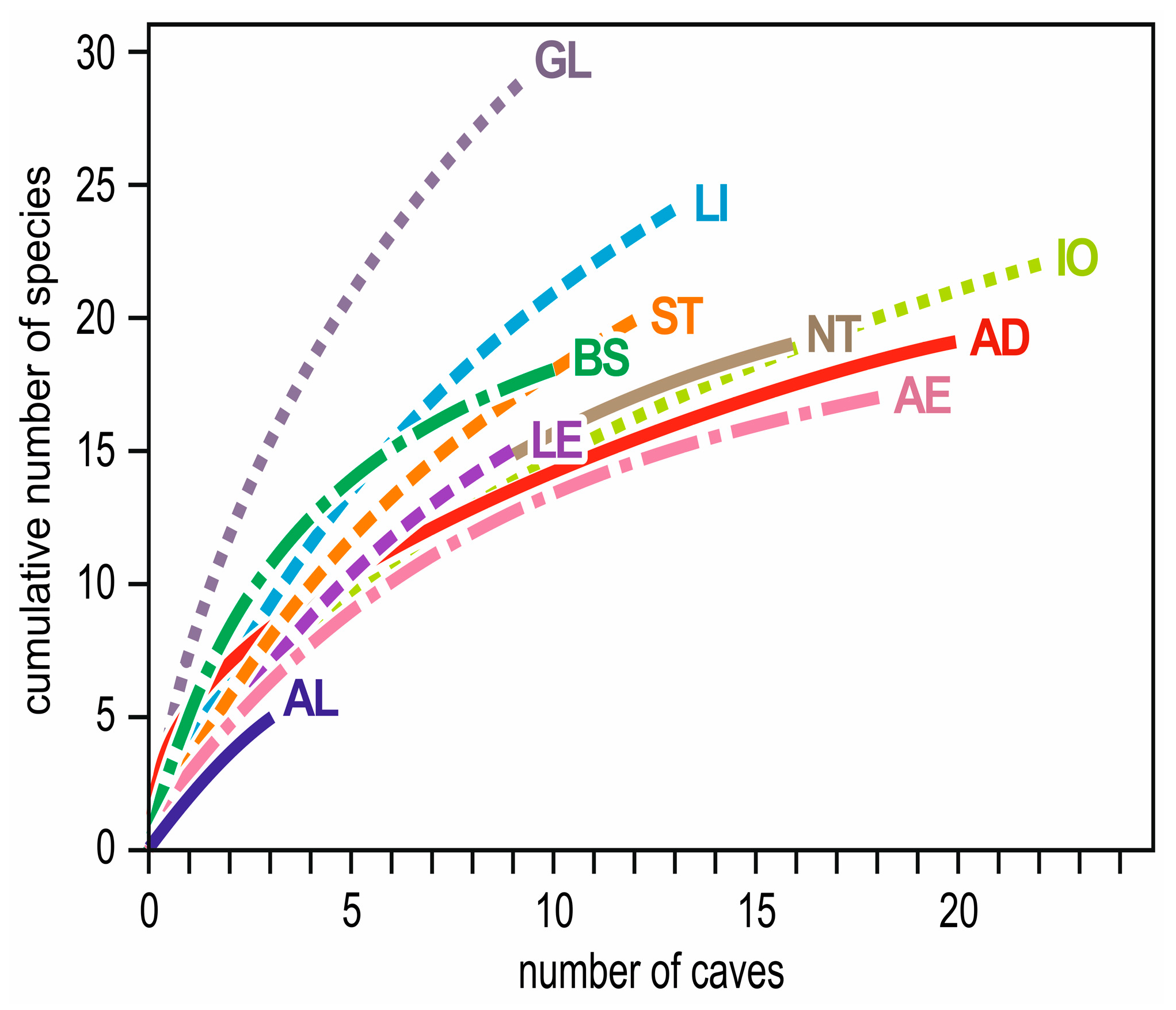
3. Regional Variations in Species Richness
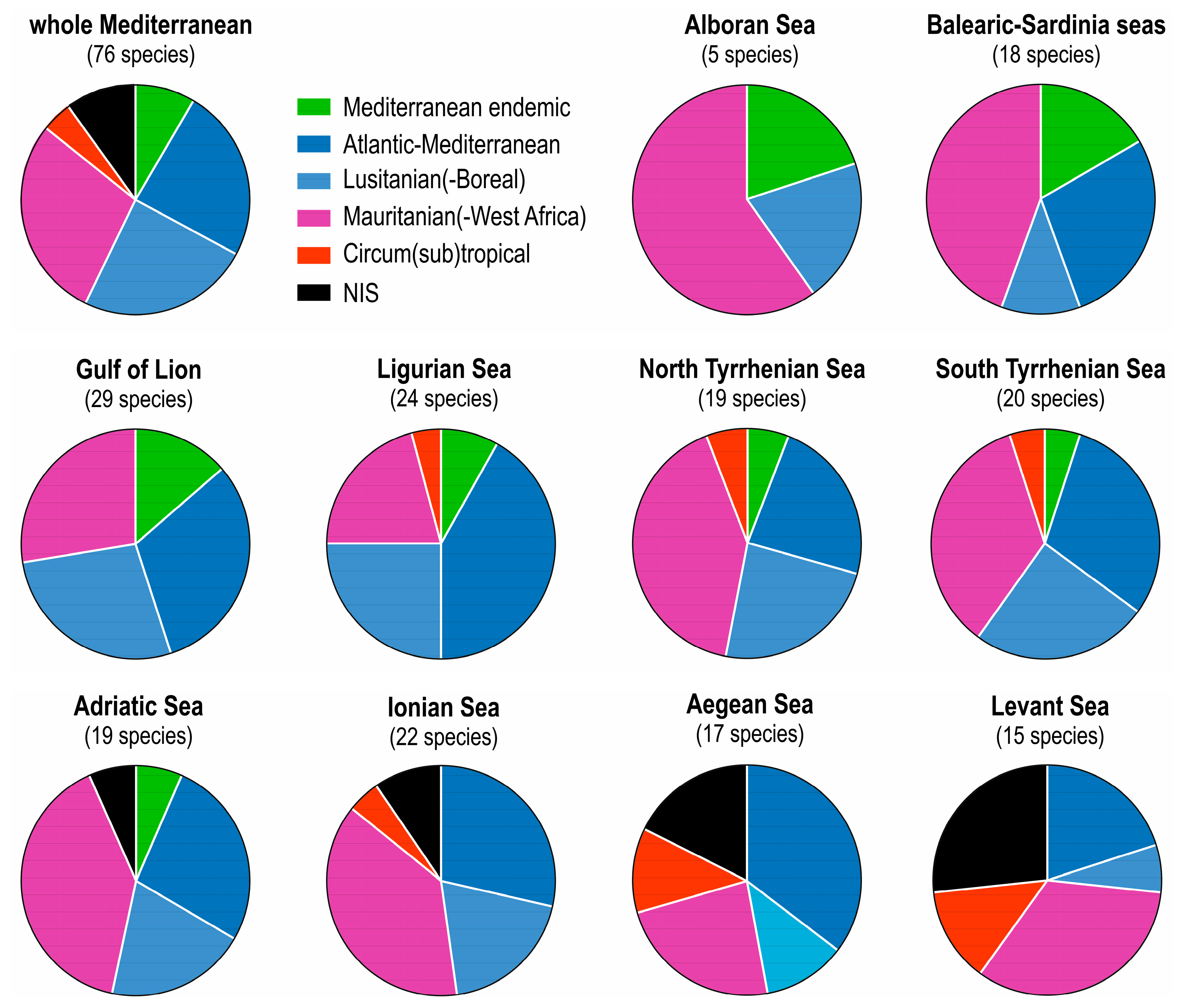
4. Zoogeography
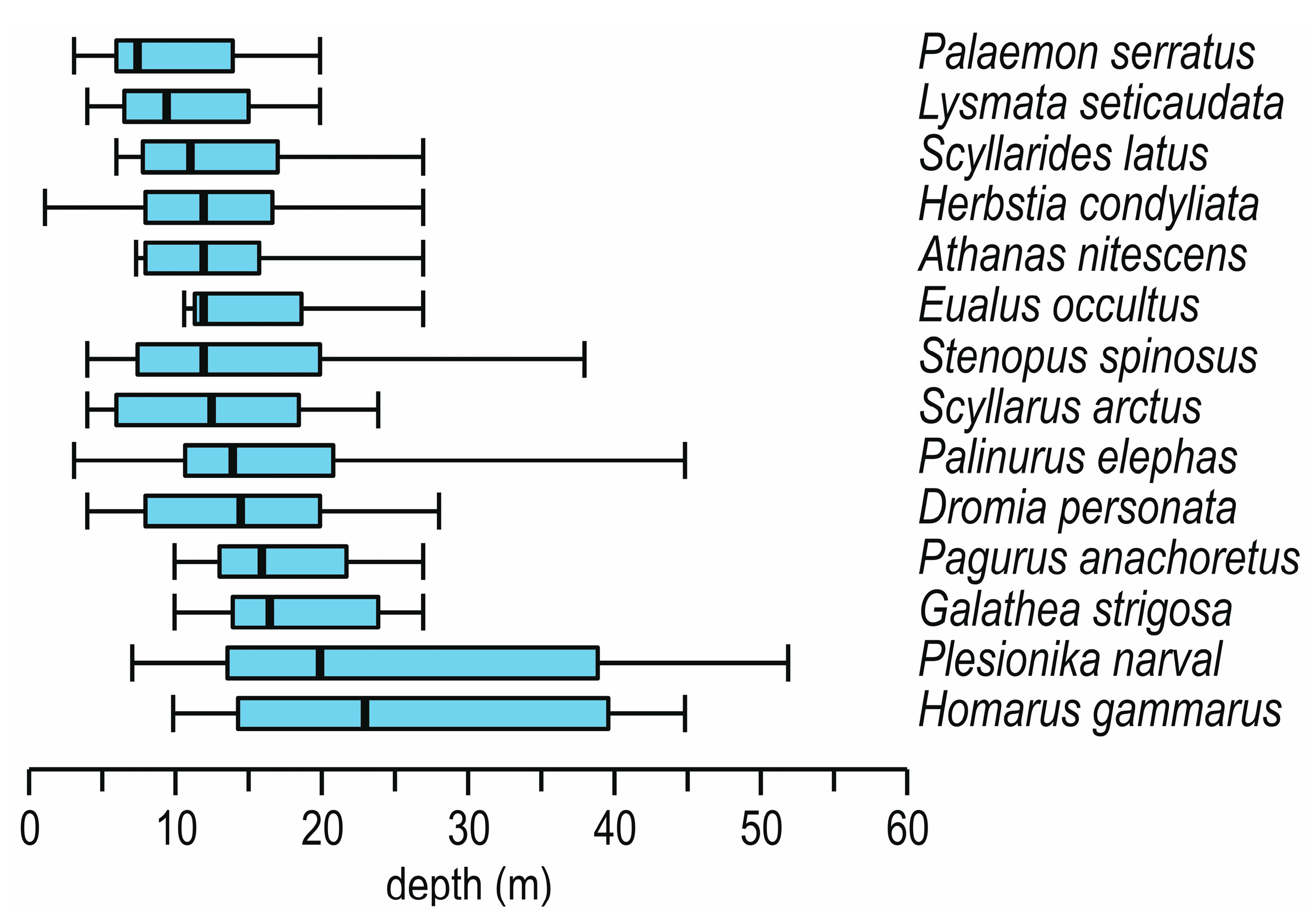
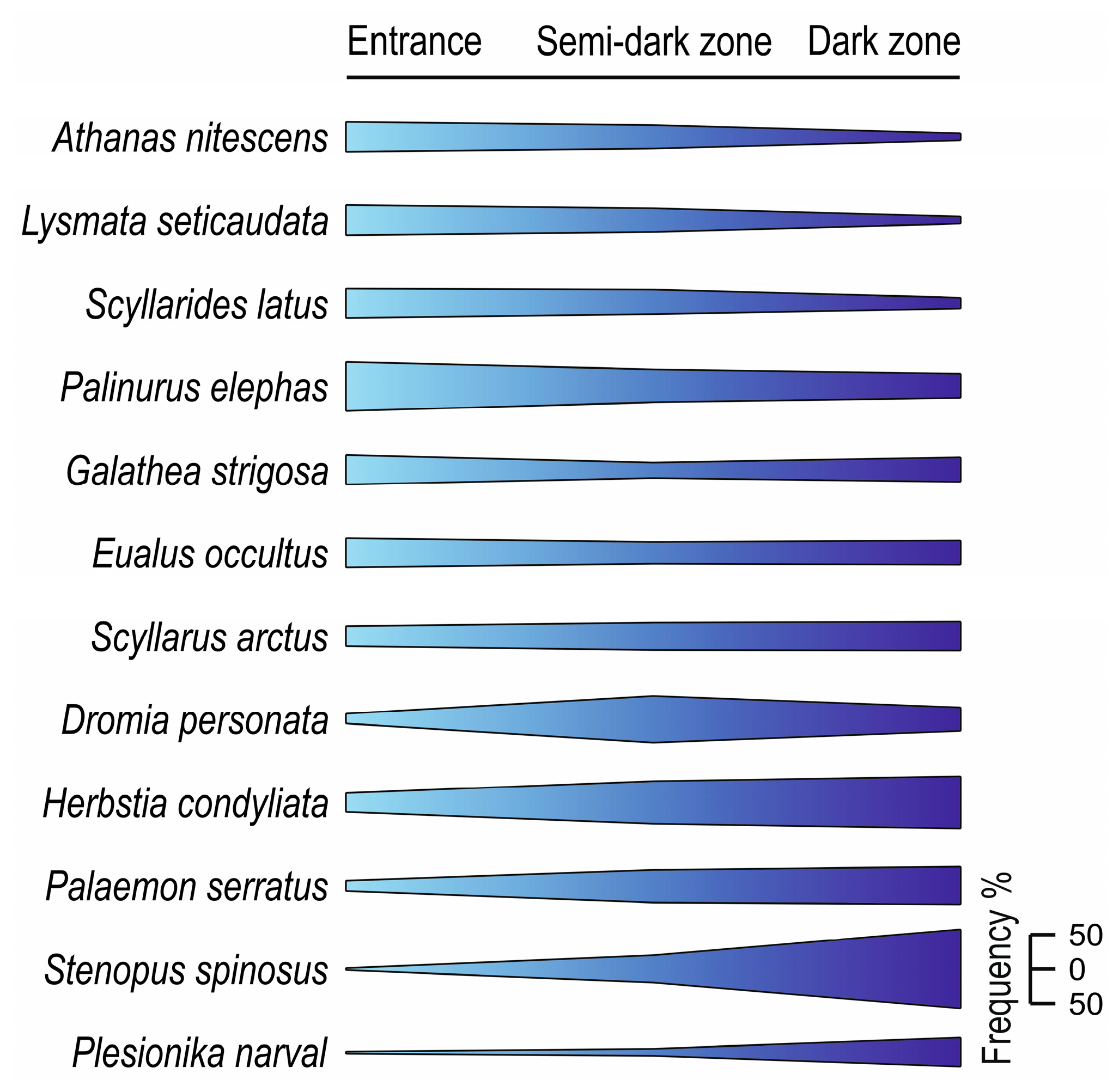
5. Depth and Cave Zone Preferences
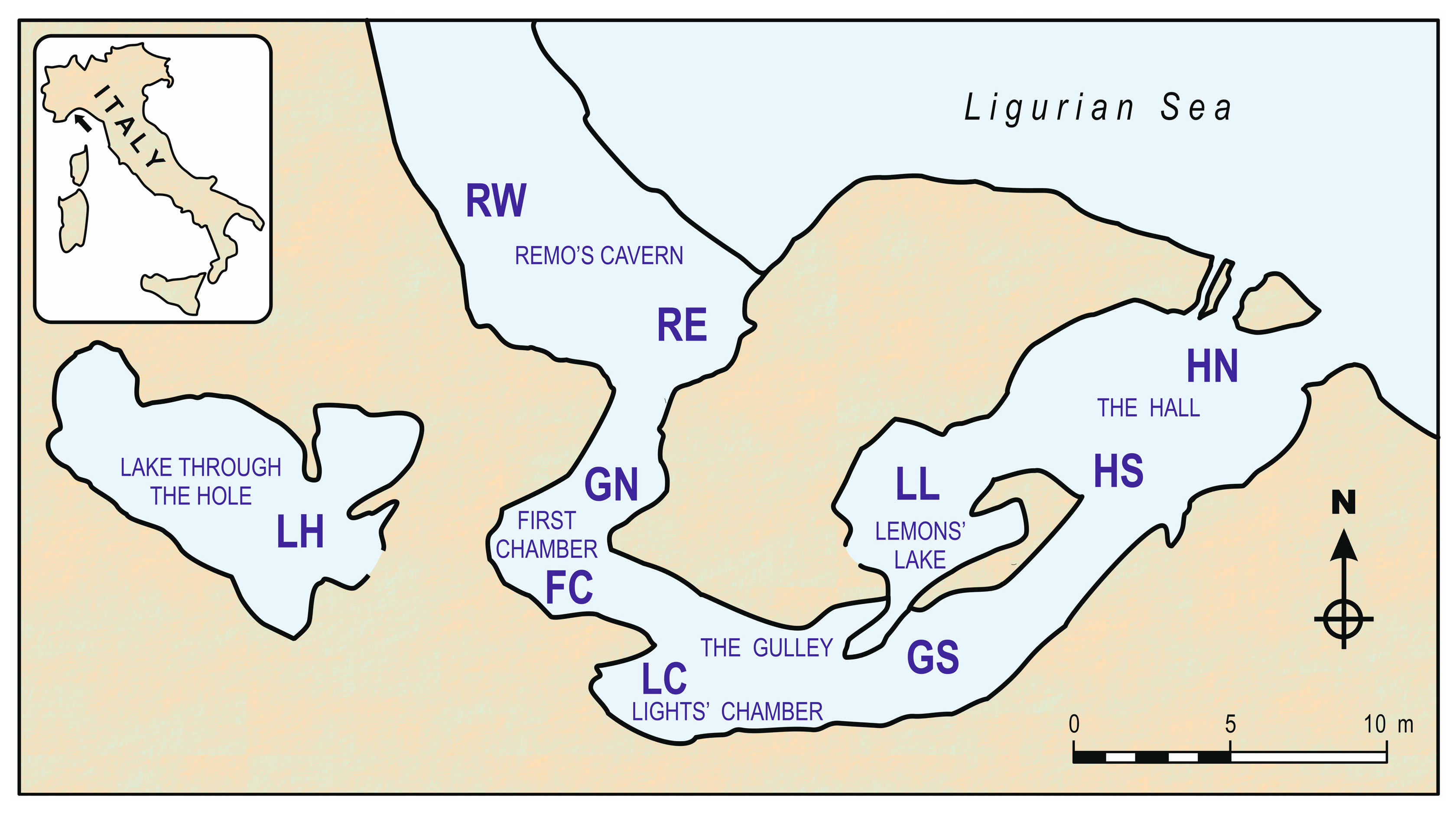
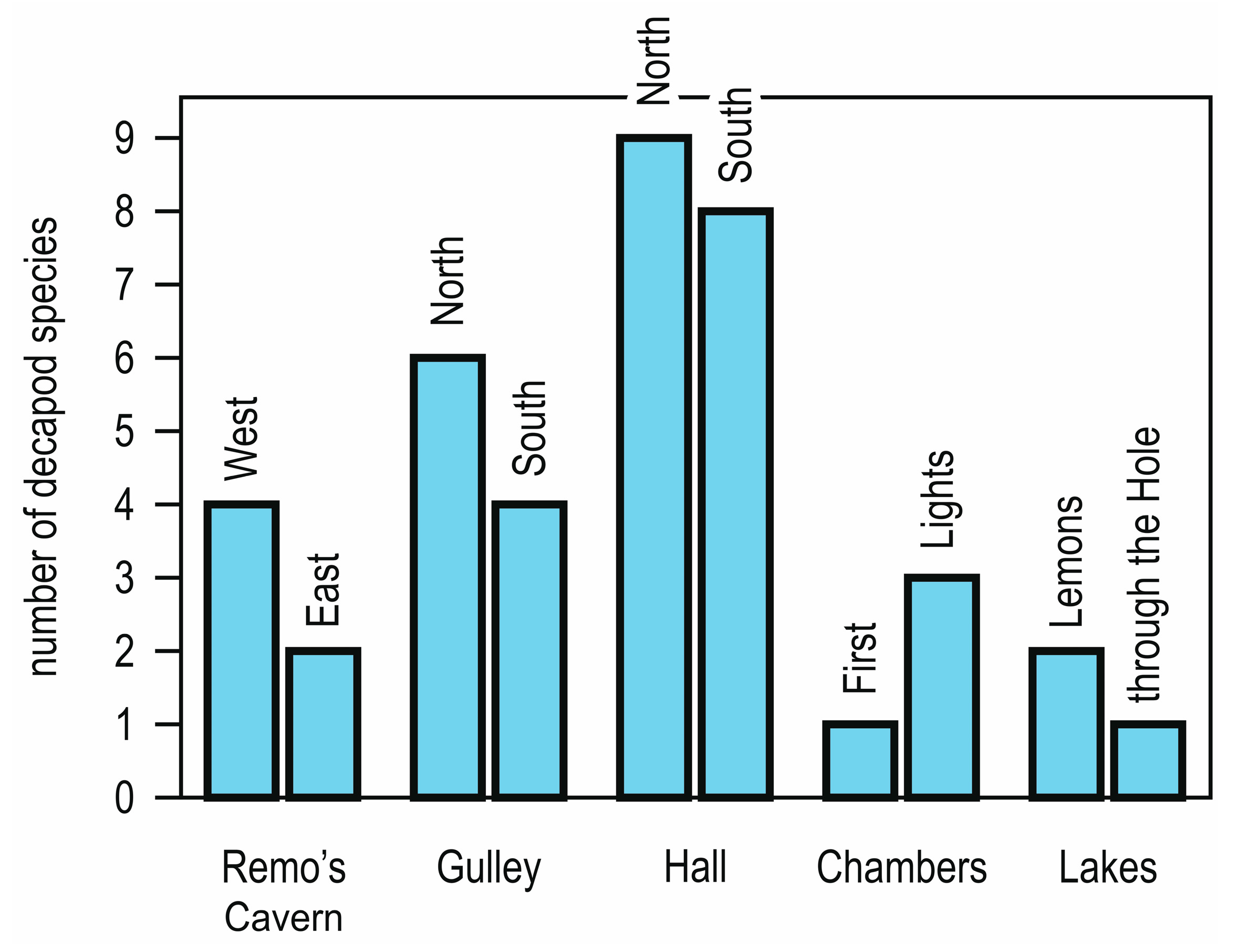
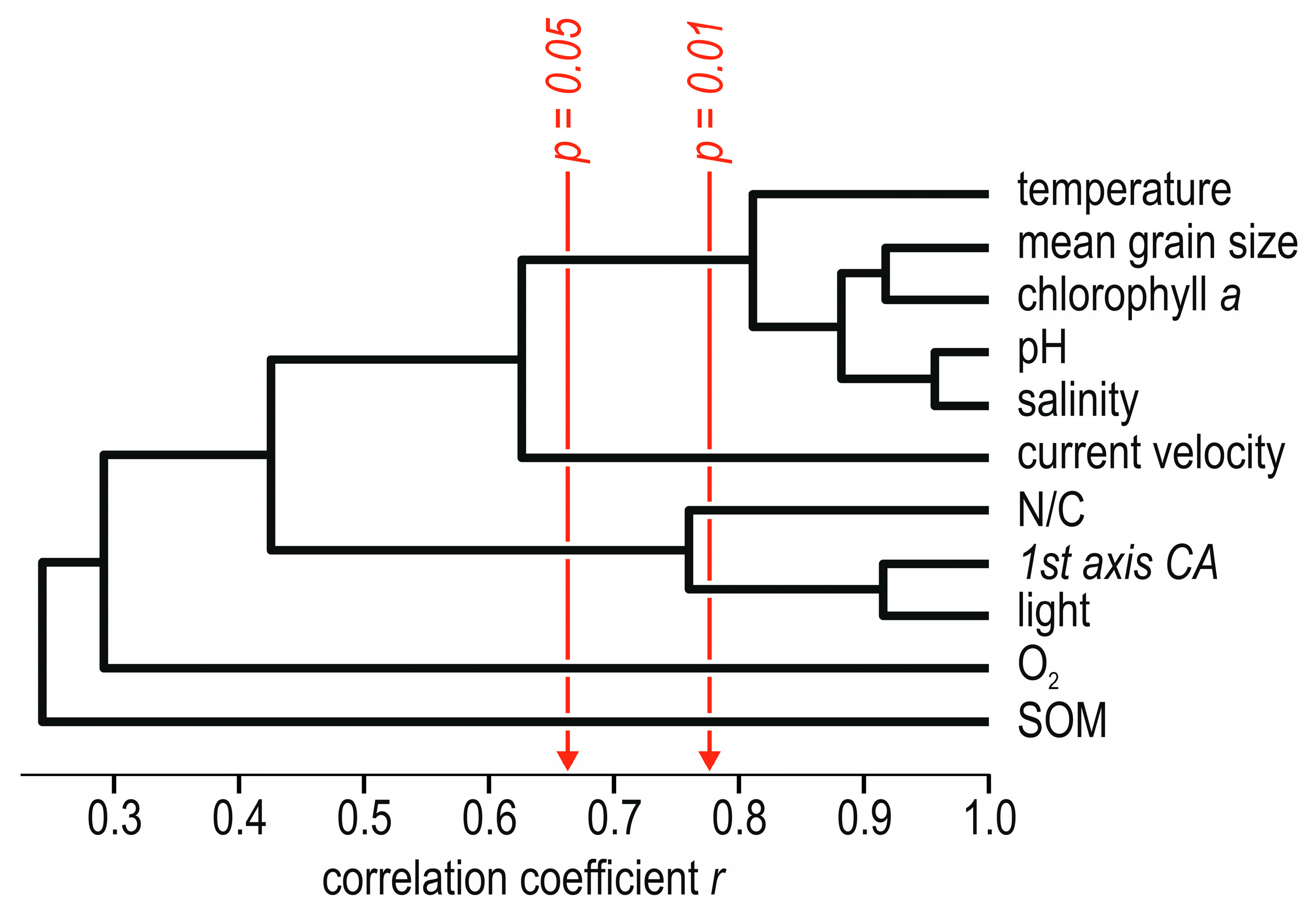
6. Ecology and Distribution in the Grotta Marina di Bergeggi
7. Final Remarks
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Anderson, S.C.; Mills Flemming, J.; Watson, R.; Lotze, H.K. Rapid global expansion of invertebrate fisheries: Trends, drivers, and ecosystem effects. PLoS ONE 2011, 6, e14735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Grave, S.; Pentcheff, N.D.; Ahyong, S.T.; Chan, T.Y.; Crandall, K.A.; Dworschak, P.C.; Felder, D.L.; Feldmann, R.M.; Fransen, C.H.J.M.; Goulding, L.Y.D.; et al. A classification of living and fossil genera of decapod crustaceans. Raffles Bull. Zool. Suppl. 2009, 21, 1–109. [Google Scholar]
- Abele, L.G. Species diversity of decapod crustaceans in marine habitats. Ecology 1974, 55, 156–161. [Google Scholar] [CrossRef]
- Noël, P.; Monod, T.; Laubier, L. Crustacea in the biosphere. In Treatise on Zoology—Anatomy, Taxonomy, Biology. The Crustacea; von Vaupel Klein, J.C., Charmantier-Daures, M., Schram, F.R., Eds.; E.J. Brill: Leiden, The Netherlands, 2014; Volume 4, part B, revised, updated edition; pp. 3–115. [Google Scholar]
- Martin, J.W.; Haney, T.A. Decapod crustaceans from hydrothermal vents and cold seeps: A review through 2005. Zool. J. Linn. Soc. 2005, 145, 445–522. [Google Scholar] [CrossRef]
- Harmelin, J.G.; Vacelet, J.; Vasseur, P. Les grottes sous-marines obscures: Un milieu extrême et un remarquable biotope refuge. Téthys 1985, 11, 214–229. [Google Scholar]
- Riedl, R. Biologie der Meereshöhlen; Paul Parey: Hamburg, Germany, 1966; pp. 1–636. [Google Scholar]
- Holthuis, L.B. Decapoda. In Stygofauna Mundi: A Faunistic, Distributional, and Ecological Synthesis of the World Fauna Inhabiting Subterranean Waters (Including the Marine Interstitial); Botosaneanu, L., Ed.; E.J. Brill: Leiden, The Netherlands, 1986; pp. 589–615. [Google Scholar]
- Anker, A. A worldwide review of stygobiotic and stygophilic shrimps of the family Alpheidae (Crustacea, Decapoda, Caridea). Subterr. Biol. 2008, 6, 1–16. [Google Scholar]
- Giakoumi, S.; Sini, M.; Gerovasileiou, V.; Mazor, T.; Beher, J.; Possingham, H.P.; Abdulla, A.; Çinar, M.E.; Dendrinos, P.; Gucu, A.C.; et al. Ecoregion-based conservation planning in the Mediterranean: Dealing with large-scale heterogeneity. PLoS ONE 2013, 8, e76449. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N.; Cattaneo-Vietti, R.; Cinelli, F.; Morri, C.; Pansini, M. Lo studio biologico delle grotte sottomarine: Conoscenze attuali e prospettive. Boll. Mus. Ist. Biol. Univ. Genova 1996, 60, 41–69. [Google Scholar]
- Gerovasileiou, V.; Chintiroglou, C.; Vafidis, D.; Koutsoubas, D.; Sini, M.; Dailianis, T.; Issaris, Y.; Akritopoulou, E.; Dimarchopoulou, D.; Voultsiadou, E. Census of biodiversity in marine caves of the Eastern Mediterranean Sea. Medit. Mar. Sci. 2015, 16, 245–265. [Google Scholar] [CrossRef] [Green Version]
- Gerovasileiou, V.; Bianchi, C.N. Mediterranean marine caves: A synthesis of current knowledge. Oceanogr. Mar. Biol. Ann. Rev. 2021, 59, 1–88. [Google Scholar]
- Gerovasileiou, V.; Voultsiadou, E. Mediterranean marine caves as biodiversity reservoirs: A preliminary overview. In Proceedings of the 1st Mediterranean Symposium on the Conservation of Dark Habitats; Langar, H., Bouafif, C., Ouerghi, A., Eds.; UNEP/MAP–RAC/SPA: Tunis, Tunisia, 2014; pp. 45–50. [Google Scholar]
- Gerovasileiou, V.; Martínez, A.; Álvarez, F.; Boxshall, G.; Humphreys, W.F.; Jaume, D.; Becking, L.E.; Muricy, G.; van Hengstum, P.J.; Dekeyzer, S.; et al. World Register of marine Cave Species (WoRCS): A new thematic species database for marine and anchialine cave biodiversity. Res. Ideas Outcomes 2016, 2, e10451. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N.; Cevasco, M.G.; Diviacco, G.; Morri, C. Primi risultati di una ricerca ecologica sulla grotta marina di Bergeggi (Savona). Boll. Mus. Ist. Biol. Univ. Genova 1986, 52, 267–293. [Google Scholar]
- Parravicini, V.; Guidetti, P.; Morri, C.; Montefalcone, M.; Donato, M.; Bianchi, C.N. Consequences of sea water temperature anomalies on a Mediterranean submarine cave ecosystem. Estuar. Coast. Shelf Sci. 2010, 86, 276–282. [Google Scholar] [CrossRef]
- Montefalcone, M.; De Falco, G.; Nepote, E.; Canessa, M.; Bertolino, M.; Bavestrello, G.; Morri, C.; Bianchi, C.N. Thirty year ecosystem trajectories in a submerged marine cave under changing pressure regime. Mar. Environ. Res. 2018, 137, 98–110. [Google Scholar] [CrossRef] [PubMed]
- Sgorbini, S.; Bianchi, C.N.; Degl’Innocenti, F.; Diviacco, G.; Forti, S.; Morri, C.; Niccolai, I. Méthodologie d’une étude hydrobiologique dans la grotte marine de Bergeggi (mer Ligure). Rapp. Comm. Int. Mer Médit. 1988, 31, 119. [Google Scholar]
- Morri, C.; Bianchi, C.N.; Degl’Innocenti, F.; Diviacco, G.; Forti, S.; Maccarone, M.; Niccolai, I.; Sgorbini, S.; Tucci, S. Gradienti fisico-chimici e ricoprimento biologico nella Grotta Marina di Bergeggi (Mar Ligure). Mem. Ist. Ital. Speleol. Ser. II 1994, 6, 85–94. [Google Scholar]
- Stock, J.H.; Iliffe, T.M.; Williams, D. The concept ‘anchialine’ reconsidered. Stygologia 1986, 2, 90–92. [Google Scholar]
- Bianchi, C.N. Flora e fauna: Lineamenti generali e prospettive. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, C.N., Ferrari, G., Forti, P., Eds.; Ministero dell’Ambiente e della Tutela del Territorio: Roma, Italy, 2003; pp. 137–146. [Google Scholar]
- Gràcia, F.; Jaume, D.; Ramis, D.; Fornós, J.J.; Bover, P.; Clamor, B.; Gual, M.À.; Vadell, M. Les coves de Cala Anguila (Manacor, Mallorca). II: La Cova Genovesa o Cova d’en Bessó. Espeleogènesi, geomorfologia, hidrologia, sedimentologia, fauna, paleontologia, arqueologia i concervació. Endins Pub. Espeleol. 2003, 25, 43–86. [Google Scholar]
- Parenzan, P. Animalia Speluncarum Italiae; Congedo: Galatina, Italy, 1989; (published posthumous in 2002); pp. 1–224. [Google Scholar]
- Gili, J.M.; Macpherson, E. Crustáceos decápodos capturados en cuevas submarinas del litoral Balear. Investig. Pesq. 1987, 51 (Suppl. 1), 285–291. [Google Scholar]
- Pessani, D.; Manconi, R. I Crostacei Decapodi delle grotte della Penisola Sorrentina. Mem. Ist. Ital. Speleol. Ser. II 1994, 6, 199–202. [Google Scholar]
- Pipitone, C.; Vaccaro, A. Studio dei crostacei decapodi dell’Isola di Ustica: Censimento faunistico, distribuzione e biogeografia. NTR-IRMA 2003, 70, 1–24. [Google Scholar]
- Manconi, R.; Pessani, D. Crostacei decapodi. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, C.N., Ferrari, G., Forti, P., Eds.; Ministero dell’Ambiente e della Tutela del Territorio: Roma, Italy, 2003; pp. 187–193. [Google Scholar]
- Denitto, F.; Moscatello, S.; Belmonte, G. Occurrence and distribution pattern of Palaemon spp. shrimps in a shallow submarine cave environment: A study case in South-eastern Italy. Mar. Ecol. 2009, 30, 416–424. [Google Scholar] [CrossRef]
- Denitto, F.; Pastore, M.; Belmonte, G. Occurrence of the Guinean species Herbstia nitida Manning & Holthuis, 1981 (Decapoda, Brachyura) in a Mediterranean submarine cave and a comparison with the congeneric H. condyliata (Fabricius, 1787). Crustaceana 2010, 83, 1017–1024. [Google Scholar]
- Mačić, V.; Panou, A.; Bundone, L.; Varda, D.; Pavićević, M. First inventory of the semi-submerged marine caves in South Dinarides Karst (Adriatic coast) and preliminary list of species. Turk. J. Fish. Aquat. Sci. 2019, 19, 765–774. [Google Scholar] [CrossRef]
- Pretus, J.L. Description of Odontozona addaia spec. nov. (Crustacea: Decapoda: Stenopodidae) from a marine cave in the island of Minorca, western Mediterranean. Zool. Meded. 1990, 63, 343–357. [Google Scholar]
- Fransen, C.H.J.M. Salmoneus sketi, a new species of alpheid shrimp (Crustacea: Decapoda: Caridea) from a submarine cave in the Adriatic. Zool. Meded. 1991, 65, 171–179. [Google Scholar]
- Chevaldonné, P.; Pretus, J.L. Rediscovery of the rare Mediterranean marine cave stenopodid shrimp Odontozona addaia Pretus, 1990, 30 years after its original description (Crustacea: Decapoda: Stenopodidea). Zootaxa 2021, 4950, 137–148. [Google Scholar] [CrossRef]
- Rastorgueff, P.A.; Chevaldonné, P.; Arslan, D.; Verna, C.; Lejeusne, C. Cryptic habitats and cryptic diversity: Unexpected patterns of connectivity and phylogeographical breaks in a Mediterranean endemic marine cave mysid. Mol. Ecol. 2014, 23, 2825–2843. [Google Scholar] [CrossRef] [Green Version]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. In Life in the Mediterranean Sea: A Look at Habitat Changes; Stambler, N., Ed.; Nova Science: New York, NY, USA, 2012; pp. 1–55. [Google Scholar]
- Dengler, J.; Oldeland, J. Effects of sampling protocol on the shapes of species richness curves. J. Biogeogr. 2010, 37, 1698–1705. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Azzola, A.; Cocito, S.; Morri, C.; Oprandi, A.; Peirano, A.; Sgorbini, S.; Montefalcone, M. Biodiversity monitoring in Mediterranean marine protected areas: Scientific and methodological challenges. Diversity 2022, 14, 43. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Dando, P.R.; Morri, C. Increased biodiversity of sessile epibenthos at subtidal hydrothermal vents: Seven hypotheses based on observations at Milos Island, Aegean Sea. Adv. Oceanogr. Limnol. 2011, 2, 1–31. [Google Scholar] [CrossRef]
- Williamson, M. Relationship of species number to area, distance and other variables. In Analytical Biogeography. An Integrated Approach to the Study of Animal and Plant Distributions; Myers, A.A., Giller, P.S., Eds.; Chapman and Hall: London, UK, 1990; pp. 92–115. [Google Scholar]
- Bianchi, C.N.; Morri, C. Marine biodiversity of the Mediterranean Sea: Situation, problems and prospects for future research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Tortonese, E. How is to be interpreted a ‘Mediterranean’ species? Thalassographica 1978, 2, 9–17. [Google Scholar]
- Tortonese, E. Distribution and ecology of endemic elements in the Mediterranean fauna (fishes and echinoderms). In Mediterranean Marine Ecosystems; Moraitou-Apostolopoulou, M., Kiortsis, V., Eds.; NATO Conference Series 8; Plenum Press: New York, NY, USA, 1985; pp. 57–83. [Google Scholar]
- Pérès, J.M.; Picard, J. Nouveau manuel de bionomie benthique de la mer Méditerranée. Rec. Trav. Stat. Mar. Endoume 1964, 31, 1–137. [Google Scholar]
- Fredj, G.; Bellan-Santini, D.; Menardi, M. État des connaissances sur la faune marine méditerranéenne. Bull. Inst. Océanogr. Monaco 1992, no. spécial 9, 133–145. [Google Scholar]
- Harmelin, J.G.; d’Hont, J.L. Transfers of bryozoan species between the Atlantic Ocean and the Mediterranean Sea via the Strait of Gibraltar. Oceanol. Acta 1993, 16, 63–72. [Google Scholar]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Çinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 1. Spatial distribution. Medit. Mar. Sci. 2010, 11, 381–493. [Google Scholar] [CrossRef] [Green Version]
- Galil, B.S. Lessepsian migration: New findings on the foremost anthropogenic change in the Levant basin fauna. In Symposium Mediterranean Seas 2000; Della Croce, N.F.R., Ed.; University of Genova, Istituto di Scienze Ambientali Marine: Santa Margherita Ligure, Italy, 1993; pp. 307–323. [Google Scholar]
- Logan, A.; Bianchi, C.N.; Morri, C.; Zibrowius, H. The present-day Mediterranean brachiopod fauna: Diversity, life habits, biogeography and paleobiogeography. Sci. Mar. 2004, 68 (Suppl. 1), 163–170. [Google Scholar] [CrossRef] [Green Version]
- D’Udekem d’Acoz, C. Inventaire et Distribution des Crustacés Décapodes de l’Atlantique Nord-Oriental, de la Méditerranée et des Eaux Continentales Adjacentes au Nord de 25° N; Patrimoines Naturels 40; Muséum National d’Histoire Naturelle: Paris, France, 1999; pp. 1–383. [Google Scholar]
- Bakir, K.; Katagan, T.; Aker, H.V.; Ozcan, T.; Sezgin, M.; Ates, A.S.; Koçak, C.; Kirmim, F. The marine arthropods of Turkey. Turk. J. Zool. 2014, 38, 765–831. [Google Scholar] [CrossRef] [Green Version]
- Marco-Herrero, E.; Abello, P.; Drake, P.; García-Raso, J.E.; Gonzalez-Gordillo, J.I.; Guerao, G.; Palero, F.; Cuesta, J.A. Annotated checklist of brachyuran crabs (Crustacea: Decapoda) of the Iberian Peninsula (SW Europe). Sci. Mar. 2015, 79, 243–256. [Google Scholar] [CrossRef] [Green Version]
- García Raso, J.E.; Cuesta, J.A.; Abello, P.; Macpherson, E. Updating changes in the Iberian decapod crustacean fauna (excluding crabs) after 50 years. Sci. Mar. 2018, 82, 207–229. [Google Scholar] [CrossRef]
- Gonzalez, J.A. Checklists of Crustacea Decapoda from the Canary and Cape Verde Islands, with an assessment of Macaronesian and Cape Verde biogeographic marine ecoregions. Zootaxa 2018, 4412, 401–448. [Google Scholar] [CrossRef] [PubMed]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terossi, M.; De Grave, S.; Mantelatto, F.L. Global biogeography, cryptic species and systematic issues in the shrimp genus Hippolyte Leach, 1814 (Decapoda: Caridea: Hippolytidae) by multimarker analyses. Sci. Rep. 2017, 7, e6697. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ledoyer, M. Écologie de la faune vagile des biotopes méditerranéens accessibles en scaphandre autonome (région de Marseille principalement). IV.—Synthèse de l’étude écologique. Rec. Trav. Stat. Mar. Endoume 1968, 44, 125–295. [Google Scholar]
- García, L.; Gràcia, F. Sobre algunes espècies de crustacis decàpodes interessants de les illes Balears (Crustacea: Decapoda). Boll. Soc. Hist. Nat. Balears 1996, 39, 177–186. [Google Scholar]
- Tortonese, E. La fauna del Mediterraneo e i suoi rapporti con quella dei mari vicini. Pubbl. Stn. Zool. Napoli 1969, 37, 369–384. [Google Scholar]
- Briggs, J.C. Marine Zoogeography; McGraw-Hill: New York, NY, USA, 1974; pp. 1–475. [Google Scholar]
- Ferreira, L.A.A.; Tavares, M. Pisidia longimana (Risso, 1816), a junior synonym of P. bluteli (Risso, 1816) (Crustacea: Decapoda: Anomura: Porcellanidae) and a species distinct from P. longicornis (Linnaeus, 1767). Pap. Avul. Zool. 2020, 60, e20206036. [Google Scholar] [CrossRef]
- Spalding, M.D.; Fox, H.E.; Allen, G.R.; Davidson, N.; Ferdaña, Z.A.; Finlayson, M.A.X.; Halpern, B.S.; Jorge, M.A.; Lombana, A.; Lourie, S.A.; et al. Marine ecoregions of the world: A bioregionalization of coastal and shelf areas. BioScience 2007, 57, 573–583. [Google Scholar] [CrossRef] [Green Version]
- Briggs, J.C.; Bowen, B.W. A realignment of marine biogeographic provinces with particular reference to fish distributions. J. Biogeogr. 2012, 39, 12–30. [Google Scholar] [CrossRef]
- Briggs, J.C.; Bowen, B.W. Marine shelf habitat: Biogeography and evolution. J. Biogeogr. 2013, 40, 1023–1035. [Google Scholar] [CrossRef]
- Morri, C.; Cattaneo-Vietti, R.; Sartoni, G.; Bianchi, C.N. Shallow epibenthic communities of Ilha do Sal (Cape Verde Archipelago, Eastern Atlantic). Arquipél. Life Mar. Sci. 2000, 2, 157–165. [Google Scholar]
- Sparrow, A.; Badalamenti, F.; Pipitone, C. Contribution to the knowledge of Percnon gibbesi (Decapoda, Grapsidae), an exotic species spreading rapidly in Sicilian waters. Crustaceana 2001, 74, 1009–1017. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; García Raso, J.E.; Çinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Medit. Mar. Sci. 2010, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Yokes, B.; Galil, B.S. New records of alien decapods (Crustacea) from the Mediterranean coast of Turkey, with a description of a new palaemonid species. Zoosystema 2006, 28, 747–755. [Google Scholar]
- Ďuriš, Z. Palaemonid shrimps (Crustacea: Decapoda) of Saudi Arabia from the ‘Red Sea biodiversity survey’ 2011–2013, with 11 new records for the Red Sea. Mar. Biodiver. 2017, 47, 1147–1161. [Google Scholar] [CrossRef]
- Gerovasileiou, V.; Voultsiadou, E.; Issaris, Y.; Zenetos, A. Alien biodiversity in Mediterranean marine caves. Mar. Ecol. 2016, 37, 239–256. [Google Scholar] [CrossRef]
- Klaoudatos, D.; Kapiris, K. Alien crabs in the Mediterranean Sea: Current status and perspectives. In Crabs: Global Diversity, Behavior and Environmental Threats; Ardovini, C., Ed.; Nova Science: New York, NY, USA, 2014; pp. 101–159. [Google Scholar]
- Morri, C.; Puce, S.; Bianchi, C.N.; Bitar, G.; Zibrowius, H.; Bavestrello, G. Hydroids (Cnidaria: Hydrozoa) from the Levant Sea (mainly Lebanon), with emphasis on alien species. J. Mar. Biol. Ass. UK 2009, 89, 49–62. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Caroli, F.; Guidetti, P.; Morri, C. Seawater warming at the northern reach for southern species: Gulf of Genoa, NW Mediterranean. J. Mar. Biol. Ass. UK 2018, 98, 1–12. [Google Scholar] [CrossRef]
- Morais, S.; Narciso, L.; Calado, R.; Nunes, M.L.; Rosa, R. Lipid dynamics during the embryonic development of Plesionika martia martia (Decapoda; Pandalidae), Palaemon serratus and P. elegans (Decapoda; Palaemonidae): Relation to metabolic consumption. Mar. Ecol. Progr. Ser. 2002, 242, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Thessalou-Legaki, M.; Frantzis, A.; Nassiokas, K.; Hatzinikolaou, S. Depth zonation in a Parapandalus narval (Crustacea, Decapoda, Pandalidae) population from Rhodos Island, Greece. Estuar. Coast. Shelf Sci. 1989, 29, 273–284. [Google Scholar] [CrossRef]
- Zariquiey Alvarez, R. Crustáceos Decápodos Ibéricos. Investig. Pesq. 1968, 32, 1–510. [Google Scholar]
- Bianchi, C.N. Biologia delle grotte marine del Mediterraneo. In Mare ed Ecologia; Fiorentini, A., Ed.; Provincia di Genova, Unione Regionale Province Liguri and Marevivo: Genoa, Italy, 1994; pp. 35–44. [Google Scholar]
- Bianchi, C.N.; Morri, C. Biologia ed ecologia delle grotte sottomarine. In Speleologia Marina; Barbieri, F., Ed.; Gribaudo: Cavallermaggiore, Italy, 1999; pp. 113–160. [Google Scholar]
- Bianchi, C.N.; Morri, C. Studio bionomico comparativo di alcune grotte marine sommerse; definizione di una scala di confinamento. Mem. Ist. Ital. Speleol. 1994, 6 (Suppl. 2), 107–123. [Google Scholar]
- Harmelin, J.G. Les peuplements des substrats durs circalittoraux. In Les Biocénoses Marines et Littorales de Méditérranée. Synthèse, Menaces et Perspectives; Bellan-Santini, D., Lacaze, J.C., Poizat, C., Eds.; Patrimoines Naturels 19; Muséum National d’Histoire Naturelle: Paris, France, 1994; pp. 118–126. [Google Scholar]
- Cinelli, F.; Fresi, E.; Mazzella, L.; Pansini, M.; Pronzato, R.; Svoboda, A. Distribution of benthic phyto- and zoocoenoses along a light gradient in a superficial marine cave. In Biology of Benthic Organisms; Keegan, B.F., Ceidigh, P.O., Boaden, P.J.S., Eds.; Pergamon Press: Oxford, UK, 1977; pp. 173–183. [Google Scholar]
- Southward, A.J.; Kennicutt, M.C.; Alcalà-Herrera, J.; Abbiati, M.; Airoldi, L.; Cinelli, F.; Bianchi, C.N.; Morri, C.; Southward, E.C. On the biology of submarine caves with sulphur springs: Appraisal of 13C/12C ratios as a guide to trophic relations. J. Mar. Biol. Ass. UK 1996, 76, 265–285. [Google Scholar] [CrossRef]
- Harmelin, J.G. Bryozoaires des grottes sous-marines obscures de la région marseillaise, faunistique et écologie. Téthys 1969, 1, 793–806. [Google Scholar]
- Morri, C. Confinamento idrologico. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, C.N., Ferrari, G., Forti, P., Eds.; Ministero dell’Ambiente e della Tutela del Territorio: Rome, Italy, 2003; pp. 291–296. [Google Scholar]
- Bianchi, C.N.; Abbiati, M.; Airoldi, L.; Alvisi, M.; Benedetti-Cecchi, L.; Cappelletti, A.; Cinelli, F.; Colantoni, P.; Dando, P.R.; Morri, C.; et al. Hydrology and water budget of a submarine cave with sulphur water spring: The Grotta Azzurra of Capo Palinuro (Southern Italy). Proc. Ital. Ass. Oceanol. Limnol. 1998, 12, 285–301. [Google Scholar]
- Bianchi, C.N.; Morri, C.; Russo, G.F. Deplezione trofica. In Grotte Marine: Cinquant’anni di Ricerca in Italia; Cicogna, F., Bianchi, C.N., Ferrari, G., Forti, P., Eds.; Ministero dell’Ambiente e della Tutela del Territorio: Rome, Italy, 2003; pp. 297–305. [Google Scholar]
- Rastorgueff, P.A.; Bellan-Santini, D.; Bianchi, C.N.; Bussotti, S.; Chevaldonné, P.; Guidetti, P.; Harmelin, J.G.; Montefalcone, M.; Morri, C.; Perez, T.; et al. An ecosystem-based approach to evaluate the ecological quality of Mediterranean undersea caves. Ecol. Indic. 2015, 54, 137–152. [Google Scholar] [CrossRef]
- Fichez, R. Les pigments chlorophylliens: Indices d’oligotrophie dans les grottes sous-marines. Compte Rendus Acad. Sci. 1990, 310, 155–161. [Google Scholar]
- Cocito, S.; Fanucci, S.; Niccolai, I.; Morri, C.; Bianchi, C.N. Relationships between trophic organization of benthic communities and organic matter content in Tyrrhenian Sea sediments. Hydrobiologia 1990, 207, 53–60. [Google Scholar] [CrossRef]
- Rastorgueff, P.-A.; Harmelin-Vivien, M.; Richard, P.; Chevaldonné, P. Feeding strategies and resource partitioning mitigate the effects of oligotrophy for marine cave mysids. Mar. Ecol. Progr. Ser. 2011, 440, 163–176. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Pronzato, R.; Cattaneo-Vietti, R.; Benedetti-Cecchi, L.; Morri, C.; Pansini, M.; Chemello, R.; Milazzo, M.; Fraschetti, S.; Terlizzi, A.; et al. Mediterranean marine benthos: A manual of methods for its sampling and study. Hard bottoms. Biol. Mar. Medit. 2004, 11, 185–215. [Google Scholar]
- Hirst, C.N.; Jackson, D.A. Reconstructing community relationships: The impact of sampling error, ordination approach, and gradient length. Divers. Distrib. 2007, 13, 361–371. [Google Scholar] [CrossRef]
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 2012; pp. 1–1006. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PaSt: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Lebart, L. Validité des résultats en analyse des données. Consommation 1977, 1, 41–69. [Google Scholar]
- Öztürk, B.; Güngör, A.; Barraud, T. Marine caves biodiversity and conservation in the Turkish part of the Mediterranean Sea: Preliminary results of East Med Cave Project. In Marine Caves of the Eastern Mediterranean Sea: Biodiversity, Threats and Conservation; Öztürk, B., Ed.; Publication no. 53; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2020; pp. 147–158. [Google Scholar]
- Crocetta, F.; Al Mabruk, S.A.A.; Azzurro, E.; Bakiu, R.; Bariche, M.; Batjakas, I.E.; Bejaoui, T.; Ben Souissi, J.; Cauchi, J.; Corsini-Foka, M.; et al. New alien Mediterranean biodiversity records (November 2021). Medit. Mar. Sci. 2021, 22, 724–746. [Google Scholar]
- Digenis, M.; Ragkousis, M.; Vasileiadou, K.; Gerovasileiou, V.; Katsanevakis, S. New records of the Indo-Pacific shrimp Urocaridella pulchella Yokes and Galil, 2006 from the Eastern Mediterranean Sea. BioInvasions Rec. 2021, 10, 295–303. [Google Scholar] [CrossRef]
- Moscatello, S.; Belmonte, G. The plankton of a shallow submarine cave (‘Grotta di Ciolo’, Salento Peninsula, SE Italy). Mar. Ecol. 2007, 28 (Suppl. 1), 47–59. [Google Scholar] [CrossRef]
- Cattaneo, R.; Pastorino, M.V. Popolamenti algali e fauna bentonica nelle cavità naturali della regione litorale mediterranea. Rass. Speleol. Ital. 1974, 12, 272–281. [Google Scholar]
- Karlovac, O. Présence du Stenopus spinosus Risso dans l’Adriatique. Bil. Inst. Oceanogr. Ribar. Split 1953, 5, 1–3. [Google Scholar]
- Ferrero, L. Parapandalus narval (Fabricius) in una grotta dell’Isola di Giannutri—Arcipelago Toscano. Boll. Pesca Piscic. Idrobiol. N.S. 1969, 23, 163–167. [Google Scholar]
- Iliffe, T.M.; Bishop, R.E. Adaptations to life in marine caves. In Fisheries and Aquaculture. Encyclopedia of Life Support Systems; Safran, P., Ed.; UNESCO and EOLSS Publishers: Oxford, UK, 2007; pp. 1–26. [Google Scholar]
- Mammola, S.; Amorim, I.R.; Bichuette, M.E.; Borges, P.A.V.; Cheeptham, N.; Cooper, S.J.B.; Culver, D.C.; Deharveng, L.; Eme, D.; Ferreira, R.L.; et al. Fundamental research questions in subterranean biology. Biol. Rev. 2020, 95, 1855–1872. [Google Scholar] [CrossRef]
- Ott, J.A.; Svoboda, A. Sea caves as model systems for energy flow studies in primary hard bottom communities. Pubbl. Stn. Zool. Napoli 1978, 40, 477–485. [Google Scholar]
- Coma, R.; Carola, M.; Riera, T.; Zabala, M. Horizontal transfer of matter by a cave-dwelling mysid. Mar. Ecol. 1997, 18, 211–226. [Google Scholar] [CrossRef]
- Bussotti, S.; Di Franco, A.; Bianchi, C.N.; Chevaldonné, P.; Egea, L.; Fanelli, Ε.; Lejeusne, C.; Musco, L.; Navarro-Barranco, C.; Pey, A.; et al. Fish mitigate trophic depletion in marine cave ecosystems. Sci. Rep. 2018, 8, 9193. [Google Scholar] [CrossRef]
- Nepote, E.; Bianchi, C.N.; Morri, C.; Ferrari, M.; Montefalcone, M. Impact of a harbour construction on the benthic community of two shallow marine caves. Mar. Pollut. Bull. 2017, 114, 35–45. [Google Scholar] [CrossRef]
- Ouerghi, A.; Gerovasileiou, V.; Bianchi, C.N. Mediterranean marine caves: A synthesis of current knowledge and the Mediterranean Action Plan for the conservation of “dark habitats”. In Marine Caves of the Eastern Mediterranean Sea: Biodiversity, Threats and Conservation; Öztürk, B., Ed.; Publication no. 53; Turkish Marine Research Foundation (TUDAV): Istanbul, Turkey, 2019; pp. 1–13. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Infraorder | Family | Genus and Species |
|---|---|---|
| Stenopodidea | Stenopodidae | Odontozona addaia Pretus, 1990 |
| Stenopus spinosus Risso, 1827 | ||
| Caridea | Palaemonidae | Balssia gasti (Balss, 1921) |
| Brachycarpus biunguiculatus (Lucas, 1846) | ||
| Gnathophyllum elegans (Risso, 1816) | ||
| Palaemon adspersus Rathke, 1837 | ||
| Palaemon elegans Rathke, 1837 | ||
| Palaemon serratus (Pennant, 1777) | ||
| Palaemon xiphias Risso, 1816 | ||
| Periclimenes amethysteus (Risso, 1827) | ||
| Periclimenes scriptus (Risso, 1822) | ||
| Urocaridella pulchella Yokes & Galil, 2006 | ||
| Alpheidae | Alpheus dentipes Guérin, 1832 | |
| Athanas nitescens (Leach, 1814) | ||
| Salmoneus sketi Fransen, 1991 | ||
| Synalpheus gambarelloides (Nardo, 1847) | ||
| Hippolytidae | Caridion sp. | |
| Hippolyte holthuisi Zariquiey Alvarez, 1953 | ||
| Saron marmoratus (Olivier, 1811) | ||
| Lysmatidae | Lysmata nilita Dohrn & Holthuis, 1950 | |
| Lysmata seticaudata (Risso, 1816) | ||
| Pandalidae | Plesionika narval (Fabricius, 1787) | |
| Thoridae | Eualus cranchii (Leach, 1817) | |
| Eualus occultus (Lebour, 1936) | ||
| Eualus sollaudi (Zariquiey Cenarro, 1936) | ||
| Astacidea | Nephropidae | Homarus gammarus (Linnaeus, 1758) |
| Achelata | Palinuridae | Palinurus elephas (Fabricius, 1787) |
| Scyllaridae | Scyllarides latus (Latreille, 1803) | |
| Scyllarus arctus (Linnaeus, 1758) | ||
| Scyllarus pygmaeus (Bate, 1888) | ||
| Anomura | Galatheidae | Galathea dispersa Bate, 1859 |
| Galathea intermedia Lilljeborg, 1851 | ||
| Galathea nexa Embleton, 1836 | ||
| Galathea strigosa (Linnaeus, 1761) | ||
| Munididae | Munida rugosa (Fabricius, 1775) | |
| Porcellanidae | Pisidia bluteli (Risso, 1816) | |
| Porcellana platycheles (Pennant, 1777) | ||
| Diogenidae | Calcinus tubularis (Linnaeus, 1767) | |
| Clibanarius erythropus (Latreille, 1818) | ||
| Dardanus arrosor (Herbst, 1796) | ||
| Dardanus calidus (Risso, 1827) | ||
| Diogenes pugilator (P. Roux, 1829) | ||
| Paguridae | Cestopagurus timidus (P. Roux, 1830) | |
| Pagurus anachoretus Risso, 1827 | ||
| Pagurus chevreuxi (Bouvier, 1896) | ||
| Pagurus cuanensis Bell, 1845 | ||
| Pagurus prideaux Leach, 1815 | ||
| Brachyura | Dromiidae | Dromia personata (Linnaeus, 1758) |
| Homolidae | Homola barbata (Fabricius, 1793) | |
| Ethusidae | Ethusa mascarone (Herbst, 1785) | |
| Eriphiidae | Eriphia verrucosa (Forskål, 1775) | |
| Leucosiidae | Ilia nucleus (Linnaeus, 1758) | |
| Progeryonidae | Paragalene longicrura (Nardo, 1869) | |
| Epialtidae | Acanthonyx lunulatus (Risso, 1816) | |
| Herbstia condyliata (Fabricius, 1787) | ||
| Herbstia nitida Manning & Holthuis, 1981 | ||
| Lissa chiragra (Fabricius, 1775) | ||
| Pisa armata (Latreille, 1803) | ||
| Pisa nodipes Leach, 1815 | ||
| Inachidae | Achaeus cranchii Leach, 1817 | |
| Inachus dorsettensis (Pennant, 1777) | ||
| Macropodia czernjawskii (A.T. Brandt, 1880) | ||
| Macropodia rostrata (Linnaeus, 1761) | ||
| Majidae | Eurynome aspera (Pennant, 1777) | |
| Maja crispata Risso, 1827 | ||
| Maja squinado (Herbst, 1788) | ||
| Pilumnidae | Pilumnus hirtellus (Linnaeus, 1761) | |
| Pilumnus minutus De Haan, 1835 | ||
| Pilumnus spinifer H. Milne Edwards, 1834 | ||
| Portunidae | Carupa tenuipes Dana, 1852 | |
| Charybdis hellerii (A. Milne-Edwards, 1867) | ||
| Portunus hastatus (Linnaeus, 1767) | ||
| Xanthidae | Xantho pilipes A. Milne-Edwards, 1867 | |
| Grapsidae | Pachygrapsus marmoratus (Fabricius, 1787) | |
| Percnidae | Percnon gibbesi (H. Milne Edwards, 1853) | |
| Pinnotheridae | Nepinnotheres pinnotheres (Linnaeus, 1758) |
| Sectors | N | SN | c | z | rdir | S∞ | rrec |
|---|---|---|---|---|---|---|---|
| Alboran Sea | 3 | 5 | 1.7 | 1.00 | 1.000 | 32 | 0.993 |
| Balearic and Sardinia seas | 10 | 18 | 5.1 | 0.58 | 0.989 | 27 | 0.999 |
| Gulf of Lion | 7 | 29 | 7.5 | 0.74 | 0.996 | 45 | 0.999 |
| Ligurian Sea | 13 | 24 | 3.8 | 0.74 | 0.996 | 48 | 1.000 |
| North Tyrrhenian Sea | 16 | 19 | 3.1 | 0.69 | 0.991 | 32 | 0.999 |
| South Tyrrhenian Sea | 12 | 20 | 3.3 | 0.74 | 0.998 | 37 | 0.999 |
| Adriatic Sea | 20 | 19 | 2.1 | 0.75 | 0.996 | 32 | 0.999 |
| Ionian Sea | 22 | 22 | 2.3 | 0.64 | 0.997 | 36 | 0.999 |
| Aegean Sea | 18 | 17 | 3.1 | 0.60 | 0.990 | 23 | 0.999 |
| Levantine Basin | 9 | 15 | 2.8 | 0.78 | 0.997 | 37 | 1.000 |
| Parameters | RW | RE | GN | GS | FC | LC | HN | HS | LL | LH |
|---|---|---|---|---|---|---|---|---|---|---|
| Light (μW·cm−2) | 29.0 | 32.5 | 0.4 | 0.1 | 0.2 | 0.0 | 1.3 | 0.2 | 0.1 | 0.0 |
| Current Velocity (cm·s−1) | 20.3 | 15.9 | 14.3 | 14.1 | 7.5 | 1.6 | 15.9 | 14.3 | 7.6 | 2.7 |
| Temperature (°C) | 19.0 | 19.0 | 18.8 | 18.5 | 19.0 | 18.5 | 18.3 | 18.0 | 17.5 | 17.0 |
| Salinity (psu) | 37.8 | 37.8 | 37.7 | 37.5 | 37.1 | 37.5 | 37.6 | 37.2 | 26.8 | 27.8 |
| O2 (ppm) | 6.25 | 5.55 | 6.49 | 6.13 | 6.35 | 6.10 | 5.35 | 5.80 | 6.45 | 3.53 |
| pH | 8.22 | 8.18 | 8.15 | 8.10 | 8.11 | 8.09 | 8.14 | 8.14 | 7.82 | 7.90 |
| Mean Grain Size (mm) | 7.5 | 10.0 | 6.9 | 7.0 | 7.9 | 4.8 | 9.0 | 7.5 | 0.5 | 3.6 |
| Suspended Organic Matter (mg·L−1) | 1.05 | 1.05 | 0.79 | 0.39 | 0.79 | 0.25 | 1.74 | 1.96 | 0.73 | 0.39 |
| Chlorophyll a (μg·L−1) | 0.30 | 0.30 | 0.30 | 0.25 | 0.30 | 0.20 | 0.30 | 0.26 | 0.14 | 0.15 |
| Nitrogen/Carbon % | 11.2 | 11.8 | 10.3 | 10.0 | 9.8 | 9.3 | 8.7 | 8.1 | 9.8 | 9.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bianchi, C.N.; Gerovasileiou, V.; Morri, C.; Froglia, C. Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review. Diversity 2022, 14, 176. https://doi.org/10.3390/d14030176
Bianchi CN, Gerovasileiou V, Morri C, Froglia C. Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review. Diversity. 2022; 14(3):176. https://doi.org/10.3390/d14030176
Chicago/Turabian StyleBianchi, Carlo Nike, Vasilis Gerovasileiou, Carla Morri, and Carlo Froglia. 2022. "Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review" Diversity 14, no. 3: 176. https://doi.org/10.3390/d14030176
APA StyleBianchi, C. N., Gerovasileiou, V., Morri, C., & Froglia, C. (2022). Distribution and Ecology of Decapod Crustaceans in Mediterranean Marine Caves: A Review. Diversity, 14(3), 176. https://doi.org/10.3390/d14030176