
COI Metabarcoding Provides Insights into the Highly Diverse Diet of a Generalist Salamander, Salamandra salamandra (Caudata: Salamandridae)


Abstract
:1. Introduction
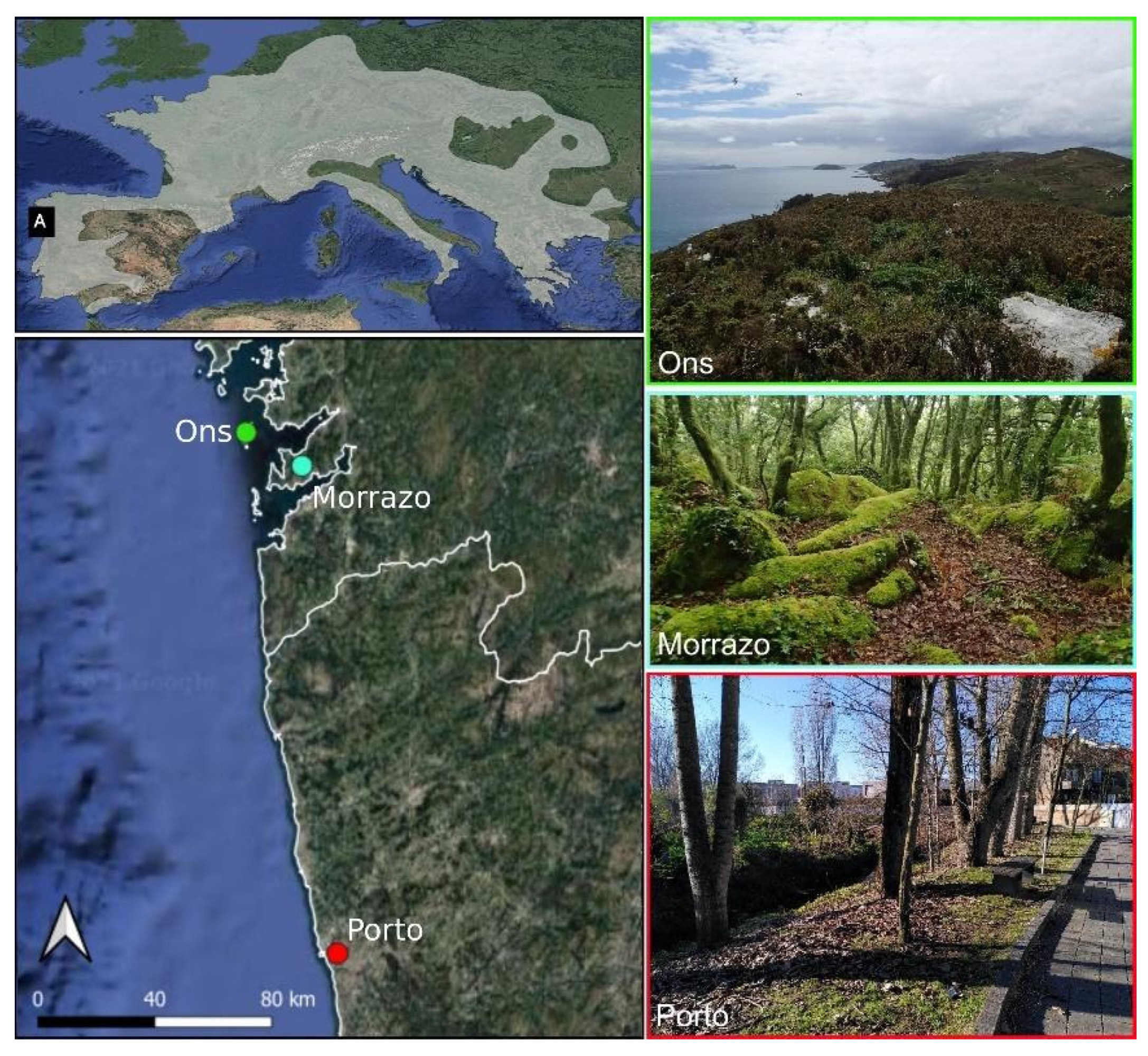
2. Materials and Methods
3. Results
3.1. Sample Collection and Sequence Amplification
3.2. Diet Characterization
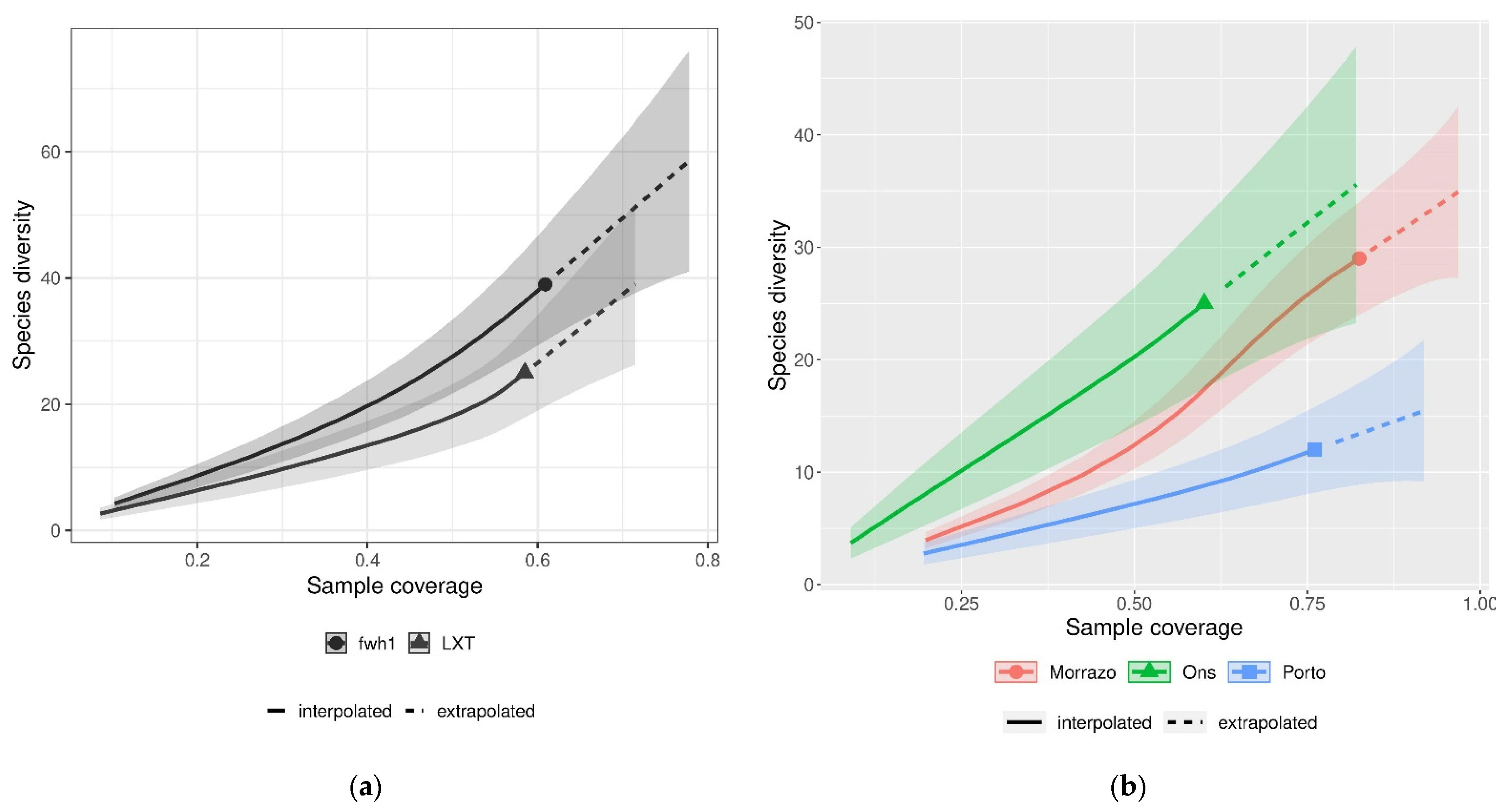
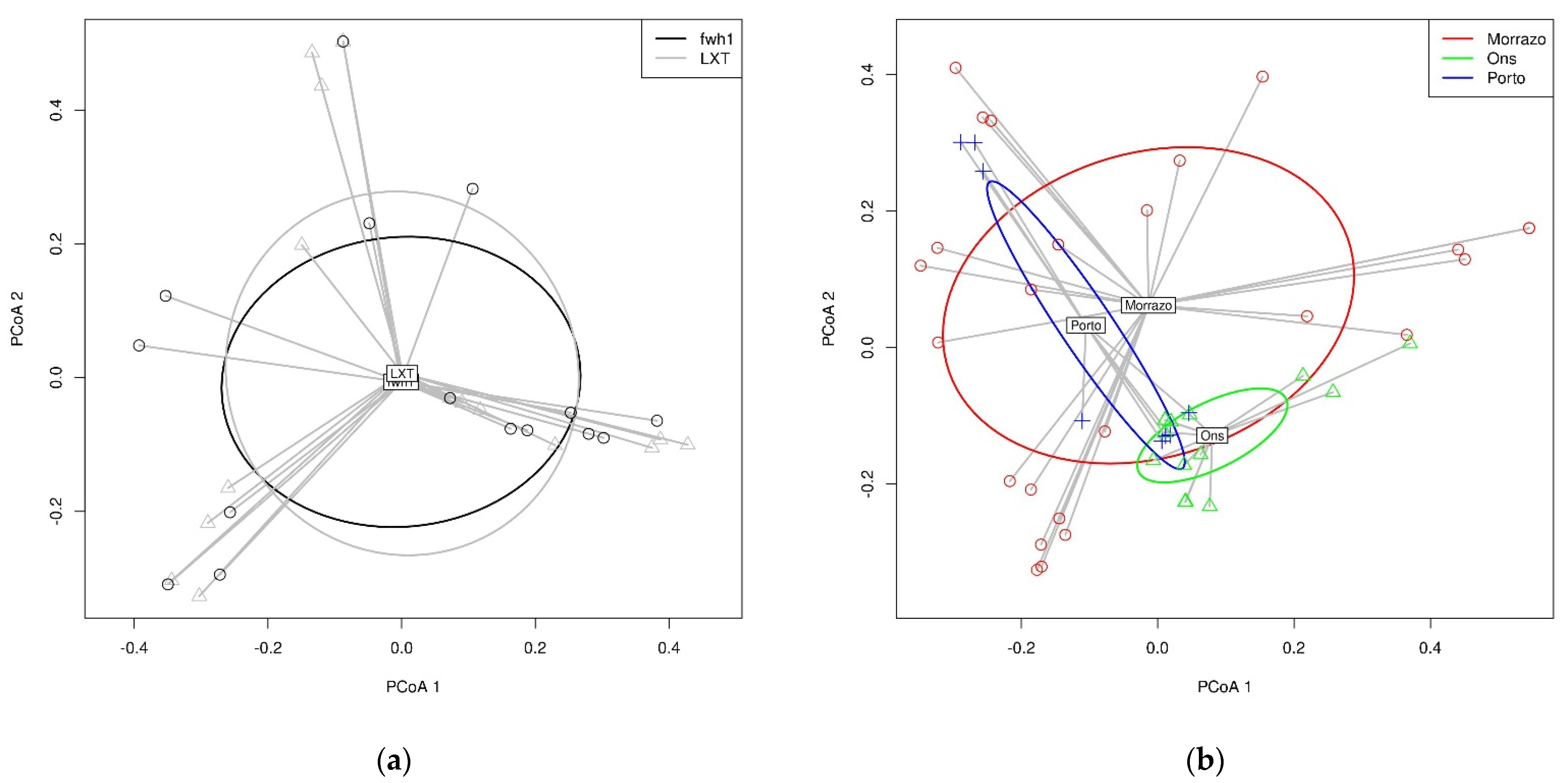
3.3. Species Richness
4. Discussion
4.1. COI Metabarcoding for Salamanders’ Diet Characterization
4.2. COI Primers as Barcodes
4.3. Dietary Variation across Regions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Losos, J.B.; Greene, H.W. Ecological and evolutionary implications of diet in monitor lizards. Biol. J. Linn. Soc. 1988, 35, 379–407. [Google Scholar] [CrossRef]
- Svanbäck, R.; Quevedo, M.; Olsson, J.; Eklöv, P. Individuals in food webs: The relationships between trophic position, omnivory and among-individual diet variation. Oecologia 2015, 178, 103–114. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Manenti, R.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Corti, C.; Ficetola, G.F.; Mancinell, G. Interspecific and inter-population variation in individual diet specialization: Do environmental factors have a role? Ecology 2020, 101, e03088. [Google Scholar] [CrossRef]
- Lunghi, E.; Cianferoni, F.; Ceccolini, F.; Veith, M.; Manenti, R.; Mancinelli, G.; Corti, C.; Ficetola, G.F. What shapes the trophic niche of European plethodontid salamanders? PLoS ONE 2018, 13, e0205672. [Google Scholar] [CrossRef]
- De Sousa, L.L.; Silva, S.M.; Xavier, R. DNA metabarcoding in diet studies: Unveiling ecological aspects in aquatic and terrestrial ecosystems. Environ. DNA 2019, 1, 199–214. [Google Scholar] [CrossRef] [Green Version]
- Ando, H.; Mukai, H.; Komura, T.; Dewi, T.; Ando, M.; Isagi, Y. Methodological trends and perspectives of animal dietary studies by noninvasive fecal DNA metabarcoding. Environ. DNA 2020, 2, 391–406. [Google Scholar] [CrossRef]
- Ficetola, G.F.; Manenti, R.; Taberlet, P. Environmental DNA and metabarcoding for the study of amphibians and reptiles: Species distribution, the microbiome, and much more. Amphibia-Reptilia 2019, 40, 129–148. [Google Scholar] [CrossRef] [Green Version]
- Laking, A.E.; Li, Z.; Goossens, E.; Miñarro, M.; Beukema, W.; Lens, L.; Bonte, D.; Verheyen, K.; Pasmans, F.; Martel, A. Salamander loss alters litter decomposition dynamics. Sci. Total Environ. 2021, 776, 145994. [Google Scholar] [CrossRef]
- Peterman, W.E.; Crawford, J.A.; Semlitsch, R.D. Productivity and significance of headwater streams: Population structure and biomass of the black-bellied salamander (Desmognathus quadramaculatus). Freshw. Biol. 2007, 53, 347–357. [Google Scholar] [CrossRef]
- Davic, R.D.; Welsh Jr, H.H. On the ecological roles of salamanders. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 405–434. [Google Scholar] [CrossRef] [Green Version]
- Walton, B.M. Salamanders in forest-floor food webs: Environmental heterogeneity affects the strength of top-down effects. Pedobiologia 2005, 49, 381–393. [Google Scholar] [CrossRef]
- Hocking, D.J.; Babbitt, K.J. Effects of red-backed salamanders on ecosystem functions. PLoS ONE 2014, 9, e86854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, A.; Salvidio, S.; Posillico, M.; Altea, T.; Matteucci, G.; Romano, A. What goes in does not come out: Different non-lethal dietary methods give contradictory interpretations of prey selectivity in amphibians. Amphibia-Reptilia 2014, 35, 255–262. [Google Scholar] [CrossRef]
- Crovetto, F.; Romano, A.; Salvidio, S. Comparison of two non-lethal methods for dietary studies in terrestrial salamanders. Wildl. Res. 2012, 39, 266–270. [Google Scholar] [CrossRef]
- Bas Lopez, S.; Gutian Rivera, J.; Castro Lorenzo, A.d.; Sanchez Canals, J.L. Data on the diet of salamander (Salamandra salamandra L.) in Galicia. Bol. Estac. Cent. Ecol. 1979, 8, 73–78. [Google Scholar]
- Çiçek, K.; Koyun, M.; Tok, C.V. Food composition of the Near Eastern Fire Salamander, Salamandra infraimmaculata Martens, 1885 (Amphibia: Urodela: Salamandridae) from Eastern Anatolia. Zool. Middle East 2017, 63, 130–135. [Google Scholar] [CrossRef]
- Solé, M.; Rödder, D. Dietary assessments of adult amphibians. In Amphibian Ecology and Conservation: A Handbook of Techniques; Dodd, C.K., Jr., Ed.; Oxford University Press: Oxford, UK, 2009; pp. 167–184. [Google Scholar]
- Salvidio, S.; Pasmans, F.; Bogaerts, S.; Martel, A.; van de Loo, M.; Romano, A. Consistency in trophic strategies between populations of the Sardinian endemic salamander Speleomantes imperialis. Anim. Biol. 2017, 67, 1–16. [Google Scholar] [CrossRef]
- Alberdi, A.; Aizpurua, O.; Bohmann, K.; Gopalakrishnan, S.; Lynggaard, C.; Nielsen, M.; Gilbert, M.T.P. Promises and pitfalls of using high-throughput sequencing for diet analysis. Mol. Ecol. Resour. 2019, 19, 327–348. [Google Scholar] [CrossRef]
- Unger, S.D.; Williams, L.A.; Diaz, L.; Jachowski, C.B. DNA barcoding to assess diet of larval eastern hellbenders in North Carolina. Food Webs 2020, 22, e00134. [Google Scholar] [CrossRef]
- Unger, S.D. Adult female Eastern hellbender Cryptobranchus alleganiensis (Cryptobranchidae) conspecific cannibalism confirmed via DNA barcoding. Herpetol. Notes 2020, 13, 169–170. [Google Scholar]
- Wang, Y.; Smith, H.K.; Goossens, E.; Hertzog, L.; Bletz, M.C.; Bonte, D.; Verheyen, K.; Lens, L.; Vences, M.; Pasmans, F.; et al. Diet diversity and environment determine the intestinal microbiome and bacterial pathogen load of fire salamanders. Sci. Rep. 2021, 11, 20493. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. B 2003, 270, S96–S99. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Herbert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Borrell, Y.J.; Miralles, L.; Do Huu, H.; Mohammed-Geba, K.; Garcia-Vazquez, E. DNA in a bottle—Rapid metabarcoding survey for early alerts of invasive species in ports. PLoS ONE 2017, 12, e0183347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grey, E.K.; Bernatchez, L.; Cassey, P.; Deiner, K.; Deveney, M.; Howland, K.L.; Lacoursière-Roussel, A.; Leong, S.C.Y.; Li, Y.; Olds, B.; et al. Effects of sampling effort on biodiversity patterns estimated from environmental DNA metabarcoding surveys. Sci. Rep. 2018, 8, 8843. [Google Scholar] [CrossRef] [Green Version]
- Wangensteen, O.S.; Palacín, C.; Guardiola, M.; Turon, X. DNA metabarcoding of littoral hard-bottom communities: High diversity and database gaps revealed by two molecular markers. PeerJ 2018, 6, e4705. [Google Scholar] [CrossRef]
- Velo-Antón, G.; Buckley, D. Salamandra común—Salamandra salamandra. In Enciclopedia Virtual de los Vertebrados Españoles; Salvador, A., Martínez-Solano, I., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2015; Available online: www.vertebradosibericos.org/ (accessed on 22 November 2021).
- Pereira, A.; Samlali, M.A.; S’Khifa, A.; Slimani, T.; Harris, D.J. A pilot study on the use of DNA metabarcoding for diet analysis in a montane amphibian population from North Africa. Afric. J. Herpetol. 2021, 70, 68–74. [Google Scholar] [CrossRef]
- Zinger, L.; Bonin, A.; Alsos, I.G.; Bálint, M.; Bik, H.; Boyer, F.; Chariton, A.A.; Creer, S.; Coissac, E.; Deagle, B.E.; et al. DNA metabarcoding-Need for robust experimental design to draw sound ecological conclusions. Mol. Ecol. 2019, 28, 1857–1862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, L.J.; Soubrier, J.; Weyrich, L.S.; Cooper, A. Environmental metabarcodes for insects: In silico PCR reveals potential for taxonomic bias. Mol. Ecol. Resour. 2014, 14, 1160–1170. [Google Scholar] [CrossRef]
- Deagle, B.E.; Jarman, S.N.; Coissac, E.; Pompanon, F.; Taberlet, P. DNA metabarcoding and the cytochrome c oxidase subunit I marker: Not a perfect match. Biol. Lett. 2014, 10, 20140562. [Google Scholar] [CrossRef] [Green Version]
- da Silva, L.P.; Mata, V.A.; Lopes, P.B.; Lopes, R.J.; Beja, P. High-resolution multi-marker DNA metabarcoding reveals sexual dietary differentiation in a bird with minor dimorphism. Ecol. Evol. 2020, 10, 10364–10373. [Google Scholar] [CrossRef]
- Vamos, E.; Elbrecht, V.; Leese, F. Short COI markers for freshwater macroinvertebrate metabarcoding. PeerJ Prepr. 2017, 1, e14625. [Google Scholar] [CrossRef] [Green Version]
- Geller, J.; Meyer, C.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Lopes, R.J.; Pinho, C.J.; Santos, B.; Seguro, M.; Mata, V.A.; Egeter, B.; Vasconcelos, R. Intricate trophic links between threatened vertebrates confined to a small island in the Atlantic Ocean. Ecol. Evol. 2019, 9, 4994–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adey, A.; Morrison, H.G.; Asan; Xun, X.; Kitzman, J.O.; Turner, E.H.; Stackhouse, B.; MacKenzie, A.P.; Caruccio, N.C.; Zhang, X.; et al. Rapid, low-input, low-bias construction of shotgun fragment libraries by high-density in vitro transposition. Genome Biol. 2010, 11, R119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. PEAR: A fast and accurate Illumina Paired-End reAd merger. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [Green Version]
- Boyer, F.; Mercier, C.; Bonin, A.; Le Bras, Y.; Taberlet, P.; Coissac, E. obitools: A unix-inspired software package for DNA metabarcoding. Mol. Ecol. Resour. 2016, 16, 176–182. [Google Scholar] [CrossRef] [PubMed]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Mata, V.A.; Ferreira, S.; Campos, R.M.; da Silva, L.P.; Veríssimo, J.; Corley, M.F.V.; Beja, P. Efficient assessment of nocturnal flying insect communities by combining automatic light traps and DNA metabarcoding. Environ. DNA 2020, 3, 398–408. [Google Scholar] [CrossRef]
- Frøslev, T.G.; Kjøller, R.; Bruun, H.H.; Ejrnæs, R.; Brunbjerg, A.K.; Pietroni, C.; Hansen, A.J. Algorithm for post-clustering curation of DNA amplicon data yields reliable biodiversity estimates. Nat. Commun. 2017, 8, 1188. [Google Scholar] [CrossRef]
- Buchner, D.; Leese, F. BOLDigger—A Python package to identify and organise sequences with the Barcode of Life Data systems. Metabarcoding Metagenom. 2020, 4, 19–21. [Google Scholar] [CrossRef]
- Deagle, B.E.; Thomas, A.C.; McInnes, J.C.; Clarke, L.J.; Vesterinen, E.J.; Clare, E.L.; Kartzinel, T.R.; Paige, E. Counting with DNA in metabarcoding studies: How should we convert sequence reads to dietary data? Mol. Ecol. 2019, 28, 391–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill, numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Jost, L. Coverage-based rarefaction and extrapolation: Standardizing samples by completeness rather than size. Ecology 2012, 93, 2533–2547. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-7. 2020. Available online: https://CRAN.R-project.org/package=vegan (accessed on 30 December 2021).
- Clarke, K.R. Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 1993, 18, 117–143. [Google Scholar] [CrossRef]
- Balogová, M.; Miková, E.; Orendáš, P.; Uhrin, M. Trophic spectrum of adult Salamandra salamandra in the Carpathians with the first note on food intake by the species during winter. Herpetol. Notes 2015, 8, 371–377. [Google Scholar]
- Piñol, J.; Senar, M.A.; Symondson, W.O.C. The choice of universal primers and the characteristics of the species mixture determine when DNA metabarcoding can be quantitative. Mol. Ecol. 2019, 28, 407–419. [Google Scholar] [CrossRef]
- Taberlet, P.; Bonin, A.; Zinger, L.; Coissac, E. DNA sequencing. In Environmental DNA: For Biodiversity Research and Monitoring; Oxford University Press: Oxford, UK, 2018. [Google Scholar] [CrossRef]
- Andújar, C.; Creedy, T.J.; Arribas, P.; López, H.; Salces-Castellano, A.; Pérez-Delgado, A.J.; Vogler, A.P.; Emerson, B.C. Validated removal of nuclear pseudogenes and sequencing artefacts from mitochondrial metabarcode data. Mol. Ecol. Resour. 2021, 21, 1772–1787. [Google Scholar] [CrossRef]
- Weisrock, D.W.; Hime, P.M.; Nunziata, S.O.; Jones, K.S.; Murphy, M.O.; Hotaling, S.; Kratovil, J.D. Surmounting the Large-Genome “Problem” for Genomic Data Generation in Salamanders. In Population Genomics: Wildlife; Hohenlohe, P.A., Rajora, O.P., Eds.; Springer International Publishing AG: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Russell, J.C.; Kueffer, C. Island Biodiversity in the Anthropocene. Annu. Rev. Environ. Resour. 2019, 44, 31–60. [Google Scholar] [CrossRef]
- Steinmetz, R.; Seuaturien, N.; Intanajitjuy, P.; Inrueang, P.; Prempree, K. The effects of prey depletion on dietary niches of sympatric apex predators in Southeast Asia. Integr. Zool. 2021, 16, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Ferenti, S.; David, A.; Nagy, D. Feeding-behaviour responses to anthropogenic factors on Salamandra salamandra (Amphibia, Caudata). Biharean Biol. 2010, 4, 139–143. [Google Scholar]
- Van Gestel, C.; Loureiro, S.; Idar, P. Terrestrial isopods as model organisms in soil ecotoxicology: A review. ZooKeys 2018, 801, 127–162. [Google Scholar] [CrossRef] [PubMed]
- Kuzmin, S.L. Feeding ecology of Salamandra and Mertensiella: A review of data and ontogenetic evolutionary trends. Mertensiella 1994, 4, 271–286. [Google Scholar]
- Mulder, K.P.; Alarcón-Ríos, L.; Nicieza, A.G.; Fleischer, R.C.; Bell, R.C.; Velo-Antón, G. Independent evolutionary transitions to pueriparity across multiple timescales in the viviparous genus Salamandra. Mol. Phylogenet. Evol. 2022, 167, 107347. [Google Scholar] [CrossRef]
- Alarcón-Ríos, L.; Nicieza, A.G.; Kaliontzopoulou, A.; Buckley, D.; Velo-Antón, G. Evolutionary history and not heterochronic modifications associated with viviparity drive head shape differentiation in a reproductive polymorphic species, Salamandra salamandra. Evol. Biol. 2020, 47, 43–55. [Google Scholar] [CrossRef]
- Steinfartz, S.; Weitere, M.; Tautz, D. Tracing the first step to speciation: Ecological and genetic differentiation of a salamander population in a small forest. Mol. Ecol. 2007, 16, 4550–4561. [Google Scholar] [CrossRef]
- Manenti, R.; Denoël, M.; Ficetola, G.F. Foraging plasticity favours adaptation to new habitats in fire salamanders. Anim. Behav. 2013, 86, 375–382. [Google Scholar] [CrossRef]
- Velo-Antón, G.; Cordero-Rivera, A. Ethological and phenotypic divergence in insular fire salamanders: Diurnal activity mediated by predation? Acta Ethol. 2017, 20, 243–253. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Phylum | Class | Order | Family | Genus | Morrazo | Ons | Porto | Total |
|---|---|---|---|---|---|---|---|---|
| Annelida | Clitellata | Haplotaxida | Almidae | Alma | 30 * | 9 | ||
| Lumbricidae | Aporrectodea | 20 | 6 | |||||
| Dendrobaena | 18 | 9 | ||||||
| Eisenia | 12 | 6 | ||||||
| Lumbricus | 35 | 30 | 26 | |||||
| Octolasion | 6 | 3 | ||||||
| Arthropoda | 6 | 3 | ||||||
| Arachnida | Opiliones | Ischyropsalididae | Ischyropsalis | 12 | 6 | |||
| Trombidiformes | Eupodidae | 10 | 3 | |||||
| Chilopoda | Scolopendromorpha | Cryptopidae | Cryptops | 6 | 3 | |||
| Collembola | Entomobryomorpha | Entomobryidae | 10 | 3 | ||||
| Isotomidae | 10 | 3 | ||||||
| Poduromorpha | 10 | 3 | ||||||
| Hypogastruridae | Hypogastrura | 30 * | 9 | |||||
| Symphypleona | Bourletiellidae | 20 | 6 | |||||
| Dicyrtomidae | Dicyrtomina | 10 | 3 | |||||
| Sminthurididae | 10 | 3 | ||||||
| Diplopoda | Glomerida | Glomeridae | Glomeris | 65 | 31 | |||
| Julida | Julidae | Cylindroiulus | 12 | 6 | ||||
| Ommatoiulus | 47 | 13 | 26 | |||||
| Platydesmida | Andrognathidae | 18 | 9 | |||||
| Polydesmida | 18 | 9 | ||||||
| Paradoxosomatidae | Oxidus | 25 * | 6 | |||||
| Polydesmidae | Polydesmus | 88 | 38 | 51 | ||||
| Insecta | Coleoptera | Cantharidae | Cantharis | 12 | 6 | |||
| Curculionidae | Caenopsis | 12 | 6 | |||||
| Histeridae | Pactolinus | 10 | 3 | |||||
| Tenebrionidae | Nalassus | 24 | 11 | |||||
| Dermaptera | Forficulidae | Forficula | 10 | 3 | ||||
| Diptera | Dolichopodidae | Condylostylus | 12 | 6 | ||||
| Ephydridae | Scatella | 12 | 6 | |||||
| Psychodidae | Bichromomyia | 10 | 3 | |||||
| Sciaridae | Sciaridae sp. | 12 | 6 | |||||
| Sepsidae | Meropliosepsis | 6 | 3 | |||||
| Syrphidae | Eupeodes | 13 * | 3 | |||||
| Hemiptera | Aphididae | Chaitophorus | 13 * | 3 | ||||
| Hymenoptera | 25 * | 6 | ||||||
| Lepidoptera | Noctuidae | Omphaloscelis | 10 | 3 | ||||
| Peridroma | 13 * | 3 | ||||||
| Sphingidae | Manduca | 10 | 3 | |||||
| Orthoptera | Tettigoniidae | Cyrtaspis | 6 | 3 | ||||
| Malacostraca | Isopoda | Armadillidiidae | Armadillidium | 10 | 38 * | 11 | ||
| Eluma | 6 | 63 * | 17 | |||||
| Oniscidae | Oniscus | 6 | 3 | |||||
| Porcellionidae | Porcellio | 18 | 9 | |||||
| Mollusca | Gastropoda | Pulmonata | Cochlicellidae | Cochlicella | 40 * | 11 | ||
| Stylommatophora | Agriolimacidae | Deroceras | 12 | 10 | 13 | 11 | ||
| Arionidae | Arion | 82 | 30 | 49 | ||||
| Geomalacus | 6 | 3 | ||||||
| Helicidae | Oestophora | 40 * | 11 | |||||
| Hygromiidae | Portugala | 25 * | 6 | |||||
| Lauriidae | Lauria | 10 | 3 | |||||
| Limacidae | Lehmannia | 20 | 6 | |||||
| Milacidae | Milax | 40 * | 11 | |||||
| Geomitridae | Ponentina | 13 | 3 | |||||
| Oxychilidae | Oxychilus | 12 | 6 | |||||
| Testacellidae | Testacella | 10 | 3 | |||||
| Urocyclidae | Microkerkus | 6 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marques, A.J.D.; Mata, V.A.; Velo-Antón, G. COI Metabarcoding Provides Insights into the Highly Diverse Diet of a Generalist Salamander, Salamandra salamandra (Caudata: Salamandridae). Diversity 2022, 14, 89. https://doi.org/10.3390/d14020089
Marques AJD, Mata VA, Velo-Antón G. COI Metabarcoding Provides Insights into the Highly Diverse Diet of a Generalist Salamander, Salamandra salamandra (Caudata: Salamandridae). Diversity. 2022; 14(2):89. https://doi.org/10.3390/d14020089
Chicago/Turabian StyleMarques, Adam J. D., Vanessa A. Mata, and Guillermo Velo-Antón. 2022. "COI Metabarcoding Provides Insights into the Highly Diverse Diet of a Generalist Salamander, Salamandra salamandra (Caudata: Salamandridae)" Diversity 14, no. 2: 89. https://doi.org/10.3390/d14020089
APA StyleMarques, A. J. D., Mata, V. A., & Velo-Antón, G. (2022). COI Metabarcoding Provides Insights into the Highly Diverse Diet of a Generalist Salamander, Salamandra salamandra (Caudata: Salamandridae). Diversity, 14(2), 89. https://doi.org/10.3390/d14020089