
The Impact of Different Biotopes and Management Practices on the Burden of Parasites in Artificial Nests of Osmia spp. (Megachilidae) Bees

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
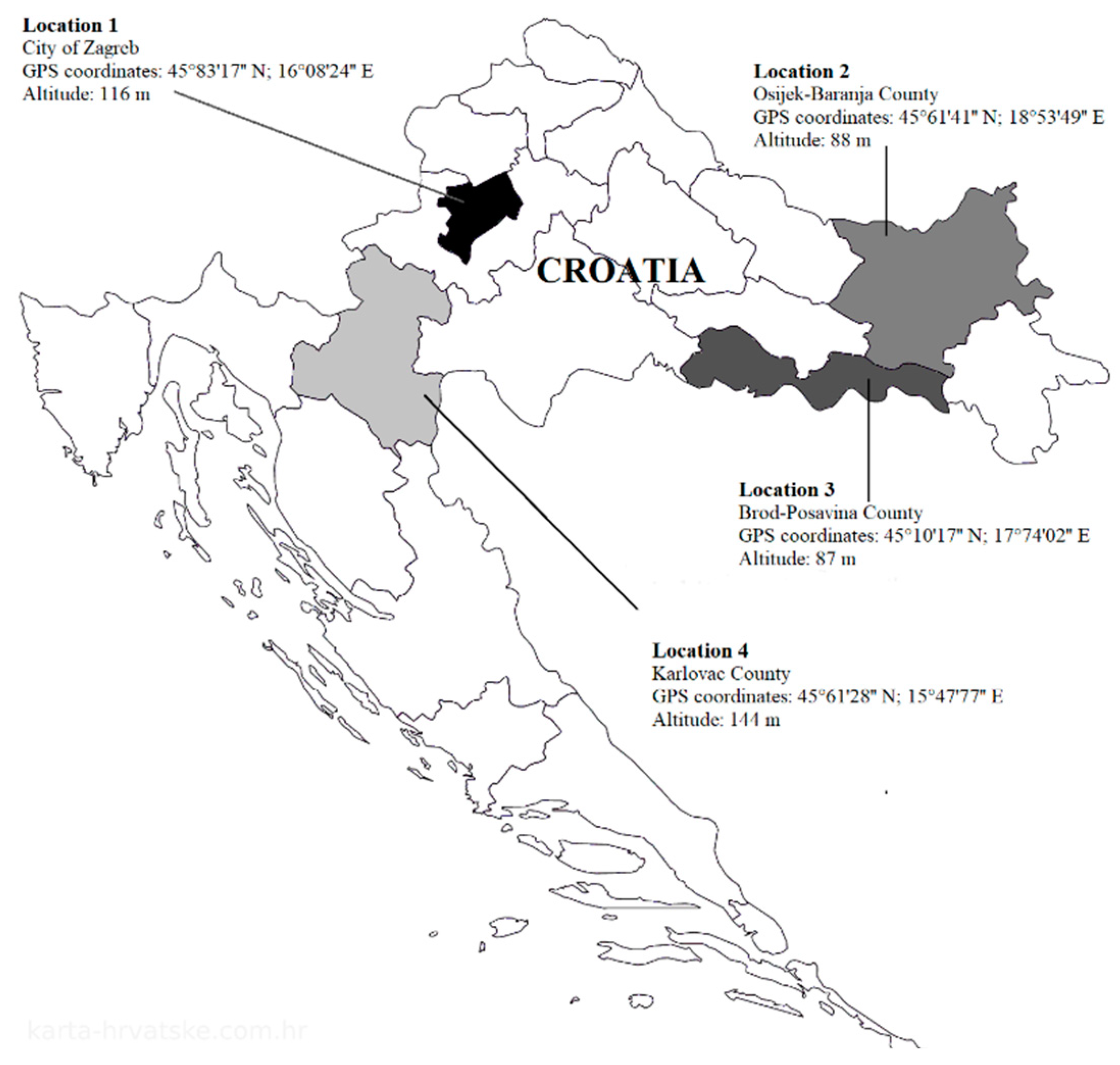
2.1. Artificial Solitary Bee’s Nests Sampling
2.2. Artificial Nests Section Analyses
2.3. Estimation of the Osmia spp. Bee’s Survival Level
- I.
- Failures included:
- The number of nest brood chambers containing the undeveloped bee’s offspring, e.g., mummified and dry larvae and pupae specimens;
- The number of nest brood chambers containing the individual dead adult bees outside of their cocoons;
- The number of nest brood chambers containing parasites, predators or pests;
- The number of nest brood chambers containing unused pollen pellets.
- II.
- The number of live and healthy adult bee specimens (non-symptomatic bees, bees free of parasites and predation, bees without visible characteristic clinical symptoms of diseases) which are fully developed in cocoons.
2.4. Laboratory Microscopic and Molecular Diagnostic of Nosema spp.
2.5. Statistical Analyses
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krunić, M.; Stanisavljević, L.J. Augmentation of managed populations of Osmia cornuta and O. rufa (Hymenoptera: Megachilidae) in Southeastern Europe. Eur. J. Entomol. 2006, 103, 695–697. [Google Scholar] [CrossRef] [Green Version]
- Maclvor, J.S.; Packer, L. The bees among us: Modelling Occupancy of Solitary Bees. PLoS ONE 2016, 11, e0164764. [Google Scholar] [CrossRef]
- Sedivy, C.; Dorn, S. Towards a sustainable management of bees of the subgenus Osmia (Megachilidae; Osmia) as fruit tree pollinators. Apidologie 2014, 45, 88–105. [Google Scholar] [CrossRef] [Green Version]
- Bosch, J.; Kemp, W.P. Developing and establishing bee species as crop pollinators: The example of Osmia spp. (Hymenoptera: Megachilidae) and fruit trees. Bull. Entomol. Res. 2002, 92, 3–16. [Google Scholar] [PubMed]
- Łoś, A.; Skórka, P.; Strachecka, A.; Winiarczyk, S.; Adaszek, L.; Winiarczyk, M.; Wolski, D. The associations among the breeding performance of Osmia bicornis L. (Hymenoptera: Megachilidae), burden of pathogens and nest parasites along urbanisation gradient. Sci. Total Environ. 2020, 710, 135520. [Google Scholar] [CrossRef] [PubMed]
- Carvalheiro, L.G.; Kunin, W.E.; Keil, P.; Aguirre-Gutiérrez, J.; Ellis, W.N.; Fox, R.; Groom, Q.; Hennekens, S.; Van Landuyt, W.; Maes, D.; et al. Species richness declines and biotic homogenisation have slowed down for NW-European pollinators and plants. Ecol. Lett. 2013, 16, 1416–1417. [Google Scholar] [CrossRef]
- Nieto, A.; Roberts, S.P.M.; Kemp, J.; Rasmont, P.; Kuhlmann, M.; García Criado, M.; Biesmeijer, J.C.; Bogusch, P.; Dathe, H.H.; De la Rúa, P.; et al. European Red List of Bees; Publication Office of the European Union: Luxembourg, 2014. [Google Scholar]
- Wagner, D.L. Insects declines in the Anthropocene. Ann. Rev. Entomol. 2020, 65, 457–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tlak Gajger, I.; Sakač, M.; Gregorc, A. Impact of Thiamethoxam on Honey Bee Qeeen (Apis mellifera) Reproductive Morphology and Physiology. Bull. Environ. Contam. Toxicol. 2017, 99, 297–302. [Google Scholar] [CrossRef]
- Laurino, D.; Lioy, S.; Carisio, L.; Manino, A.; Porporato, M. Vespa velutina: An Alien Driver of Honey Bee Colony Losses. Diversity 2020, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Strobl, V.; Bruckner, S.; Radford, S.; Wolf, S.; Albrecht, M.; Villamar-Bouza, L.; Maitip, J.; Kolari, E.; Chantawannakul, P.; Glauser, G.; et al. No impact of neonicotinoids on male solitary bees Osmia cornuta under semi-field conditions. Physiol. Entomol. 2021, 46, 105–109. [Google Scholar] [CrossRef]
- Tlak Gajger, I.; Šimenc, l.; Toplak, I. The First Detection and Genetic Characterization of Four Different Honeybee Viruses in Wild Bumblebees from Croatia. Pathogens 2021, 10, 808. [Google Scholar] [CrossRef] [PubMed]
- González-Varo, J.P.; Biesmeijer, J.C.; Bommarco, R.; Potts, S.G.; Schweiger, O.; Smith, H.G.; Steffan-Dewenter, I.; Szentgyörgyi, H.; Woyciechowski, M.; Vilà, M. Combined effects of global change pressures on animal-mediated pollination. Trends Ecol. Evol. 2013, 28, 524–530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation: The impacts of urbanization on native species are poorly studied but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Giraudeau, M.; Mousel, M.; Earl, S.; McGraw, K. Parasites in the City: Degree of Urbanization Predicts Poxvirus and Coccidian Infections in House Finches (Haemorhous mexicanus). PLoS ONE 2014, 9, e86747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giejdasz, K.; Fliszkiewicz, M. Effect of temperature treatment during development of Osmia rufa L., on mortality, emergence and longevity of adults. J. Apic. Sci. 2016, 60, 221–232. [Google Scholar] [CrossRef] [Green Version]
- Kierat, J.; Szentgyörgyi, H.; Czarnoleski, M.; Woyciechowski, M. The thermal environment of the nest affects body and cell size in the solitary red mason bee (Osmia bicornis L.). J. Therm. Biol. 2017, 68, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Isaksson, C. Urbanization, oxidative stress and inflammation: A question of evolving, acclimatizing or coping with urban environmental stress. Funct. Ecol. 2015, 29, 913–923. [Google Scholar] [CrossRef]
- Kierat, J.; Filipiak, M.; Szentgyörgyi, H.; Woyciechowski, M. Predation Cues in Solitary bee Nests. J. Insect Behav. 2017, 30, 385–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krunić, M.; Stanisavljević, L.J.; Pinzauti, M.; Felicioli, A. The accompanying fauna of Osmia cornuta and Osmia rufa and effective measures of protection. Bull. Insectol. 2005, 58, 141–152. [Google Scholar]
- OIE—Office International des Epizooties. Chapter 2.2.4., Nosemosis of Honey Bees. In Manual of Diagnostic Tests and Vaccines for Terrestrial Animals; OIE: Pairs, France, 2013; Available online: http://www.oie.int/international-standard-setting/terrestrial-manual/access-online (accessed on 26 June 2021).
- Maccagnani, B.; Giacomello, F.; Fanti, M.; Gobbin, D.; Maini, S.; Angeli, G. Apis mellifera and Osmia cornuta as carriers for the secondary spread of Bacillus subtilis on apple flowers. BioControl 2009, 54, 123. [Google Scholar] [CrossRef]
- Graystock, P.; Goulson, D.; Hughes, W.O.H. Parasites in bloom: Flowers aid dispersal and transmission of pollinator parasites within and between bee species. Proc. R. Soc. B 2015, 282, 20151371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Egerer, M.; Cecala, J.; Cohen, H. Wild bee conservation within urban gardens and nurseries: Effects of local and landscape management. Sustainability 2020, 12, 293. [Google Scholar] [CrossRef] [Green Version]
- Voulgari-Kokota, A.; Grimmer, G.; Steffan-Dewenter, I.; Keller, A. Bacterial community structure and succession in nests of two megachilid bee genera. FEMS Microbiol. Ecol. 2019, 95, fiy218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, K.E.; Ricigliano, V.A. Honey bee gut dysbiosis: A novel context of disease ecology. Curr. Opin. Insect Sci. 2017, 22, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Voulgari-Kokota, A.; Steffan-Dewenter, I.; Keller, A. Susceptibility of Red Mason Bee Larvae to Bacterial Threats Due to Microbiome Exchange with Imported Pollen Provisions. Insects 2020, 11, 373. [Google Scholar] [CrossRef] [PubMed]
- Gierer, F.; Vaughan, S.; Slater, M.; Thompson, H.M.; Elmore, J.S.; Girling, R.D. A review of the factors that influence pesticide residues in pollen and nectar: Future research requirements for optimizing the estimation of pollinator exposure. Environ. Pollut. 2019, 249, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowles, R.S.; Eitzer, B.D. Residues of neonicotinoid insecticides in pollen and nectar from model plants. J. Environ. Hortic. 2017, 35, 24–34. [Google Scholar] [CrossRef]
- Seidelman, K. Zur parazitenkontrolle in stammzuchten der roten mauerbiene Osmia rufa L. Wissen-Schaften Z. Univ. Halle 1990, 39, 25–34. [Google Scholar]
- Stanisavljević, L.J. The Impact of Accompaining Fauna on the Populations of Newly Domesticated Solitary Bees Osmia cornuta (Latr.) and O. rufa (L.) (Megachilidae, Hymenoptera). Master’s Thesis, Faculty of Biology, University of Belgrade, Belgrade, Serbia, 1996. [Google Scholar]
- Krunić, M.; Stanisavljević, L.J.; Brajković, M.; Tomanović, Ž.; Radović, I. Ecological studies of Osmia cornuta (Latr.) (Megachilidae, Hymenoptera) populations in Yugoslavia with special attention to their diapause. Acta Hortic. 2001, 561, 297–301. [Google Scholar] [CrossRef]
- Madras-Majewska, B.; Zajdel, B.; Boczkowska, B. The influence of nests usage on mason bee (Osmia rufa L.) survival. Ann. Warsaw Agricult. Univ.-SGGW Anim. Sci. 2011, 49, 115–119. [Google Scholar]
- Zajdel, B.; Kucharska, K.; Kucharski, D.; Fliszkiewicz, M.; Gąbka, J. Accompanying fauna of red mason bees in annual and perennial nesting sites. Med. Weter 2014, 70, 745–749. [Google Scholar]
- Zajdel, B.; Borański, M.; Kucharska, K.; Teper, D. Reproduction and Accompanying Fauna of Red Mason Bee Osmia rufa L. (syn. Osmia bicornis L.) in Areas with Different Levels of Urbanization. J. Apic. Sci. 2021, 65, 123–137. [Google Scholar] [CrossRef]
- Ravoet, J.; De Smet, L.; Meeus, I.; Smagghe, G.; Wenseleers, T.; De Graaf, D.C. Widespread occurrence of honey bee pathogens in solitary bees. J. Invertebr. Pathol. 2014, 122, 55–58. [Google Scholar] [CrossRef] [PubMed]
- Hedtke, S.M.; Blitzer, E.J.; Montgomery, G.A.; Danforth, B.N. Introduction of Non-Native Pollinators Can Lead to Trans-Continental Movement of Bee-Associated Fungi. PLoS ONE 2015, 10, e0130560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopit, A.M.; Pitts-Singer, T.L. Routes of Pesticide Exposure in Solitary, Cavity-Nesting Bees. Environ. Entomol. 2018, 47, 499–510. [Google Scholar] [CrossRef]
- Cecala, J.M.; Wilson Rankin, E.E. Pollinators and plant nurseries: How irrigation and pesticide treatment of native ornamental plants impact solitary bees. Proc. R. Soc. B 2021, 288, 20211287. [Google Scholar] [CrossRef] [PubMed]
- Stuligross, C.; Williams, N.M. Pesticide and resource stressors additively impair wild bee reproduction. Proc. R. Soc. B 2020, 287, 20201390. [Google Scholar] [CrossRef] [PubMed]
- Klaus, F.; Tscharntke, T.; Bischoff, G.; Grass, I. Floral resource diversification promotes solitary bee reproduction and may offset insecticide effects—Evidence from a semi-field experiment. Ecol. Lett. 2021, 24, 668–675. [Google Scholar] [CrossRef] [PubMed]
- Mokkapati, J.S.; Bednarska, A.J.; Laskowski, R. The development of the solitary bee Osmia bicornis is affected by some insecticide agrochemicals at environmentally relevant concentrations. Sci. Total Environ. 2021, 775, 145588. [Google Scholar] [CrossRef] [PubMed]
- Wilson Rankin, E.E.; Barney, S.K.; Lozano, G.E. Reduced water negatively impacts social bee survival and productivity via shifts in floral nutrition. J. Insect Sci. 2020, 20, 15. [Google Scholar] [CrossRef]
- Fliszkiewicz, M.; Kuśnierczak, A.; Szymaś, B. The accompanying fauna of solitary bee Osmia bicornis (L.) syn. Osmia rufa (L.) nests settled in different biotopes. J. Apic. Sci. 2012, 56, 51–58. [Google Scholar] [CrossRef] [Green Version]
- Palladini, J.D.; Maron, J.L. Reproduction and survival of a solitary bee along native and exotic floral resource gradients. Oecologia 2014, 176, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Bosch, J.; Vicens, N. Body size as an estimator of production costs in a solitary bee. Ecol. Entomol. 2002, 27, 129–137. [Google Scholar] [CrossRef]
- Schenk, M.; Mitesser, O.; Hovestadt, T.; Holzschuh, A. Overwintering temperature and body condition shift emergence dates of spring-emerging solitary bees. PeerJ 2018, 16, e4721. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
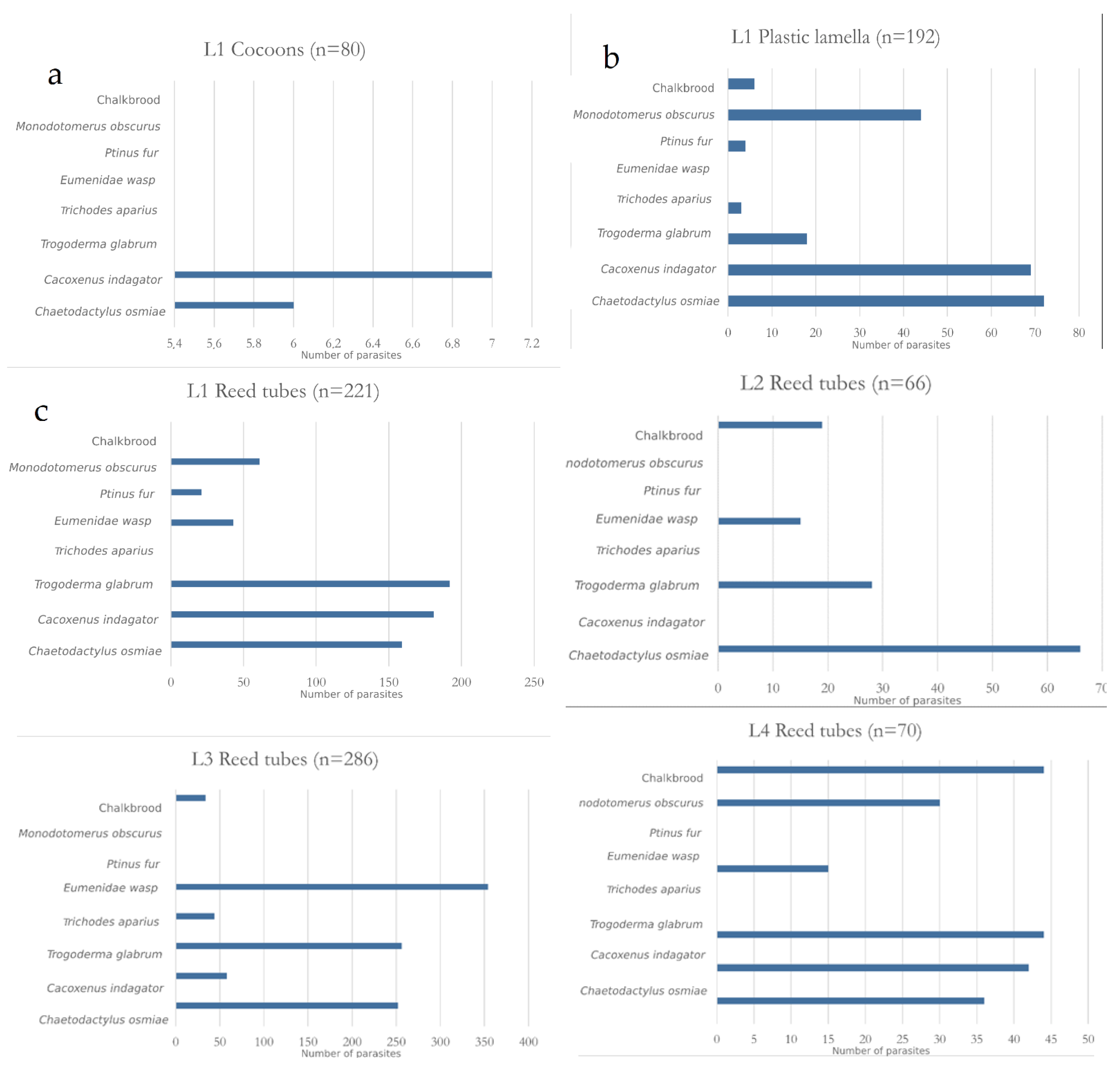{kind=link}
{kind=link}
{kind=link}
| Number of Brood Chambers | Bee Rearing Station Location | |||
|---|---|---|---|---|
| L1 (a + b + c) | L2 | L3 | L4 | |
| Empty | 0 + 403 + 592 Σ995 | 328 | 563 | 122 |
| Brood failures | 22 + 643 + 1166 Σ1831 | 218 | 1100 | 229 |
| Healthy bee | 58 + 453 + 404 Σ915 | 85 | 176 | 118 |
| Species | Location | Sample Size (n) | Mean (g) | S.D. |
|---|---|---|---|---|
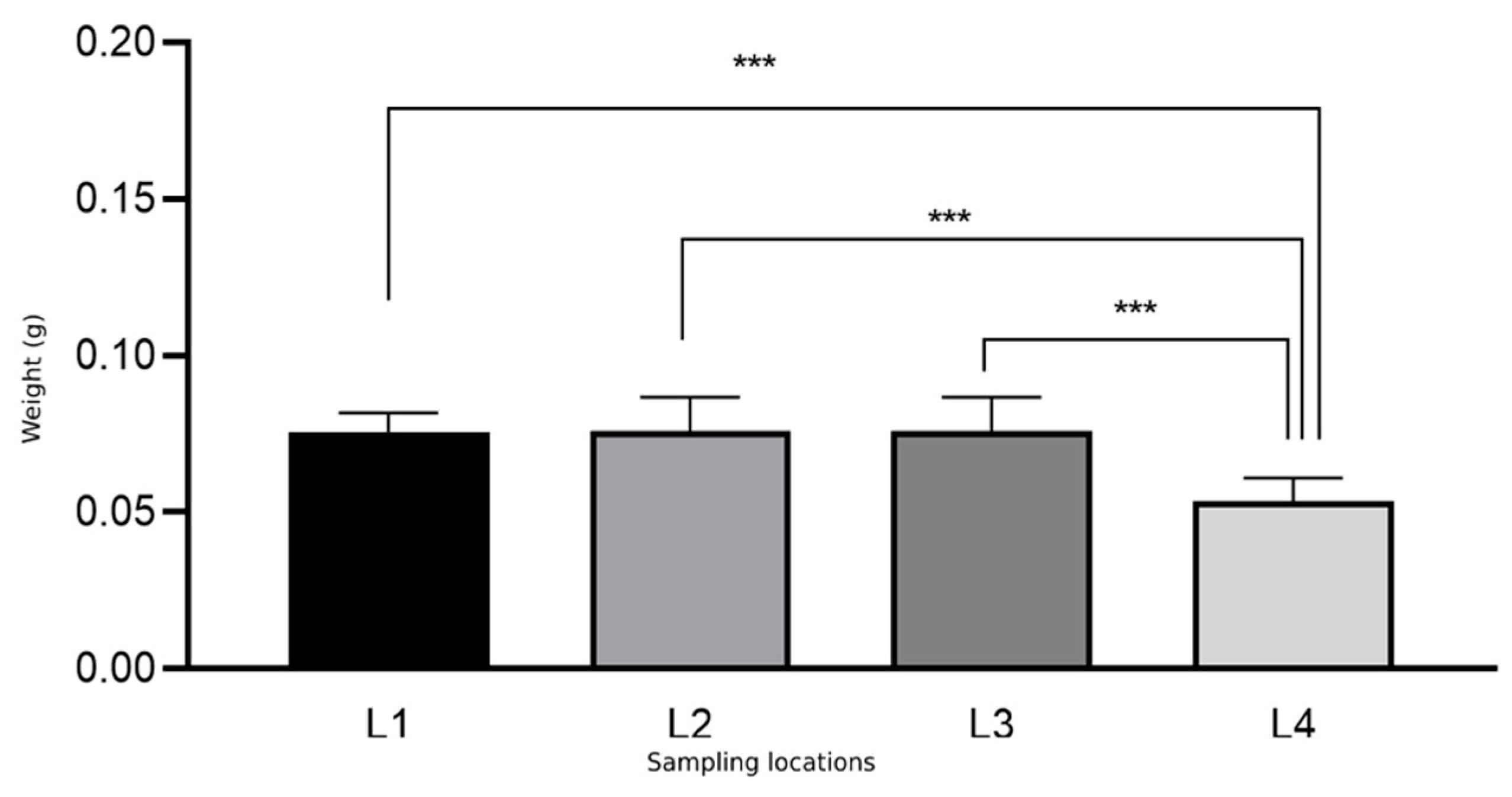
| Osmia bicornis | L1 (a + b + c) | 613 (45 + 341 + 227) | 0.075 | 0.006 |
| L2 | 38 | 0.076 | 0.010 | |
| L3 | 122 | 0.076 | 0.010 | |
| L4 | 66 | 0.053 | 0.007 | |
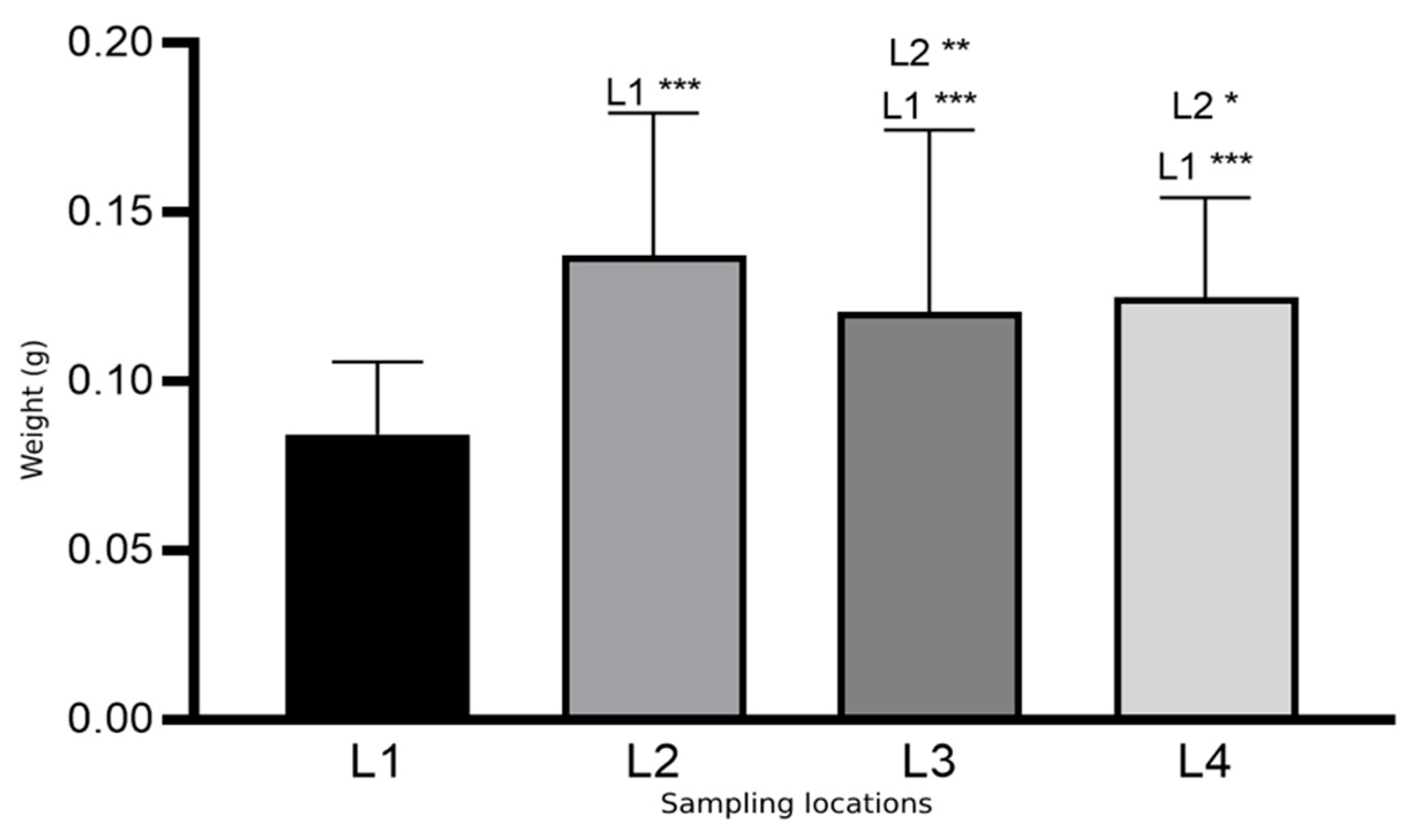
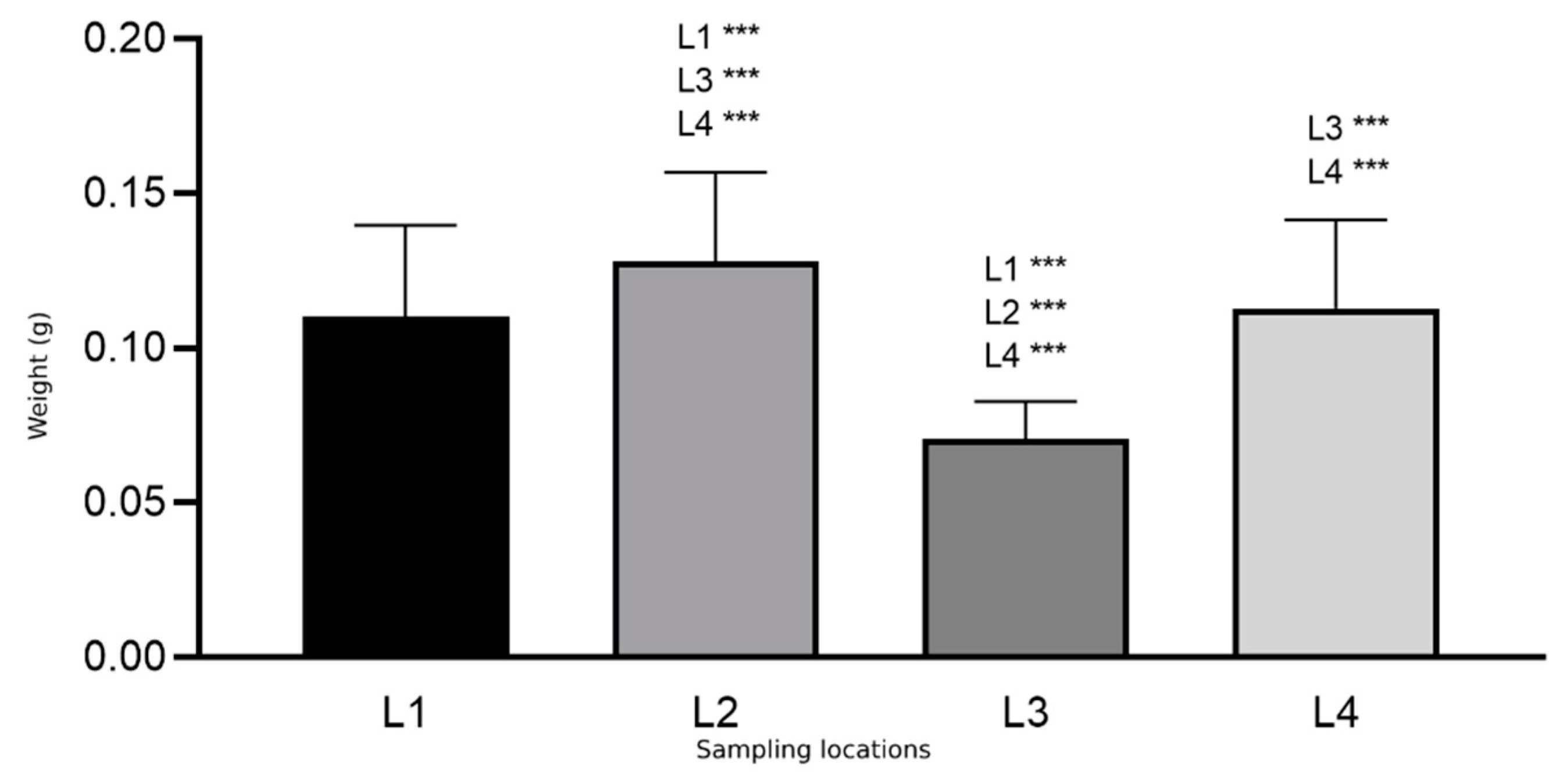
| Osmia cornuta | L1 (a + b + c) | 302 (13 + 112 + 177) | 0.110 | 0.029 |
| L2 | 47 | 0.128 | 0.028 | |
| L3 | 53 | 0.075 | 0.012 | |
| L4 | 52 | 0.112 | 0.028 |
| Location | Brood Chambers Contain | SL (%) | ||||
|---|---|---|---|---|---|---|
| Mummified Larvae and Pupae | Dead Adult Bees | Parasites, Predators or Pests | Unused Pollen Pellets | Live and Healthy Adult Bees | ||
| L1a | 2 | 7 | 13 | - | 58 | 72.5 |
| L1b | 256 | 152 | 216 | 19 | 453 | 41.33 |
| L1c | 286 Σ613 | 108 Σ267 | 657 Σ886 | 115 Σ134 | 404 Σ915 | 34.64 |
| L2 | 30 | 16 | 128 | 44 | 85 | 28.05 |
| L3 | 34 | 31 | 1028 | 7 | 176 | 13.80 |
| L4 | 9 | 3 | 211 | 6 | 118 | 34.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tlak Gajger, I.; Laklija, I.; Jurković, M.; Košćević, A.; Dar, S.A.; Ševar, M. The Impact of Different Biotopes and Management Practices on the Burden of Parasites in Artificial Nests of Osmia spp. (Megachilidae) Bees. Diversity 2022, 14, 226. https://doi.org/10.3390/d14030226
Tlak Gajger I, Laklija I, Jurković M, Košćević A, Dar SA, Ševar M. The Impact of Different Biotopes and Management Practices on the Burden of Parasites in Artificial Nests of Osmia spp. (Megachilidae) Bees. Diversity. 2022; 14(3):226. https://doi.org/10.3390/d14030226
Chicago/Turabian StyleTlak Gajger, Ivana, Ivana Laklija, Mirko Jurković, Anja Košćević, Showket Ahmad Dar, and Marija Ševar. 2022. "The Impact of Different Biotopes and Management Practices on the Burden of Parasites in Artificial Nests of Osmia spp. (Megachilidae) Bees" Diversity 14, no. 3: 226. https://doi.org/10.3390/d14030226
APA StyleTlak Gajger, I., Laklija, I., Jurković, M., Košćević, A., Dar, S. A., & Ševar, M. (2022). The Impact of Different Biotopes and Management Practices on the Burden of Parasites in Artificial Nests of Osmia spp. (Megachilidae) Bees. Diversity, 14(3), 226. https://doi.org/10.3390/d14030226