1. Introduction
The genus
Ishige Yendo (Phaeophyceae, Ishigeaceae) have three species (
I. okamurae Yendo,
I. foliacea Okamura and
I. sinicola (Setchell and N.L. Gardner) Chihara) reported from Korea, Japan and Mexico [
1]. Yendo found two life forms on a single plant body, a filiform frond with abnormal foliose fronds as one of its segments. He described this alga as
I. okamurae having different morphological forms: the filiform frond (cylindrical or terete) and the foliose frond (flat and leaf-like) [
2].
However, Okamura in Segawa [
3] treated the foliose form isolated from
I. okamurae as a new species as
I. foliacea. Moreover, the foliose type was also placed on the type specimen of
I. okamurae in the Herbarium, Graduate School of Science, Hokkaido Univ. (SAP) Japan ([
4] Figure 17). Additionally, this specimen of
I. okamurae had a stamp of “TYPUS” indicating a type specimen.
Later, a lectotype specimen of
I. foliacea was proposed on the basis of a personal communication with Kazuhiro Kogame [
5]. However, this specimen was an independent frond that was not epiphytic on the plant body of
I. okamurae and therefore did not represent
I. foliacea as originally described [
3].
Herbarium specimens include molecular information that helps to analyze taxonomic problems such as the recognition of cryptic species or species boundaries [
6,
7]. Therefore, DNA barcoding for herbarium specimens can provide valuable information to identify species boundary and to find new species [
8].
In the present study, we examined the taxonomic identity of Ishige specimens with two life forms on a single plant body of I. okamurae. Molecular analyses were conducted to clarify the taxonomic relationships between the two Ishige species. In addition, we proposed a new type specimen of I. foliacea that shows typical morphological characteristics that fit the original description of I. okamurae and I. foliacea.
2. Materials and Methods
We examined 1200 sheets of the herbarium specimens of
Ishige species collected in Korea during 2008–2020 and deposited in the National Institute of Biological Resources (NIBR; KB), Korea. These specimens have the same morphological characteristics as the filiform type of frond with abnormal foliose branches (
Table 1,
Figure 1), present in
I. okamurae as described by [
2] and in
I. foliacea following the research in [
3]. Morphological analysis was conducted using a light microscope (BX50; Olympus, Tokyo, Japan) with a digital camera (C-4040 Zoom; Olympus). We used a scanner (Epson, Seiko Epson, Nagano, Japan) to obtain images of the herbarium specimens.
Among 21 sheets of
I. okamurae specimens with epiphytic
I. foliacea, three specimens from different collection sites were selected for molecular analysis (
Table 1,
Figure 1). Each sample of the
I. okamurae and the epiphytic
I. foliacea on it was cut from a single plant body and three identical processes were conducted using three NIBR herbarium specimens.
We also re-examined the specimens of
I. okamurae and
I. sinicola analyzed by the authors in [
4]. A molecular analysis of herbarium specimens was conducted as a follow-up to previous studies [
8]. We isolated and separated the two life forms of specimens obtained from a single herbarium specimen and removed a small piece (<0.5 cm
2) for molecular phylogenetic analysis. DNA extraction, polymerase chain reaction (PCR), and sequencing were conducted following the protocols reported by Lee and Lee (2018).
We selected
cox3 region to compare with the DNA sequences of the
Ishige species previously reported [
5]. We designed PCR primers to amplify the
cox3 region from
Ishige samples (forward primer, cox3-Ish-44F, 5′-TAGTTTCTCGAAGCCCTTGG-3′; reverse primer, cox3-Ish-641R, 5′-TGGAAGCCRTGRAAACCTGT-3′), using the
cox3 DNA sequences deposited in NCBI GenBank (
www.ncbi.nlm.nih.gov/Genbank).
The PCR conditions were as follows: 3 min at 95 °C, 40 cycles of 30 s at 94 °C, 30 s at 50 °C, and 30 s at 72 °C, with a final 7 min extension at 72 °C. A commercial sequencing service (Macrogen, Seoul, Korea) was used to determine DNA sequences and Sequencher 5.4.6 (Gene Codes, Ann Arbor, MI, USA) was used to assemble the chromatograms. Phylogenetic analysis of the taxonomic relationships among
Ishige species was performed using MEGA ver. 6 [
9].
4. Discussion
Ishige okamurae was described as two life forms (the filiform type (cylindrical or terete) and the foliose type) on one plant body by [
2]. However, this type of
I. okamurae was rarely collected from Korea. Among 1200 sheets of
Ishige specimens collected from around the Korean peninsula during 2008-2020, only fifteen specimens had two life forms. Moreover, [
5] taxonomically treated Korean and Japan
I. foliacea (
I. sinicola auct. japon. and corea.) based on free-living specimens and not with epiphytic form on
I. okamurae, and designated the free-living form of the specimen as the lectotype for
I. foliacea [
5].
In the original description of the new species,
Ishige okamurae and
I. foliacea, indicated the presence of epiphytic foliose fronds and filiform fronds as the main distinguishing traits [
2,
3]. The study in [
4] recognized the absence of the type specimen of
I. okamurae and designated a lectotype of
I. okamurae consistent with Yendo’s original description and drawings; this typification was later confirmed by the authors in [
5].
Okamura in [
3] examined the form of two individuals adhering together among Yendo’s specimens, and recognized them as different species, with one attached to the other. He described the foliose form as a new species,
I. foliacea. Therefore, the lectotype of
I. foliacea should be a specimen in which two individuals are attached and corresponds more closely to the original description and drawings by the author in [
2] and Okamura in [
3].
The lectotype of
I. okamurae (in [
4]) selected among Yendo specimens is consistent with the original description of Okamura in [
3] and that of the study in [
2]. Therefore, the foliose part of the herbarium sheet, i.e., the epiphytic specimen living on the filiform specimen already designated as the lectotype of
I. okamurae should also be the lectotype of
I. foliacea. That is, the collection item in which two species are attached to each other must also be the type of
I. foliacea to match the original description and drawings of the two species. This lectotype designation fits the original description and drawings of the two species and is therefore in accordance with Art. 9.3 and Art. 9.4 of the International Code of Nomenclature (ICN) for algae, fungi, and plants [
10]. Moreover, the lectotype designation should reflect on the species description, including original materials (e.g., hand drawing, messages in notes) and carefully selected among syntypes, if possible [e.g., typification of brown alga
Laminaria rodriguezii in [
11]). However, the previous proposed lectotype specimen of
I. foliacea in [
5] could not satisfied for the species description in [
2,
3].
The lectotype of
I. foliacea was designated on the base of the annotation of Yendo’s specimen by Tadao Yoshida [
5]. However, this annotation by Yoshida did not mean any indication of intent by the original author who named the species. The lectotype designation should be based on the original materials, which were associated with the preparation of description, diagnosis, or illustration with analysis (Art. 9.3 and Art. 9.4 in the ICN). Therefore, the lectotype of
I. foliacea should follow the original description by [
2] and Okamura in [
3]. In addition, the study in [
5] analyzed only those samples in which
I. foliacea and
I. okamurae were growing independently and designated a specimen of
I. foliacea showing the free-living form as a lectotype. This is clearly inconsistent with [
2] and Okamura’s original description [
3]; that is, the foliose fronds of
I. foliacea were epiphytic on the branch of
I. okamurae.
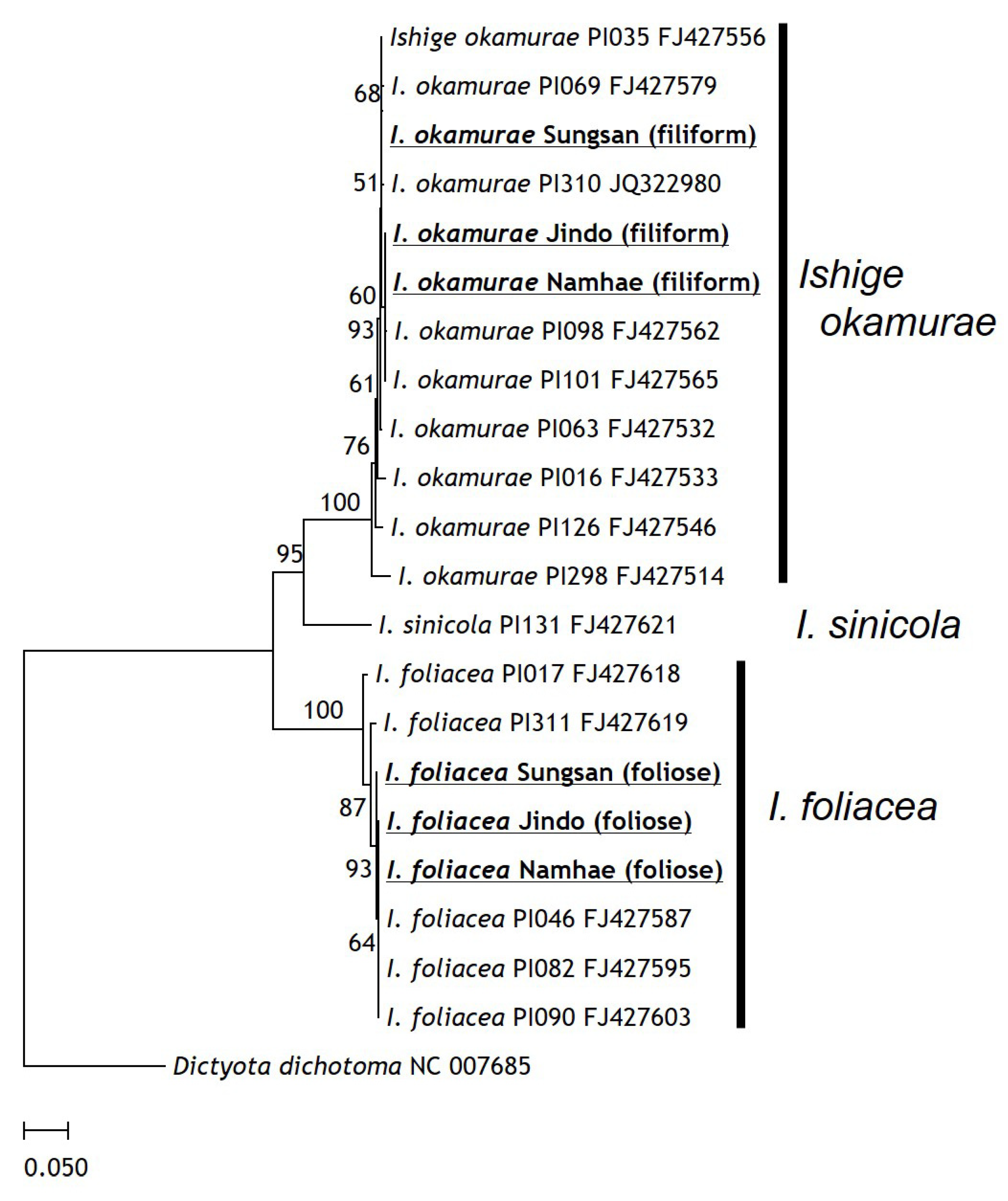
The foliose branch of filiform
I. okamurae was identified as
I. foliacea. The
cox3 sequences of the foliose fronds in the herbarium specimens showed 97.5–100% sequence similarity with individuals of
I. foliacea in free-living specimens and formed a clade apart from
I. okamurae and
I. sinicola. The filiform
I. okamurae with foliose branches showed high similarity with
I. okamurae reported from Japan and Taiwan (96.6–100%). Therefore, the rarely reported individuals with two life forms consisted of two species (
I. foliacea is epiphytic on
I. okamurae). These results are congruent with the morphological descriptions reported by the authors in [
2]. Moreover, a molecular taxonomic study based on the
cox3 showed high sequence similarity between the free-living and epiphytic
I. foliacea [
5]. This finding is interesting because it confirms that
I. foliacea is present in two life forms (independent and epiphytic on
I. okamurae). The
cox3 gene region was taxonomically sufficient to classify the three species of the genus
Ishige.
To confirm the taxonomic relationship among the three species of the genus
Ishige, molecular taxonomic analyses should be conducted on type specimens of the three species:
I. foliacea,
I. sinicola, and
I. okamurae. Studies on the taxonomic status of
Ishige sinicola in Korea should be on conducted because many algae show biogeographical distribution on both the coast of California and the coast of Northeast Asia. For example,
Padina durvillei was reported originally from Chile; however, it is distributed in cosmopolitan regions, including Asia and Africa. The type of localities of
Erithro tetraseriata and Rhodymenia californica are the coast of California; they were also discovered on the Asian coast [
1]. Even though no DNA sequence of the genus
Ishige was successfully produced from the type of specimen, the exact lectotype designation is required to prevent taxonomic confusion.
The original description given by the authors in [
2] should be a key reference for determining the type specimen. In this study, we analyzed DNA sequences from two life forms of morphotypes, including
I. okamurae and
I. foliacea. This generic information presents a useful taxonomic guideline for further studies. Moreover, the specimen of
I. foliacea epiphytic on
I. okamurae deposited at the Hokkaido University Herbarium (SAP) was observed only from the photocopy of the specimen provided by SAP. Thus, the analysis of the Japanese samples of epiphytic
I. foliacea on
I. okamurae, including the lectotype, need to be further studied.






{kind=link}
{kind=link}