
Island–to–Island Vicariance, Founder–Events and within–Area Speciation: The Biogeographic History of the Antillattus Clade (Salticidae: Euophryini)

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
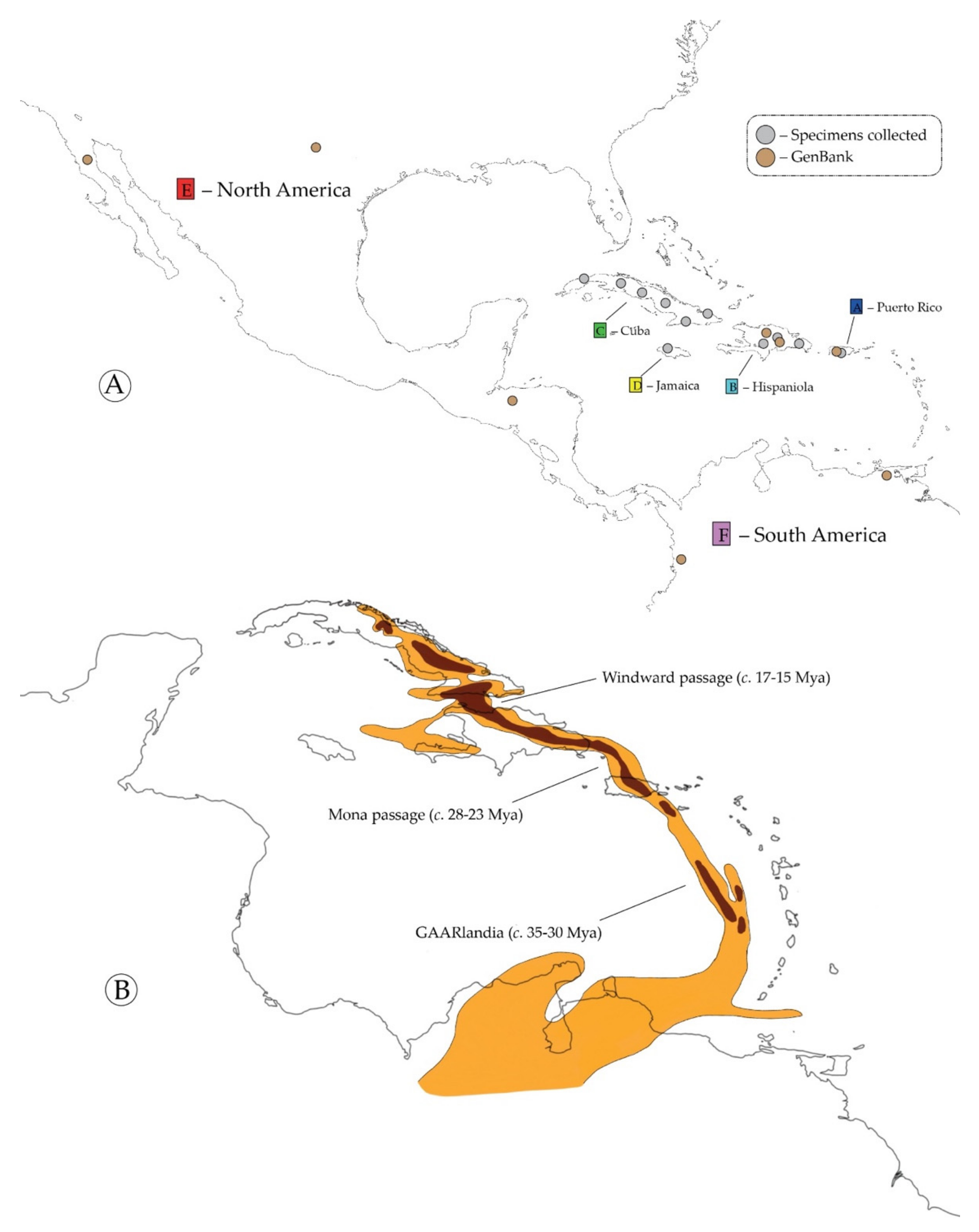
Study Group and Taxon Sample
3. DNA Extraction, Amplification and Sequencing
3.1. Phylogenetic Inference
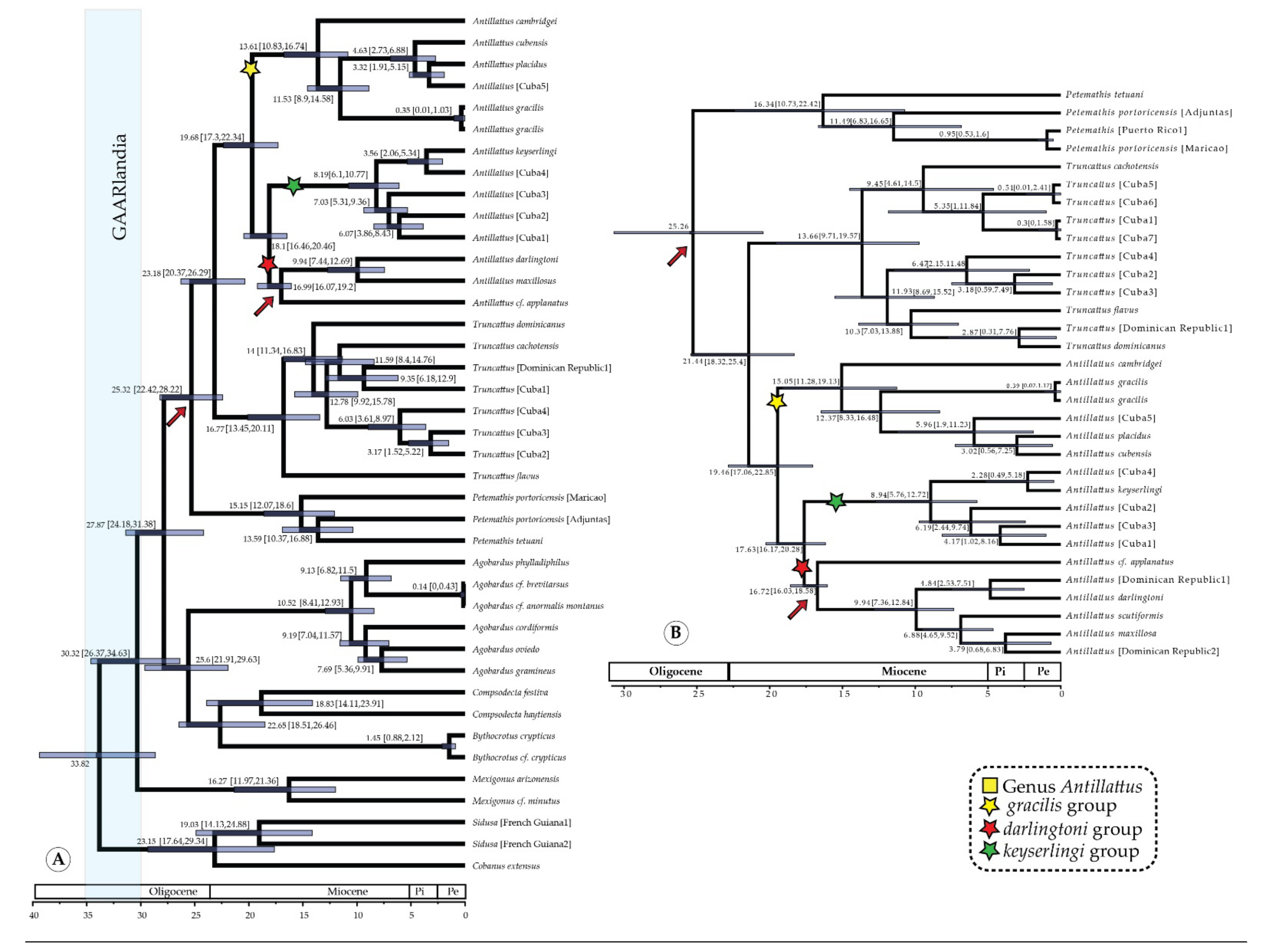
3.2. Time Calibration and Divergence Estimation
4. Biogeographical Estimation
5. Results
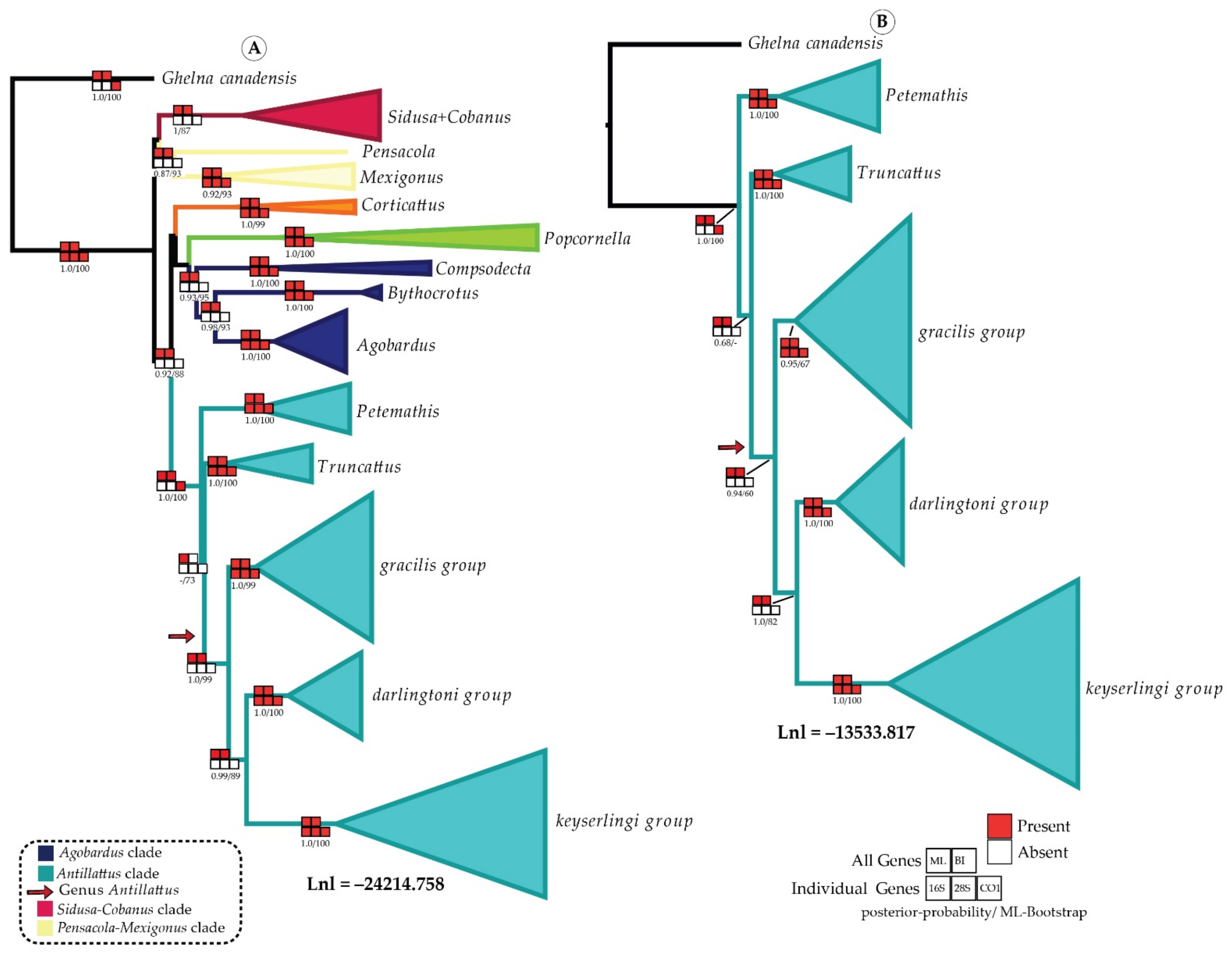
Phylogeny and Divergence Time
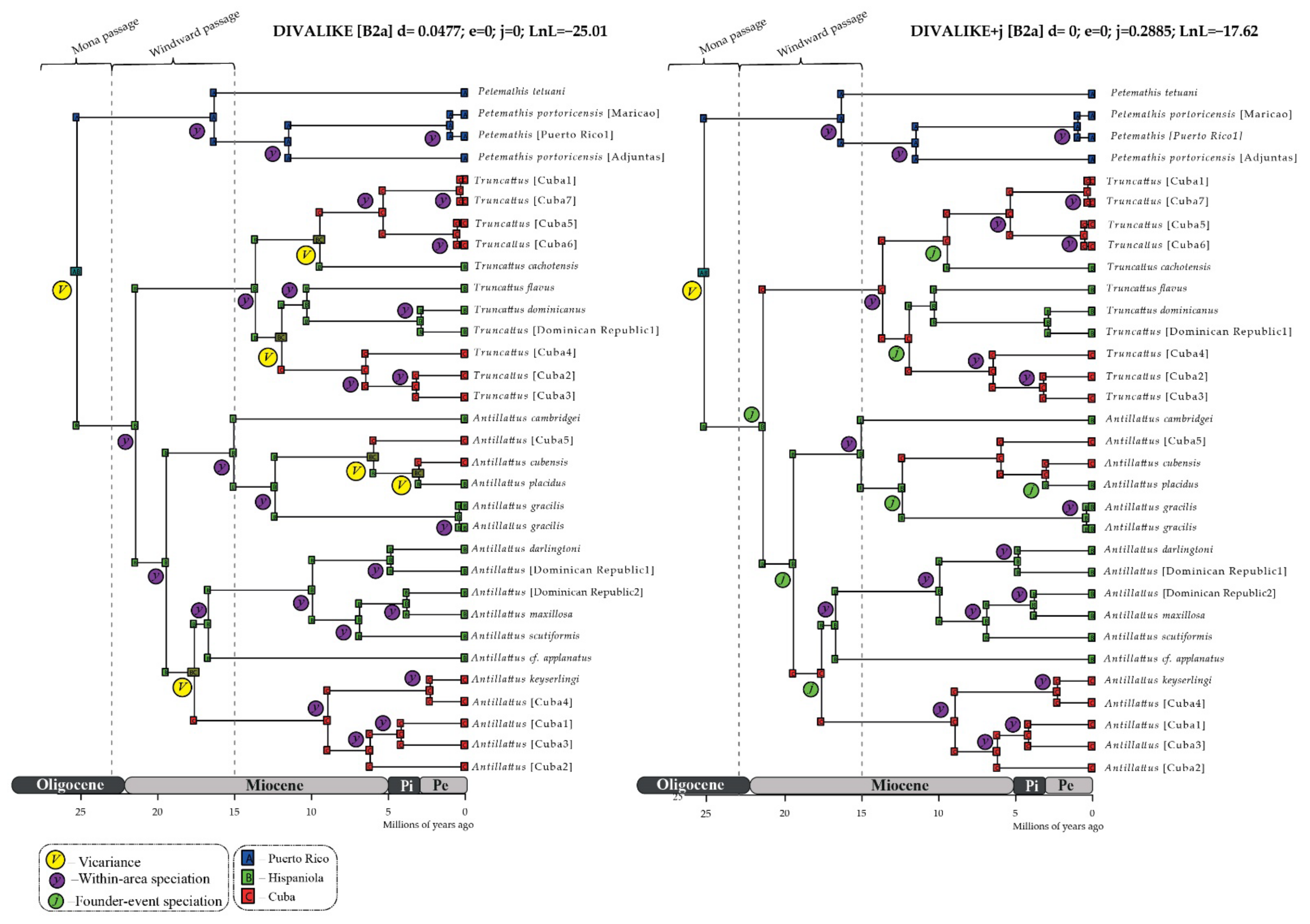
6. Model Selection and Ancestral Range Estimation
Estimation of Biogeographical Events
7. Discussion
7.1. Ancestral Range of the Antillattus Clade and GAARlandia
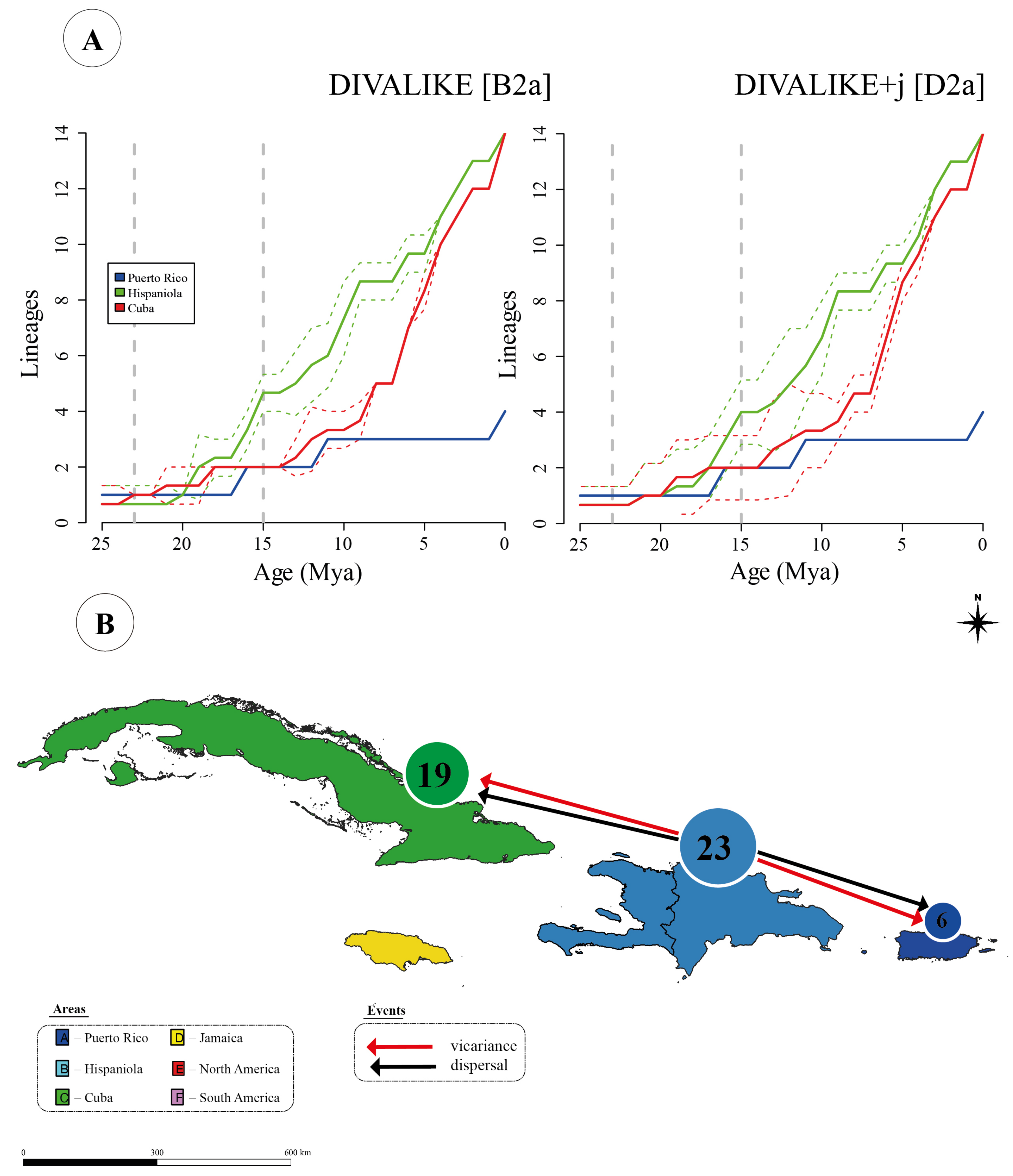
7.2. Inter-Island Biogeographical History
7.3. From Hispaniola to Cuba and Puerto Rico
8. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dávalos, L.M. Phylogeny and biogeography of Caribbean mammals. Biol. J. Linn. Soc. 2004, 8, 373–394. [Google Scholar] [CrossRef]
- Ricklefs, R.; Bermingham, E. The West Indies as a laboratory of biogeography and evolution. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2008, 363, 2393–2413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnarsson, I.; Kuntner, M. The generation of a biodiversity hotspot: Biogeography and phylogeography of the western Indian Ocean islands. In Current Topics in Phylogenetics and Phylogeography of Terrestrial and Aquatic Systems; Anamthawat-Jonsson, K., Ed.; Tech Publishers Press: Rijeka, Croatia, 2012; pp. 33–82. [Google Scholar]
- Chamberland, L.; McHugh, A.; Kechejian, S.; Binford, G.J.; Bond, J.E.; Coddington, J.; Dolman, G.; Hamilton, C.A.; Harvey, M.S.; Kuntner, M.; et al. From Gondwana to GAAR landia: Evolutionary history and biogeography of ogre-faced spiders (Deinopis). J. Biogeogr. 2018, 45, 2442–2457. [Google Scholar] [CrossRef] [Green Version]
- Matos-Maraví, P.; Águila, R.N.; Peña, C.; Miller, J.Y.; Sourakov, A.; Wahlberg, N. Causes of endemic radiation in the Caribbean: Evidence from the historical biogeography and diversification of the butterfly genus Calisto (Nymphalidae: Satyrinae: Satyrini). BMC Evol. Biol. 2014, 14, 199. [Google Scholar] [CrossRef] [Green Version]
- Gillespie, R.G.; Roderick, G.K. Arthropods on islands: Colonization, speciation, and conservation. Annu. Rev. Entomol. 2002, 47, 595–632. [Google Scholar] [CrossRef]
- De Queiroz, K. A unified species concept and its consequences for the future of taxonomy. Proc. Calif. Acad. Sci. 2005, 56, 196–215. [Google Scholar]
- Cowie, R.H.; Holland, B.S. Dispersal is fundamental to biogeography and the evolution of biodiversity on oceanic islands. J. Biogeogr. 2006, 33, 193–198. [Google Scholar] [CrossRef]
- Gillespie, R.G.; Baldwin, B.G.; Waters, J.M.; Fraser, C.I.; Nikula, R.; Roderick, G.K. Long-distance dispersal: A framework for hypothesis testing. Trends Ecol. Evol. 2012, 27, 47–55. [Google Scholar] [CrossRef]
- Schönhofer, A.L.; McCormack, M.; Tsurusaki, N.; Martens, J.; Hedin, M. Molecular phylogeny of the harvestmen genus Sabacon (Arachnida: Opiliones: Dyspnoi) reveals multiple Eocene–Oligocene intercontinental dispersal events in the Holarctic. Mol. Phylogenet. Evol. 2013, 66, 303–315. [Google Scholar] [CrossRef]
- Agnarsson, I.; Cheng, R.C.; Kuntner, M. A Multi-Clade Test Supports the Intermediate Dispersal Model of Biogeography. PLoS ONE 2014, 91, e86780. [Google Scholar] [CrossRef]
- Toussaint, E.F.A.; Fikacek, M.; Short, A.E.Z. India-Madagascar vicariance explains cascade beetle bio-geography. Biol. J. Linn. Soc. 2016, 118, 982–991. [Google Scholar] [CrossRef] [Green Version]
- Weaver, P.F.; Cruz, A.; Johnson, S.; Dupin, J.; Weaver, K.F. Colonizing the Caribbean: Biogeography and evolution of livebearing fishes of the genus Limia (Poeciliidae). J. Biogeogr. 2016, 43, 1808–1819. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Robles-Gil, P.; Hoffman, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; da Fonseca, G.A.B. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; CEMEX, Agrupación Sierra Madre: Monterrey, Mexico, 2004. [Google Scholar]
- Diamond, J.M.; Gilpin, M.E.; Mayr, E. Species-Distance Relation for Birds of Solomon Archipelago, and Paradox of Great Speciators. Proc. Natl. Acad. Sci. USA 1976, 73, 2160–2164. [Google Scholar] [CrossRef] [Green Version]
- Glor, R.E.; Losos, J.B.; Larson, A. Out of Cuba: Overwater dispersal and speciation among lizards in the Anolis carolinensis subgroup. Mol. Ecol. 2005, 14, 2419–2432. [Google Scholar] [CrossRef]
- Iturralde-Vinent, M.A. Meso-Cenozoic Caribbean paleogeography: Implications for the historical biogeography of the region. Int. Geol. Rev. 2006, 48, 791–827. [Google Scholar] [CrossRef] [Green Version]
- Crews, S.C.; Gillespie, R.G. Molecular systematics of Selenops spiders (Araneae: Selenopidae) from North and Central America: Implications for Caribbean biogeography. Biol. J. Linn. Soc. 2010, 101, 288–322. [Google Scholar] [CrossRef]
- Claramunt, S.; Derryberry, E.P.; Remsen, J.V., Jr.; Brumfield, R.T. High dispersal ability inhibits speciation in a continental radiation of passerine birds. Proc. Royal Soc. B 2012, 279, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Algar, A.C.; Mahler, D.L.; Glor, R.E.; Losos, J.B. Niche incumbency, dispersal limitation and climate shape geographical distributions in a species-rich island adaptive radiation. Glob. Ecol. Biogeogr. 2013, 22, 391–402. [Google Scholar] [CrossRef]
- Helmus, M.R.; Mahler, D.L.; Losos, J.B. Island biogeography of the Anthropocene. Nature 2014, 513, 543–546. [Google Scholar] [CrossRef]
- Dziki, A.; Binford, G.J.; Coddington, J.A.; Agnarsson, I. Spintharus flavidus in the Caribbean—A 30 million year biogeographical history and radiation of a ‘widespread species’. PeerJ 2015, 3, e1422. [Google Scholar] [CrossRef] [Green Version]
- Esposito, L.A.; Bloom, T.; Caicedo, L.; Alicia-Serrano, A.; Sanchez-Ruiz, J.; May-Collado, L.J.; Binford, G.; Agnarsson, I. Islands within islands: Diversification of tailless whip spiders (Amblypygi, Phrynus) in Caribbean caves. Mol. Phylogenet. Evol. 2015, 93, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Deler-Hernández, A.; Sýkora, V.; Seidel, M.; Cala-Riquelme, F.; Fikáček, M. Multiple origins of the Phaenonotum beetles in the Greater Antilles (Coleoptera: Hydrophilidae): Phylogeny, biogeography and systematics. Zool. J. Linnean Soc. 2017, 183, 97–120. [Google Scholar] [CrossRef]
- Reynolds, R.G.; Strickland, T.R.; Kolbe, J.J.; Falk, B.G.; Perry, G.; Revell, L.J.; Losos, J.B. Archipelagic genetics in a widespread Caribbean anole. J. Biogeogr. 2017, 44, 2631–2647. [Google Scholar] [CrossRef]
- Chamberland, L.; Salgado-Roa, F.C.; Basco, A.; Crastz-Flores, A.; Binford, G.J.; Agnarsson, I. Phylogeography of the widespread Caribbean spiny orb weaver Gasteracantha cancriformis. PeerJ 2020, 8, e8976. [Google Scholar] [CrossRef]
- Ali, J.R.; Hedges, S.B. Colonizing the Caribbean: New geological data and an updated land-vertebrate colonization record challenge the GAARlandia land-bridge hypothesis. J. Biogeogr. 2021, 48, 2699–2707. [Google Scholar] [CrossRef]
- Iturralde-Vinent, M.A. Aspectos geológicos de la biogeografía de Cuba. Cienc. Tierra Espacio 1982, 5, 85–100. [Google Scholar]
- Mann, P.; Schubert, C.; Burke, K. Review of Caribbean neotectonics. In The Caribbean Region: Geological Society of America; Dengo, G., Case, J.E., Eds.; The Geology of North America: Boulder, CO, USA, 1990; pp. 307–338. [Google Scholar]
- MacPhee, R.D.E.; Iturralde-Vinent, M.A. First Tertiary land mammal fossils from Greater Antilles: An Early Miocene sloth (Xenarhra, Megalonychidae) from Cuba. Am. Mus. Novit. 1994, 3094, 1–13. [Google Scholar]
- MacPhee, R.D.E.; Iturralde-Vinent, M.A. Origin of the Greater Antilles land mammal fauna 1: New Tertiary land mammals from Cuba and Puerto Rico. Am. Mus. Novit. 1995, 314, 1–31. [Google Scholar]
- Iturralde-Vinent, M.A.; MacPhee, R.D.E. Paleogeography of the Caribbean region: Implications for Cenozoic biogeography. Bull. Am. Mus. Natl. Hist. 1999, 238, 1–95. [Google Scholar]
- MacPhee, R.D.E.; Iturralde-Vinent, M.A. A short history of Greater Antillean land mammals: Biogeography, paleogeography, radiation, and extinctions. Tropics 2000, 10, 145–154. [Google Scholar] [CrossRef] [Green Version]
- White, J.L.; MacPhee, R.D.E. The sloths of the West Indies: A systematic and phylogenetic review. In Biogeography of the West Indies: Patterns and Perspectives; Woods, C.A., Sergile, F.E., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 201–235. [Google Scholar]
- Pindell, J.L.; Barrett, S.F. Geological evolution of the Caribbean region: A plate tectonic perspective. In The Caribbean Region: Geological Society of America; Dengo, G., Case, J.E., Eds.; The Geology of North America: Boulder, CO, USA, 1990; pp. 405–432. [Google Scholar]
- Van Gestel, J.P.; Mann, P.; Grindlay, N.R.; Dolan, J.F. Three-phase tectonic evolution of the northern margin of Puerto Rico as inferred from an integration of seismic reflection, well, and outcrop data. Mar. Geol. 1999, 161, 257–286. [Google Scholar] [CrossRef]
- MacPhee, R.D.E.; Iturralde-Vinent, M.A.; Gaffney, E.S. Domo de Zaza, an early Miocene vertebrate locality in south-central Cuba, with notes on the tectonic evolution of Puerto Rico and the Mona Passage. Am. Mus. Novit. 2003, 3394, 1–42. [Google Scholar] [CrossRef]
- Alonso, R.; Crawford, A.J.; Bermingham, E. Molecular phylogeny of an endemic radiation of Cuban toads (Bufonidae: Peltophryne) based on mitochondrial and nuclear genes. J. Biogeogr. 2012, 39, 434–451. [Google Scholar] [CrossRef]
- Rosen, D.E. A vicariance model of Caribbean biogeography. Syst. Zool. 1975, 24, 431–464. [Google Scholar] [CrossRef]
- Rosen, D.E. Geological hierarchies and biogeographical congruence in the Caribbean. Ann. Mo. Bot. Gard. 1985, 72, 636–659. [Google Scholar] [CrossRef] [Green Version]
- Tada, R.; Iturralde-Vinent, M.; Matsui, T.; Tajika, E.; Oji, T.; Goto, K.; Nakano, Y.; Takayama, H.; Yamamoto, S.; Toyoda, K.; et al. K/T Boundary deposits in the Paleo-western Caribbean basin. AAPG Mem. 2003, 26, 23. [Google Scholar]
- Hedges, S.B.; Hass, C.; Maxson, L. Caribbean biogeography: Molecular evidence for dispersal in West Indian terrestrial vertebrates. Proc. Natl. Acad. Sci. USA 1992, 89, 1909–1913. [Google Scholar] [CrossRef] [Green Version]
- Hedges, S.B. Biogeography of the West Indies: An overview. In Biogeography of the West Indies: Patterns and Perspectives; Woods, C.A., Sergile, F.E., Eds.; CRC Press: Baton Rouge, LA, USA, 2001; pp. 15–33. [Google Scholar]
- Hedges, S.B. Paleogeography of the Antilles and origin of West Indian terrestrial vertebrates. Ann. Missouri Bot. Gard. 2006, 93, 231–244. [Google Scholar] [CrossRef]
- Sato, J.J.; Ohdachi, S.D.; Echenique-Diaz, L.M.; Borroto-Páez, R.; Begué-Quiala, G.; Delgado-Labañino, L.J.; Gámez-Díez, J.; Álbarez-Lemus, J.; Nguyen, T.S.; Yamaguchi, N.; et al. Molecular phylogenetic analysis of nuclear genes suggests a Cenozoic overwater dispersal origin for the Cuban solenodon. Sci. Rep. 2016, 6, 31173. [Google Scholar] [CrossRef] [Green Version]
- Uit de Weerd, D.R.; Robinson, D.G.; Rosenberg, G. Evolutionary and biogeographical history of the land snail family Urocoptidae (Gastropoda: Pulmonata) across the Caribbean region. J. Biogeogr. 2016, 43, 763–777. [Google Scholar] [CrossRef]
- Yusseff-Vanegas, S.; Agnarsson, I. Molecular phylogeny of the forensically important genus Cochliomyia (Diptera: Calliphoridae). ZooKeys 2016, 609, 107–120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mchugh, A.; Yablonsky, C.; Binford, G.; Agnarsson, I. Molecular phylogenetics of Caribbean Micrathena (Araneae: Araneidae) suggests multiple colonization events and single island endemism. Invertebr. Syst. 2014, 28, 337–349. [Google Scholar] [CrossRef]
- Agnarsson, I.; LeQuier, S.M.; Kuntner, M.; Cheng, R.C.; Coddington, J.A.; Binford, G. Phylogeography of a good Caribbean disperser: Argiope argentata (Araneae, Araneidae) and a new ‘cryptic’ species from Cuba. ZooKeys 2016, 2016, 25–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čandek, K.; Agnarsson, I.; Binford, G.J.; Kuntner, M. Biogeography of the Caribbean Cyrtognatha spiders. Sci. Rep. 2019, 9, 397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Čandek, K.; Binford, G.J.; Agnarsson, I.; Kuntner, M. Caribbean golden orbweaving spiders maintain gene flow with North America. Zool. Scr. 2020, 49, 210–221. [Google Scholar] [CrossRef]
- Crews, S.C.; Esposito, L.A. Towards a synthesis of the Caribbean biogeography of terrestrial arthropods. BMC Evol. Biol. 2020, 20, 12. [Google Scholar] [CrossRef]
- Ali, J.R. Colonizing the Caribbean: Is the GAARlandia land-bridge hypothesis gaining a foothold? J. Biogeogr. 2012, 39, 431–433. [Google Scholar] [CrossRef]
- Philippon, M.; Cornée, J.-J.; Münch, P.; van Hinsbergen, D.J.J.; BouDagher-Fadel, M.; Gailler, L. Eocene intra-plate shortening responsible for the rise of a faunal pathway in the northeastern Caribbean realm. PLoS ONE 2020, 15, e0241000. [Google Scholar] [CrossRef]
- Murphy, W.J.; Collier, G.E. Phylogenetic relationships within the aplocheiloid fish genus Rivulus (Cyprinodontiformes, Rivulidae): Implications for Caribbean and Central American biogeography. Mol. Biol. Evol. 1996, 13, 642–649. [Google Scholar] [CrossRef] [Green Version]
- Dávalos, L.M. Short-faced bats (Phyllostomidae: Stenodermatina): A Caribbean radiation of strict frugivores. J. Biogeogr. 2007, 34, 364–375. [Google Scholar] [CrossRef]
- Santiago-Valentin, E.; Olmstead, R.G. Phylogenetics of the Antillean Goetzeoideae (Solanaceae) and their relationships within the Solanaceae based on chloroplast and ITS DNA sequence data. Am. Soc. Plant Taxon. 2003, 28, 452–460. [Google Scholar]
- Tong, Y.; Binford, G.; Rheims, A.R.; Kuntner, M.; Liu, J.; Agnarsson, I. Huntsmen of the Caribbean: Multiple tests of the GAARlandia hypothesis. Mol. Phylogenet. Evol. 2019, 130, 259–268. [Google Scholar] [CrossRef]
- Censky, E.J.; Hodge, K.; Dudley, J. Over-water dispersal of lizards due to hurricanes. Nature 1998, 395, 556. [Google Scholar] [CrossRef]
- Hall, J.P.W.; Robbins, R.K.; Harvey, D.J. Extinction and biogeography in the Caribbean: New evidence from a fossil riodinid butterfly in Dominican amber. Proc. Soc. R. Soc. B-Biol. Sci. 2004, 271, 797–801. [Google Scholar] [CrossRef]
- Wahlberg, N. That awkward age for butterflies: Insights from the age of the butterfly subfamily Nymphalinae (Lepidoptera: Nymphalidae). Syst. Biol. 2006, 55, 703–714. [Google Scholar] [CrossRef]
- Wahlberg, N.; Freitas, A.V. Colonization of and radiation in South America by butterflies in the subtribe Phyciodina (Lepidoptera: Nymphalidae). Mol. Phylogenet. Evol. 2007, 44, 1257–1272. [Google Scholar] [CrossRef]
- Sourakov, A.; Zakharov, E.V. “Darwin’s butterflies”? DNA barcoding and the radiation of the endemic Caribbean butterfly genus Calisto (Lepidoptera, Nymphalidae, Satyrinae). Comp. Cytogenet. 2011, 5, 191–210. [Google Scholar] [CrossRef] [Green Version]
- Ceccarelli, F.S.; Zaldívar-Riverón, A. Broad polyphyly and historical biogeography of the Neotropical wasp genus Notiospathius (Braconidae: Doryctinae). Mol. Phylogenet. Evol. 2013, 69, 142–152. [Google Scholar] [CrossRef]
- Lewis, D.S.; Sperling, F.A.H.; Nakahara, S.; Cotton, A.M.; Kawahara, A.Y.; Condamine, F.L. Role of Caribbean Islands in the diversification and biogeography of Neotropical Heraclides swallowtails. Cladistics 2015, 31, 291–314. [Google Scholar] [CrossRef]
- Rodriguez, J.; Pitts, J.P.; von Dohlen, C.D. Historical biogeography of the widespread spider wasp tribe Aporini (Hymenoptera: Pompilidae). J. Biogeogr. 2015, 42, 495–506. [Google Scholar] [CrossRef]
- Zhang, G.; Basharat, U.; Matzke, N.; Franz, N.M. Model selection in statistical historical biogeography of Neotropical insects—the Exophthalmus genus complex (Curculionidae: Entiminae). Mol. Phylogenet. Evol. 2017, 109, 226–239. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.X.; Maddison, W.P. Molecular phylogeny, divergence times and biogeography of spiders of the subfamily Euophryinae (Araneae: Salticidae). Mol. Phylogenet. Evol. 2013, 68, 81–92. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarty, P. Systematics and historical biogeography of Greater Antillean Cichlidae. Mol. Phylogenet. Evol. 2006, 39, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Říčan, O.; Piálek, L.; Zardoya, R.; Doadrio, I.; Zrzavý, J. Biogeography of the Mesoamerican Cichlidae (Teleostei: Heroini): Colonization through the GAARlandia land bridge and early diversification. J. Biogeogr. 2013, 40, 579–593. [Google Scholar] [CrossRef] [Green Version]
- World Spider Catalog. World Spider Catalog. Version 22.5. Natural History Museum Bern. Available online: http://wsc.nmbe.ch (accessed on 17 November 2021).
- Spagna, J.C.; Peattie, A.M. Terrestrial locomotion in arachnids. J. Insect Physiol. 2012, 58, 599–606. [Google Scholar] [CrossRef]
- Kropf, C. Hydraulic system of locomotion. In Spider Ecophysiology; Nentwig, W., Ed.; Springer: Berlin, Germany, 2013; pp. 43–56. [Google Scholar]
- Brandt, E.E.; Roberts, K.T.; Williams, C.M.; Elias, D.O. Low temperatures impact species distributions of jumping spiders across a desert elevational cline. J. Insect. Physiol. 2020, 122, 104037. [Google Scholar] [CrossRef] [Green Version]
- Bodner, M.R.; Maddison, W.P. The biogeography and age of salticid spider radiations (Araneae: Salticidae). Mol. Phylogenet. Evol. 2012, 65, 213–240. [Google Scholar] [CrossRef]
- Maddison, W.P.; Hedin, M.C. Jumping spider phylogeny (Araneae: Salticidae). Invertebr. Syst. 2003, 17, 529–549. [Google Scholar] [CrossRef] [Green Version]
- Maddison, W.P.; Bodner, M.R.; Needham, K.M. Salticid spider phylogeny revisited, with the discovery of a large Australasian clade (Araneae: Salticidae). Zootaxa 2008, 1893, 46–64. [Google Scholar] [CrossRef] [Green Version]
- Bryant, E.B. The salticid spiders of Hispaniola. Bull. Mus. Comp. Zool. 1943, 92, 445–529. [Google Scholar]
- Zhang, J.X.; Maddison, W.P. New euophryine jumping spiders from the Dominican Republic and Puerto Rico (Araneae: Salticidae: Euophryinae). Zootaxa. 2012, 3476, 1–54. [Google Scholar] [CrossRef]
- Prószyński, J.; Deeleman-Reinhold, C.L. Description of some Salticidae (Aranei) from the Malay Archipelago. II. Salticidae of Java and Sumatra, with comments on related species. Arthropoda Sel. 2012, 21, 29–60. [Google Scholar] [CrossRef]
- Zhang, J.X.; Maddison, W.P. Genera of euophryine jumping spiders (Araneae: Salticidae), with a combined molecular-morphological phylogeny. Zootaxa 2015, 3938, 1–147. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Bio. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Hedin, M.C.; Maddison, W.P. A combined molecular approach to phylogeny of the jumping spider subfamily Dendryphantinae (Araneae: Salticidae). Mol. Phylogenet. Evol. 2001, 18, 386–403. [Google Scholar] [CrossRef] [Green Version]
- Green, P. Phrap, Version 0.990329. 1999. Available online: http://www.phrap.org (accessed on 2 June 2021).
- Green, P.; Ewing, B. Phred, Version 0.020425 c. 2002. Available online: http://www.phrap.org (accessed on 2 June 2021).
- Maddison, D.R.; Maddison, W.P. 2021. Chromaseq: A Mesquite Package for Analyzing Sequence Chromatograms. Version 1.31. 2018. Available online: http://chromaseq.mesquiteproject.org (accessed on 2 June 2021).
- Maddison, W.P.; Maddison, D.R. Mesquite: A Modular System for Evolutionary Analysis. Version 3.6. 2018. Available online: http://www.mesquiteproject.org (accessed on 2 June 2021).
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [Green Version]
- Posada, D.; Buckley, T.R. Model selection and model averaging in phylogenetics: Advantages of Akaike Information Criterion and Bayesian Approaches over Likelihood Ratio Tests. Syst. Biol. 2004, 53, 793–808. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.F.K.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast model selection for accurate phylogenetic estimates. Nat. Methods 2015, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Altekar, G.; Dwarkadas, S.; Huelsenbeck, J.P.; Ronquist, F. Parallel Metropolis coupled Markov chain Monte Carlo for Bayesian phylogenetic inference. Bioinformatics 2004, 20, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for inference of large phylogenetic tres. In Proceedings of the 2010 Gateway Computing Environments Workshop (GCE 2010), New Orleans, LA, USA, 14 November 2010; pp. 1–8. [Google Scholar] [CrossRef] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Suchard, M.A.; Xie, D.; Drummond, A.J. Tracer v1.6. 2014. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 2 June 2021).
- Drummond, A.J.; Suchard, M.A.; Xie, D.; Rambaut, A. Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol. Biol. Evol. 2012, 29, 1969–1973. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistuzzi, F.U.; Billing-Ross, P.; Paliwal, A.; Kumar, S. Fast and slow implementations of relaxed-clock methods show similar patterns of accuracy in estimating divergence times. Mol. Biol. Evol. 2011, 28, 2439–2442. [Google Scholar] [CrossRef] [PubMed]
- Bidegaray-Batista, L.; Arnedo, M.A. Gone with the plate: The opening of the Western Mediterranean basin drove the diversification of ground-dweller spiders. MBC Evol. Biol. 2011, 11, 317. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Lewis, P.O.; Fan, Y.; Kuo, L.; Chen, M.H. Improving marginal likelihood estimation for Bayesian phylogenetic model selection. Syst. Biol. 2011, 60, 150–160. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Gernhard, T. The conditioned reconstructed process. J. Theor. Biol. 2008, 253, 769–778. [Google Scholar] [CrossRef] [Green Version]
- Stadler, T. On incomplete sampling under birth–death models and connections to the sampling-based coalescent. J. Theor. Biol. 2009, 261, 58–66. [Google Scholar] [CrossRef]
- Wunderlich, J. Die Fossilen Spinnen im Dominikanischen Bernstein; Published by the author Straubenhardt: Baden-Württemberg, Germany, 1988. [Google Scholar]
- Penney, D. Dominican Amber Spiders: A Comparative Palaeontological-Neontological Approach to Identification, Faunistics, Ecology and Biogeography; Siri Scientific Press: Rochdale, UK, 2008. [Google Scholar]
- Peckham, G.W.; Peckham, E.G. On some new genera and species of Attidae from the eastern part of Guatemala. Proc. Nat. Hist. Soc. Wisconsin. 1885, 2, 62–86. [Google Scholar]
- Zhang, J. Phylogeny and Systematics of the Jumping Spider Subfamily Euophryinae (Araneae: Salticidae), with Consideration of Biogeography and Genitalic Evolution. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2012. Available online: https://open.library.ubc.ca/collections/ubctheses/24/items/1.0072804 (accessed on 17 November 2021).
- Matzke, N.J. Model selection in historical biogeography reveals that founder-event speciation is a crucial process in Island Clades. Syst. Biol. 2014, 63, 951–970. [Google Scholar] [CrossRef]
- Matzke, N.J. BioGeoBEARS: BioGeography with Bayesian (and Likelihood) Evolutionary Analysis with R Scripts. Version 1.1.1. 2018. Available online: https://doi.org/10.5281/zenodo.1478250 (accessed on 4 September 2021).
- Ree, R.H.; Smith, S.A. Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst. Biol. 2008, 57, 4–14. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F. Dispersal-vicariance analysis: A new approach to the quantification of historical biogeography. Syst. Biol. 1997, 46, 195–203. [Google Scholar] [CrossRef]
- Landis, M.J.; Matzke, N.J.; Moore, B.R.; Huelsenbeck, J.P. Bayesian analysis of biogeography when the number of áreas is large. Syst. Biol. 2013, 62, 789–804. [Google Scholar] [CrossRef] [Green Version]
- Klaus, K.V.; Matzke, N.J. Statistical Comparison of Trait-Dependent Biogeographical Models Indicates That Podocarpaceae Dispersal Is Influenced by Both Seed Cone Traits and Geographical Distance. Syst. Biol. 2020, 69, 61–75. [Google Scholar] [CrossRef]
- Ree, R.H.; Sanmartín, I. Conceptual and statistical problems with the DEC+J model of founder-event speciation and its comparison with DEC via model selection. J. Biogeogr. 2018, 45, 741–749. [Google Scholar] [CrossRef]
- Matzke, N.J. Statistical comparison of DEC and DEC+J is identical to comparison of two ClaSSE submodels, and is therefore valid. OSF Preprint 2021, 1–40. [Google Scholar] [CrossRef]
- Matzke, N.J. Probabilistic historical biogeography: New models for founder-event speciation, imperfect detection, and fossils allow improved accuracy and model-testing. Front. Biogeogr. 2013, 5, 242–248. [Google Scholar] [CrossRef] [Green Version]
- Matzke, N.J. Stochastic Mapping under Biogeographical Models. PhyloWiki BioGeoBEARS Website. 2016. Available online: http://phylo.wikidot.com/biogeobears#stochastic_mapping (accessed on 27 September 2021).
- Magalhaes, I.L.F.; Santos, A.J.; Ramírez, M.J. Incorporating topological and age uncertainty into event-based biogeography supports paleo-islands in Galapagos and ancient connections among Neotropical dry forests. Diversity 2021, 13, 418. [Google Scholar] [CrossRef]
- Shapiro, L.; Binford, G.J.; Agnarsson, I. Single-Island Endemism despite Repeated Dispersal in Caribbean Micrathena (Araneae: Araneidae): An Updated Phylogeographic Analysis. Diversity 2022, 14, 128. [Google Scholar] [CrossRef]
- Loureiro, L.O.; Engstrom, M.D.; Lim, B.K. Optimization of Genotype by Sequencing data for phylogenetic purposes. MethodsX 2020, 7, 100892. [Google Scholar] [CrossRef] [PubMed]
- Hedges, S.B.; Woods, C.A. Caribbean hot spot. Nature 1993, 364, 375. [Google Scholar] [CrossRef]
- Woods, C.; Borroto, P.R.; Kilpatrick, C. Insular patterns and radiation of West Indian rodents. In Biogeography of the West Indies: Patterns and Perspectives; Woods, C.A., Sergile, F.E., Eds.; CRC Press: Boca Raton, FL, USA, 2001; pp. 333–351. [Google Scholar]
- Fabre, P.H.; Vilstrup, J.T.; Raghavan, M.; Der Sarkissian, C.; Willerslev, E.; Douzery, E.J.P.; Orlando, L. Rodents of the Caribbean: Origin and diversification of hutias unravelled by next-generation museomics. Biol. Lett. 2014, 10, 20140266. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | CO1 | 16S-ND1 | 28S |
|---|---|---|---|---|---|
| Agobardus anormalis montanus | JXZ357 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615636 | KC615802 | KC615376 |
| Agobardus bahoruco | JXZ324 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615844 | KC615417 | |
| Agobardus cf. brevitarsus | JXZ311 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615637 | KC615803 | KC615637 |
| Agobardus cordiformis | JXZ358 | DOMINICAN REPUBLIC: Pedernales, (N17.965, W71.635) | KC615634 | KC615800 | KC615374 |
| Agobardus gramineus | JXZ314 | DOMINICAN REPUBLIC: Pedernales, (N17.965, W71.635) | KC615635 | KC615801 | KC615375 |
| Agobardus oviedo | JXZ312 | DOMINICAN REPUBLIC: Pedernales, (N17.802, W71.349) | KC615638 | KC615804 | KC615378 |
| Antillattus [Cuba1] | CU787945 CU00107A | CUBA: Granma, Bartolomé Maso, (N20.009, W76.894) | √ | √ | √ |
| Antillattus [Cuba2] | CU00025A CU00086A CU00090A CU00004A CU00016A | CUBA: Granma, Bartolomé Maso, (N20.013,W76.834) | √ | √ | |
| Antillattus [Cuba3] | CU787957 CU03506A | CUBA: Pinar del Rio, Viñales, (N22.657, W83.701) | √ | ||
| Antillattus [Cuba4] | CU00100A CU03361A CU03317A | CUBA: Guantánamo, Baracoa, (N20.331, W74.569) | √ | ||
| Antillattus [Cuba4] | CU03121A | CUBA: Guantánamo, Nibujón, (N20.052, W76.502) | √ | ||
| Antillattus cambridgei | JXZ321 | DOMINICAN REPUBLIC: La Vega, (N19.033, W70.543) | KC615646 | KC615818 | KC615392 |
| Antillattus cambridgei | DR784676 DR785410 DR785798 DR782454 | DOMINICAN REPUBLIC: La Alta Gracia, (N19.067, W69.463) | √ | √ | |
| Antillattus cambridgei | DR785494 DR782541 | DOMINICAN REPUBLIC: La Alta Gracia, (N19.893, W71.653) | √ | √ | |
| Antillattus cambridgei | DR782541 DR785783 DR785508 | DOMINICAN REPUBLIC: La Alta Gracia, (N19.355, W070.111) | √ | √ | |
| Antillattus cambridgei | DR782598 | DOMINICAN REPUBLIC: La Alta Gracia, (19.355N, W70.111) | √ | √ | √ |
| Antillattus cambridgei | DR784852 DR785098 DR785696 DR785438 DR787296 | DOMINICAN REPUBLIC: La Alta Gracia, (19.067, W69.463) | √ | √ | |
| Antillattus cambridgei | DR787296 DR787293 DR787254 DR787223 | DOMINICAN REPUBLIC: La Vega, (N19.036, W70.543) | √ | √ | |
| Antillattus cambridgei | DR787328 DR787252 DR787285 DR787207 DR787327 DR787319 DR787324 | DOMINICAN REPUBLIC: Santo Domingo, (19.051 N, W70.888) | √ | √ | |
| Antillattus cambridgei | DR787105 | DOMINICAN REPUBLIC: San Juan, (N19.175, W71.049) | √ | √ | |
| Antillattus cf. applanatus | JXZ336 | DOMINICAN REPUBLIC: Barahona, Cachote (N18.101, W71.194) | KC615699 | KC615911 | KC615699 |
| Antillattus cubensis | CU003076 CU002975 CU003097 CU003360 CU002456 CU003486 CU02560A CU02975A CU03033A CU03076A CU03097A CU03360A | CUBA: Cienfuegos, Soledad, (N22.124, W80.325) | √ | ||
| Antillattus cubensis | CU03417A CU03488A | CUBA: Santiago de Cuba, San Luis, (N20.179, W75.783) | √ | ||
| Antillattus cubensis | CU3075A | CUBA: Santiago de Cuba, (N20.010, W76.037) | √ | ||
| Antillattus cubensis | CU02583A | CUBA: Guantánamo, Baracoa, (N20.331, W74.569) | √ | ||
| Antillattus cubensis | CU787598 CU783280 CU787621 CU787283 CU787277 | CUBA: Granma, Bartolomé Maso, (N20.009, W76.894) | √ | ||
| Antillattus darlingtoni | JXZ341 | DOMINICAN REPUBLIC: La Vega, Ébano Verde, (N19.033,W70.543) | KC615762 | KC616005 | KC615583 |
| Antillattus darlingtoni | DR787120 | DOMINICAN REPUBLIC: San Juan, Pico Duarte | √ | √ | |
| Antillattus darlingtoni | DR786937 DR784873 | DOMINICAN REPUBLIC: Valle nuevo | √ | √ | |
| Antillattus darlingtoni | DR784828 DR784873 | DOMINICAN REPUBLIC: La Vega, Ébano Verde, (N19.026, W19.0264) | √ | √ | |
| Antillattus gracilis | JXZ320 | DOMINICAN REPUBLIC: La Vega, P.N.Armando Bermúdez, (N19.06, W70.86) | KC615817 | KC615391 | |
| Antillattus gracilis | DR782845 DR787278 | DOMINICAN REPUBLIC: Santo Domingo, Los Tablones (N19.051, W70.888) | √ | √ | |
| Antillattus keyserlingi | CU03135A | CUBA: Holguin, Frank Pais, (N20.529N, N75.768) | √ | ||
| Antillattus keyserlingi | CU02571A | CUBA: Santiago de Cuba, Gran Piedra, (N20.011, W75.623) | √ | ||
| Antillattus keyserlingi | CU787312 | CUBA: Guantánamo, Baracoa, (20.331N, W74.569) | √ | ||
| Antillattus keyserlingi | CU00081A CU00088A CU02951A CU02985A CU03043A CU782822 CU783187 CU783232 CU783245 CU783281 CU783404 CU783425 CU787302 CU787433 CU787625 | CUBA: Granma, Bartolomé Maso, (N20.052, W76.502) | √ | √ | √ |
| Antillattus keyserlingi | CU02467A CU03538A CU03395A | CUBA: Holguin, Frank Pais,(N20.529, W75.768) | √ | √ | √ |
| Antillattus keyserlingi | CU03036A CU03274A | CUBA: Granma, Bartolomé Maso, (N20.015–W76.839) | √ | ||
| Antillattus maxillosus | JXZ335 | DOMINICAN REPUBLIC: La Vega, road Constanza to Ocoa, Valle Nuevo (N18.700, W70.606) | KC615708 | KC615935 | KC615510 |
| Antillattus maxillosus | DR786952 DR786992 DR786981 | DOMINICAN REPUBLIC: Valle nuevo, Villa Pajón (N18.82208, W070.6838) | √ | √ | |
| Antillattus [Cuba5] | CU03373A CU03396A CU03539A CU03534A | CUBA: Pinar del Rio, Viñales, (N22.653, W83.699) | √ | ||
| Antillattus placidus | DR787249 | DOMINICAN REPUBLIC: La Vega, Jarabacoa, (N19.036, W70.543) | √ | √ | |
| Antillattus placidus | DR782502 DR785683 DR785081 | DOMINICAN REPUBLIC: La Alta Gracia, Yuma, (N19.355, W70.111) | √ | √ | |
| Antillattus scutiformis | JXZ326 | DOMINICAN REPUBLIC: La Vega, road Constanza to Ocoa, Valle Nuevo (N18.848, W70.720) | KC615860 | KC615433 | |
| Bythocrotus cf. crypticus | JXZ323 | DOMINICAN REPUBLIC: El Seibo, Pedro Sanchez, (N18.86, W69.11) | KC615661 | KC615839 | KC615412 |
| Bythocrotus crypticus | JXZ322 | DOMINICAN REPUBLIC: Barahona, (N18.424, W71.112) | KC615660 | KC615838 | KC615411 |
| Cobanus cambridgei | JXZ122 | COSTA RICA: Prov. San José, (N9.65, W83.97) | KC615872 | KC615445 | |
| Cobanus extensus | JXZ122 | ECUADOR: Pichincha, near El Cisne, (N0.1493, W79.0317) | KC615872 | KC615445 | |
| Cobanus mandibularis | JXZ245 | PANAMA: Panamá: Gamboa, Pipeline Road, (N9.15840, W79.74252) | KC615876 | KC615449 | |
| Cobanus unicolor | JXZ244 | PANAMA: Chiriqui: Fortuna, Quebrada Samudio, (N8.73464, W82.24839) | KC615878 | KC615451 | |
| Compsodecta festiva | JAM4122A | JAMAICA: Portland, Millbank, (N18.013, W76.379) | √ | ||
| Compsodecta haytiensis | JXZ325 | DOMINICAN REPUBLIC: Barahona, Highway 44 south of Barahona (N18.138, W71.070) | KC615671 | KC615859 | KC615432 |
| Compsodecta peckhami | JXZ327 | DOMINICAN REPUBLIC: Pedernales, Rio Mulito (N18.155, W71.758) | KC615884 | KC615457 | |
| Corticattus guajataca | JXZ305 | PUERTO RICO: Isabela: Bosque de Guajataca (N18.421, W66.966) | KC615715 | KC615945 | KC615521 |
| Corticattus latus | JXZ337 | DOMINICAN REPUBLIC: Pedernales: Laguna de Oviedo (N17.802 W71.349) | KC615698 | KC615908 | KC615483 |
| Mexigonus arizonenzis | JXZ163 | USA: Arizona: Yavapai Co., Iron Springs (N34.58476, W112.57071) | KC615747 | KC615988 | KC615564 |
| Mexigonus cf. minuta | d117 | ECUADOR: Pichincha: Quito | √ | √ | √ |
| Mexigonus morosus | JXZ362 | USA: California: San Mate Co.,(N37.434, W122.311) | KC615990 | KC615566 | |
| Pensacola signata | JXZ371 | GUATEMALA: Depto. Petén, Reserva Natural Ixpanpajul | KC616006 | KC615584 | |
| Petemathis portoricensis | PR782206 | PUERTO RICO: Villalba: Toro negro, El Bolo Trail (N18.1777401, W66.488319) | √ | √ | |
| Petemathis portoricensis [Adjuntas] | JXZ306 | PUERTO RICO: Adjuntas, HWY143 to Cerro Punta (N18.167, W66.576) | KC615716 | KC615946 | KC615522 |
| Petemathis portoricensis [Maricao] | JXZ303 | PUERTO RICO: Maricao, Bosque de Maricao (N18.150, W66.994) | KC615711 | KC615940 | KC615515 |
| Petemathis tetuani | JXZ303 | PUERTO RICO: Maricao, Bosque de Maricao (N18.150, W66.994) | KC615711 | KC615940 | KC615515 |
| Petemathis tetuani | PR782277 | PUERTO RICO: Villalba: Toro negro, El Bolo Trail, (N18.177, W66.488) | √ | √ | |
| Petemathis tetuani | PR392859 | PUERTO RICO: Rio Grande, El Yunque, Mt. Britton, (N18.2957, W65.7906) | √ | √ | |
| Popcornella furcata | JXZ334 | DOMINICAN REPUBLIC: La Vega, Reserva Científica Ébano Verde, (N19.04, W70.518) | KC615714 | KC615944 | KC615520 |
| Popcornella spiniformis | JXZ339 | DOMINICAN REPUBLIC: Barahona, Cachote (N18.098, W71.187) | KC615914 | KC615489 | |
| Popcornella yunque | JXZ309 | PUERTO RICO: Río Grande, El Yunque Nat. Forest, (N18.3174, W65.8314) | KC615937 | KC615512 | |
| Sidusa [French guiana1] | JXZ128 | FRENCH GUIANA: Commune Règina, les Nourages Field Station (N4.069, W52.669) | KC615770 | KC616015 | KC615593 |
| Sidusa [French guiana2] | JXZ100 | FRENCH GUIANA: Commune Règina, les Nourages Field Station, (N4.069, W52.669) | KC615679 | KC615871 | KC615444 |
| Truncattus [Cuba1] | CU3492A | CUBA: Granma, Bartolomé Maso, National Park Pico Turquino (N 20.0526, W76.502) | √ | ||
| Truncattus [Cuba2] | CU787947 CU03405A | CUBA: Granma, Bartolomé Maso, National Park Pico Turquino (N20.0526, W76.5029) | √ | √ | √ |
| Truncattus [Cuba3] | CU787949 CU00083A CU03065A | CUBA: Granma, Bartolomé Maso, National Park Pico Turquino (N20.0526, W76.5029) | √ | ||
| Truncattus [Cuba4] | CU00014A | CUBA: Granma, Bartolomé Maso, National Park Pico Turquino (N20.052, W76.502) | √ | ||
| Truncattus [Dominican Republic1] | DR787029 | DOMINICAN REPUBLIC: Valle nuevo, Villa Pajón, (N18.82208, W070.6838) | √ | ||
| Truncattus cachotensis | JXZ338 | DOMINICAN REPUBLIC: Barahona, Cachote, (N18.101, W71.194) | KC615701 | KC615913 | KC615488 |
| Truncattus dominicanus | JXZ340 | DOMINICAN REPUBLIC: La Vega, P.N.Armando Bermúdez,(N19.06, W70.86) | KC615703 | KC615920 | KC615495 |
| Truncattus dominicanus | DR787325 | DOMINICAN REPUBLIC: San Juan, Los tablones,(N19.0511, W70.888) | √ | √ | |
| Truncattus flavus | JXZ332 | DOMINICAN REPUBLIC: La Vega, P.N.Armando Bermúdez, (N19.06, W70.86) | KC615707 | KC615933 | KC615508 |
| Outgroups | |||||
| Agobardus anormalis montanus | JXZ357 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615636 | KC615802 | KC615376 |
| Agobardus bahoruco | JXZ324 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615844 | KC615417 | |
| Agobardus cf. brevitarsus | JXZ311 | DOMINICAN REPUBLIC: Pedernales, (N18.128, W71.558) | KC615637 | KC615803 | KC615637 |
| Agobardus cordiformis | JXZ358 | DOMINICAN REPUBLIC: Pedernales, (N17.965, W71.635) | KC615634 | KC615800 | KC615374 |
| Agobardus gramineus | JXZ314 | DOMINICAN REPUBLIC: Pedernales, (N17.965, W71.635) | KC615635 | KC615801 | KC615375 |
| Agobardus oviedo | JXZ312 | DOMINICAN REPUBLIC: Pedernales, (N17.802, W71.349) | KC615638 | KC615804 | KC615378 |
| Bythocrotus cf. crypticus | JXZ323 | DOMINICAN REPUBLIC: El Seibo, Pedro Sanchez, (N18.86, W69.11) | KC615661 | KC615839 | KC615412 |
| Bythocrotus crypticus | JXZ322 | DOMINICAN REPUBLIC: Barahona, (N18.424, W71.112) | KC615660 | KC615838 | KC615411 |
| Cobanus cambridgei | JXZ122 | COSTA RICA: Prov. San José, (N9.65, W83.97) | KC615872 | KC615445 | |
| Cobanus extensus | JXZ122 | ECUADOR: Pichincha, near El Cisne, (N0.1493, W79.0317) | KC615872 | KC615445 | |
| Cobanus mandibularis | JXZ245 | PANAMA: Panamá: Gamboa, Pipeline Road, (N9.15840, W79.74252) | KC615876 | KC615449 | |
| Cobanus unicolor | JXZ244 | PANAMA: Chiriqui: Fortuna, Quebrada Samudio, (N8.73464, W82.24839) | KC615878 | KC615451 | |
| Compsodecta festiva | JAM4122A | JAMAICA: Portland, Millbank, (N18.013, W76.379) | √ | ||
| Compsodecta haytiensis | JXZ325 | DOMINICAN REPUBLIC: Barahona, Highway 44 south of Barahona (N18.138, W71.070) | KC615671 | KC615859 | KC615432 |
| Compsodecta peckhami | JXZ327 | DOMINICAN REPUBLIC: Pedernales, Rio Mulito (N18.155, W71.758) | KC615884 | KC615457 | |
| Corticattus guajataca | JXZ305 | PUERTO RICO: Isabela: Bosque de Guajataca (N18.421, W66.966) | KC615715 | KC615945 | KC615521 |
| Corticattus latus | JXZ337 | DOMINICAN REPUBLIC: Pedernales: Laguna de Oviedo (N17.802, W71.349) | KC615698 | KC615908 | KC615483 |
| Mexigonus arizonenzis | JXZ163 | USA: Arizona: Yavapai Co., Iron Springs (N34.58476, W112.57071) | KC615747 | KC615988 | KC615564 |
| Mexigonus cf. minuta | d117 | ECUADOR: Pichincha: Quito | KC615748 | KC615989 | KC615565 |
| Mexigonus morosus | JXZ362 | USA: California: San Mate Co.,(N37.434, W122.311) | KC615990 | KC615566 | |
| Pensacola signata | JXZ371 | GUATEMALA: Depto. Petén, Reserva Natural Ixpanpajul | KC616006 | KC615584 | |
| Popcornella furcata | JXZ334 | DOMINICAN REPUBLIC: La Vega, Reserva Científica Ébano Verde, (N19.04, W70.518) | KC615714 | KC615944 | KC615520 |
| Popcornella spiniformis | JXZ339 | DOMINICAN REPUBLIC: Barahona, Cachote (N18.098, W71.187) | KC615914 | KC615489 | |
| Popcornella yunque | JXZ309 | PUERTO RICO: Río Grande, El Yunque Nat. Forest, (N18.3174, W65.8314) | KC615937 | KC615512 | |
| Sidusa [French guiana1] | JXZ128 | FRENCH GUIANA: Commune Règina, les Nourages Field Station (N4.069, W52.669) | KC615770 | KC616015 | KC615593 |
| Sidusa [French guiana2] | JXZ100 | FRENCH GUIANA: Commune Règina, les Nourages Field Station, (N4.069, W52.669) | KC615679 | KC615871 | KC615444 |
| Ghelna canadensis | d005 | USA: North Carolina (N35.704, W82.373) | EF201651 | JQ312080 | KT462689 |
| (1) Non-GAARlandia/(2) GAARlandia | |||
| A: Dispersal probability decreases as distance increases | B: Distance does not affect dispersal probability | C: Probability of overwater dispersal is very low | |
| (A) GA1 | A1a/A2a | A1b/A2b | A1c/A2c |
| (1) Non–land connections/(2) Land connections | |||
| A: Dispersal probability decreases as distance increases | B: Distance does not affect dispersal probability | C: Probability of overwater dispersal is very low | |
| (A) MO | A1a/A2a | A1b/A2b | A1c/A2c |
| (B) MO+WI | B1a/B2a | B1b/B2b | B1c/B2c |
| Time–Constrained/GAARlandia (A2a) | |||||||
| LnL | n par | d | e | j | AIC | AICc | |
| DEC | −43.84 * | 2 | 0.021 | <0.0001 | 0 | 91.69 | 92.01 |
| DEC +J | −33.87 * | 3 | <0.0001 | <0.0001 | 0.22 | 73.73 | 74.4 |
| DIVALIKE | −47.05 | 2 | 0.035 | <0.0001 | 0 | 98.1 | 98.43 |
| DIVALIKE +J | −36 | 3 | 0.005 | <0.0001 | 0.22 | 77.99 | 78.66 |
| BAYAREALIKE | −60.83 | 2 | 0.021 | 0.031 | 0 | 125.7 | 126 |
| BAYAREALIKE +J | −36.73 | 3 | 0.0038 | <0.0001 | 0.21 | 79.45 | 80.12 |
| Time–constrained/land connections prior to the Mona Passage and the Windward Passage (B2a) | |||||||
| DEC | −25.1 | 2 | 0.033 | <0.0001 | 0 | 54.19 | 54.61 |
| DEC +J | −18.09 | 3 | <0.0001 | <0.0001 | 0.31 | 42.18 | 43.04 |
| DIVALIKE | −25.01 * | 2 | 0.048 | <0.0001 | 0 | 54.01 | 54.43 |
| DIVALIKE +J | −17.62 * | 3 | <0.0001 | <0.0001 | 0.29 | 41.25 | 42.1 |
| BAYAREALIKE | −34.22 | 2 | <0.0001 | 0.041 | 0 | 72.44 | 72.86 |
| BAYAREALIKE +J | −18.8 | 3 | <0.0001 | <0.0001 | 0.27 | 43.61 | 44.46 |
| DIVALIKE | ||||||||||||
| j | a | d | e | s | v | y | Ÿd | Ad | Ÿa | Ÿc | Total events | |
| means | 0 | 0 | 5.5 | 0 | 0 | 6.61 | 24.39 | 5.5 | 5.5 | 5.5 | 31 | 36.5 |
| stdevs | 0 | 0 | 0.64 | 0 | 0 | 0.67 | 0.67 | 0.64 | 0.64 | 0.64 | 0 | 0.64 |
| sums | 0 | 0 | 550 | 0 | 0 | 661 | 2439 | 550 | 550 | 550 | 3100 | 3650 |
| DIVALIKE+j | ||||||||||||
| j | a | d | e | s | v | y | Ÿd | Ad | Ÿa | Ÿc | Total events | |
| means | 6.63 | 0 | 0 | 0 | 0 | 0.82 | 23.55 | 6.63 | 0 | 0 | 31 | 31 |
| stdevs | 1.04 | 0 | 0 | 0 | 0 | 0.64 | 0.94 | 1.04 | 0 | 0 | 0 | 0 |
| sums | 663 | 0 | 0 | 0 | 0 | 82 | 2355 | 663 | 0 | 0 | 3100 | 3100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cala-Riquelme, F.; Wiencek, P.; Florez-Daza, E.; Binford, G.J.; Agnarsson, I. Island–to–Island Vicariance, Founder–Events and within–Area Speciation: The Biogeographic History of the Antillattus Clade (Salticidae: Euophryini). Diversity 2022, 14, 224. https://doi.org/10.3390/d14030224
Cala-Riquelme F, Wiencek P, Florez-Daza E, Binford GJ, Agnarsson I. Island–to–Island Vicariance, Founder–Events and within–Area Speciation: The Biogeographic History of the Antillattus Clade (Salticidae: Euophryini). Diversity. 2022; 14(3):224. https://doi.org/10.3390/d14030224
Chicago/Turabian StyleCala-Riquelme, Franklyn, Patrick Wiencek, Eduardo Florez-Daza, Greta J. Binford, and Ingi Agnarsson. 2022. "Island–to–Island Vicariance, Founder–Events and within–Area Speciation: The Biogeographic History of the Antillattus Clade (Salticidae: Euophryini)" Diversity 14, no. 3: 224. https://doi.org/10.3390/d14030224
APA StyleCala-Riquelme, F., Wiencek, P., Florez-Daza, E., Binford, G. J., & Agnarsson, I. (2022). Island–to–Island Vicariance, Founder–Events and within–Area Speciation: The Biogeographic History of the Antillattus Clade (Salticidae: Euophryini). Diversity, 14(3), 224. https://doi.org/10.3390/d14030224