
Distinct Reproductive Strategy of Two Endemic Amazonian Quillworts

 , , , ,
, , , , 
 , , , , and
, , , , and 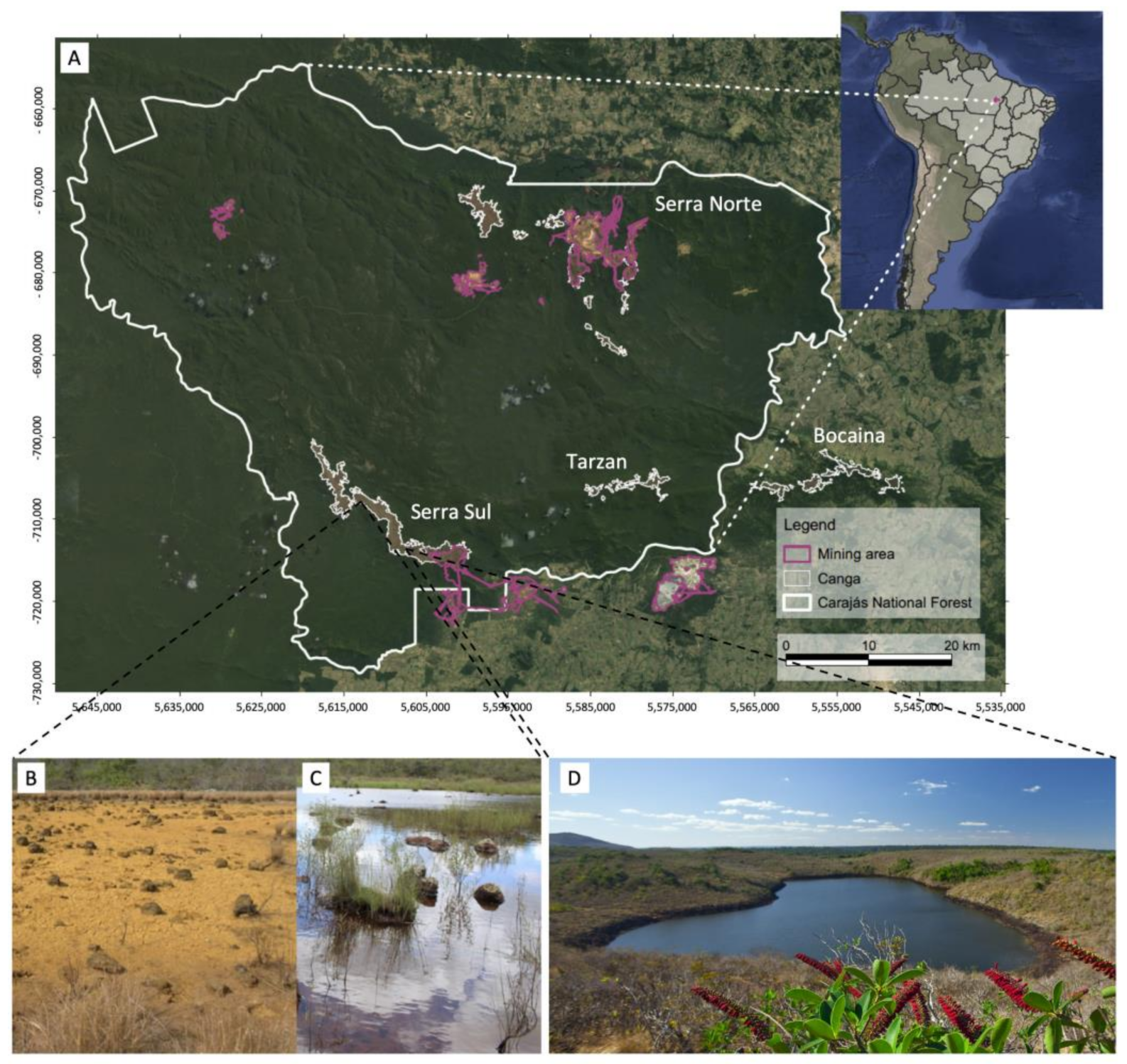{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Spore Germination from Sporangia Collected during the Dry and Rainy Seasons
2.3. Sexual Propagation Assays
Nuclear DNA Extraction and PCR Amplification of ISSR Markers
2.4. Isoëtes cangae Ex Situ Growth and Sporangia Production
2.5. Vegetative or Asexual Propagation Assays
2.6. Data Analysis
3. Results
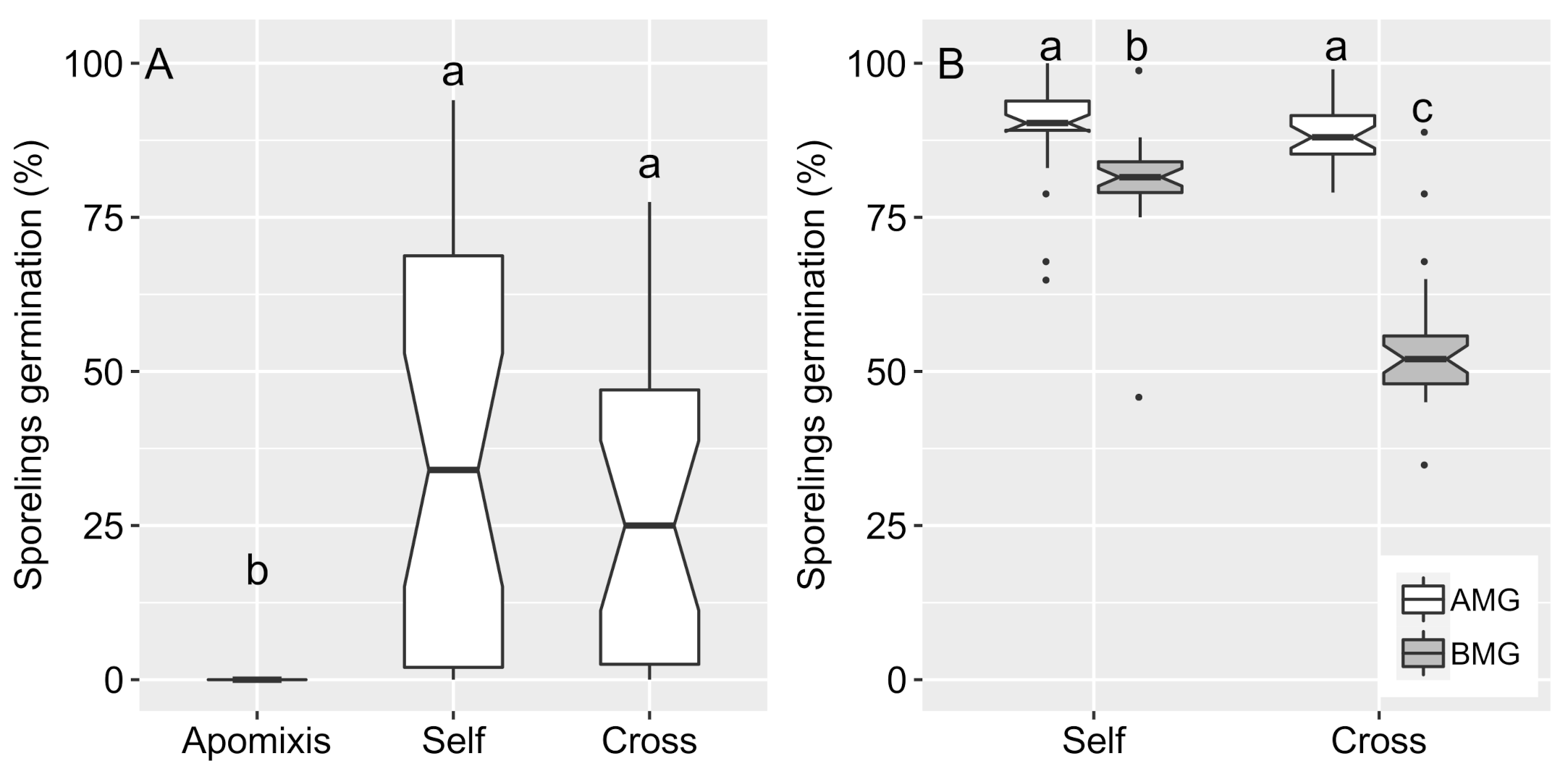
3.1. Spore Germination and Sporophyte Growth from Sporangia Collected during the Dry and Rainy Seasons
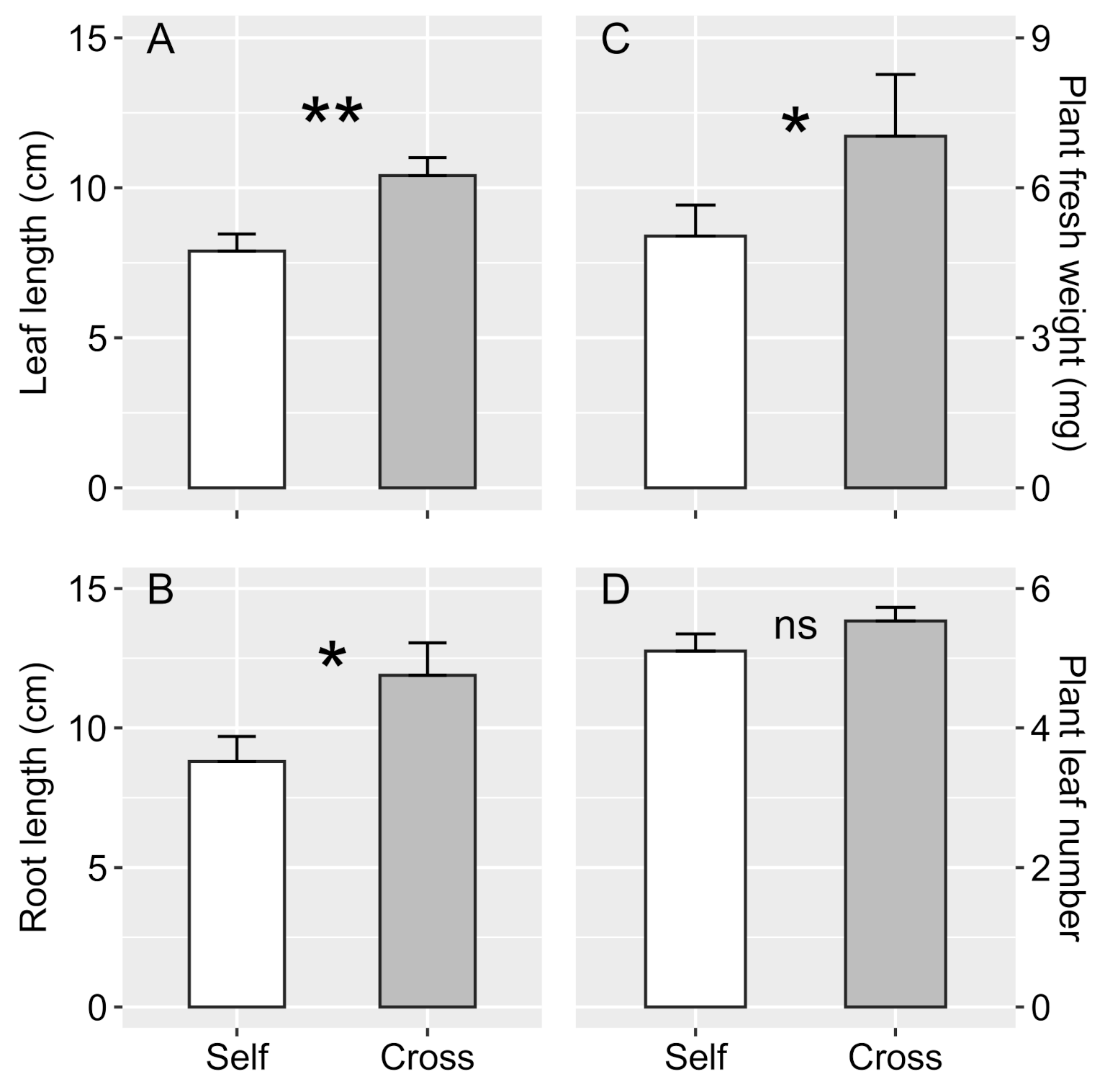
3.2. Sexual Propagation and Progenies Growth
3.3. Isoëtes cangae Ex Situ Growth and Sporangia Production
3.4. Vegetative or Asexual Reproduction
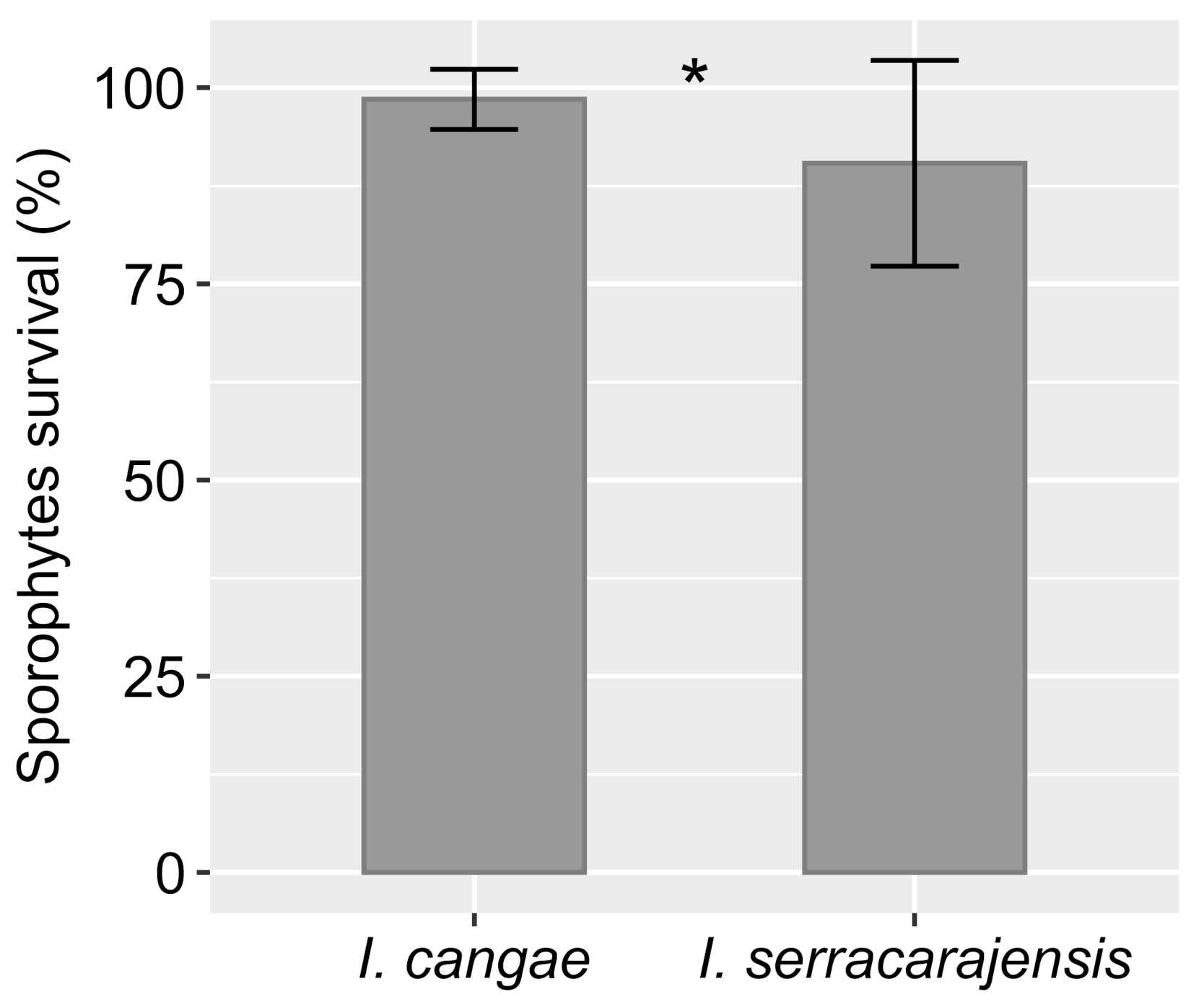
3.5. Sporophytes Survival
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pereira, J.B.S.; Salino, A.; Arruda, A.; Stutzel, T. Two New Species of Isoetes (Isoetaceae) from Northern Brazil. Phytotaxa 2016, 272, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Smolders, A.J.P.; Lucassen, E.C.H.E.T.; Roelofs, J.G.M. The Isoetid Environment: Biogeochemistry and Threats. Aquat. Bot. 2002, 73, 325–350. [Google Scholar] [CrossRef]
- Hetherington, A.J.; Berry, C.M.; Dolan, L. Networks of Highly Branched Stigmarian Rootlets Developed on the First Giant Trees. Proc. Natl. Acad. Sci. USA 2016, 113, 6695–6700. [Google Scholar] [CrossRef] [Green Version]
- Hickey, R.J.; Macluf, C.; Taylor, W.C. A Re-Evaluation of Isoetes savatieri Franchet in Argentina and Chile. Am. Fern J. 2003, 93, 126–136. [Google Scholar] [CrossRef]
- Troia, A.; Pereira, J.B.; Kim, C.; Taylor, W.C. The Genus Isoetes (Isoetaceae): A Provisional Checklist of the Accepted and Unresolved Taxa. Phytotaxa 2016, 277, 101–145. [Google Scholar] [CrossRef]
- Nunes, G.L.; Oliveira, R.R.M.; Guimarães, J.T.F.; Giulietti, A.M.; Caldeira, C.; Vasconcelos, S.; Pires, E.; Dias, M.; Watanabe, M.; Pereira, J.; et al. Quillworts from the Amazon: A Multidisciplinary Populational Study on Isoetes serracarajensis and Isoetes cangae. PLoS ONE 2018, 13, e0201417. [Google Scholar] [CrossRef]
- De Oliveira Mota, N.F.; Watanabe, M.T.C.; Zappi, D.C.; Hiura, A.L.; Pallos, J.; Viveros, R.S.; Giulietti, A.M.; Viana, P.L. Amazon Canga: The Unique Vegetation of Carajás Revealed by the List of Seed Plants. Rodriguésia 2018, 69, 1435–1488. [Google Scholar] [CrossRef] [Green Version]
- Skirycz, A.; Castilho, A.; Chaparro, C.; Carvalho, N.; Tzotzos, G.; Siqueira, J.O. Canga Biodiversity, a Matter of Mining. Front. Plant Sci. 2014, 5, 653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giulietti, A.M.; Giannini, T.C.; Mota, N.F.O.; Watanabe, M.T.C.; Viana, P.L.; Pastore, M.; Silva, U.C.S.; Siqueira, M.F.; Pirani, J.R.; Lima, H.C.; et al. Edaphic Endemism in the Amazon: Vascular Plants of the Canga of Carajás, Brazil. Bot. Rev. 2019, 85, 357–383. [Google Scholar] [CrossRef] [Green Version]
- Souza-Filho, P.W.M.; de Souza, E.B.; Silva Júnior, R.O.; Nascimento, W.R.; de Versiani Mendonça, B.R.; Guimarães, J.T.F.; Dall’Agnol, R.; Siqueira, J.O. Four Decades of Land-Cover, Land-Use and Hydroclimatology Changes in the Itacaiúnas River Watershed, Southeastern Amazon. J. Environ. Manag. 2016, 167, 175–184. [Google Scholar] [CrossRef]
- Lansdown, R.V. Isoetes cangae. The IUCN Red List of Threatened Species 2019. 2019. Available online: https://www.iucnredlist.org/ (accessed on 20 May 2021).
- Abeli, T.; Caldeira, C.F.; Barni, E.; Siniscalco, C.; Parco, V.; Rossi, G. The Ecology of the Endemic Quillwort Isoëtes malinverniana: From Basic Research to Legal and in Situ Conservation. Aquat. Conserv. Mar. Freshw. Ecosyst. 2020, 30, 1719–1722. [Google Scholar] [CrossRef]
- Abeli, T.; Cauzzi, P.; Rossi, G.; Pistoja, F.; Mucciarelli, M. A Gleam of Hope for the Critically Endangered Isoëtes malinverniana: Use of Small-Scale Translocations to Guide Conservation Planning. Aquat. Conserv. Mar. Freshw. Ecosyst. 2018, 28, 501–505. [Google Scholar] [CrossRef]
- Caldeira, C.F.; Abranches, C.B.; Gastauer, M.; Ramos, S.J.; Guimarães, J.T.F.; Pereira, J.B.S.; Siqueira, J.O. Sporeling Regeneration and Ex Situ Growth of Isoëtes cangae (Isoetaceae): Initial Steps towards the Conservation of a Rare Amazonian Quillwort. Aquat. Bot. 2019, 152, 51–58. [Google Scholar] [CrossRef]
- Oh, M.J.; Kim, C.; Na, H.R.; Shin, H.; Liu, J.R.; Choi, H.-K.; Kim, S.W. High Frequency Sporophytes Regeneration from the Spore Culture of the Endangered Aquatic Fern Isoetes coreana. Am. J. Plant Sci. 2013, 4, 14–20. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.P.; Araujo, J.V.S.R.; Lopes, A.V.S.; Vettorazzi, J.C.F.; Boechat, M.S.B.; Arêdes, F.A.; Campos, N.V.; Calderon, E.N.; Santos, F.M.G.; Fernandes, T.N.; et al. The Genetic Diversity and Population Structure of Two Endemic Amazonian Quillwort (Isoetes L.) Species. PeerJ 2020, 8, e10274. [Google Scholar] [CrossRef]
- Čtvrtlíková, M.; Znachor, P.; Vrba, J. The Effect of Temperature on the Phenology of Germination of Isoetes lacustris. Preslia 2014, 86, 279–292. [Google Scholar]
- Karrfalt, E. Some Observations on the Reproductive Anatomy of Isoetes andicola. Am. Fern J. 1999, 89, 198–203. [Google Scholar] [CrossRef]
- Taylor, W.C.; Luebke, N.T. Germinating Spores and Growing Sporelings of Aquatic Isoetes. Am. Fern J. 1986, 76, 21–24. [Google Scholar] [CrossRef]
- Huang, Y.-M.; Chang, Y.-L.; Chiou, W.-L. Soil Spore Bank of Isoetes taiwanensis DeVol (Isoetaceae). Int. J. Plant Reprod. Biol. 2015, 7, 1–7. [Google Scholar] [CrossRef]
- Doyle, J.J.; Doyle, J.L. Isolation of Plant DNA from Fresh Tissue. Focus 1990, 12, 13–15. [Google Scholar]
- Zandonadi, D.B.; Duarte, H.M.; Santos, M.P.; Prado, L.A.S.; Martins, R.L.; Calderon, E.N.; Fernandes, A.C.A.; Santos, Q.S.; Nunes, F.J.G.; Ribeiro, L.C.F.; et al. Ecophysiology of Two Endemic Amazon Quillworts. Aquat. Bot. 2021, 170, 103350. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Abeli, T.; Mucciarelli, M. Notes on the Natural History and Reproductive Biology of Isoëtes malinverniana. Am. Fern J. 2010, 100, 235–237. [Google Scholar] [CrossRef]
- Magrini, S.; Azzella, M.M.; Bolpagni, R.; Zucconi, L. In Vitro Propagation of Isoëtes sabatina (Isoetaceae): A Key Conservation Challenge for a Critically Endangered Quillwort. Plants 2020, 9, 887. [Google Scholar] [CrossRef] [PubMed]
- Dalapicolla, J.; Alves, R.; Jaffé, R.; Vasconcelos, S.; Pires, E.S.; Nunes, G.L.; de Souza Pereira, J.B.; Guimarães, J.T.F.; Dias, M.C.; Fernandes, T.N.; et al. Conservation Implications of Genetic Structure in the Narrowest Endemic Quillwort from the Eastern Amazon. Ecol. Evol. 2021, ece3.7812. [Google Scholar] [CrossRef]
- Renzaglia, K.S.; Garbary, D.J. Motile Gametes of Land Plants: Diversity, Development, and Evolution. Crit. Rev. Plant Sci. 2001, 20, 107–213. [Google Scholar] [CrossRef]
- Sahoo, P.K.; Guimarães, J.T.F.; Souza-Filho, P.W.M.; Silva, M.S.D.A.; Silva Junior, R.O.; Pessim, G.; Moraes, B.C.D.E.; Pessoa, P.F.P.; Rodrigues, T.M.; Costa, M.F.D.A.; et al. Influence of Seasonal Variation on the Hydro-Biogeochemical Characteristics of Two Upland Lakes in the Southeastern Amazon, Brazil. An. Acad. Bras. Ciênc. 2016, 88, 2211–2227. [Google Scholar] [CrossRef] [Green Version]
- Caplen, C.A.; Werth, C.R. Isozymes of the Isoetes riparia Complex, I. Genetic Variation and Relatedness of Diploid Species. Syst. Bot. 2000, 25, 235. [Google Scholar] [CrossRef]
- Taylor, W.C.; Hickey, R.J. Habitat, Evolution, and Speciation in Isoetes. Ann. Mo. Bot. Gard. 1992, 79, 613–622. [Google Scholar] [CrossRef]
- Chen, Y.-Y.; Liao, L.; Li, W.; Li, Z.-Z. Genetic Diversity and Population Structure of the Endangered Alpine Quillwort Isoetes hypsophila Hand.-Mazz. Revealed by AFLP Markers. Plant Syst. Evol. 2010, 290, 127–139. [Google Scholar] [CrossRef]
- Arrigo, N.; Therrien, J.; Anderson, C.L.; Windham, M.D.; Haufler, C.H.; Barker, M.S. A Total Evidence Approach to Understanding Phylogenetic Relationships and Ecological Diversity in Selaginella Subg. Tetragonostachys. Am. J. Bot. 2013, 100, 1672–1682. [Google Scholar] [CrossRef]
- DiMichele, W.A.; Davis, J.I.; Olmstead, R.G. Origins of Heterospory and the Seed Habit: The Role of Heterochrony. Taxon 1989, 38, 1–11. [Google Scholar] [CrossRef]
- Herlihy, C.R.; Eckert, C.G. Genetic Cost of Reproductive Assurance in a Self-Fertilizing Plant. Nature 2002, 416, 320–323. [Google Scholar] [CrossRef]
- Caldeira, C.F.; Jeanguenin, L.; Chaumont, F.; Tardieu, F. Circadian Rhythms of Hydraulic Conductance and Growth Are Enhanced by Drought and Improve Plant Performance. Nat. Commun. 2014, 5, 5365. [Google Scholar] [CrossRef] [Green Version]
- Gifford, R.M.; Thorne, J.H.; Hitz, W.D.; Giaquinta, R.T. Crop Productivity and Photoassimilate Partitioning. Science 1984, 225, 801–808. [Google Scholar] [CrossRef] [PubMed]
- Koester, R.P.; Skoneczka, J.A.; Cary, T.R.; Diers, B.W.; Ainsworth, E.A. Historical Gains in Soybean (Glycine max Merr.) Seed Yield Are Driven by Linear Increases in Light Interception, Energy Conversion, and Partitioning Efficiencies. J. Exp. Bot. 2014, 65, 3311–3321. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Caldeira, C.F.; Lopes, A.V.S.; Aguiar, K.C.; Ferreira, A.L.; Araujo, J.V.S.; Gomes, V.M.S.; Zandonadi, D.B.; Abranches, C.B.; Ramos, S.J.; Gastauer, M.; et al. Distinct Reproductive Strategy of Two Endemic Amazonian Quillworts. Diversity 2021, 13, 348. https://doi.org/10.3390/d13080348
Caldeira CF, Lopes AVS, Aguiar KC, Ferreira AL, Araujo JVS, Gomes VMS, Zandonadi DB, Abranches CB, Ramos SJ, Gastauer M, et al. Distinct Reproductive Strategy of Two Endemic Amazonian Quillworts. Diversity. 2021; 13(8):348. https://doi.org/10.3390/d13080348
Chicago/Turabian StyleCaldeira, Cecilio F., Arthur V. S. Lopes, Keyvilla C. Aguiar, Aline L. Ferreira, João V. S. Araujo, Vinnícius M. S. Gomes, Daniel B. Zandonadi, Cinthia B. Abranches, Silvio J. Ramos, Markus Gastauer, and et al. 2021. "Distinct Reproductive Strategy of Two Endemic Amazonian Quillworts" Diversity 13, no. 8: 348. https://doi.org/10.3390/d13080348
APA StyleCaldeira, C. F., Lopes, A. V. S., Aguiar, K. C., Ferreira, A. L., Araujo, J. V. S., Gomes, V. M. S., Zandonadi, D. B., Abranches, C. B., Ramos, S. J., Gastauer, M., Campos, N. V., Gestinari, L. M. S., Prado, L. A., Santos, F. M. G., Martins, R. L., Esteves, F. A., Oliveira, G., & Santos, M. P. (2021). Distinct Reproductive Strategy of Two Endemic Amazonian Quillworts. Diversity, 13(8), 348. https://doi.org/10.3390/d13080348