
Diversity and Diet Differences of Small Mammals in Commensal Habitats




Abstract
:
1. Introduction
2. Materials and Methods
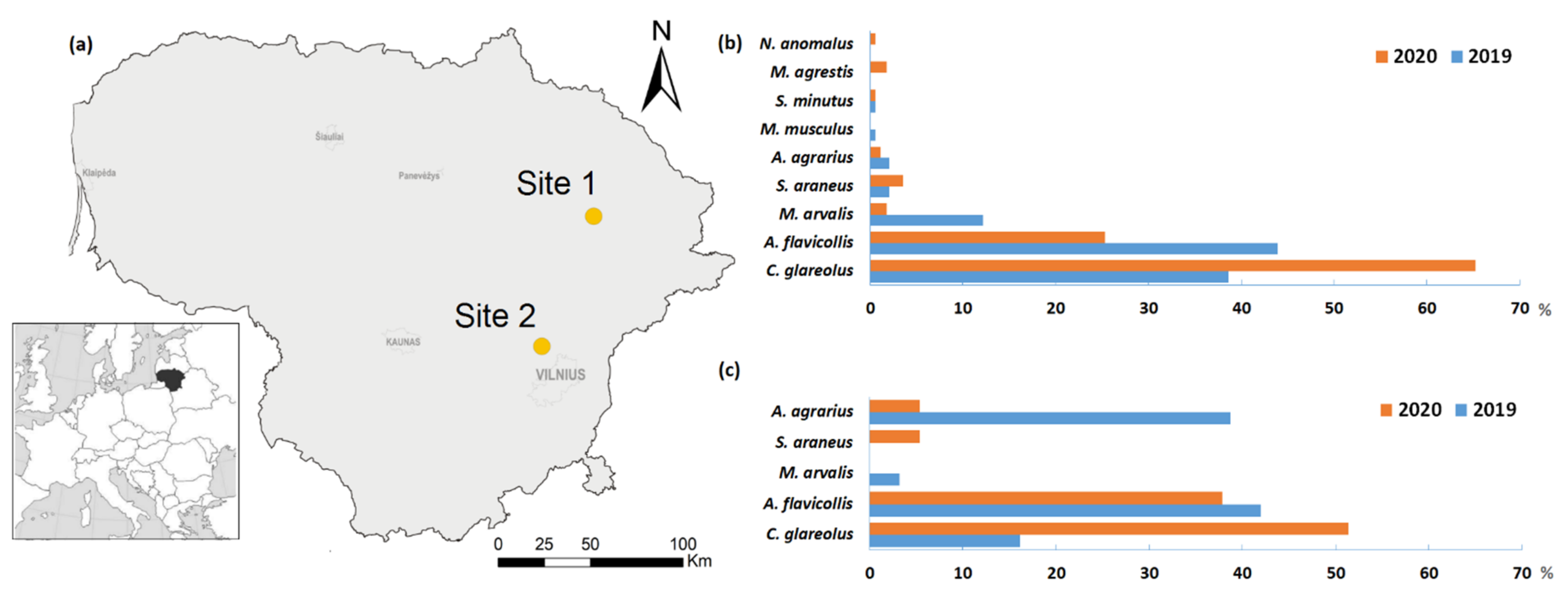
2.1. Study Site
2.2. Small Mammal Trapping
2.3. Stable Isotope Analysis
2.4. Statistical Analyses
3. Results
3.1. Species Composition and Diversity
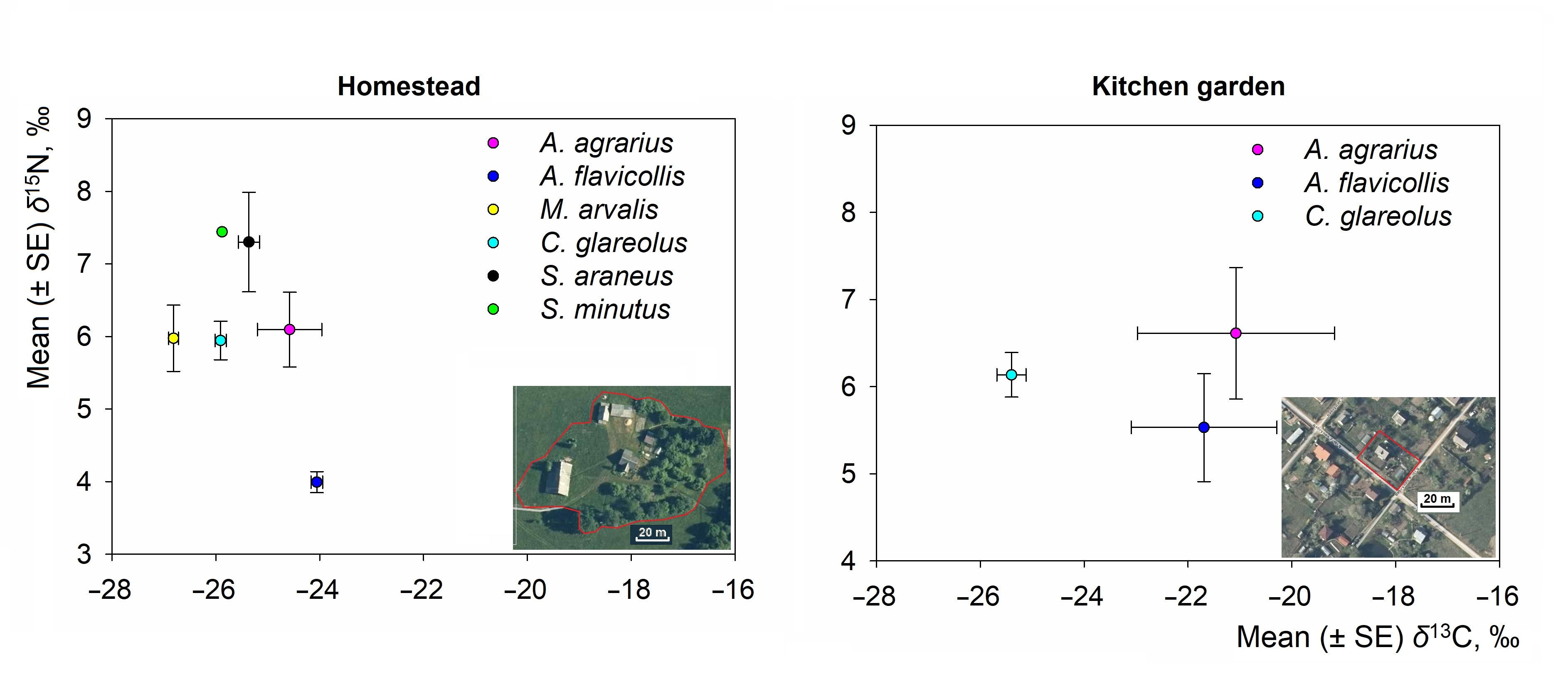
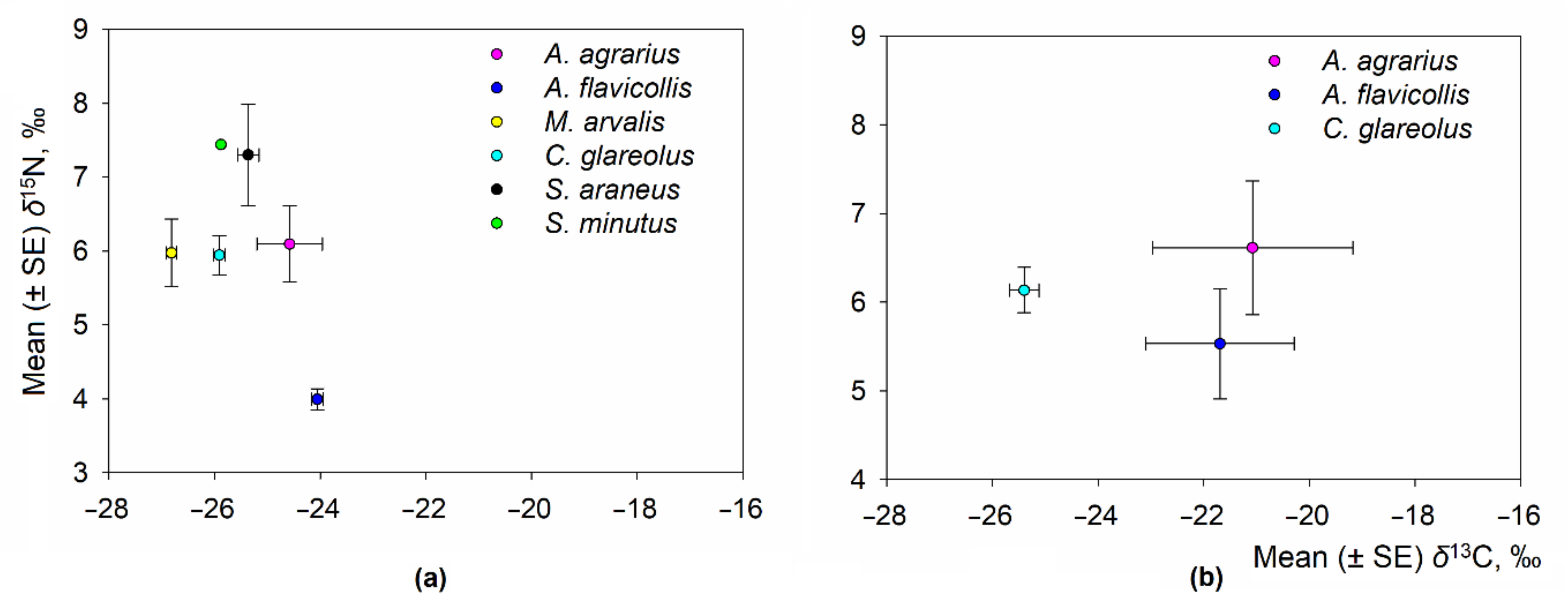
3.2. Interspecific Differences of Isotopic Niche of Small Mammals in Commensal Habitats
4. Discussion
5. Conclusions
- (1).
- We present the first data on small mammal trophic ecology in commensal habitats (homestead and kitchen garden) in the northern part of the middle latitudes.
- (2).
- The highest small mammal species richness occurred in commensal habitats that provided food. It was low in the kitchen garden, which was under the highest human influence.
- (3).
- The most numerous small mammal species in the homestead had a higher degree of dietary separation (central ellipses not overlapping) than the kitchen garden (1.27% to 35.97% overlap of SEAc between species).
- (4).
- Temporal changes of δ13C and δ15N values in the hair were not equally expressed in different species.
- (5).
- Human influence in commensal habitats may have different effects on the diets of different species of small mammals, where separation according to δ15N is better expressed.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Régnier, R. Attack on Rodents in Rural Areas. In Annales d’Hygiene Publique, Industrielle et Sociale; J.-B. Baillière et Fils: Paris, France, 1928; Volume 6, pp. 427–433. [Google Scholar]
- Khlyap, L.A.; Warshavsky, A.A. Synanthropic and agrophilic rodents as invasive alien mammals. Russ. J. Biol. Invasions 2010, 1, 301–312. [Google Scholar] [CrossRef]
- Paliy, A.P.; Sumakova, N.V.; Antoniuk, A.A.; Behas, V.L.; Panasenko, A.S. Development and effectiveness of domestic bait in mouse-like rodents control. Ukr. J. Ecol. 2021, 11, 209–215. [Google Scholar] [CrossRef]
- Hornok, S.; Földvári, G.; Rigó, K.; Meli, M.L.; Gönczi, E.; Répási, A.; Farkas, R.; Papp, I.; Kontschán, J.; Hofmann-Lehmann, R. Synanthropic rodents and their ectoparasites as carriers of a novel haemoplasma and vector-borne, zoonotic pathogens indoors. Parasite. Vector. 2015, 8, 27. [Google Scholar] [CrossRef] [Green Version]
- Jahan, N.A.; Lindsey, L.L.; Larsen, P.A. The role of peridomestic rodents as reservoirs for zoonotic foodborne pathogens. Vector-Borne Zoonot. 2021, 21, 133–148. [Google Scholar] [CrossRef]
- Meheretu, Y.; Granberg, Å.; Berhane, G.; Khalil, H.; Lwande, O.W.; Mitiku, M.; Welegerima, K.; Bellocq, J.G.d.; Bryja, J.; Abreha, H.; et al. Prevalence of Orthohantavirus-Reactive Antibodies in Humans and Peri-Domestic Rodents in Northern Ethiopia. Viruses 2021, 13, 1054. [Google Scholar] [CrossRef]
- Panti-May, J.A.; Sodá-Tamayo, L.; Gamboa-Tec, N.; Cetina-Franco, R.; Cigarroa-Toledo, N.; Machaín-Williams, C.; Robles, M.d.; Hernández-Betancourt, S.F. Perceptions of rodent-associated problems: An experience in urban and rural areas of Yucatan, Mexico. Urban. Ecosyst. 2017, 20, 983–988. [Google Scholar] [CrossRef]
- Govinda Raj, G. Rodents. In Pests and Their Management, Omkar; Springer: Singapore, 2018; pp. 973–1013. [Google Scholar] [CrossRef]
- Khan, W.; Das, S.N.; Ullah, H.; Panhwar, W.A.; Ahmed, S.; Ahmad, M.S.; Kamal, M.; Ahmad, A.; Mohsin, M.U.; Hussain, A.; et al. Distribution of commensal rodents in some shops of three districts in Malakand region, Pakistan. Braz. J. Biol. 2021, 82. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Guevara, J.N.A.; Ball, B.A. Urbanization alters small rodent community composition but not abundance. PeerJ 2018, 6, e4885. [Google Scholar] [CrossRef]
- Mazza, V.; Dammhahn, M.; Lösche, E.; Eccard, J.A. Small mammals in the big city: Behavioural adjustments of non-commensal rodents to urban environments. Glob. Chang. Biol. 2020, 26, 6326–6337. [Google Scholar] [CrossRef]
- Baker, P.J.; Harris, S. Urban mammals: What does the future hold? An analysis of the factors affecting patterns of use of residential gardens in Great Britain. Mammal. Rev. 2007, 37, 297–315. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Balčiauskienė, L. On the Doorstep, Rodents in Homesteads and Kitchen Gardens. Animals 2020, 10, 856. [Google Scholar] [CrossRef]
- Santini, L.; González-Suárez, M.; Russo, D.; Gonzalez-Voyer, A.; von Hardenberg, A.; Ancillotto, L. One strategy does not fit all: Determinants of urban adaptation in mammals. Ecol. Lett. 2019, 22, 365–376. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.; Vial, F.; Pietravalle, S.; Cowan, D. Results of a 15-year systematic survey of commensal rodents in English dwellings. Sci. Rep. 2017, 7, 15882. [Google Scholar] [CrossRef] [Green Version]
- Assefa, A.; Chelmala, S. Comparison of rodent community between natural and modified habitats in Kafta-Sheraro National Park and its adjoining villages, Ethiopia: Implication for conservation. J. Basic Appl. Zool. 2019, 80, 59. [Google Scholar] [CrossRef] [Green Version]
- Mayamba, A.; Byamungu, R.M.; Broecke, B.V.; Leirs, H.; Hieronimo, P.; Nakiyemba, A.; Isabirye, M.; Kifumba, D.; Kimaro, D.N.; Mdangi, M.E.; et al. Factors influencing the distribution and abundance of small rodent pest species in agricultural landscapes in Eastern Uganda. J. Vertebr. Biol. 2020, 69, 20002. [Google Scholar] [CrossRef]
- Pocock, M.J.; Searle, J.B.; White, P.C. Adaptations of animals to commensal habitats: Population dynamics of house mice Mus musculus domesticus on farms. J. Anim. Ecol. 2004, 73, 878–888. [Google Scholar] [CrossRef]
- Hope, A.G.; Gragg, S.F.; Nippert, J.B.; Combe, F.J. Consumer roles of small mammals within fragmented native tallgrass prairie. Ecosphere 2021, 12, e03441. [Google Scholar] [CrossRef]
- Ward-Fear, G.; Brown, G.P.; Pearson, D.; Shine, R. Untangling the influence of biotic and abiotic factors on habitat selection by a tropical rodent. Sci. Rep. 2021, 11, 12895. [Google Scholar] [CrossRef]
- Hunter, C.M.; Williamson, D.H.Z.; Pearson, M.; Saikawa, E.; Gribble, M.O.; Kegler, M. Safe community gardening practices: Focus groups with garden leaders in Atlanta, Georgia. Local Environ. 2020, 25, 18–35. [Google Scholar] [CrossRef]
- Chalmin-Pui, L.S.; Griffiths, A.; Roe, J.; Heaton, T.; Cameron, R. Why garden?–Attitudes and the perceived health benefits of home gardening. Cities 2021, 112, 103118. [Google Scholar] [CrossRef]
- Vávra, J.; Smutná, Z.; Hruška, V. Why I Would Want to Live in the Village If I Was Not Interested in Cultivating the Plot? A Study of Home Gardening in Rural Czechia. Sustainability 2021, 13, 706. [Google Scholar] [CrossRef]
- Ciftcioglu, G.C. Social preference-based valuation of the links between home gardens, ecosystem services, and human well-being in Lefke Region of North Cyprus. Ecosyst. Serv. 2017, 25, 227–236. [Google Scholar] [CrossRef]
- Bimonte, S.; Billaud, O.; Fontaine, B.; Martin, T.; Flouvat, F.; Hassan, A.; Rouillier, N.; Sautotd, L. Collect and analysis of agro-biodiversity data in a participative context: A business intelligence framework. Ecol. Inform. 2021, 61, 101231. [Google Scholar] [CrossRef]
- Ebel, R.; Menalled, F.; Ahmed, S.; Gingrich, S.; Baldinelli, G.M.; Félix, G.F. How biodiversity loss affects society. In Handbook on the Human Impact of Agriculture; James, H.S., Ed.; Edward Elgar Publishing: Cheltenham, UK, 2021; pp. 352–376. [Google Scholar] [CrossRef]
- Khanam, S.; Mushtaq, M.; Kayani, A.R.; Nadeem, M.S.; Beg, M.A. Small mammal community composition and abundance in rural human habitations of Pothwar, Pakistan. Trop. Ecol. 2017, 58, 515–524. [Google Scholar]
- Mariën, J.; Kourouma, F.; Magassouba, N.F.; Leirs, H.; Fichet-Calvet, E. Movement patterns of small rodents in Lassa fever-endemic villages in Guinea. EcoHealth 2018, 15, 348–359. [Google Scholar] [CrossRef]
- Peng, H.; Hou, D.; Zhu, W. Study on the changes of population of main rodents in Jianchuan area in different years and habitats. Octa J. Biosci. 2019, 7, 33–36. [Google Scholar]
- Singleton, G.R.; Lorica, R.P.; Htwe, N.M.; Stuart, A.M. Rodent management and cereal production in Asia–balancing food security and conservation. Pest. Manag. Sci. 2021. [Google Scholar] [CrossRef]
- Kays, R.; Parsons, A.W. Mammals in and around suburban yards, and the attraction of chicken coops. Urban. Ecosyst. 2014, 17, 691–705. [Google Scholar] [CrossRef]
- Steinberg, M.K. Neotropical kitchen gardens as a potential research landscape for conservation biologists. Conserv. Biol. 1998, 12, 1150–1152. [Google Scholar] [CrossRef]
- Bastien, M.; Vaniscotte, A.; Combes, B.; Umhang, G.; Raton, V.; Germain, E.; Villenaag, I.; Auberta, D.; Boué, F.; Poulleab, M.-L. Identifying drivers of fox and cat faecal deposits in kitchen gardens in order to evaluate measures for reducing contamination of fresh fruit and vegetables. Food Waterborne Parasitol. 2019, 14, e00034. [Google Scholar] [CrossRef]
- Balčiauskas, L. Methods of Investigation of Terrestrial Ecosystems; Part. I. Animal Surveys; VU Leidykla: Vilnius, Lithuania, 2004; p. 183. [Google Scholar]
- Balciauskas, L.; Balciauskiene, L.; Janonyte, A. Reproduction of the root vole (Microtus oeconomus) at the edge of its distribution range. Turk. J. Zool. 2012, 36, 668–675. [Google Scholar] [CrossRef]
- Metcalf, D. Multiple Thymus Grafts in Aged Mice. Nature 1965, 208, 87–88. [Google Scholar] [CrossRef] [PubMed]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Balčiauskienė, L.; Remeikis, V. Immediate increase in isotopic enrichment in small mammals following the expansion of a great cormorant colony. Biogeosciences 2018, 15, 3883–3891. [Google Scholar] [CrossRef] [Green Version]
- Balčiauskas, L.; Skipitytė, R.; Jasiulionis, M.; Trakimas, G.; Balčiauskienė, L.; Remeikis, V. The impact of Great Cormorants on biogenic pollution of land ecosystems: Stable isotope signatures in small mammals. Sci. Total Environ. 2016, 565, 376–383. [Google Scholar] [CrossRef]
- Jackson, A.L.; Inger, R.; Parnell, A.C.; Bearhop, S. Comparing isotopic niche widths among and within communities: SIBER—Stable Isotope Bayesian Ellipses in R. J. Anim. Ecol. 2011, 80, 595–602. [Google Scholar] [CrossRef] [PubMed]
- Krebs, C.J. Ecological Methodology, 2nd ed.; Addison-Wesley Educational Publishers, Inc.: Menlo Park, CA, USA, 1999; 620p. [Google Scholar]
- Balčiauskas, L.; Balčiauskienė, L.; Stirkė, V. Mow the Grass at the Mouse’s Peril: Diversity of Small Mammals in Commercial Fruit Farms. Animals 2019, 9, 334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guiry, E.; Buckley, M. Urban rats have less variable, higher protein diets. Proc. R. Soc. B 2018, 285, 20181441. [Google Scholar] [CrossRef] [Green Version]
- Galetti, M.; Guevara, R.; Neves, C.L.; Rodarte, R.R.; Bovendorp, R.S.; Moreirac, M.; Hopkins, J.B.; Yeakelf, D.J. Defaunation affects the populations and diets of rodents in Neotropical rainforests. Biol. Conserv. 2015, 190, 2–7. [Google Scholar] [CrossRef] [Green Version]
- Nakagawa, M.; Hyodo, F.; Nakashizuka, T. Effect of forest use on trophic levels of small mammals: An analysis using stable isotopes. Can. J. Zool. 2007, 85, 472–478. [Google Scholar] [CrossRef]
- Dammhahn, M.; Randriamoria, T.M.; Goodman, S.M. Broad and flexible stable isotope niches in invasive non-native Rattus spp. in anthropogenic and natural habitats of central eastern Madagascar. BMC Ecol. 2017, 17, 16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Camargo, N.F.; Reis, G.G.; Camargo, A.C.L.; Nardoto, G.B.; Kneitel, J.M.; Vieira, E.M. Seasonal isotopic niche of a rodent: High between-individual variation but no changes in individual niche width during the rich-resource period. Biotropica 2021, 53, 966–975. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Stable Isotopes Reveal the Dominant Species to Have the Widest Trophic Niche of Three Syntopic Microtus Voles. Animals 2021, 11, 1814. [Google Scholar] [CrossRef]
- Balčiauskas, L.; Skipitytė, R.; Garbaras, A.; Stirkė, V.; Balčiauskienė, L.; Remeikis, V. Isotopic niche of syntopic granivores in commercial orchards and meadows. Animals 2021, in press. [Google Scholar]
- Balčiauskas, L.; Skipitytė, R.; Balčiauskienė, L.; Jasiulionis, M. Resource partitioning confirmed by isotopic signatures allows small mammals to share seasonally flooded meadows. Ecol. Evol. 2019, 9, 5479–5489. [Google Scholar] [CrossRef]
- Forbes, K.M.; Stuart, P.; Mappes, T.; Hoset, K.S.; Henttonen, H.; Huitu, O. Diet quality limits summer growth of field vole populations. PLoS ONE 2014, 9, e91113. [Google Scholar] [CrossRef] [Green Version]
- Loza, E.; Cotton, J.M.; Smiley, T.M.; Terry, R.C. Using Small Mammals to Understand the Effects of Urbanization in Southern California over the Last 100 Years. In Proceedings of the AGU Fall Meeting Abstracts, New Orleans, LA, USA, 11–15 December 2017; Volume 2017, p. GC11B-0736. Available online: https://agu.confex.com/agu/fm17/meetingapp.cgi/Paper/259271 (accessed on 11 May 2021).
- Gortat, T.; Barkowska, M.; Tkowska, A.G.S.; Pieniążek, A.; Kozakiewicz, A.; Kozakiewicz, M. The effects of urbanization—small mammal communities in a gradient of human pressure in Warsaw city, Poland. Pol. J. Ecol. 2014, 62, 163–172. [Google Scholar] [CrossRef]
- DePasquale, C.; Li, X.; Harold, M.; Mueller, S.; McLaren, S.; Mahan, C. Selection for increased cranial capacity in small mammals during a century of urbanization. J. Mammal. 2020, 101, 1706–1710. [Google Scholar] [CrossRef]
- Moll, R.J.; Cepek, J.D.; Lorch, P.D.; Dennis, P.M.; Robison, T.; Montgomery, R.A. At what spatial scale (s) do mammals respond to urbanization? Ecography 2020, 43, 171–183. [Google Scholar] [CrossRef] [Green Version]
- White, A.J.; Poulin, R.G.; Wissel, B.; Doucette, J.L.; Somers, C.M. Agricultural land use alters trophic status and population density of deer mice (Peromyscus maniculatus) on the North American Great Plains. Can. J. Zool. 2012, 90, 868–874. [Google Scholar] [CrossRef]
- Fedriani, J.M.; Fuller, T.K.; Sauvajot, R.M. Does availability of anthropogenic food enhance densities of omnivorous mammals? An example with coyotes in southern California. Ecography 2001, 24, 325–331. [Google Scholar] [CrossRef]
- Crowley, B.E.; Castro, I.; Soarimalala, V.; Goodman, S.M. Isotopic evidence for niche partitioning and the influence of anthropogenic disturbance on endemic and introduced rodents in central Madagascar. Sci. Nat. 2018, 105, 44. [Google Scholar] [CrossRef]
- Soininen, E.M.; Ehrich, D.; Lecomte, N.; Yoccoz, N.G.; Tarroux, A.; Berteaux, D.; Gauthier, G.; Gielly, L.; Brochmann, C.; Gussarova, G.; et al. Sources of variation in small rodent trophic niche: New insights from DNA metabarcoding and stable isotope analysis. Isot. Environ. Health Stud. 2014, 50, 361–381. [Google Scholar] [CrossRef] [Green Version]
- Langton, S.D.; Cowan, D.P.; Meyer, A.N. The occurrence of commensal rodents in dwellings as revealed by the 1996 English House Condition Survey. J. Appl. Ecol. 2001, 38, 699–709. [Google Scholar] [CrossRef]
- Hulme-Beaman, A.; Dobney, K.; Cucchi, T.; Searle, J.B. An ecological and evolutionary framework for commensalism in anthropogenic environments. Trends Ecol. Evol. 2016, 31, 633–645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baltensperger, A.P.; Huettmann, F.; Hagelin, J.C.; Welker, J.M. Quantifying trophic niche spaces of small mammals using stable isotopes (δ15N and δ13C) at two scales across Alaska. Can. J. Zool. 2015, 93, 579–588. [Google Scholar] [CrossRef] [Green Version]
- Mori, E.; Ferretti, F.; Fattorini, N. Alien war: Ectoparasite load, diet and temporal niche partitioning in a multi-species assembly of small rodents. Biol. Invasions 2019, 21, 3305–3318. [Google Scholar] [CrossRef]
- Pardo, L.H.; Nadelhoffer, K.J. Using nitrogen isotope ratios to assess terrestrial ecosystems at regional and global scales. In Isoscapes: Understanding Movement, Pattern, and Process on Earth through Isotope Mapping; West, J.B., Bowen, G.J., Dawson, T.E., Tu, K.P., Eds.; Springer: Dordrecht, The Netherlands, 2010; pp. 221–249. [Google Scholar] [CrossRef]
- Bertolino, S.; Colangelo, P.; Mori, E.; Capizzi, D. Good for management, not for conservation: An overview of research, conservation and management of Italian small mammals. Hystrix Ital. J. Mammal. 2015, 26, 25–35. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Homesteads | Kitchen Gardens | ||
|---|---|---|---|---|
| 2019 | 2020 | 2019 | 2020 | |
| Species richness, S | 7 (4–7) | 8 (5–8) | 4 (4–4) | 4 (4–4) |
| Dominance, D | 0.35 (0.31–0.41) | 0.48 (0.40–0.58) * | 0.36 (0.31–0.40) | 0.42 (0.36–0.47) |
| Diversity, H | 1.23 (1.03–1.36) | 1.05 (0.80–1.23) | 1.13 (1.03–1.23) | 1.01 (0.87–1.15) |
| Species | N | δ13C Values, ‰ | δ15N Values, ‰ | ||||
|---|---|---|---|---|---|---|---|
| Mean ± SE | Min–Max | Range | Mean ± SE | Min–Max | Range | ||
| Homestead habitat | |||||||
| A. agrarius | 2 | −24.58 ± 0.62 | −25.20–(−23.96) | 1.24 | 6.10 ± 0.52 | 5.58–6.61 | 1.03 |
| A. flavicollis | 57 | −24.06 ± 0.11 | −27.13–(−22.85) | 4.28 | 3.99 ± 0.14 | 0.15–8.35 | 8.20 |
| M. arvalis | 18 | −26.82 ± 0.09 | −27.59–(−26.22) | 1.37 | 5.98 ± 0.46 | 2.92–9.09 | 6.17 |
| C. glareolus | 56 | −25.91 ± 0.11 | −27.90–(−23.98) | 3.92 | 5.94 ± 0.26 | 2.44–10.70 | 8.26 |
| S. araneus | 3 | −25.36 ± 0.20 | −25.64–(−24.97) | 0.67 | 7.30 ± 0.69 | 6.27–8.60 | 2.33 |
| Kitchen garden habitat | |||||||
| A. agrarius | 7 | −21.08 ± 1.90 | −24.69–(−13.35) | 11.34 | 6.61 ± 0.76 | 3.70–9.60 | 5.90 |
| A. flavicollis | 11 | −21.69 ± 1.40 | −25.56–(−12.24) | 13.32 | 5.53 ± 0.62 | 2.40–9.16 | 6.76 |
| C. glareolus | 5 | −25.40 ± 0.28 | −27.43–(−24.57) | 2.86 | 6.14 ± 0.26 | 4.78–7.06 | 2.28 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Balčiauskas, L.; Balčiauskienė, L.; Garbaras, A.; Stirkė, V. Diversity and Diet Differences of Small Mammals in Commensal Habitats. Diversity 2021, 13, 346. https://doi.org/10.3390/d13080346
Balčiauskas L, Balčiauskienė L, Garbaras A, Stirkė V. Diversity and Diet Differences of Small Mammals in Commensal Habitats. Diversity. 2021; 13(8):346. https://doi.org/10.3390/d13080346
Chicago/Turabian StyleBalčiauskas, Linas, Laima Balčiauskienė, Andrius Garbaras, and Vitalijus Stirkė. 2021. "Diversity and Diet Differences of Small Mammals in Commensal Habitats" Diversity 13, no. 8: 346. https://doi.org/10.3390/d13080346
APA StyleBalčiauskas, L., Balčiauskienė, L., Garbaras, A., & Stirkė, V. (2021). Diversity and Diet Differences of Small Mammals in Commensal Habitats. Diversity, 13(8), 346. https://doi.org/10.3390/d13080346