
Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies



Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Helminth Examination
2.3. Data Analysis
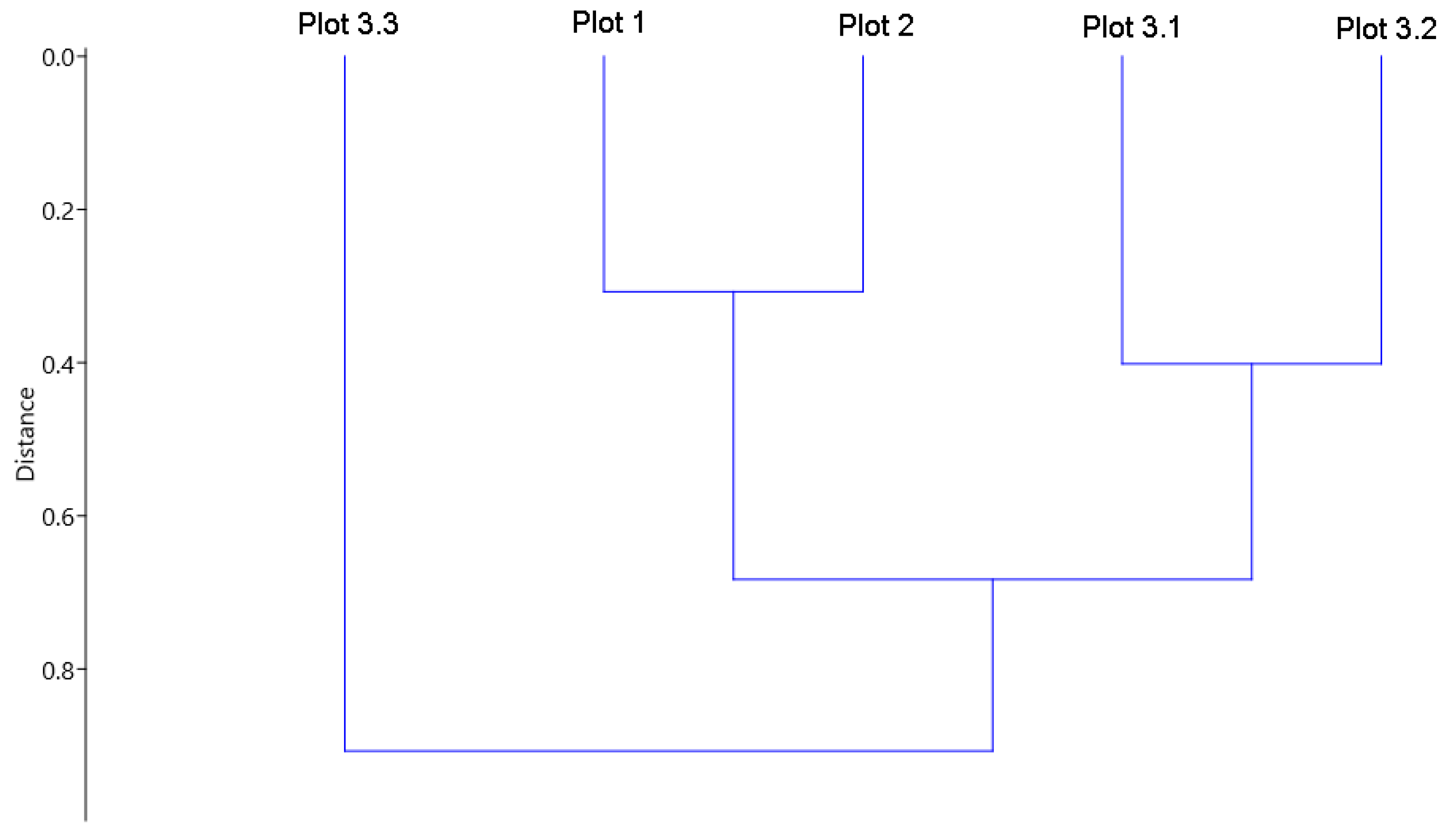
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubois, A.; Ohler, A. Frogs of the subgenus Pelophylax (Amphibia, Anura, Genus Rana): A catalogue of available and valid scientific names, with comments on name-bearing types, complete synonymies, proposed common names, and maps showing all type localities. Zool. Pol. 1994, 39, 139–204. [Google Scholar]
- Kuzmin, Y.; Dmytrieva, I.; Marushchak, O.; Morozov-Leonov, S.; Oskyrko, O.; Nekrasova, O. Helminth species and infracommunities in frogs Pelophylax ridibundus and P. esculentus (Amphibia: Ranidae) in Northern Ukraine. Acta Parasitol. 2020, 65, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Wells, K.D. The Ecology and Behavior of Amphibians; The University of Chicago Press: Chicago, IL, USA, 2007; p. 1162. [Google Scholar] [CrossRef]
- David, A.; Sas, I.; Cicort-Lucaciu, A.-Ş.; Lezau, O. Food composition of some green water frog populations from Livada Forest, Satu Mare County, Romania. Muz. Olten. Craiova. Oltenia. Stud. Şi Comunicări. Ştiinţele Naturii. 2010, 26, 169–174. [Google Scholar]
- Paunovic, A.; Bjelić-Čabrilo, O.; Šimić, S. The diet of water frogs (Pelophylax esculentus ‘complex’) from the Petrovaradinski Rit marsh (Serbia). Arch. Biol. Sci. 2010, 62, 797–804. [Google Scholar] [CrossRef]
- Chikhlyaev, I.V.; Ruchin, A.B.; Fayzulin, A.I. Short communication: An overview of the trematodes fauna of pool frog Pelophylax lessonae (Camerano, 1882) in the Volga basin, Russia: 2. Larval stages. Nusant. Biosci. 2019, 11, 106–111. [Google Scholar] [CrossRef]
- Bjelić-Čabrilo, O.; Popović, E.; Paunović, A. Helminthofauna of Pelophylax kl. esculentus (Linne, 1758) from Petrovaradinski Rit Marsh (Serbia). Helminthologia 2009, 46, 107–111. [Google Scholar] [CrossRef] [Green Version]
- Popiołek, M.; Rozenblut-Kościsty, B.; Kot, M.; Nosal, W.; Ogielska, M. Endoparasitic helminths of water frog complex in Poland: Do differences exist between the parental species Pelophylax ridibundus and Pelophylax lessonae, and their natural hybrid Pelophylax esculentus? Helminthologia 2011, 48, 108–115. [Google Scholar] [CrossRef]
- Chikhlyaev, I.V.; Ruchin, A.B.; Fayzulin, A.I. An overview of the trematodes fauna of pool frog Pelophylax lessonae (Camerano, 1882) in the Volga basin, Russia: 1. Adult stages. Nusant. Biosci. 2018, 10, 256–262. [Google Scholar] [CrossRef]
- Čeirāns, A.; Gravele, E.; Gavarane, I.; Pupins, M.; Mezaraupe, L.; Rubenina, I.; Kvach, Y.; Skute, A.; Oskyrko, O.; Nekrasova, O.; et al. Helminth communities in amphibians from Latvia, with an emphasis on their connection to host ecology. J. Helmint. 2021, 95, e48. [Google Scholar] [CrossRef]
- Niewiadomska, K.; Pojmanska, T. Multiple strategies of digenean trematodes to complete their life cycles. Wiad. Parazytol. 2011, 57, 233–241. [Google Scholar]
- Galaktionov, K.V.; Dobrovolskij, A.A. The trematode life cycle as a system of adaptations. In The Biology and Evolution of Trematodes. Chapter 2; Kluwer Academic Publishers: London, UK, 2003; pp. 215–317. [Google Scholar] [CrossRef]
- Chikhlyaev, I.V.; Ruchin, A.B.; Fayzulin, A.I. Parasitic nematodes of Pool frog (Pelophylax lessonae) in the Volga basin. J. MVZ Cordoba 2019, 24, 7314–7321. [Google Scholar] [CrossRef] [Green Version]
- Kirillova, N.Y.; Kirillov, A.A. The life cycle of Cosmocerca ornata (Dujardin, 1845) (Nematoda: Cosmocercidae)—A parasite of amphibians. Inland Water Biol. 2021, 3, 286–301. [Google Scholar] [CrossRef]
- Chikhlyaev, I.V.; Ruchin, A.B.; Alekseev, S.K.; Korzikov, V.A. On the helminth fauna of the European common frog, Rana temporaria Linnaeus, 1768 (Amphibia: Anura) from different habitats in Kaluga Region. Curr. Stud. Herpetol. 2013, 13, 58–63. (In Russian) [Google Scholar]
- Chikhlyaev, I.V.; Kirillova, N.Y.; Kirillov, A.A. Ecological analysis of trematodes (Trematoda) of marsh frog Pelophylax ridibundus (Ranidae, Anura) from various habitats of the National Park “Samarskaya Luka” (Russia). Nat. Conserv. Res. 2018, 3 (Suppl. 1), 36–50. [Google Scholar] [CrossRef] [Green Version]
- Chikhlyaev, I.V.; Ruchin, A.B. Helminths of amphibians (Amphibia) in beaver ponds in the Central Russia. AACL Bioflux 2020, 13, 3810–3821. [Google Scholar]
- Rubenina, I.; Kirjusina, M.; Čeirāns, A.; Gravele, E.; Gavarane, I.; Pupins, M.; Krasnov, B.R. Environmental, anthropogenic, and spatial factors affecting species composition and species associations in helminth communities of water frogs (Pelophylax esculentus complex) in Latvia. Parasitol. Res. 2021, 120, 3461–3474. [Google Scholar] [CrossRef]
- Johnson, P.T.J.; Wood, C.L.; Joseph, M.B.; Preston, D.L.; Haas, S.E.; Springe, Y.P. Habitat heterogeneity drives the host-diversity-begets parasite-diversity relationship: Evidence from experimental and field studies. Ecol. Lett. 2016, 19, 752–761. [Google Scholar] [CrossRef] [Green Version]
- Chikhlyaev, I.V.; Ruchin, A.B.; Kirillov, A.A. Ecological analysis of the helminth fauna in Bufo bufo (Amphibia: Anura) from various habitats. Nat. Conserv. Res. 2020, 5 (Suppl. 2), 1–10. [Google Scholar] [CrossRef]
- Kirillov, A.A.; Kirillova, N.Y. Helminth fauna of reptiles in the National Park “Smolny”, Russia. Nat. Conserv. Res. 2021, 6, 9–22. [Google Scholar] [CrossRef]
- Koprivnikar, J.; Marcogliese, D.J.; Rohr, J.R.; Orlofske, S.A.; Raffel, T.R.; Johnson, P.T.J. Macroparasite infections of amphibians: What can they tell us? EcoHealth 2012, 9, 342–360. [Google Scholar] [CrossRef]
- Buss, N.; Hua, J. Parasite susceptibility in an amphibian host is modified by salinization and predators. Environ. Pollut. 2018, 236, 754–763. [Google Scholar] [CrossRef] [PubMed]
- Hamann, M.I.; Kehr, A.I.; González, C.E. Species affinity and infracommunity ordination of helminths of Leptodactylus chaquensis (Anura: Leptodactylidae) in two contrasting environments from northeastern Argentina. J. Parasitol. 2006, 92, 1171–1179. [Google Scholar] [CrossRef] [PubMed]
- King, K.C.; Mclaughlin, J.D.; Gendron, A.D.; Pauli, B.D.; Giroux, I.; Rondeau, B.; Boily, M.; Juneau, P.; Marcogliese, D.J. Impacts of agriculture on the parasite communities of northern leopard frogs (Rana pipiens) in southern Quebec, Canada. Parasitology 2007, 134, 2063–2080. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, K.C.; Mclaughlin, J.D.; Boily, M.; Marcogliese, D.J. Effects of agricultural landscape and pesticides on parasitism in native bullfrogs. Biol. Conserv. 2010, 143, 302–310. [Google Scholar] [CrossRef]
- Campião, K.M.; Ribas, A.C.A.; Silva, I.C.O.; Dalazen, G.T.; Tavares, L.E.R. Anuran helminth communities from contrasting nature reserve and pasture sites in the Pantanal wetland, Brazil. J. Helminthol. 2016, 1, 1–6. [Google Scholar] [CrossRef]
- Kuzmin, S.; Beebee, T.; Adreone, F.; Niström, P.; Antony, B.P.; Schmidt, B.R.; Ogrodowczyk, A.; Ogielska, M.; Cogalniceanu, D.; Kovácz, T.; et al. Pelophylax lessonae, Pool Frog. In The IUCN Red List of Threatened Species; IUCN Red List: London, UK, 2009; pp. 1–9. [Google Scholar] [CrossRef]
- Fayzulin, A.I.; Zamaletdinov, R.I.; Litvinchuk, S.N.; Rosanov, J.M.; Borkin, L.J.; Ermakov, O.A.; Ruchin, A.B.; Lada, G.A.; Svinin, A.O.; Bashinsky, I.V.; et al. Species composition and distributional peculiarities of green frogs (Pelophylax esculentus complex) in Protected Areas of the Middle Volga Region (Russia). Nat. Conserv. Res. 2018, 3 (Suppl. 1), 1–16. [Google Scholar] [CrossRef] [Green Version]
- Zeisset, I.; Hoogesteger, T. A reassessment of the biogeographic range of northern clade pool frogs (Pelophylax lessonae). Herpetol. J. 2018, 28, 63–72. [Google Scholar]
- Ruchin, A.; Artaev, O.; Sharapova, E.; Ermakov, O.; Zamaletdinov, R.; Korzikov, V.; Bashinsky, I.; Pavlov, A.; Svinin, A.O.; Ivanov, A.; et al. Occurrence of the amphibians in the Volga, Don River basins and adjacent territories (Russia): Research in 1996–2020. Biodivers. Data J. 2020, 8, e61378. [Google Scholar] [CrossRef]
- Dedukh, D.; Litvinchuk, J.; Svinin, A.; Litvinchuk, S.; Rosanov, J.; Krasikova, A. Variation in hybridogenetic hybrid emergence between populations of water frogs from the Pelophylax esculentus complex. PLoS ONE 2019, 14, e0224759. [Google Scholar] [CrossRef]
- Ivanov, A.Y.; Ruchin, A.B.; Fayzulin, A.I.; Chikhlyaev, I.V.; Litvinchuk, S.N.; Kirillov, A.A.; Svinin, A.O.; Ermakov, O.A. The first record of natural transfer of mitochondrial DNA from Pelophylax cf. bedriagae into P. lessonae (Amphibia, Anura). Nat. Conserv. Res. 2019, 4, 125–128. [Google Scholar] [CrossRef] [Green Version]
- Herczeg, D.; Voros, J.; Vegvari, Z.; Brooks, D.R. Helminth parasites of the Pelophylax esculentus complex (Anura: Ranidae) in Hortobágy National Park (Hungary). Comp. Parasitol. 2016, 83, 36–48. [Google Scholar] [CrossRef] [Green Version]
- Skryabin, K.I. Method of Complete Helminthological Dissection of Vertebrates, Including Man; Moscow University: Moscow, Russia, 1928; p. 45. (In Russian) [Google Scholar]
- Bykhovskaya-Pavlovskaya, I.E. Parasites of Fishes, a Study Guide; Nauka: Leningrad, Russia, 1985; p. 123. (In Russian) [Google Scholar]
- Ryzhikov, K.M.; Sharpilo, V.P.; Shevchenko, N.N. Amphibian Helminths of the USSR Fauna; Nauka: Moscow, Russia, 1980; p. 279. (In Russian) [Google Scholar]
- Sudarikov, V.E.; Shigin, A.A.; Kurochkin, Y.V.; Lomakin, V.V.; Stenko, R.P.; Yurlova, N.I. Metacercariae of Trematodes—Parasites of Freshwater Aquatic Organisms in Central Russia; Nauka: Moscow, Russia, 2002; p. 298. (In Russian) [Google Scholar]
- Fauna Europaea. Available online: https://fauna-eu.org/cdm_dataportal/search/results/taxon?ws=portal%2Ftaxon%2Ffind&query=Coenagrion+pulchellum+&form_build_id=form-I7SfdygUtDbRDakknqqgPvI9jKid3ZL2_iwbWDOcTMQ&form_id=cdm_dataportal_search_taxon_form&search%5BdoTaxaByCommonNames%5D=&search%5BdoSynonyms%5D=&search%5BdoTaxa%5D=1&search%5BpageSize%5D=25&search%5BpageNumber%5D=0 (accessed on 14 January 2022).
- Tkach, V.V.; Pawlowski, J.; Mariaux, J.; Swiderski, Z. Molecular phylogeny of the suborder Plagiorchiata and its position in the system of Digenea. In Interrelations of the Platyhelminthes; Littlewood, D.T.J., Bray, R.A., Eds.; Taylor & Francis: London, UK, 2001; pp. 186–193. [Google Scholar]
- Gibson, D.I.; Jones, A.; Bray, R.A. Keys to the Trematoda; CABI Publishing and London, The Natural History Museum: Wallingford, UK, 2002; Volume 1, p. 521. [Google Scholar]
- Leon-Regagnon, V.; Brooks, D. Molecular phylogeny of Haematoloechus Looss, 1899. J. Parasitol. 2003, 89, 1206–1211. [Google Scholar] [CrossRef] [Green Version]
- Olson, P.D.; Cribb, T.H.; Tkach, V.V.; Bray, R.A.; Littlewood, D.T.J. Phylogeny and classification of the Digenea (Platyhelminthes: Trematoda). Int. J. Parasitol. 2003, 33, 733–755. [Google Scholar] [CrossRef]
- Bray, R.A.; Gibson, D.I.; Jones, A. Keys to the Trematoda; CABI Publishing and London, The Natural History Museum: Wallingford, UK, 2008; Volume 3, p. 848. [Google Scholar]
- Hodda, M. Phylum Nematoda Cobb 1932. In: Zhang, Z.-Q. (Ed.) Animal biodiversity: An outline of higher-level classification and survey of taxonomic richness. Zootaxa 2011, 3148, 63–95. [Google Scholar] [CrossRef]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. revisited. J. Parasitol. 1997, 83, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Kirillov, A.A. Helminth communities of grass snake Natrix natrix L. (Reptilia: Colubridae) from South of Middle Volga region. Izv. Samara Sci. Cent. Russ. Acad. Sci. 2011, 13, 127–134. (In Russian) [Google Scholar]
- Bakanov, A.I. Quantitated Estimation of Dominance in Ecological Communities; Institute of Inland Water Biology, USSR Academy of Sciences: Borok, Russia, 1987; p. 64. (In Russian) [Google Scholar]
- Magurran, A.E. Ecological Diversity and its Measurement; Mir: Moscow, Russia, 1992; p. 182. (In Russian) [Google Scholar]
- Pigulevsky, S.V. The family Gorgoderidae Looss, 1901. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1952; Volume 7, pp. 605–760. (In Russian) [Google Scholar]
- Pigulevsky, S.V. The family Gorgoderidae Looss, 1901. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1953; Volume 8, pp. 251–615. (In Russian) [Google Scholar]
- Vojtková, L. Motolice obojživelníků ČSSR. I. Dospĕlé motolice. In Folia Facultatis Scientiarum Naturalium Universitatis Purkynianae Brunensis. Biologia; Universita, J.E. Purkynĕ: Brno, Czechoslovakia, 1974; Volume 25, p. 133. (In Czech) [Google Scholar]
- Kalabekov, A. Development cycles of some trematodes of longlegged wood frog (Rana macrocnemis Boul.). In Ecology and Biology of Animals on the Northern Slopes Central Caucasus, Collection of Zoological works; Naniev, V.I., Ed.; North Ossetian State University: Ordzhonikidze, Russia, 1976; pp. 3–42. (In Russian) [Google Scholar]
- Skryabin, K.I.; Antipin, D.N. Superfamily Plagiorchioidea Dollfus, 1930. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; Nauka: Moscow, Russia, 1962; Volume 20, pp. 49–166. (In Russian) [Google Scholar]
- Thiel, P.H. Die Entwicklung von Agamodistomum anopheles zum Pneumonoeces variegatus Rud. Zentralbl. Bakter. 1930, 117, 103–112. (In German) [Google Scholar]
- Dobrowolsky, A.A. The life cycle of Pneumonoeces asper Looss, 1899 (Plagiorchiidae, Pneumonoecinae). In Proceedings of the Scientific Conference of the All-Union Society of Helminthologists. Part 4; USSR Academy of Sciences Publishing House: Moscow, Russia, 1965; pp. 59–64. (In Russian) [Google Scholar]
- Grabda, B. Life cycle of Haematoloechus similis (Looss, 1899) (Trematoda: Plagiorchidae). Acta Parasitol. Polon. 1960, 8, 357–366. [Google Scholar]
- Dobrowolsky, A.A. Some information about the life cycle of the trematode Opisthioglyphe ranae (Froelich, 1791) (Plagiorchiidae). Helminthologia 1965, 3, 205–221. [Google Scholar]
- Grabda-Kazubska, B. Studies on abbreviation of the life-cycle in Opisthioglyphe ranae (Froelich, 1791) and O. rastellus (Olsson, 1876) (Trematoda, Plagiorchiidae). Acta Parasitol. Polon. 1969, 16, 20–27. [Google Scholar]
- Khotenovsky, I.A. The family Pleurogenidae Looss, 1899. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; Nauka: Moscow, Russia, 1970; Volume 23, pp. 136–297. (In Russian) [Google Scholar]
- Grabda-Kazubska, B. Life cycle of Pleurogenes claviger (Rudolphi, 1819) (Trematoda: Pleurogenidae). Acta Parasitol. Polon. 1971, 19, 337–348. [Google Scholar]
- Neuhaus, W. Entwicklung und Biologie von Pleurogenoides medians Olss. Zool. Yahrb. Syst. 1940, 74, 207–242. (In German) [Google Scholar]
- Shevchenko, N.N.; Vergun, G.I. On the life cycle of amphibian trematodes Prosotocus confusus (Looss, 1894) Looss, 1899. Helminthologia 1961, 3, 294–298. [Google Scholar]
- Skryabin, K.I. Suborder Paramphistomatata (Szidat, 1936) Skrjabin et Schulz, 1937. In Trematodes of Animals and Humans. Basics of Trematodology; Skryabin, K.I., Ed.; Publishing House of the USSR Academy of Sciences: Moscow, Russia, 1949; Volume 3, p. 624. (In Russian) [Google Scholar]
- Grabda-Kazubska, B. Observations on the life cycle of Diplodiscus subclavatus (Pallas, 1760) (Trematoda, Diplodiscidae). Acta Parasitol. Polon. 1980, 27, 261–271. [Google Scholar]
- Dobrowolsky, A.A. The life cycle of Paralepoderma cloacicola (Lühe, 1909) Dollfus, 1950 (Trematoda, Plagiorchiidae). Leningr. Univ. Bull. 1969, 9, 28–38. (In Russian) [Google Scholar]
- Grabda-Kazubska, B. A study of the trematode genus Paralepoderma Dollfus, 1950 (Trematoda: Plagiorchiidae). Acta Parasitol. Polon. 1975, 23, 463–484. [Google Scholar]
- Sudarikov, V.E. Order Strigeidida (La Rue, 1926) Sudarikov, 1959. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; USSR Academy of Sciences Publishing House: Moscow, Russia, 1959; Volume 16, pp. 219–631. (In Russian) [Google Scholar]
- Sudarikov, V.E. The biology of trematodes Strigea strigis (Schr., 1788) and S. sphaerula (Rud., 1803). In Works of the Helminthology Laboratory of the USSR Academy of SCIENCES (GELAN); USSR Academy of Sciences Publishing House: Moscow, Russia, 1960; Volume 10, pp. 217–226. (In Russian) [Google Scholar]
- Odening, K. Der Lebenszyklus des Trematoden Strigea strigis (Schrank) im Raum Berlin. Monatsber. Deutsch. Acad. Wissensch. Zu Berl. 1966, 8, 696–697. (In German) [Google Scholar]
- Odening, K. Die lebenszyklen von Strigea falconispalumbi (Viborg), S. strigis (Schrank) und S. sphaerula (Rudolphi) (Trematoda, Strigeida) im Raum Berlin. Zool. Yahrb. Syst. 1967, 94, 1–67. (In German) [Google Scholar]
- Odening, K. Der Lebenszyklus des Trematoden Strigea sphaerula (Rudolphi) im Raum Berlin. Monatsber. Deutsch. Acad. Wissensch. Zu Berl. 1966, 8, 695–696. (In German) [Google Scholar]
- Sudarikov, V.E. Order Strigeidida (La Rue, 1926) Sudarikov, 1959. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; USSR Academy of Sciences Publishing House: Moscow, Russia, 1960; Volume 17, pp. 157–533. (In Russian) [Google Scholar]
- Odening, K. Der Lebenszyklus von Neodiplostomum spathoides Dubois (Trematoda, Strigeida) im Raum Berlin nebst Baitragen zur Entwicklungsweise verwandter Arten. Zool Yahrb. Syst. 1965, 92, 523–624. (In German) [Google Scholar]
- Odening, K. Die Lebenszyklen der Trematoda Neodiplostomum spathoides Dubois und N. attenuatum (v. Linstow) im Raum Berlin. Monatsber. Deutsch. Acad. Wissensch. Zu Berl. 1965, 7, 952–954. (In German) [Google Scholar]
- Potekhina, L.F. The development cycle of alariosis pathogen in foxes and dogs. Rep. USSR Acad. Sci. 1951, 76, 325–327. (In Russian) [Google Scholar]
- Sudarikov, V.E. Biological features of trematodes of the genus Alaria. In Works of the Helminthology Laboratory of the USSR Academy of Sciences (GELAN); USSR Academy of Sciences Publishing House: Moscow, Russia, 1959; Volume 9, pp. 326–332. (In Russian) [Google Scholar]
- Sudarikov, V.E. Suborder Strigeata La Rue, 1926. Superfamily Diplostomatoidea Nicoll, 1937. In Trematodes of Animals and Human. Basics of Trematodology; Skryabin, K.I., Ed.; USSR Academy of Sciences Publishing House: Moscow, Russia, 1960; Volume 18, pp. 453–694. (In Russian) [Google Scholar]
- Hendrix, W.M.L. Observations of the routes of infection of Oswaldocruzia filiformis (Nematoda, Trichostrongylidae) in amphibian. Z. Parasitenk. 1983, 69, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Moravec, F.; Vojtkova, L. Variabilität von zwei Nematodenarten Oswaldocruzia filiformis (Goeze, 1782) und Oxysomatium brevicaudatum (Zeder, 1800). In Der Gemeinsamen Parasiten der Europäischen Amphibien und Reptilien; Scripta Facultatis Scientiarum Naturalium Universitatis Purkynianae Brunensis. Biologia; Universita J.E. Purkynĕ: Brno, Czechoslovakia, 1975; Volume 2, pp. 61–76. (In German) [Google Scholar]
- Kirillova, N.Y.; Kirillov, A.A.; Shchenkov, S.V.; Chikhlyaev, I.V. Oswaldocruzia filiformis sensu lato (Nematoda: Molineidae) from amphibians and reptiles in European Russia: Morphological and molecular data. Nat. Conserv. Res. 2020, 5, 41–56. [Google Scholar] [CrossRef]
- Skryabin, K.I.; Shikhobalova, N.P.; Lagodovskaya, E.A. Oxyuridae of animals and humans. Part 2. In Basics of Nematodology; Skryabin, K.I., Ed.; USSR Academy of Sciences Publishing House: Moscow, Russia, 1961; Volume 10, p. 500. (In Russian) [Google Scholar]
- Desportes, C. Forcipomyia velox Winn et Sycorax silacea Curtis, vecteurs d’Icosiella neglecta (Diesing, 1850) filaire commune de la grenouille verte. Ann. Parasit. Hum. Et Comp. 1942, 19, 53–68. (In French) [Google Scholar] [CrossRef]
- Mikulíček, P.; Mešková, M.; Cyprich, M.; Jablonski, D.; Papežík, P.; Hamidi, D.; Pekşen, C.; Vörös, J.; Herczeg, D.; Benovics, M. Weak population-genetic structure of a widely distributed nematode parasite of frogs in the western Palearctic. J. Zool. Syst. Evol. Res. 2021, 59, 1689–1702. [Google Scholar] [CrossRef]
- Yoder, H.R.; Coggins, J.R. Helminth communities in five species of sympatric amphibians from three adjacent ephemeral ponds in southeastern Wisconsin. J. Parasitol. 2007, 93, 755–760. [Google Scholar] [CrossRef]
- Campião, K.M.; Da Silva, R.J.; Ferreira, V.L. Helminth parasite communities of allopatric populations of the frog Leptodactylus podicipinus from Pantanal, Brazil. J. Helmint. 2014, 88, 13–19. [Google Scholar] [CrossRef]
- Kuzmenkin, D.V. The habitat distribution of freshwater mollusks in the plain part of Upper Ob basin. Izv. Altai State Univ. 2013, 3, 80–85. (In Russian) [Google Scholar]
- Bogatov, V.V.; Kijashko, P.V. Class Bivalvia Linnaeus, 1758. In Key for Determination of Freshwater Zooplankton and Zoobenthos of the European Russia. Vol. 2. Zoobenthos; Tsalolikhin, S.Y., Ed.; KMK Scientific Press: Moscow, Russia, 2016; pp. 286–334. (In Russian) [Google Scholar]
- Vinarski, M.V.; Kantor, Y.I. Analytical Catalogue of Fresh and Brackish Water Molluscs of Russia and Adjacent Countries; A.N. Severtsov Institute of Ecology and Evolution of RAS: Moscow, Russia, 2016; p. 544. [Google Scholar]
- Grishutkin, G.F.; Lapshin, A.S.; Spiridonov, S.N.; Artaev, O.N.; Ruchin, A.B.; Kuznetsov, V.A.; Andreychev, A.V. Vertebrate animals of “Smolny” National Park; RAS Commission on Biodiversity Conservation: Moscow, Russia, 2013; p. 56, [Flora and fauna of National parks. Issue 9]. (In Russian) [Google Scholar]
- Vedernikov, A.A.; Svinin, A.O.; Ermakov, O.A.; Chelyadnikova, Y.A.; Musatov, G.A.; Drobot, G.P. Granulomatous inflammations in the intestine of Pelophylax ridibundus (Anura: Ranidae) caused by Brandesia turgida (Plathelminthes: Digenea). Nat. Conserv. Res. 2020, 5 (Suppl. 2), 72–76. [Google Scholar] [CrossRef]
- Belevich, O.E.; Yurchenko, Y.A. Assemblages of bloodsucking mosquito larvae (Diptera: Culicidae) in water bodies of the Northern Kulunda steppe. Parazitologiya 2011, 45, 182–193. (In Russian) [Google Scholar]
- Khlyzova, T.A.; Fiodorova, O.A. Breeding habitats of blood-sucking mosquitoes (Diptera, Culicidae) in the south of Tyumen oblast. Ukr. J. Ecol. 2018, 8, 166–170. (In Russian) [Google Scholar] [CrossRef]
- Kijashko, P.V.; Soldatenko, E.V.; Vinarski, M.V. Class Gastropoda Cuvier, 1797. In Key for Determination of Freshwater Zooplankton and Zoobenthos of the European Russia. Vol. 2. Zoobenthos; Tsalolikhin, S.Y., Ed.; KMK Scientific Press: Moscow, Russia, 2016; pp. 335–438, (In Russian). [Google Scholar] [CrossRef]
- Shimkovich, E.D. Ecological features of Lymnaea stagnalis s.l. (Gastropoda, Lymnaeidae) in the waters of Tatarstan. Univ. Proc. Volga Region. Nat. Sciences. Biol. 2016, 2, 11–18. (In Russian) [Google Scholar] [CrossRef]
- Yavorskaya, N.M. Zoobenthos in watercourses and water bodies in the “Bolonsky” State Nature Reserve (Russia). Nat. Conserv. Res. 2020, 5, 64–79. [Google Scholar] [CrossRef]
- Frolova, E.N. Larvae of Trematodes of Mollusks in South Karelia Lakes; Nauka: Leningrad, Russia, 1975; p. 184. (In Russian) [Google Scholar]
- Faltýnková, A. Larval trematodes (Digenea) in molluscs from small water bodies near České Budšjovice, Czech Republic. Acta Parasitol. 2005, 50, 49–55. [Google Scholar]
- Faltýnková, A.; Sures, B.; Kostadinova, A. Biodiversity of trematodes in their intermediate mollusc and fish hosts in the freshwater ecosystems of Europe. Syst. Parasitol. 2016, 93, 283–293. [Google Scholar] [CrossRef]
- Kolomyts, E.G. Regional Model of Global Changes in the Natural Environment; Nauka: Moscow, Russia, 2003; p. 371. (In Russian) [Google Scholar]
- Kirillov, A.A.; Kirillova, N.Y. Comparative analysis of the helminth fauna of Natrix natrix and Natrix tessellata (Reptilia, Colubridae) in the Samarskaya Luka National Park (Russia). Nat. Conserv. Res. 2019, 4, 12–25. (In Russian) [Google Scholar] [CrossRef]
- Ieshko, E.P.; Novokhatskaya, O.V. Trends in succession of parasitofauna of fish of eutrophied water bodies. J. Ichthyol. 2008, 48, 665–670. [Google Scholar] [CrossRef]


{kind=link}
| Helminths Species | Floodplain Water Bodies | Forest Water Bodies | |||
|---|---|---|---|---|---|
| Plot 1 | Plot 2 | Plot 3.1 | Plot 3.2 | Plot 3.3 | |
| Gorgodera cygnoides | 4.35 ± 2.45 (1–1) 0.04 ± 0.02 | 6.67 ± 3.22 (1–1) 0.07 ± 0.03 | |||
| Gorgodera microovata | 1.45 ± 1.44 (1) 0.01 ± 0.01 | 6.67 ± 3.22 (2–6) 0.22 ± 0.12 | 2.70 ± 2.51 (2) 0.05 ± 0.05 | 5.00 ± 4.88 (2) 0.10 ± 0.10 | 12.25 ± 4.68 (1–5) 0.27 ± 0.13 |
| Gorgodera asiatica | 2.70 ± 2.51 (1) 0.03 ± 0.03 | ||||
| Gorgoderina vitelliloba | 1.45 ± 1.44 (1) 0.01 ± 0.01 | ||||
| Haematoloechus variegatus | 5.80 ± 2.81 (1–2) 0.10 ± 0.05 | 8.33 ± 3.57 (1–3) 0.17 ± 0.08 | 16.22 ± 6.06 (1–4) 0.24 ± 0.12 | 15.00 ± 7.99 (1–2) 0.20 ± 0.12 | 6.12 ± 3.42 (1–2) 0.10 ± 0.07 |
| Haematoloechus asper | 2.90 ± 2.02 (1–2) 0.04 ± 0.03 | ||||
| Skrjabinoeces similis | 4.35 ± 2.45 (1–1) 0.04 ± 0.02 | 1.67 ± 1.65 (5) 0.08 ± 0.08 | 4.08 ± 2.83 (1–1) 0.04 ± 0.03 | ||
| Opisthioglyphe ranae | 62.32 ± 5.83 (1–126) 8.41 ± 2.12 | 5.00 ± 2.81 (1–7) 0.18 ± 0.12 | 5.41 ± 3.72 (1–2) 0.08 ± 0.06 | 10.00 ± 6.71 (2–4) 0.30 ± 0.22 | 6.12 ± 3.42 (1–1) 0.06 ± 0.03 |
| Brandesia turgida | 5.00 ± 4.88 (2) 0.10 ± 0.10 | ||||
| Pleurogenes claviger | 1.45 ± 1.44 (1) 0.01 ± 0.01 | 5.00 ± 2.81 (1–4) 0.10 ± 0.07 | 2.70 ± 2.51 (1) 0.03 ± 0.03 | ||
| Pleurogenoides medians | 44.93 ± 5.99 (1–540) 35.58 ± 11.21 | 18.33 ± 4.99 (1–36) 1.00 ± 0.62 | 8.11 ± 4.49 (1–27) 0.78 ± 0.73 | 25.00 ± 9.69 (1–3) 0.50 ± 0.22 | 2.04 ± 2.03 (7) 0.14 ± 0.14 |
| Prosotocus confusus | 39.13 ± 5.87 (1–137) 9.22 ± 2.77 | 6.67 ± 3.22 (1–6) 0.18 ± 0.11 | 13.51 ± 5.62 (1–4) 0.24 ± 0.12 | 10.00 ± 6.71 (1–1) 0.10 ± 0.07 | |
| Diplodiscus subclavatus | 18.84 ± 4.71 (1–8) 0.42 ± 0.14 | 20.00 ± 5.16 (1–4) 0.47 ± 0.13 | 2.70 ± 2.51 (1) 0.03 ± 0.03 | 10.00 ± 6.71 (1–1) 0.10 ± 0.07 | |
| Paralepoderma cloacicola, larvae | 42.03 ± 5.94 (1–35) 4.33 ± 0.99 | 1.67 ± 1.65 (12) 0.20 ± 0.20 | 10.00 ± 6.71 (4–7) 0.55 ± 0.39 | ||
| Strigea strigis, larvae | 2.90 ± 2.02 (1–3) 0.06 ± 0.05 | 3.33 ± 2.32 (4–20) 0.40 ± 0.34 | 2.70 ± 2.51 (5) 0.14 ± 0.14 | 10.00 ± 6.71 (1–5) 0.30 ± 0.25 | 4.08 ± 2.83 (1–3) 0.08 ± 0.06 |
| Strigea sphaerula, larvae | 2.90 ± 2.02 (1–3) 0.06 ± 0.05 | 5.00 ± 2.81 (1–7) 0.17 ± 0.12 | 2.70 ± 2.51 (1) 0.03 ± 0.03 | ||
| Strigea falconis, larvae | 2.70 ± 2.51 (1) 0.03 ± 0.03 | ||||
| Neodiplostomum spathoides, larvae | 1.67 ± 1.65 (15) 0.25 ± 0.25 | ||||
| Tylodelphys excavata, larvae | 1.45 ± 1.44 (3) 0.04 ± 0.04 | ||||
| Alaria alata, larvae | 49.28 ± 6.02 (1–37) 3.65 ± 0.79 | 1.67 ± 1.65 (99) 1.65 ± 1.65 | 8.11 ± 4.49 (1–35) 1.05 ± 0.95 | 10.00 ± 6.71 (9–26) 1.75 ± 1.35 | |
| Oswaldocruzia filiformis | 21.74 ± 4.96 (1–20) 0.65 ± 0.31 | 8.33 ± 3.57 (1–1) 0.08 ± 0.04 | 5.41 ± 3.72 (1–1) 0.05 ± 0.04 | 25.00 ± 9.69 (1–2) 0.40 ± 0.17 | 16.33 ± 5.28 (1–3) 0.20 ± 0.08 |
| Cosmocerca ornata | 11.59 ± 3.85 (1–2) 0.16 ± 0.06 | 5.00 ± 4.88 (1) 0.05 ± 0.05 | 6.12 ± 3.42 (1–7) 0.18 ± 0.14 | ||
| Oxysomatium brevicaudatum | 8.33 ± 3.57 (1–4) 0.13 ± 0.07 | 8.11 ± 4.49 (1–7) 0.24 ± 0.19 | 30.00 ± 10.25 (1–2) 0.40 ± 0.15 | 2.04 ± 2.03 (2) 0.04 ± 0.04 | |
| Icosiella neglecta | 27.54 ± 5.38 (1–21) 1.13 ± 0.39 | 66.67 ± 6.08 (1–20) 3.78 ± 0.61 | 37.84 ± 7.98 (1–18) 1.89 ± 0.60 | 95.00 ± 4.88 (2–22) 8.45 ± 1.26 | 59.18 ± 7.02 (1–10) 2.35 ± 0.42 |
| Total species | 19/5 | 17/5 | 15/4 | 14/3 | 10/1 |
| Trematoda | 16/5 | 14/5 | 12/4 | 10/3 | 6/1 |
| Chromadorea | 3 | 3 | 3 | 4 | 4 |
| Shannon diversity index, H′ | 1.09 | 1.51 | 1.45 | 0.93 | 1.21 |
| Pielou’s evenness index, E | 0.37 | 0.53 | 0.54 | 0.35 | 0.53 |
| Simpson index, 1-D | 0.46 | 0.38 | 0.34 | 0.64 | 0.49 |
| Total examined, specimens | 69 | 60 | 37 | 20 | 49 |
| Helminths Species | Life Cycle | Citation |
|---|---|---|
| Gorgodera cygnoides | Sphaeriidae 1–Corduliidae 3–Ranidae 5 | [50] |
| Gorgodera microovata | unknown | none |
| Gorgodera asiatica | Sphaeriidae 1–Calopterygidae, Corduliidae, Limnephilidae 3–Ranidae 5 | [50] |
| Gorgoderina vitelliloba | Sphaeriidae 1–Ranidae (larvae), Sialidae 3–Anura 5 | [51,52,53] |
| Haematoloechus variegatus | Planorbidae 1–Culicidae, Calopterygidae, Libellulidae 3–Anura 5 | [54,55] |
| Haematoloechus asper | Planorbidae 1–Calopterygidae, Lestidae 3–Anura 5 | [56] |
| Skrjabinoeces similis | Planorbidae 1–Aeshnidae, Calopterygidae, Coenagrionidae, Corduliidae, Lestidae, Libellulidae 3–Ranidae 5 | [57] |
| Opisthioglyphe ranae | Lymnaeidae) 1–Lymnaeidae, Anura (larvae) 3–Anura 5 | [58,59] |
| Brandesia turgida | unknown | none |
| Pleurogenes claviger | Bithyniidae 1–Odonata, Coleoptera, Ephemeroptera, Trichoptera, Sialidae, Gammaridae, Asellidae 3–Anura 5 | [60,61] |
| Pleurogenoides medians | Bithyniidae 1–Odonata, Coleoptera, Ephemeroptera, Trichoptera, Sialidae, Chironomidae, Gammaridae, Asellidae 3–Anura 5 | [60,62] |
| Prosotocus confusus | Bithyniidae 1–Odonata, Coleoptera, Trichoptera, Sialidae, Gammaridae 3–Anura 5 | [60,63] |
| Diplodiscus subclavatus | Planorbidae 1–water–Amphibia 5 | [64,65] |
| Paralepoderma cloacicola, larvae | Planorbidae 1–Anura 3–Colubridae 5 | [66,67] |
| Strigea strigis, larvae | Planorbidae 1–Anura (larvae) 2–Anura 3, 4–Colubridae, Insectivora, Mustelidae, Canidae 4–Strigidae 5 | [68,69,70,71] |
| Strigea sphaerula, larvae | Planorbidae 1–Anura (larvae) 2–Anura 3, 4–Colubridae 4–Corvidae 5 | [68,69,71,72] |
| Strigea falconis, larvae | Planorbidae 1–Anura (larvae) 2–Anura 3, 4–Colubridae, Insectivora, Mustelidae, Canidae 4–Accipitridae 5 | [68,71] |
| Neodiplostomum spathoides, larvae | Planorbidae 1–Anura 3–Colubridae, Corvidae, Laridae, Anatidae 4–Accipitridae 5 | [73,74,75] |
| Tylodelphys excavata, larvae | Planorbidae 1–Anura 3–Ardeidae 5 | [73] |
| Alaria alata, larvae | Planorbidae 1–Anura 2–Ranidae, Colubridae, Viperidae, Corvidae, Laridae, Anatidae, Strigidae, Accipitridae, Rodentia, Insectivora, Carnivora 4–Canidae 5 | [76,77,78] |
| Oswaldocruzia filiformis | soil–Amphibia 5 | [79,80,81] |
| Cosmocerca ornata | water–Anura 5 | [14] |
| Oxysomatium brevicaudatum | soil–Anura 5 | [80,82] |
| Icosiella neglecta | Ceratopogonidae 1–Ranidae 5 | [83,84] |
| Plot 1 | Plot 2 | Plot 3.1 | Plot 3.2 | Plot 3.3 | |
|---|---|---|---|---|---|
| Gorgodera cygnoides | 0.07 | 0.73 | – | – | – |
| Gorgodera microovata | 0.02 | 2.37 | 1.10 | 0.76 | 7.65 |
| Gorgodera asiatica | – | – | 0.55 | – | – |
| Gorgoderina vitelliloba | 0.02 | – | – | – | – |
| Haematoloechus variegatus | 0.16 | 1.83 | 4.95 | 1.51 | 2.94 |
| Haematoloechus asper | 0.07 | – | – | – | – |
| Skrjabinoeces similis | 0.07 | 0.91 | – | – | 1.18 |
| Opisthioglyphe ranae | 13.14 | 2.01 | 1.65 | 2.26 | 1.77 |
| Brandesia turgida | – | – | – | 0.38 | – |
| Pleurogenes claviger | 0.02 | 1.10 | 0.55 | – | – |
| Pleurogenoides medians | 55.61 | 10.95 | 15.93 | 3.77 | 4.12 |
| Prosotocus confusus | 14.41 | 2.01 | 4.95 | 0.76 | – |
| Diplodiscus subclavatus | 0.66 | 5.11 | 0.55 | 0.76 | – |
| Paralepoderma cloacicola, larvae | 6.77 | 2.19 | – | 4.15 | – |
| Strigea strigis, larvae | 0.09 | 4.38 | 2.75 | 2.26 | 2.35 |
| Strigea sphaerula, larvae | 0.09 | 1.83 | 0.55 | – | – |
| Strigea falconis, larvae | – | – | 0.55 | – | – |
| Neodiplostomum spathoides, larvae | – | 2.74 | – | – | – |
| Tylodelphys excavata, larvae | 0.07 | – | – | – | – |
| Alaria alata, larvae | 5.71 | 18.07 | 21.43 | 13.21 | – |
| Oswaldocruzia filiformis | 1.02 | 0.91 | 1.10 | 3.02 | 5.88 |
| Cosmocerca ornata | 0.25 | – | – | 0.38 | 5.29 |
| Oxysomatium brevicaudatum | – | 1.46 | 4.95 | 3.02 | 1.18 |
| Icosiella neglecta | 1.77 | 41.42 | 38.46 | 63.77 | 67.65 |
| Plot 1 | Plot 2 | Plot 3.1 | Plot 3.2 | Plot 3.3 | |
|---|---|---|---|---|---|
| Plot 1 | 857.0 (<0.0001) | 371.0 (<0.0001) | 406.5 (0.0054) | 475.0 (<0.0001) | |
| Plot 2 | 857.0 (<0.0001) | 842.5 (0.0459) | 367.0 (0.0096) | 1095.0 (0.0213) | |
| Plot 3.1 | 371.0 (<0.0001) | 842.5 (0.0459) | 139.0 (0.0001) | 890.0 (0.8877) | |
| Plot 3.2 | 406.5 (0.0054) | 367.0 (0.0096) | 139.0 (0.0001) | 162.5 (<0.0001) | |
| Plot 3.3 | 475.0 (<0.0001) | 1095.0 (0.0213) | 890.0 (0.8877) | 162.5 (<0.0001) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chikhlyaev, I.V.; Ruchin, A.B. Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies. Diversity 2022, 14, 247. https://doi.org/10.3390/d14040247
Chikhlyaev IV, Ruchin AB. Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies. Diversity. 2022; 14(4):247. https://doi.org/10.3390/d14040247
Chicago/Turabian StyleChikhlyaev, Igor V., and Alexander B. Ruchin. 2022. "Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies" Diversity 14, no. 4: 247. https://doi.org/10.3390/d14040247
APA StyleChikhlyaev, I. V., & Ruchin, A. B. (2022). Ecological Analysis and Biodiversity of the Helminth Community of the Pool Frog Pelophylax lessonae (Amphibia: Anura) from Floodplain and Forest Water Bodies. Diversity, 14(4), 247. https://doi.org/10.3390/d14040247