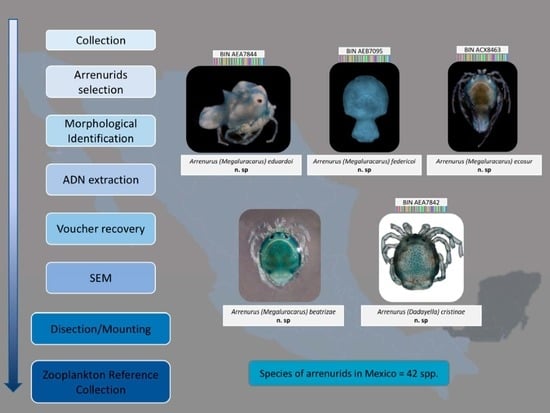
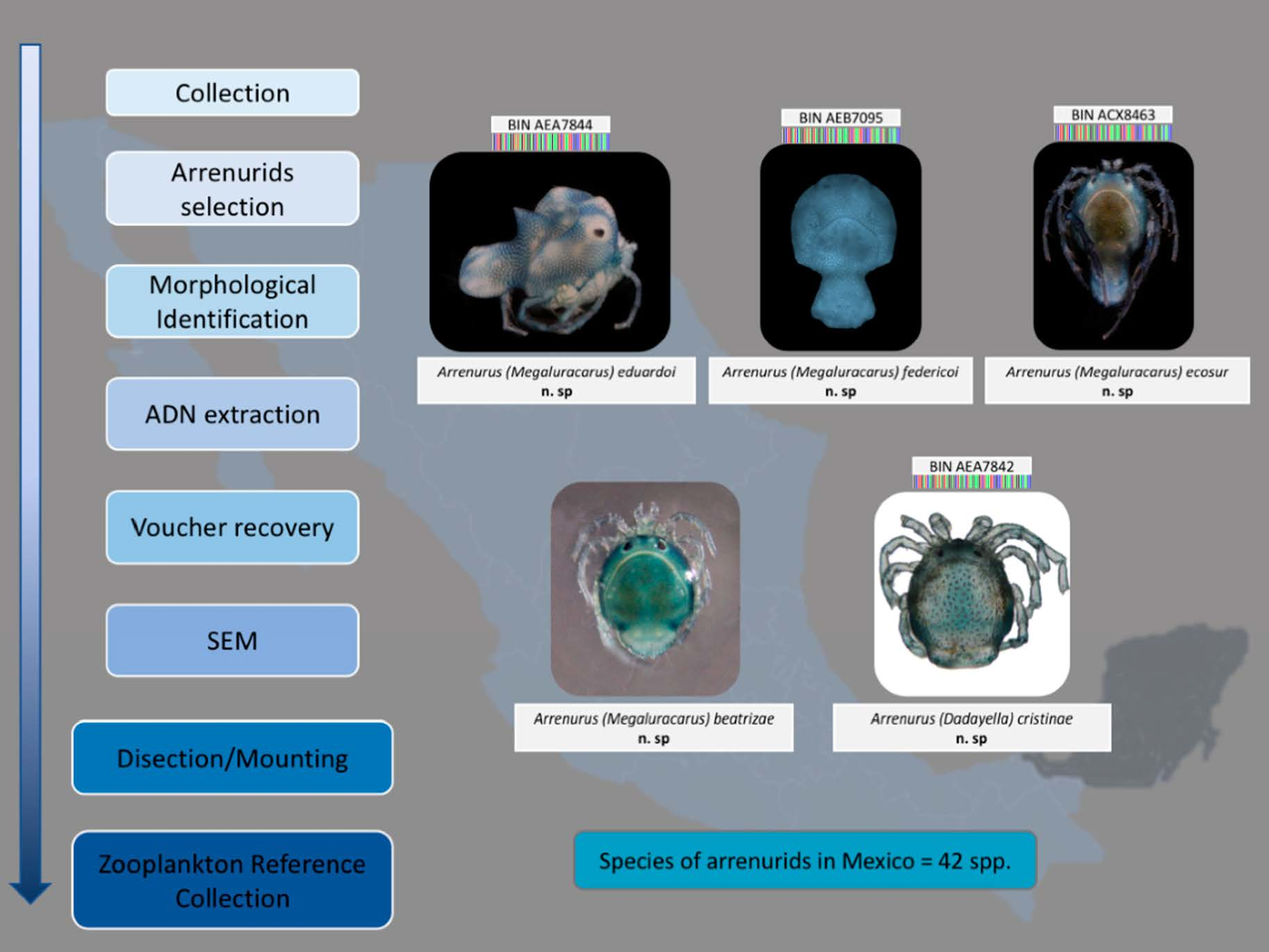
Systematic Part
Family Arrenuridae (Thor, 1900)
Genus Arrenurus (Dugés, 1834)
Subgenus Arrhenuropsis (Viets, 1954)
Arrenurus (?
Arrhenuropsis)
mexicanus (Cramer and Cook, 1992), (
Figure 3).
Material examined: One male from Ramonal pond (access number: ECO-CH-Z-10608), Quintana Roo, 19°23′31″ N, −82°37′27″ W; emergent vegetation, 14 April 2019.
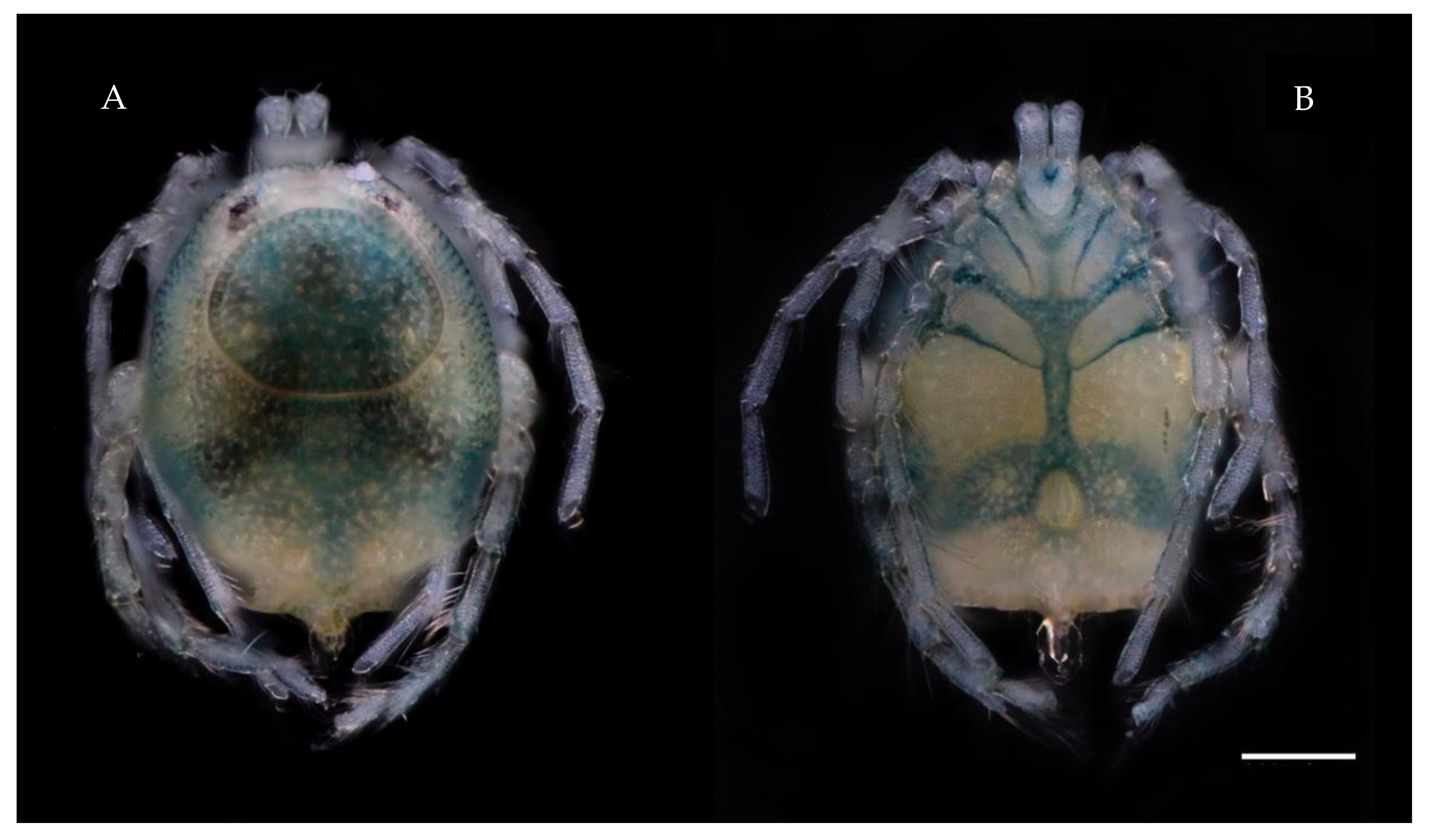
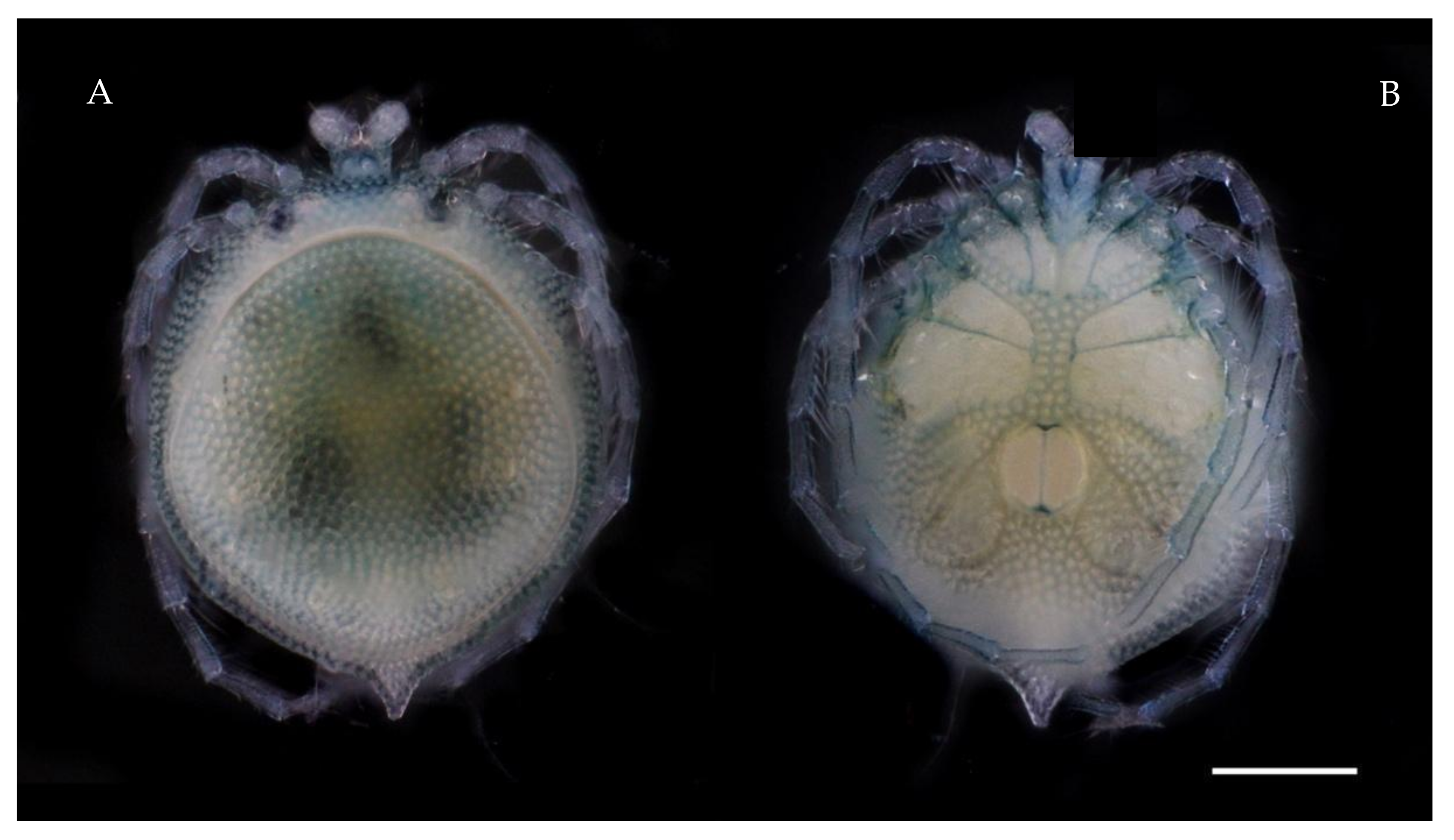
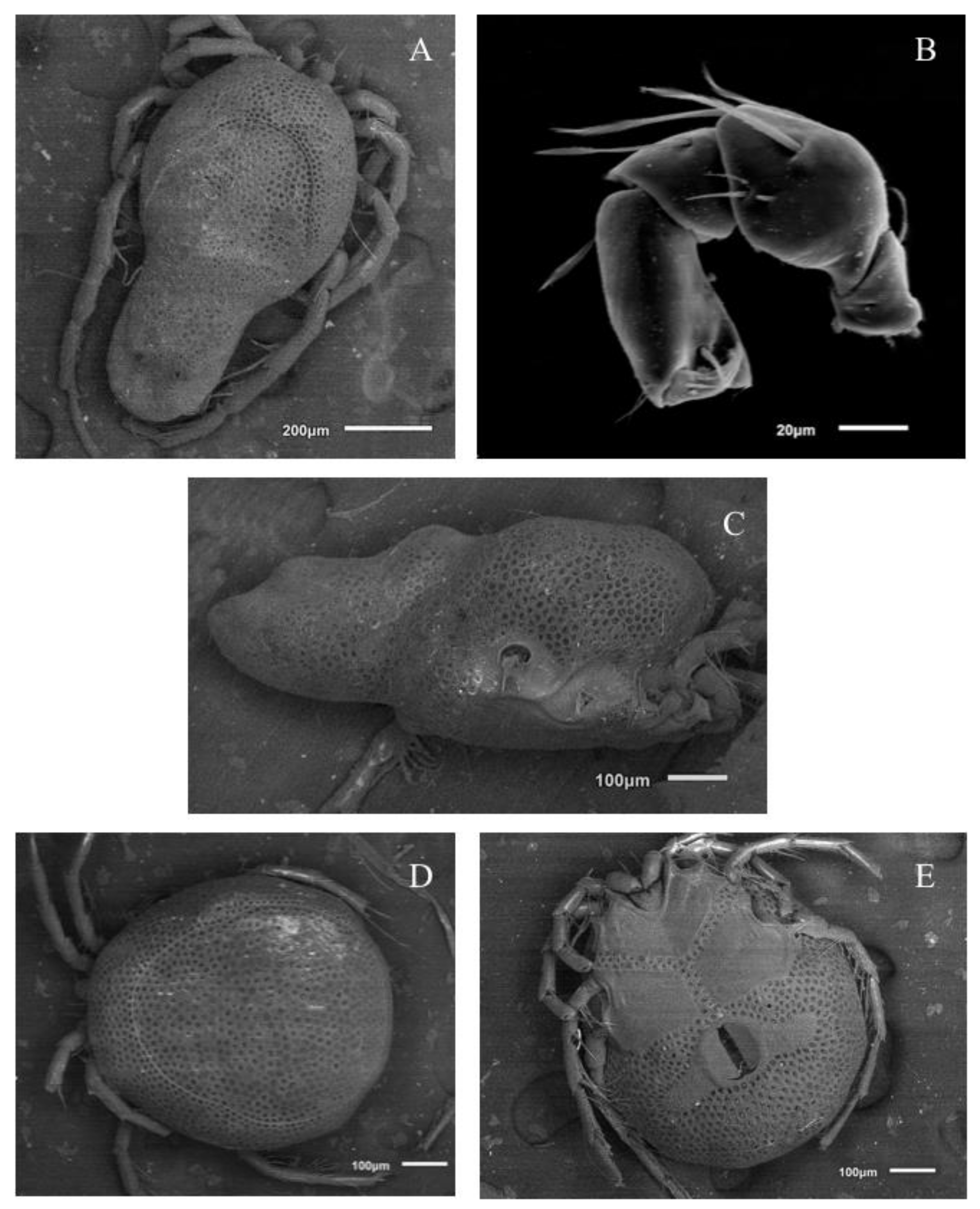
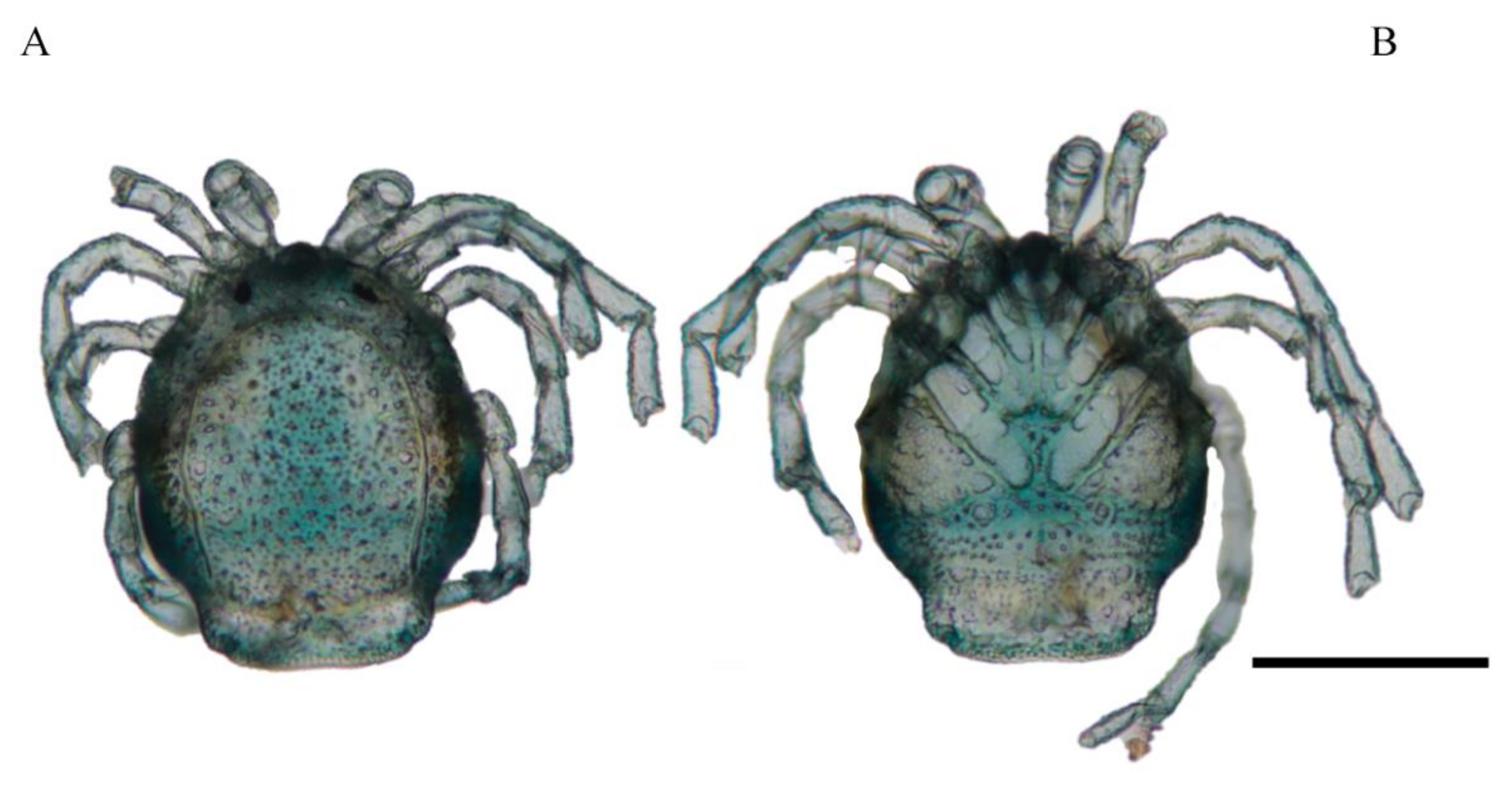
Description. MALE: Idiosoma blue-greenish with white areas in the Dgl 1–4 regions, 799 L without petiole and 493 W. Dorsal shield small, oval, and located in the anterior part of the dorsum, 296 L and 345 W (
Figure 3A). Genital field 394 W, gonopore 69 L and 48 W (
Figure 3B). Dorsal L of palpal segments L: P1: 27; P2: 74; P3: 29; P4: 84; P5: 84. Dorsal L of fourth leg segments: IV-Leg-1: 32; IV-Leg-2: 104; IV-Leg-3: 101; IV-Leg-4: 148; IV-Leg-5: 151; IV-Leg-6: 109.
Remarks. Male and female were described by Cramer and Cook [
4]. Therefore, we only give some diagnostic measurements. This record represents the second of this species for the country.
Distribution. Previously known from the Champayan lagoon, Altamira, Tamaulipas state, 22°22′49″ N, −97°58′34″ W (Mexico).
Material examined: One female from Ramonal pond (access number: ECO-CH-Z-10609), Quintana Roo state, 19°23′31″ N, −82°37′27″ W; emergent vegetation, 14 April 2019.
Description. FEMALE: Idiosoma bluish-green with white areas in the Dgl 1–4 regions, 680 L and 552 W; dorsal furrow complete, dorsal shield 512 L and 483 W (
Figure 4A). Genital field 305 W, gonopore 99 L and 116 W (
Figure 4B). Dorsal L of fourth leg segments: IV-Leg-1: 74; IV-Leg-2: 101; IV-Leg-3: 99; IV-Leg-4: 119; IV-Leg-5: 106; IV-Leg-6: 116.
Sequence: ATCTATGATACCATTGGAACAGCCCATGCTTTGATTATAATTTTCTTTATAGTCATACCCATCATAATTGGAGGATTTGGAAATTGACTCGTTCCCCTCATGTTAGAAGCTCCAGATATAGCATTCCCACGAATAAATAACATAAGATTTTGATTACTTCCACCCCCCTTAACACTCCTTCTATCAAGATCATTAACTTCTTCAGGAGCAGGAACTGGATGAACAGTTTACCCTCCTTTATCAAGAAATATCGCCCATGGAGGACCTTCAGTAGACCTAGCAATTTTCTCCCTACACCTTGCAGGTGTGTCCTCAATTTTAGGAGCAATTAACTTCTTGGCTACATTTATAAACATAAAACCTAAACATATAAAATATGACCGAATCCCCCTATTTGTAATTTCTATTTTTATCACCGTAATCCTCCTTCTTCTTTCCCTCCCCGTATTAGCTGGAGCCATTACTATACTTCTTACTGATCGAAATTTTAATACTTCATTTTTTGACCCGGCGGGGGGAGGAGATCCCATCCTTTACCAACATCTATTT.
Remarks. Female and male were described by Cramer and Cook [
4]. We provide some additional measurement data. The chaetotaxy of the palp and IV-Leg-5–6, as well as the position of ventral and dorsal glandularia, agree with the original description. The only noticeable difference is that the first and second coxae tips extend slightly beyond the body proper in our specimen. The associated sequence was obtained, representing a unique BIN (BOLD: AEA8234).
Distribution. Previously known from the Champayan lagoon, Altamira, Tamaulipas state (Mexico). This record represents the second of this species for the country.
Material examined: One male, one female, and one nymph from Silvituc lagoon (access number: ECO-CH-Z-10610-10611), Escarcega municipality, Campeche state, 18°37′26″ N, −90°17′5.9″ W, 18 March 2020.
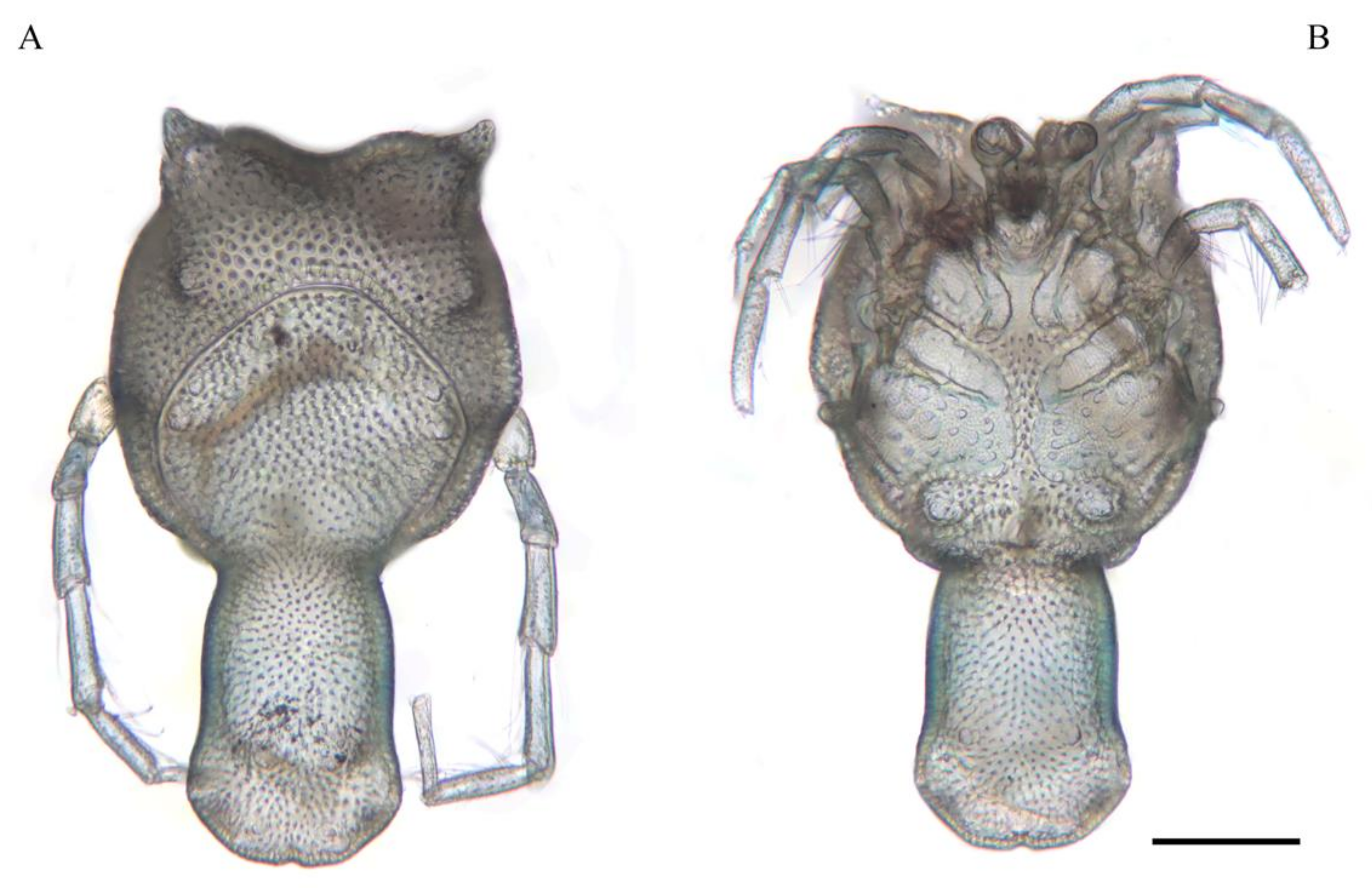
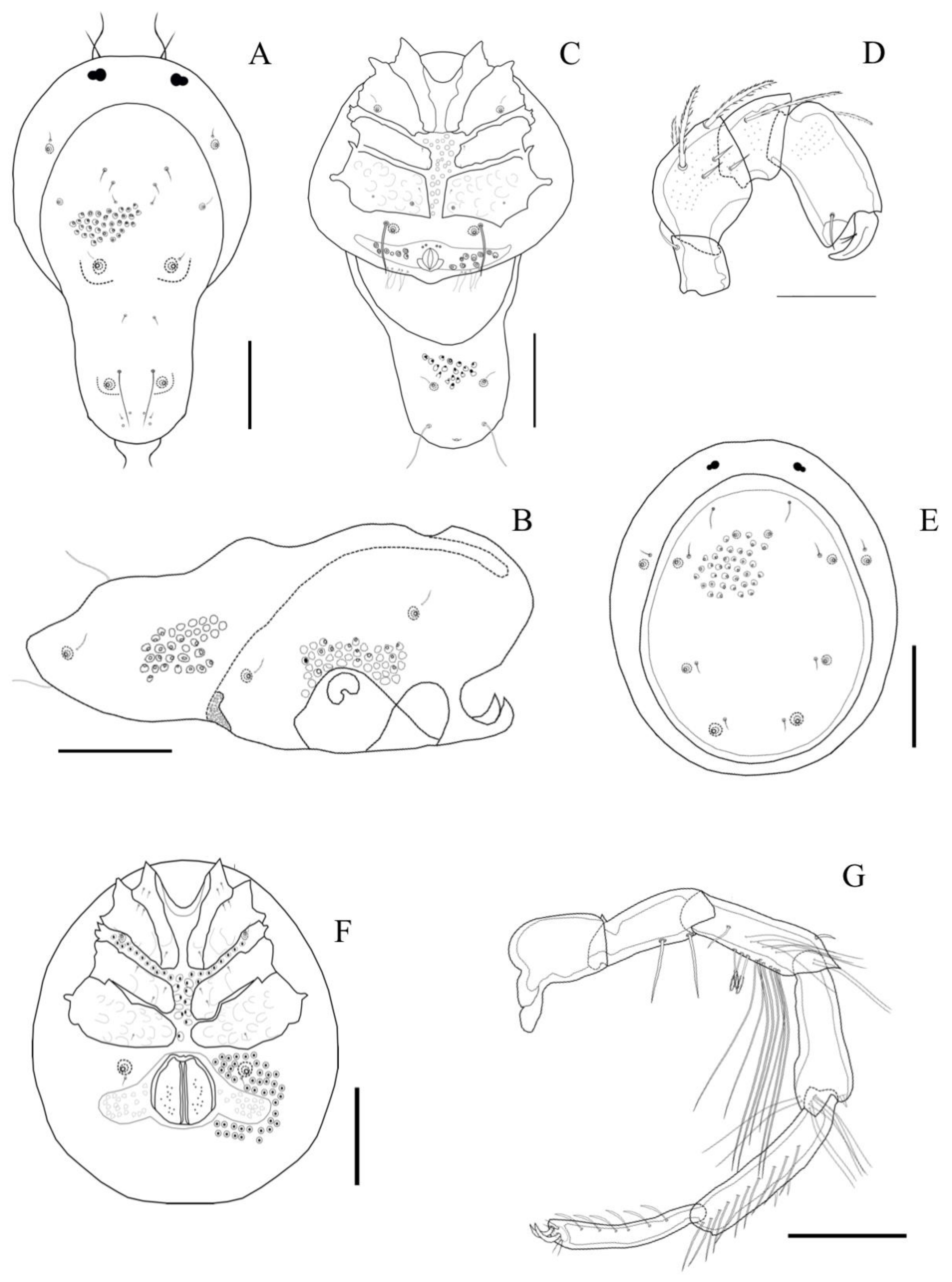
Description. MALE: Idiosoma light-bluish, 962 L (including cauda) and 560 W; dorsal shield 776 L (including cauda) and 422 W (
Figure 5A). Genital field 281 W, gonopore 47 L and 61 W (
Figure 5B). Dorsal L of palpal segments: P1: 34; P2: 63; P3: 33; P4: 66; P5: 47. Dorsal L of fourth leg segments: IV-Leg-1: 86; IV-Leg-2: 128; IV-Leg-3: 151; IV-Leg-4: 178; IV-Leg-5: 165; IV-Leg-6: 138.
FEMALE: Idiosoma light bluish, 986 L and 907 W; dorsal furrow complete, dorsal shield 719 L and 680 W. Genital field 454 W, gonopore 138 L and 140 W.
Consensus sequence: ACATTATACTTCGCATTCGGAGCTTGATCGGGTATAGTAGGAGCAAGACTTAGAAGTCTAATCCGACTAGAATTAGGGCAACCAGGAAGACTTTTAGGAAATGATCAAATTTACAACACCATTGTTACAGCGCATGCTTTCATTATAATCTTCTTTATAGTTATACCAATTATAATCGGAGGATTCGGAAACTGATTAGTACCCCTAATACTAGCCGCCCCTGATATGGCATTCCCACGAATAAATAATATAAGATTCTGACTTCTACCGCCAGCCTTAACACTTCTTTTATCAAGATCGTTAACTTCAGTAGGAGCAGGAACCGGATGAACAGTCTACCCTCCCCTATCCAGAAACATTGCACATGGTGGACCTTCAGTAGATATAGCTATCTTCTCATTACATTTAGCAGGAGTCTCCTCAATTTTAGGAGCTATCAATTTTCTAGCTACAATTTTAAATATAAAGCCTAAACATATAAAATATGACAGAATTCCATTATTTGTAGTTTCAATTTTTATTACAGTAATTCTTCTTTTACTTTCACTGCCTGTATTAGCAGGAGCTATTACTATACTTCTTACAGATCGAAATTTTAACACCTCTTTCTTCGATCCAGCTGGAGGAGGAGATCCTATTTTATACCAA.
Remarks. Our specimens agree with the descriptions given by Marshall (1903) and Wilson (1961). According to Cook [
20], the status of
A. marshallae is complex because the species is a member of a closely related group characterized by the possession of a long cauda and horn-like projections over the eyes. It can be separated from other species (
A. megalurus megalurus,
A. megalurus intermedius) by the slightly indented posterior end of the cauda.
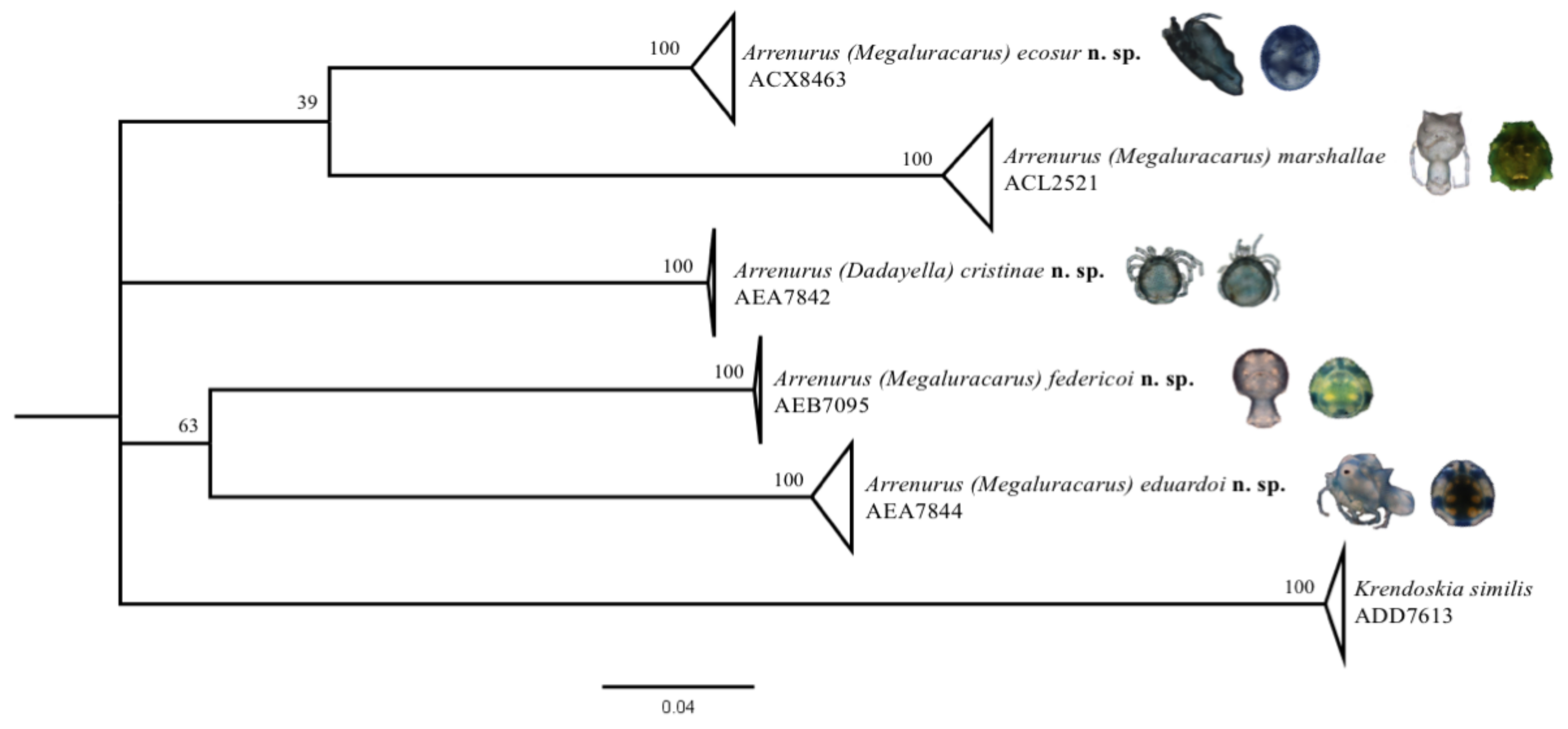
The sequences obtained in this study with the BIN ACL2521 agree with another 14 public sequences from
A. marshallae. Some of these were identified morphologically by Bruce Smith. Morphological and molecular identification agree (
Figure 2). These public sequences in the BOLD database integrated with the morphology make it possible to verify the records of putative
A. marshallae in other localities and other members of this complex group.
Distribution. Previously known from the United States and Canada. This record constitutes the first for Mexico.
Holotype: Male from Acapulquito stream, Rivera del Río hondo, Othon P. Blanco municipality, Quintana Roo state (access number: ECO-CH-Z-10612), 18° 25′ 55″ N, 88° 31′51″ W; emergent vegetation and submerged roots, 11 April 2019, coll. L. Montes.
Paratypes: Two females and two males, same data as holotype (access number: ECO-CH-Z-10613-10614).
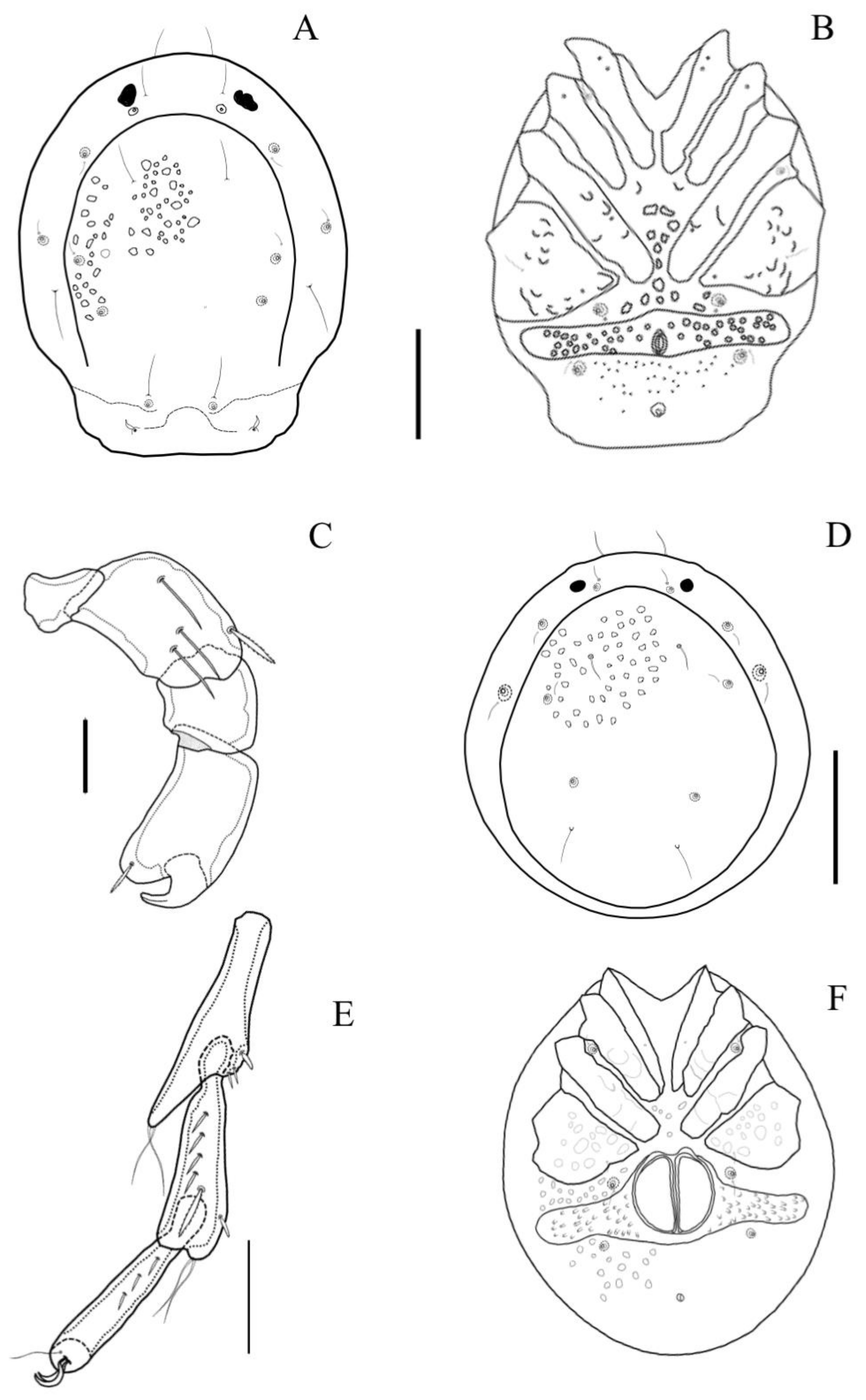
Diagnosis. Male with a large, thorn-shaped hump in the middle of the dorsal shield (
Figure 6C,D), falcate setae on Dgl-2 and Dgl-3 (
Figure 7A); three pinnate setae on P2 (two in anterolateral position and one in anteromedial position) and one falcate seta on medial position on P3. Bipectinate setae on all lateral IV-Leg-3 segment and serrate setae in the anterolateral position of IV-Leg-2 segment (
Figure 7G).
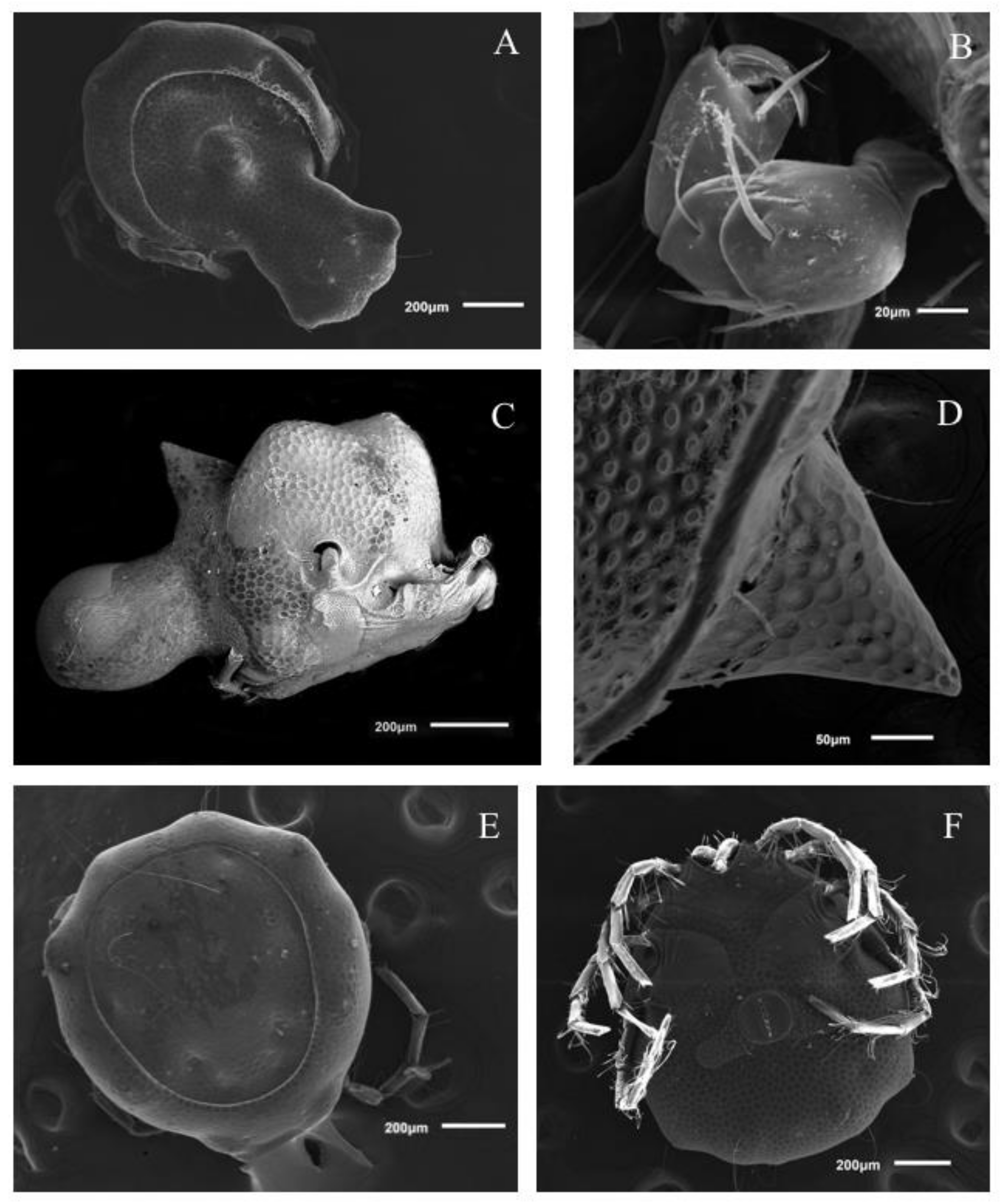
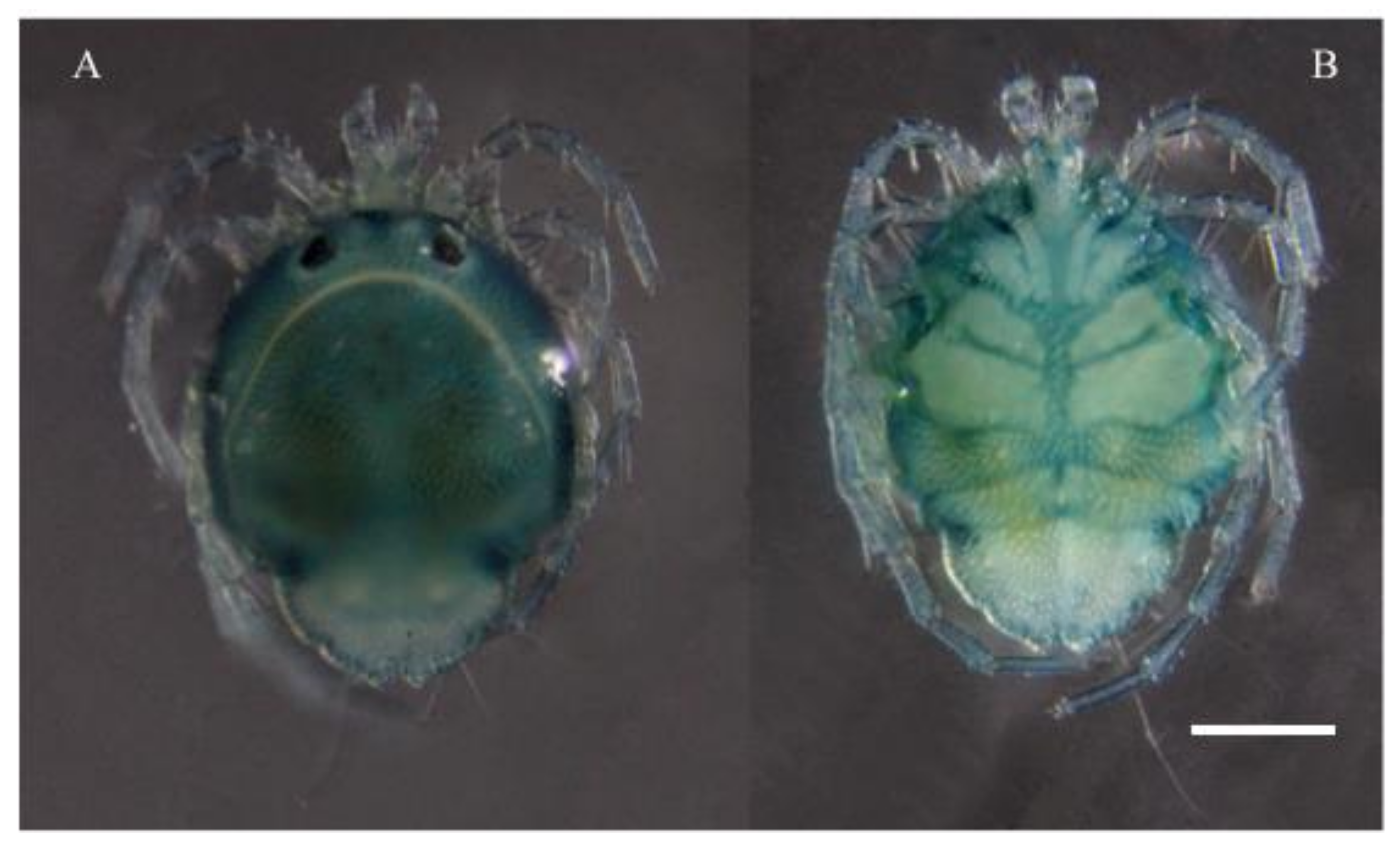
Description. MALE: Idiosoma bluish with yellow spots in the Dgl-1–4 regions (
Figure 7A), 1178 L and 785 W; anterior part of idiosoma very wide (
Figure 6 and
Figure 7A). Dgl-2 and Dgl-3 setae falcate. Dorsal shield 1000 L (cauda included dorsal portion), 571 W. Cauda long, representing almost half of the total body length, 470 L and 478 W, small humps in Lgl-4 region. Dorsal furrow complete, passing ventrally at base of cauda and continuing immediately posterior to the acetabular plates. In lateral view, there is a large, thorn-shaped hump centrally on the dorsum (212 height) (
Figure 6C,D). Anterior and posterior coxal groups separated, Cxgl-1 between Cx-II and Cx-III, Cx-IV laterally slightly extending beyond the idiosoma, posterior region concave. Cxgl-2 is located between Cx-IV and the acetabular plates (
Figure 7C). Genital field 457 W, gonopore 113 L and 102 W, acetabular plate extending laterally from the gonopore region with two setae posterior to each plate (
Figure 7C). Dorsal L of palpal segments: P1: 21; P2: 73; P3: 47; P4: 79; P5: 47; P3 with a long, falcate seta on anterolateral position (
Figure 6B D). Dorsal L of fourth leg segments: IV-Leg-1 120; IV-Leg-2: 155; IV-Leg-3: 196; IV-Leg-4: 210; IV-Leg-5: 189; IV-Leg-6: 172; IV-Leg-2 with three serrate setae in anterolateral position and IV-Leg-3 with ten bipectinate setae on IV-Leg-4 and IV-Leg-5 with 10 and 11 small, pinnate setae, respectively (
Figure 7G). IV-Leg-2–6 bear numerous swimming setae.
FEMALE: Idiosoma 1000 L and 948 W, dorsal shield 800 L and 680 W, bears the postocularia and four pairs of glandularia. Anterior idiosoma margin rounded with distinctive posterolateral projections (
Figure 6E and
Figure 7F). Acetabular plates wing-shaped, laterally directed, narrow and slightly bowed. Genital field 514 W, gonopore 182 L and 187 W. Anterior and posterior coxal group separated, Cx-I and Cx-II extending beyond the anterior margin of idiosoma (
Figure 6 and
Figure 7F). Idiosoma and legs are bluish with yellow areas on Dgl-1–4 regions (
Figure 7E).
Consensus sequence: ACTCTATACTTCGCTTTTGGCGCTTGATCAGGCATAATCGGAGCTAGCCTTAGAAGTCTTATCCGTTTAGAACTTGGACAACCTGGTAATCTTTTAGGAAACGATCAAATATACAACACAATTGTCACTGCCCACGCATTTGTTATAATCTTTTTCATAGTTATGCCAATCATAATCGGAGGATTCGGAAACTGATTAGTTCCTATTATACTAGCAGCCCCAGATATAGCTTTCCCACGAATAAATAATATAAGATTTTGACTTCTTCCCCCCGCTTTAACTCTTCTACTNTCAAGATCTCTATCTTCTTCAGGAGCGGGGACTGGCTGAACAGTTTACCCCCCTCTTTCNAGTAATATCGCTCACGGAGGACCATCTGTCGATATAGCTATTTTCTCACTTCACTTAGCAGGAGTTTCGTCTATTCTAGGGGCAATTAACTTCTTAGCCACAACTATAAACATAAAGCCAAAATATATAAAATATGACCGAATCCCCTTATTTGTAGTCTCAATTTTCATCACAGTCATTCTCCTCCTCTTATCATTACCAGTCTTAGCTGGAGCTATCACAATACTATTAACTGATCGAAACTTTAACACATCATTCTTTGACCCTGCCGGAGGGGGAGACCCAATTCTTTACCAA.
Etymology. This species is named after Eduardo Montes, brother of the first author, for his empathy, solidarity, and for the lovingly provided support.
Remarks.
A. (
Megaluracarus)
eduardoi n. sp. is similar to
A. campechensis (Cook, 1980) and
A. maya (Ramírez-Sánchez and Rivas, 2013) in terms of overall shape and sturdy idiosoma. However, males of
A. eduardoi n. sp. present a distinctive, large, thorn-shaped hump in the middle of the dorsal shield (in lateral view) that easily separates this species from the latter two. This hump resembles
A. gibberifer (Viets, 1933), originally described from Uruguay. Nevertheless, the shape of both species is quite different, especially in the dorsal view of cauda;
A. eduardoi n. sp. presents a trapezoidal shape (
Figure 6A), while
A. gibberifer has a quadrangular shape. Additionally, the reported size for
A. gibberifer is much smaller (742 L and 528 W) than that registered for
A. eduardoi n. sp. Furthermore, the palp chaetotaxy of these two species is distinct. The BOLD database assigned the unique BIN AEA7844 (
Table 1), used to pair the sexes. The result of the ML tree (
Figure 2) and the NJ tree (
Supplementary File S1) separates
A. eduardoi n. sp. from the others registered in the database and strongly supports the status of these new species.
Distribution. So far only known from the type locality, Acapulquito stream, Riviera del Río Hondo, Quintana Roo (Mexico).
Type material. Holotype: Male from Acapulquito stream, Riviera del Río Hondo, Othon P. Blanco, Quintana Roo (access number: ECO-CH-Z-10615), 18°25′55″ N, 88°31′51″ W; emergent vegetation and submerged roots, 11 April 2019, coll. L. Montes.
Paratypes: Two females and one male, same data as holotype (access number: ECO-CH-Z-10616-10617).
Diagnosis. Pores huge (on the idiosoma as well as the legs and palps), Dgl-1 and Cxgl-2 on distinct humps in males. Numerous setae surround the acetabular field in both sexes.
Description. MALE: Idiosoma 1037 L and 693 W, uniformly bluish with large pores. The anterior part of the idiosoma is wide, with noticeable humps in the Dgl-1 area which are visible in the lateral view (
Figure 8C and
Figure 9B). Dorsal shield 718 L (cauda included dorsal portion) and 436 W. Cauda of medium length, representing a third of the total length of the body, 365 L and 394 W (
Figure 8 and
Figure 9A), with lobes posterolaterally directed and Lgl-4 on small humps. Dorsal furrow complete, passing ventrally at base of cauda and continuing immediately posterior to the acetabular plates. In lateral view, a big hump is visible in the anterior part of the idiosoma in the Dgl-1 region (
Figure 8C and
Figure 9B). Coxae with a porous surface, anterior and posterior coxal groups separated, Cxgl-1 located in the middle of Cx-II and Cx-III; Cx-II and Cx-IV slightly extending beyond the anterolateral margin of the idiosoma; Cx-III slightly overlapping Cx-IV (
Figure 8B and
Figure 9C). Cxgl-2 is located between Cx-IV and the acetabular plates. Genital field 403 W, gonopore 102 L and 75 W. Acetabular plates extending laterally from the gonopore and surrounded by numerous setae (anterior ones small, 24 L, posterior ones longer, 82 L) (
Figure 8D and
Figure 9C). Dorsal L of palpal segments: P1: 37, P2: 63, P3: 41, P4: 63, P5: 38 (
Figure 9D). Dorsal L of fourth leg segments: IV-Leg-1: 125, IV-Leg-2: 165, IV-Leg-3: 209, IV-Leg-4: 159, IV-Leg-5: 193, IV-Leg-6: 165: IV-Leg-5 bears six swimming setae, IV-Leg-4 distal process bears nine short swimming setae, IV-Leg-3 bears 12 swimming setae, both IV-Leg-2 and IV-Leg-3 bear three tiny, spine-like setae on lateral surface (
Figure 9G).
FEMALE: Idiosoma bearing huge pores, bluish with yellow spots on the region of Dgl-1–4 and eyes (
Figure 9E), 1170 L and 1066 W, dorsal shield 714 L and 790 W, bears the postocularia and three pairs of glandularia. Idiosoma rounded in the anterior margin and with posterolateral lobes (
Figure 8F and
Figure 9E). Acetabular plates curved and anterolaterally directed, narrow in telation to the gonopore length. Genital field 499 W surrounded by small setae (38–52 L), gonopore 190 L and 204 W. (
Figure 8E). The anterior and posterior coxal groups separated, Cx-II and Cx-IV extending slightly beyond the margin of the idiosoma (
Figure 8 and
Figure 9G).
Consensus sequence: CTCTATTTCGCCTTTGGAGCATGATCAGGAATAATTGGAGCAAGATTAAGAAGTCTAATTCGCTTAGAACTAGGACAACCAGGAAGACTATTAGGAAACGATCAAATTTATAACACTATTGTTACAGCTCATGCATTCATCATAATTTTCTTCATAGTAATACCTATCATAATCGGAGGATTTGGAAATTGATTAGTACCATTAATACTAGCTGCTCCAGATATAGCTTTTCCTCGAATAAATAATATAAGATTTTGACTATTACCCCCAGCATTATCCCTTCTACTAGCAAGCTCCCTTTCTTCATCAGGAGCAGGAACAGGATGAACAGTTTACCCCCCATTATCAAGAAATATCGCACACGGAGGACCTTCAGTTGATATAGCTATCTTCTCACTCCACCTAGCAGGAGTATCTTCAATTCTAGGAGCCATCAATTTTCTAGCAACAATCATAAATATAAAACCTAAATACATAAAATATGATCGAATCCCTTTATTTGTTATCTCTATCTTTATCACAGTAATCTTACTCTTATTATCCTTACCAGTTTTAGCTGGAGCTATCACTATACTATTAACAGATCGAAACTTTAATACATCATTCTTCGACCCAGCAGGAGGAGGAGATCCTATCCTATACCAACAT.
Etymology. This species is named after Federico Montes, father of the first author, in the form of gratitude for bringing her closer to science since childhood.
Remarks.
Arrenurus (
Megaluracarus)
federicoi n. sp. is similar to
A. maya (Ramírez-Sánchez and Rivas, 2013), described from a cenote in Yucatan, in the shape of idiosoma, the pattern of dorsoglandularia position, and in the presence of setae surrounding the genital field. The significant difference is in the palp chaetotaxy;
A. maya presents three long, thickened setae while
A. federicoi n. sp. does not. In IV-Leg-6,
A. federicoi n. sp. presents four spine-like setae while
A. maya presents ten. Furthermore,
A. maya has very small, Dgl-4 associated setae, while
A. federicoi n. sp. Dgl-4 associated setae are at least four times longer than in
A. maya (
Figure 9A). Additionally, the cauda in
A. maya is more slender than in
A. federicoi n. sp. Both
A. catoi (Cramer and Cook, 1992) and
A. campechensis (Cook, 1980) are similar to the new species in the shape of the anterior idiosoma in dorsal view and Dgl-1 over humps. However,
Arrenurus federicoi n. sp. can be separated from both latter species by the chaetotaxy of the palp, IV-Leg, the distinctive shape of cauda in dorsal view, and especially the integument with large pores. The BOLD database assigned the unique BIN AEB7095 (
Table 1), used to pair the sexes. The ML tree (
Figure 2) and the NJ tree (
Supplementary Material) separate
A. federicoi n. sp. from the others registered in the database and strongly support the status of these new species.
Distribution. So far only known from the type locality, Acapulquito stream, Riviera del Río Hondo, Quintana Roo (Mexico).
Holotype: Male from Mis Casas, Bacalar lagoon, Bacalar, Quintana Roo (access number: ECO-CH-Z-10618), 18°25′55 N, 88°31′51 W; littoral, emergent vegetation, 14 April 2019, coll. L. Montes.
Paratypes: Three males and one female, with same data as the holotype. Six females and one male from Chichancanab lagoon, José María Morelos, Quintana Roo (access number: ECO-CH-Z-10619-10622), 19°55′26 N, 88°36′14 W.
Diagnosis. Male with cauda of moderate length (330) with Dgl-3 and Dgl-4 on distinct humps. P2 with three long, pinnate setae laterally and three medial, short, spine-like setae in the posterior margin; P3 with one thin and long, pinnate seta lateromedially situated; IV-Leg-3 with three pilose setae lateromedially situated. Considerably long setae of Cxgl-2.
Description. MALE: Idiosoma 864 L and 483 W, light blue, some specimens with purple legs. Dorsal shield 729 L (including cauda) and 374 W. Dorsal furrow complete, continuing posterior to genital field. The non-caudal portion of the dorsal shield bearing two pairs of glandularia, Dgl-3 on a hump, each one. (
Figure 10C and
Figure 11B). Cauda is relatively short, representing one-third of the total length of idiosoma, with a rounded posterior margin. Dgl-4 on small humps. In lateral view, the base of the cauda is thicker than the anterior idiosoma (
Figure 10C and
Figure 11B). Anterior and posterior coxal groups separated. Cx-I and Cx-II extend slightly beyond the idiosoma margin. Cxgl-2 between Cx-IV and the acetabular plates, with the associated setae considerably long (146 L) (
Figure 11C). Genital field 293 W, gonopore 58 L and 56 W. Acetabular plates extending laterally from the gonopore region with numerous long (50 L) setae along their posterior margin (
Figure 11C). Dorsal L of palpal segments: P1: 29; P2: 58; P3: 31; P4: 62; P5: 25 (
Figure 10B and
Figure 11D). Dorsal L of fourth leg segments: IV-Leg-1: 151, IV-Leg-2: 119, IV-Leg-3: 112, IV-Leg-4: 135, IV-Leg-5: 154, IV-Leg-6: 109. IV-Leg-3 bears eight swimming setae, three small, pilose setae, and six medium-length, swimming setae on the dorsal surface.
Female: Idiosoma oval, uniformly bluish, 655 L and 590 W, with the postocularia and four pairs of glandularia, dorsal shield 773 L and 716 W (
Figure 10D and
Figure 11E). Acetabular plates wing-shaped, laterally directed, narrow in relation to gonopore length. Genital field 378 W, gonopore 138 L and 141 W. Anterior and posterior coxal groups separated, Cx-I slightly reaching the margin of the ventral shield (
Figure 10E and
Figure 11F).
Consensus sequence: ACACTTTATTTTGCATTTGGAGCTTGATCAGGTATAGTAGGAGCTAGACTAAGAAGTCTAATTCGCCTAGAACTAGGACAACCAGGAAATCTTTTAGGAAACGATCAAATTTACAACACAATTGTAACAGCTCACGCTTTTATTATAATCTTTTTCATAGTTATACCAATCATAATCGGAGGATTCGGAAACTGACTAGTTCCATTAATACTAGCAGCCCCAGACATAGCGTTCCCACGAATAAACAATATAAGATTCTGACTTTTACCACCTGCCCTTACACTCCTACTATCTAGATCACTATCATCCACTGGAGCAGGAACAGGGTGAACTGTTTATCCACCCCTTTCAAGAAACATTGCCCATGGAGGACCGTCAGTAGACATAGCAATCTTCTCACTACACTTAGCAGGTGTGTCATCAATTTTAGGAGCTATCAACTTTTTAGCCACAATCATAAACATAAAACCTAAACACATAAAATACGATCGAATTCCCCTTTTTGTTGTATCAATTTTTATTACTGTTATCCTACTTCTTCTCTCACTTCCAGTTTTAGCAGGAGCTATTACAATGCTACTAACAGATCGAAATTTCAATACATCATTCTTTGACCCAGCCGGGGGGGGAGACCCTATCTTATACCAA.
Etymology. This species is named in honor of El Colegio de la Frontera Sur (ECOSUR), the research center where the first author completed her graduate studies.
Remarks. Arrenurus ecosur n. sp. is similar to A. tabascoensis (Cook, 1980) and A. birgei (Marshall, 1903), both known from Tabasco (Mexico), mainly in the distinct hump in the area of Dgl-3 (when viewed laterally). However, the cauda of the new species is slightly tapering, contrary to A. tabascoenis. Moreover, the posterior margin of the cauda is convex in A. ecosur n. sp. and straight in A. tabascoensis. Aditionally the principal difference among these species is the chaetotaxy of the palps. The new species presents three distinct, pinnate setae on P2. Arrenurus ecosur n. sp. is also similar to A. urbanus (Ramírez-Sánchez and Rivas, 2013) in the overall shape of the idiosoma in the lateral and dorsal view. Nevertheless, the cauda of the new species is longer and thinner.
Additionally,
A. urbanus possesses a characteristic patch of two types of seta medially on P2, which are absent in
A. ecosur n. sp. The BOLD database assigned the BIN ACX8463 (
Table 1), which was used to pair the sexes. The result of the ML tree (
Figure 2) and the NJ tree (
Supplementary Material) separate
A. eduardoi n. sp. from the others registered in the database and support the status of this new species.
Distribution. Wide regional distribution in the Yucatan Peninsula: Bacalar lagoon, Chichancanab lagoon, Muyil lagoon, Cenote Azul, Cenote Chancah Veracruz, Cenote Sijil Noh Ha, and Cenote del Padre, Quintana Roo (
Table 1).
Holotype: One male from Ramonal wetland, Quintana Roo (access number: ECO-CH-Z-10623), 19°23′31″ N, –82°37′27″ W, emergent vegetation, 14 April 2019. Coll. L. Montes and T. Goldschmidt.
Paratypes: Three males, one with the same data as the holotype (access number: ECO-CH-Z-10624), the other two from San Pedrito lagoon, Pantanos de Centla, Tabasco (access number: AAL00273, AAL00274), 18°21′58.7″ N, −92°36′03.6″ W, 6 February 2002. Coll. M. Ramírez-Sánchez.
Diagnosis. Characteristic short cauda with two pairs of lateral notches, tips of Cx-II significantly protruding beyond the anterior margin of the idiosoma, P3 presents a long, pinnate seta located medially.
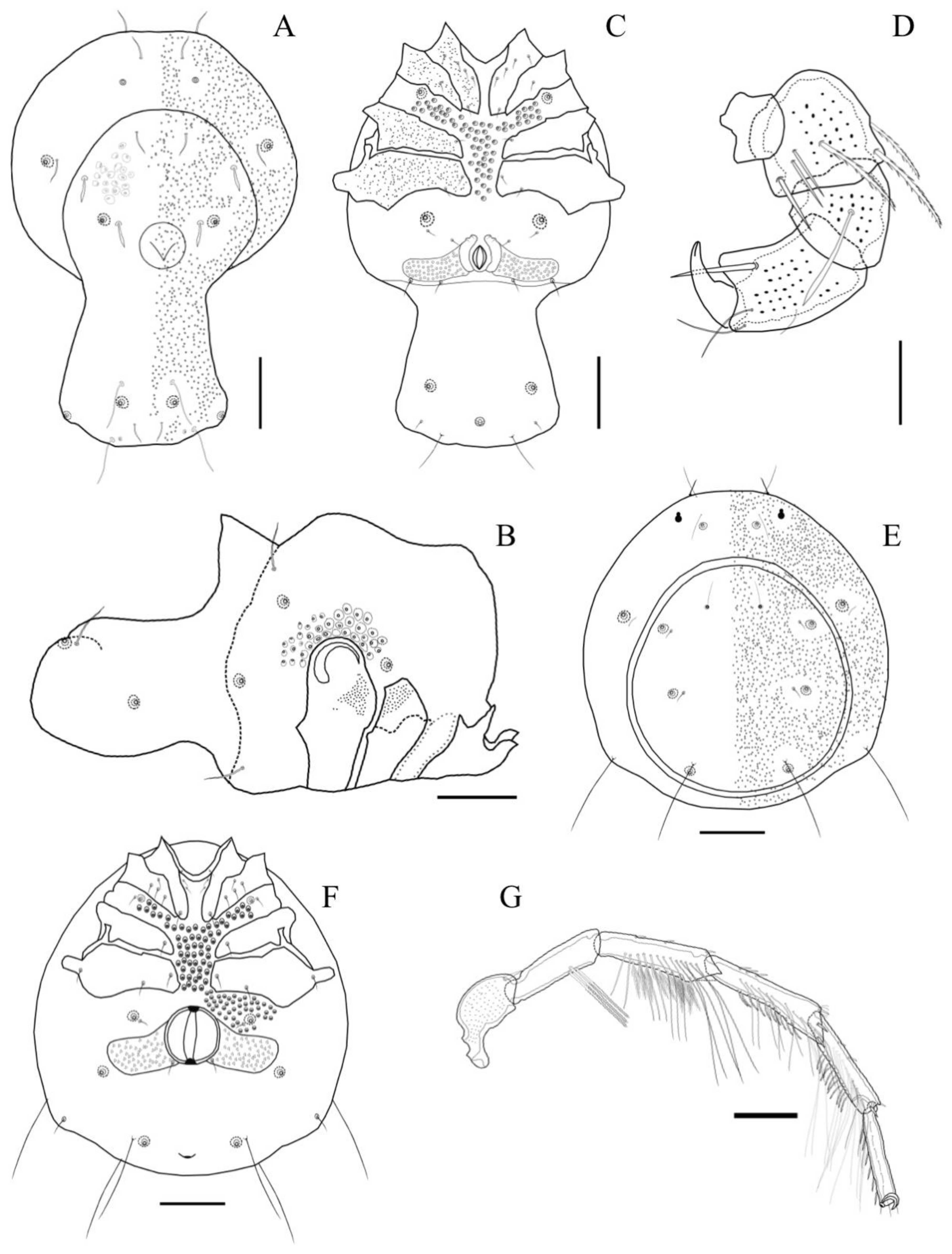
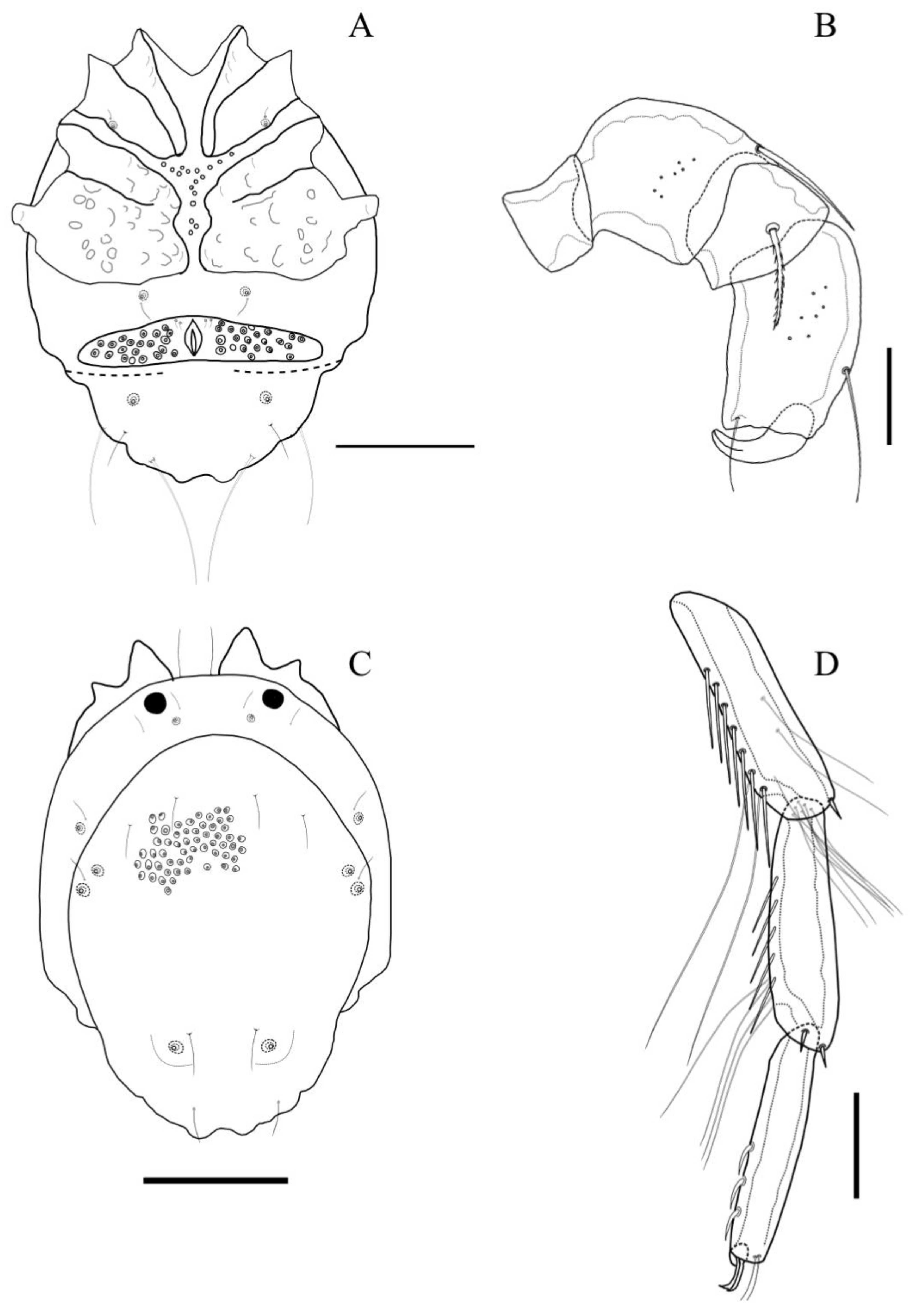
Description. MALE: Idiosoma 640 L, 512 W, dark blue with whitish cauda. Dorsal shield 581 L, 423 W. Dorsal furrow incomplete but continuing posterior to genital field. Cauda is short, 187 L and 285 W, bearing one medial and two pairs of lateral notches (
Figure 12A and
Figure 13C). The anterior part of the idiosoma is wide with a slight constriction at the base of the cauda. Dgl-2 and Dgl-3 are close to each other. Dgl-4 is located at the end of the cauda on small humps. The anterior coxal group with complete suture lines, Cx-III and Cx-IV, separated with an incomplete suture line. Tips of Cx-II significantly protrude beyond the idiosoma’s anterior margin (
Figure 13A). Cxgl-1 is located posteromedially in the margin of Cx-I. Apodemes of Cx-IV protrude slightly beyond the lateral part of the idiosoma. Cxgl-2 with an associated seta posteriorly to Cx-IV (
Figure 13A). Genital field 315 W, gonopore 69 L and 27 W. Dorsal L of palpal segments: P1: 27; P2: 47; P3: 41; P4: 58; P5: 33, P3 with a long, pinnate seta located medially (
Figure 13B). Dorsal L of fourth leg segments: IV-Leg-3: 104, IV-Leg-4: 126, IV-Leg-5: 119, IV-Leg-6: 116, IV-Leg-4-5, with numerous swimming setae and lateral, spine-like setae (seven on IV-Leg-4 and five on IV-Leg-5) (
Figure 13D). FEMALE: Unknown.
Etymology. This species is named after Beatriz Rosso de Ferradás for her invaluable contributions to water mite acarology in South America.
Remarks. This species belongs to the subgenus Megaluracarus. However, the cauda is relatively short compared with other members of the subgenus. The short cauda is a particular characteristic only shared by A. olmeca (Ramírez-Sánchez and Rivas, 2013) from Mexico and A. amazonicus (Viets, 1954) from Brazil. However, both A. olmeca and A. amazonicus have a patch of spatulate setae on the medial side of P2, while A. beatrizae exhibits only one long, pinnate seta. Additionally, the cauda posterior margin in both A. olmeca and A. amazonicus is not indented. Finally, the number of swimming setae on IV-Leg-4 is reduced in A. olmeca compared with A. beatrizae n. sp.
Distribution. So far only known from el Ramonal, Quintana Roo and San Pedrito lagoon, Tabasco.
Holotype: Male from Ramonal wetland, Quintana Roo (access number: ECO-CH-Z-10625), 19°23′31″ N, −82°37′27″ W; emergent vegetation, 14 April 2019. Coll. L Montes and T. Goldschmidt.
Paratypes: One male and two females. Same data as holotype (access number: ECO-CH-Z-10626-10627).
Diagnosis. Male cauda with two falcate setae located posterolaterally, P2 medially with three simple setae, and one pinnate seta on the anterolateral part.
Description. MALE: Idiosoma 364 L and 295 W, uniformly dark blue (
Figure 14A). Dorsal furrow incomplete. Dorsal shield 305 L and 207 W, short and relatively square cauda, 49 L. Dgl-4 anteriorly located on the cauda with the associated setae located on small humps and posteriorly in the idiosoma, with two small falcate setae on the posterolateral part of the cauda (
Figure 16A). Coxae are occupying two-thirds of the ventral region, suture lines complete. Suture lines of Cx-I–III are diagonally elongated. Cxgl-2 between Cx-II and Cx-IV. Posteriorly to Cx-IV, is the Cxgl-2 located (
Figure 16B). Genital field, 246 W, elongated almost reaching the sides of the ventral area, gonopore 59 L and 14 W. Dorsal L of palpal segments L: P1: 30; P2: 58; P3: 38; P4: 63; P5: 30. P2 with three simple setae medially located and one pinnate seta on the anterolateral, P4 rotated (
Figure 16C). L of fourth leg segments: IV-Leg-3: 63, IV-Leg-4: 73, IV-Leg-5: 100, IV-Leg-6: 101, IV-Leg-5 with one pinnate seta posteromedially located and four spine-like setae along the dorsal medially surface (
Figure 16E).
FEMALE: Idiosoma 522 L and 483 W, uniformly dark blue (
Figure 15A), dorsal shield oval, 463 L and 384 W (dorsal furrow complete), four dorsal pairs of glandularias present. Dgl-2 (on the ventral plate) is close to Dgl-3 (on the dorsal plate). Dgl-3 setae are located posteriorly and separated from their respective glandularia (
Figure 16D). With complete suture lines, coxae occupy half of the ventral area, Cx-I, and Cx-II, elongated and extended diagonally. Cx-III and Cx-IV separated, Cx-III elongated and diagonally located, suture lines of Cx-III−IV sloping, Cx-IV triangular without medial margin. Cxgl-1 is located between Cx-II and Cx-III. Genital field 335 W, straight and with numerous associated acetabula, gonopore 118 L and 112 W. Cxgl-2 between genital area and Cx-IV (
Figure 16F).
Consensus sequence: ACTCTTTATTTTGCCTTTGGATTCTGATCAGGTATGGTAGGTGCAAGATTAAGAAGACTAATTCGCTTAGAATTAGGACAACCAGGGAGACTCTTAGGGAGAGACCAAATTTACAACACAATCGTAACAGCTCATGCTTTTATCATAATCTTTTTTATAGTTATACCTATTATAATTGGAGGTTTCGGAAACTGACTAGTTCCTCTTATACTAGCAGCTCCAGATATGGCATTCCCACGAATAAACAATATAAGATTTTGACTTCTTCCCCCAGCTTTAATTCTCCTTCTATCTAGATCTCTCTCCTCAACAGGAGCAGGAACAGGGTGAACAGTATATCCNCCACTTTCAAGTAACATTGCACATGGAGGACCTTCAGTTGACATAGCAATCTTTTCCCTCCATTTAGCAGGAGTCTCATCAATTCTAGGTGCTATCAATTTCTTAGCTACAACCATCAATATAAAACCAAAATATATAAAATATGATCGTATTCCTCTATTTGTCATTTCAATTTTCATCACAGTTATTCTCCTTCTCCTATCTTTACCAGTCTTAGCAGGAGCCATTACCATACTTTTAACTGACCGAAACTTTAATACATCATTTTTTGATCCAGCTGGAGGAGGAGACCCAATTCTATATCAA.
Etymology. This species is named after Cristina Cramer Hemkes for her invaluable contributions to water mite acarology in Mexico.
Remarks. The present species belong to the
Dadayella subgenus, characterized by males with a small or undifferentiated cauda with an incomplete dorsal furrow and P2 with a simple chaetotaxy [
4].
Arrenurus (
Dadayella)
cristinae n. sp. is similar to
A. veracruzensis (Cramer and Cook, 1992) in the shape and size of the idiosoma, particularly in the quadrangular silhouette of the cauda. The female of
A. veracruzensis is similar to the new species. However, the dorsal shield of the A. veracruzensis female has three pairs of glandularia, while
A. cristinae n. sp. has two. Furthermore, the chaetotaxy of P2 is quite different;
A. cristinae n. sp. presents three simple, medial setae and a little, pinnate seta in the anterior-lateral part while
A. veracruzensis presents four medial, spine-like setae. Additionally,
A. cristinae n. sp. presents two falcate setae on the posterolateral part of the cauda, which are absent in
A. veracruzensis. Most of the
Dadayella species described are known from females, making comparisons difficult due to their scarce morphological variation. It was possible for
A. cristinae n. sp. to obtain the DNA barcode with the BIN AEA7842. Therefore, we could, undoubtedly, assign the female to the respective male (
Figure 2). These data represent the first sequences obtained for this subgenus.
Distribution. So far only known from the type locality (Ramonal, Quintana Roo).






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}