
The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity

 ,
,  , , ,
, , , 
Abstract
:1. Introduction
2. Materials and Methods
Molecular Analyses
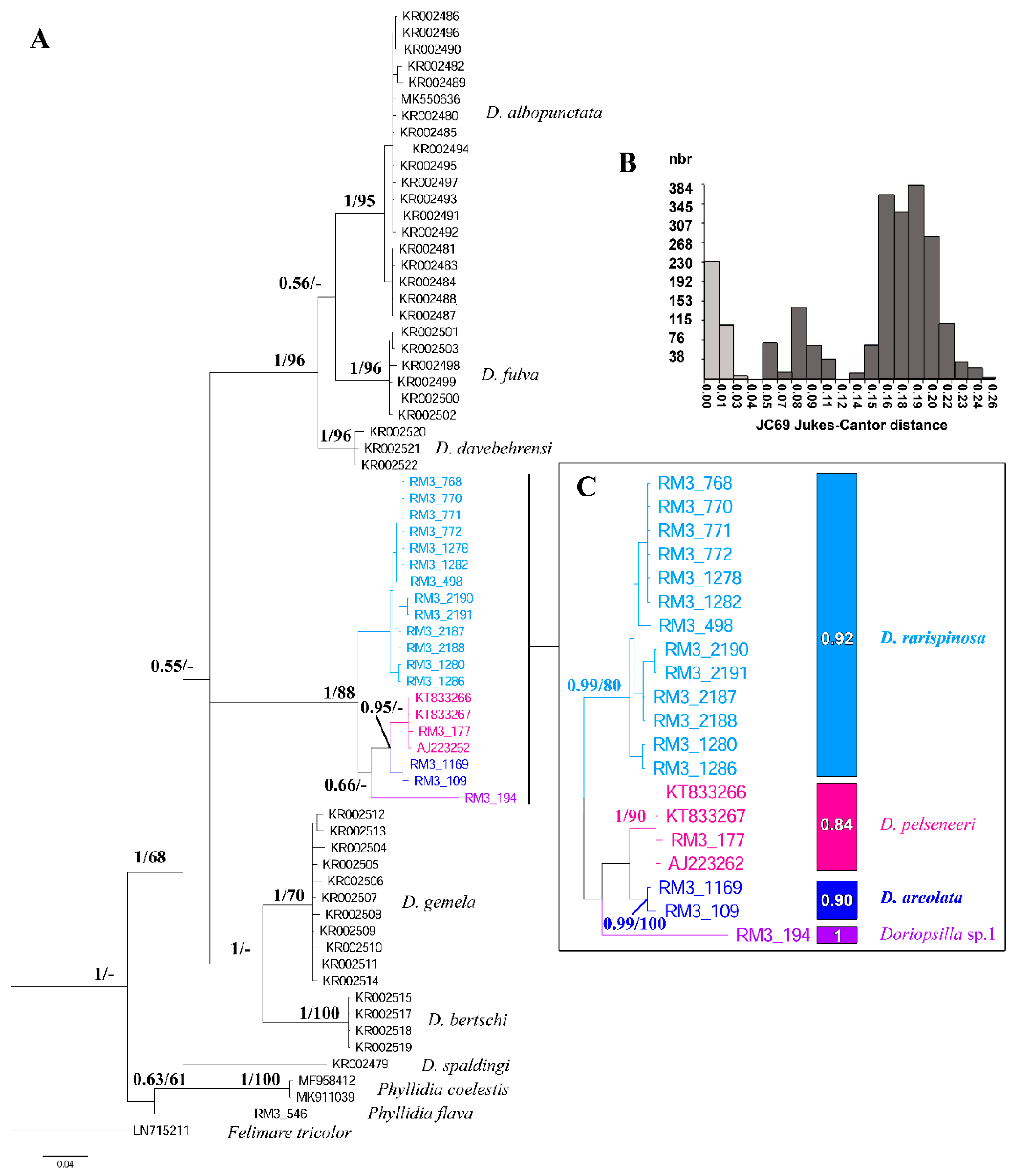
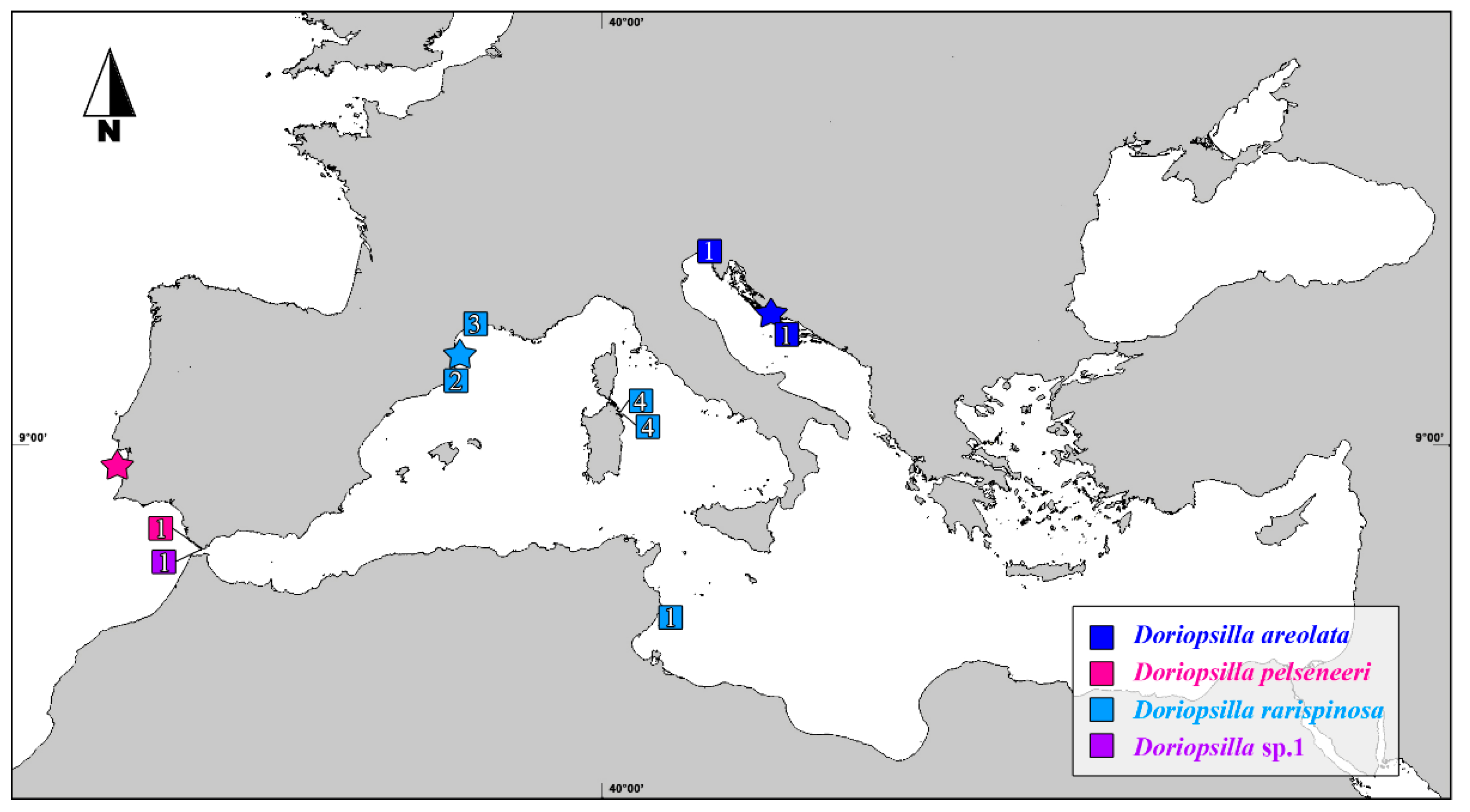
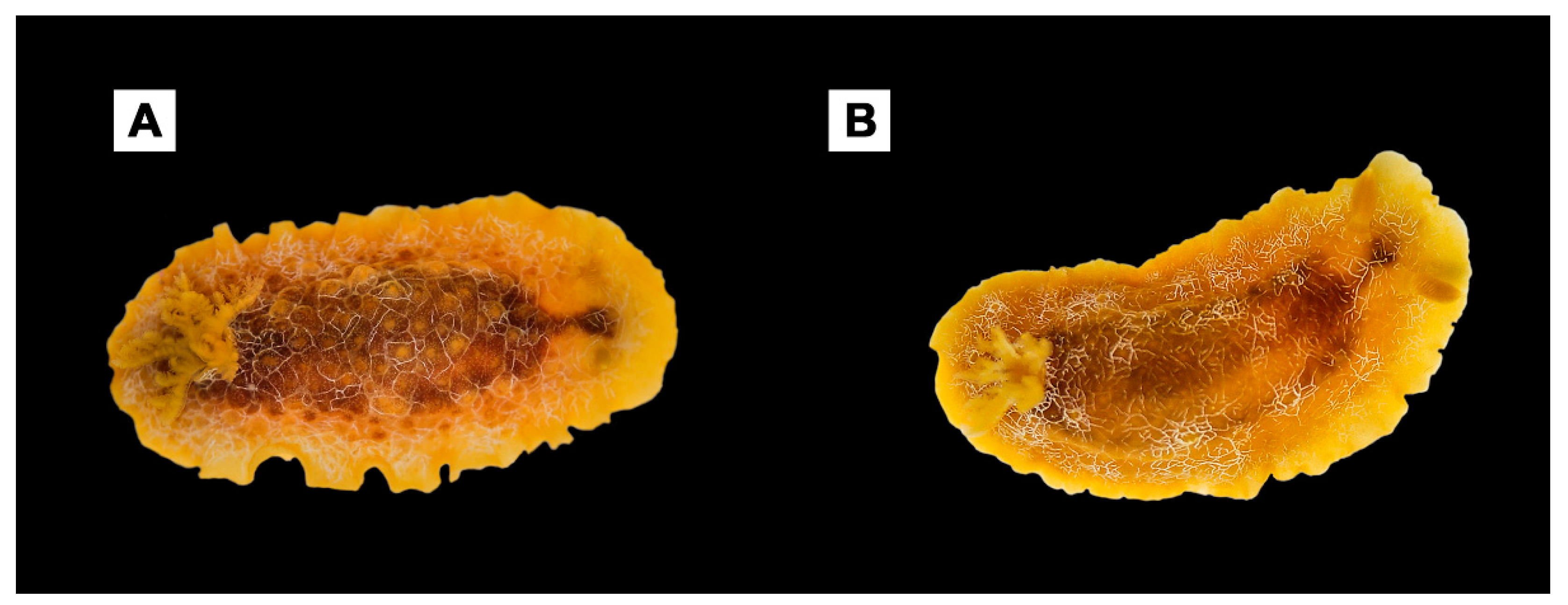
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global Diversity of Gastropods (Gastropoda; Mollusca) in Freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Brown, K.M. 10—Mollusca: Gastropoda. In Ecology and Classification of North American Freshwater Invertebrates, 2nd ed.; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 297–329. ISBN 978-0-12-690647-9. [Google Scholar]
- Wägele, H.; Willan, R.C. Phylogeny of the Nudibranchia. Zool. J. Linn. Soc. 2000, 130, 83–181. [Google Scholar] [CrossRef]
- Gavagnin, M.; Mollo, E.; Castelluccio, F.; Ghiselin, M.T.; Calado, G.; Cimino, G. Can Molluscs Biosynthesize Typical Sponge Metabolites? The Case of the Nudibranch Doriopsilla areolata. Tetrahedron 2001, 57, 8913–8916. [Google Scholar] [CrossRef]
- Valdés, A. Preliminary Molecular Phylogeny of the Radula-Less Dorids (Gastropoda: Opisthobranchia), Based on 16S MtDNA Sequence Data. J. Molluscan Stud. 2003, 69, 75–80. [Google Scholar] [CrossRef] [Green Version]
- Soares, C.; Calado, G. Spawn and Early Development of Two Sympatric Species of the Genus Doriopsilla (Opisthobranchia: Nudibranchia) with Contrasting Development Strategies. Rec. Aust. Mus. 2006, 69, 103–109. [Google Scholar] [CrossRef] [Green Version]
- Healy, J.M.; Willan, R.C. Nudibranch Spermatozoa: Comparative Ultrastructure and Systematic Importance. Veliger 1991, 34, 134–165. [Google Scholar]
- Brunckhorst, D.J. The Systematics and Phylogeny of Phyllidiid Nudibranchs (Doridoidea). Rec. Aust. Mus. 1993, 16 (Suppl. 16), 1–107. [Google Scholar] [CrossRef] [Green Version]
- Valdés, A. Revisión de La Superfamilia Porodoridoidea Odhner En Franc, 1968 (Mollusca: Nudibranchia) En El Océano Atlántico. Ph.D. Thesis, Universidad de Oviedo, Oviedo, Spain, 1996. [Google Scholar]
- Valdés, Á.; Gosliner, T.M. Phylogeny of the Radula-Less Dorids (Mollusca, Nudibranchia), with the Description of a New Genus and a New Family. Zool. Scr. 1999, 28, 315–360. [Google Scholar] [CrossRef]
- Pruvot-Fol, A. Mollusques Opisthobranches. In Faune de France; Paul Lechevalier: Paris, France, 1954; Volume 58. [Google Scholar]
- Burn, R.F. Notes on a Collection of Nudibranchia (Gastropoda: Dorididae and Dendrodorididae) from South Australia with Remarks on the Species of Basedow and Hedley, 1905. Mem. Natl. Mus. Vic. 1962, 25, 149–171. [Google Scholar] [CrossRef]
- Valdés, A.; Ortea, J.; Ávila, C.; Ballesteros, M. Review of the Genus Dendrodoris (Ehrenberg, 1831) (Gastropoda: Nudibranchia) in the Atlantic Ocean. J. Molluscan Stud. 1996, 62, 1–31. [Google Scholar] [CrossRef]
- World Register of Marine Species. Available online: https://www.marinespecies.org (accessed on 30 March 2022).
- Valdés, A.; Ortea, J. Review of the Genus Doriopsilla Bergh, 1880 (Gastropoda: Nudibranchia) in the Atlantic Ocean. Veliger 1997, 40, 240–254. [Google Scholar]
- Yonow, N. Opisthobranchs from the Western Indian Ocean, with Descriptions of Two New Species and Ten New Records (Mollusca, Gastropoda). ZooKeys 2012, 197, 1–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoover, C.; Lindsay, T.; Goddard, J.H.R.; Valdés, Á. Seeing Double: Pseudocryptic Diversity in the Doriopsilla albopunctata–Doriopsilla gemela Species Complex of the North-Eastern Pacific. Zool. Scr. 2015, 44, 612–631. [Google Scholar] [CrossRef]
- Ballesteros, M.; Ortea, J. Contribución al Conocimiento de Los Dendrodorididae (Moluscos: Opistobranquios: Doridáceos) Del Litoral Ibérico; Publicaciones del Departamento de Zoología, Universidad de Barcelona, Facultad de Biología: Barcelona, Spain, 1980; Volume 5, pp. 25–37. [Google Scholar]
- d’Oliveira, M.P. Opisthobranches Du Portugal de La Collection de M. Paulino d’Oliveira. O Instituto Rev. Sci. E Litt. 1895, 42, 574–592. [Google Scholar]
- Sea Slug Forum. Available online: http://seaslugforum.net/showall/doripels (accessed on 30 March 2022).
- García-Gómez, J.C.; Cervera, J.L.; García, F.J.; López de la Cuadra, C.M. Resultados de La Campaña Internacional de Biología Marina “Ceuta-86”: Moluscos Opistobranquios. Boll. Malac. 1989, 25, 223–232. [Google Scholar]
- Domènech, A.; Ávila, C.; Ballesteros, M. Opisthobranch Molluscs from the Subtidal Trawling Grounds off Blanes (Girona, North-East Spain). J. Mar. Biol. Assoc. U. K. 2006, 86, 383–389. [Google Scholar] [CrossRef] [Green Version]
- Templado, J.; Guerra, A.; Bedoya, J.; Moreno, D.; Remón, J.M.; Maldonado, M.; Ramos, M.A. Fauna Marina Circalitoral Del Sur de La Península Ibérica: Resultados de La Campaña Oceanográfica “Fauna I”; Museo Nacional de Ciencias Naturales (CSIC): Madrid, Spain, 1993; ISBN 978-84-00-07380-0. [Google Scholar]
- Schick, K.L. Atlas Submarino de La Costa Del Sol; Marbella: Málaga, Spain, 1998; ISBN 84-605-7552-7. [Google Scholar]
- Trainito, E.; Doneddu, M. Nudibranchi del Mediterraneo, 2nd ed.; Il Castello: Milan, Italy, 2014; ISBN 978-88-6520-480-1. [Google Scholar]
- DORIS. Available online: https://doris.ffessm.fr/ref/specie/1320 (accessed on 30 March 2022).
- Bergh, L.S.R. Die Doriopsen Des Mittelmeeres. Jahrb. Dtsch. Malakozool. Ges. 1880, 7, 297–328. [Google Scholar]
- Valdés, Á.; Hamann, J. Two New Species of Doriopsilla from the Tropical Western Atlantic with Remarks on Cariopsillidae Ortea & Espinosa, 2005. Veliger 2008, 50, 210–218. [Google Scholar]
- Bergh, L.S.R. Die Nudibranchiata Holohepatica Porostomata. Verh. K. Kais. Zool. Bot. Ges. 1892, 42, 1–16. [Google Scholar]
- Young, D.K. The Functional Morphology of the Feeding Apparatus of Some Indo-West-Pacific Dorid Nudibranchs. Malacologia 1969, 9, 421–446. [Google Scholar]
- Edmunds, M. Opisthobranchiate Mollusca from Ghana. J. Moll. Stud. 1968, 38, 83–100. [Google Scholar] [CrossRef]
- Meyer, K.B. Dorid Nudibranchs of the Caribbean Coast of the Panama Canal Zone. Bull. Mar. Sci. 1977, 27, 299–307. [Google Scholar]
- Goodheart, J.; Valdés, Á. Re-Evaluation of the Doriopsilla areolata Bergh, 1880 (Mollusca: Opisthobranchia) Subspecies Complex in the Eastern Atlantic Ocean and Its Relationship to South African Doriopsilla miniata (Alder & Hancock, 1864) Based on Molecular Data. Mar. Biodivers. 2013, 43, 113–120. [Google Scholar] [CrossRef]
- Furfaro, G.; Modica, M.V.; Oliverio, M.; Mariottini, P. A DNA-Barcoding Approach to the Phenotypic Diversity of Mediterranean Species of Felimare Ev. Marcus & Er. Marcus, 1967 (Mollusca: Gastropoda), with a Preliminary Phylogenetic Analysis. Ital. J. Zool. 2016, 83, 195–207. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Picton, B.E.; Martynov, A.V.; Mariottini, P. Diaphorodoris alba Portmann & Sandmeier, 1960 Is a Valid Species: Molecular and Morphological Comparison with D. luteocincta (M. Sars, 1870) (Gastropoda: Nudibranchia). Zootaxa 2016, 4193, 304–316. [Google Scholar] [CrossRef]
- Furfaro, G.; Mariottini, P.; Modica, M.V.; Trainito, E.; Doneddu, M.; Oliverio, M. Sympatric Sibling Species: The Case of Caloria elegans and Facelina quatrefagesi (Gastropoda: Nudibranchia). Sci. Mar. 2016, 80, 511–520. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Salvi, D.; Mancini, E.; Mariottini, P. A Multilocus View on Mediterranean Aeolid Nudibranchs (Mollusca): Systematics and Cryptic Diversity of Flabellinidae and Piseinotecidae. Mol. Phylogenet. Evol. 2018, 118, 13–22. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Trainito, E.; Vitale, F.; Mariottini, P. When Morphology Does Not Match Phylogeny: The Puzzling Case of Two Sibling Nudibranchs (Gastropoda). Zool. Scr. 2021, 50, 439–454. [Google Scholar] [CrossRef]
- Reece, J.S.; Castoe, T.A.; Parkinson, C.L. Historical Perspectives on Population Genetics and Conservation of Three Marine Turtle Species. Conserv. Genet. 2005, 6, 235–251. [Google Scholar] [CrossRef]
- Luttikhuizen, P.C.; Campos, J.; van Bleijswijk, J.; Peijnenburg, K.T.C.A.; van der Veer, H.W. Phylogeography of the Common Shrimp, Crangon crangon (L.) across Its Distribution Range. Mol. Phylogenet. Evol. 2008, 46, 1015–1030. [Google Scholar] [CrossRef]
- Triantaphyllidis, A.; Apostolidis, A.P.; Katsares, V.; Kelly, E.; Mercer, J.; Hughes, M.; Jørstad, K.E.; Tsolou, A.; Hynes, R.; Triantaphyllidis, C. Mitochondrial DNA Variation in the European Lobster (Homarus gammarus) throughout the Range. Mar. Biol. 2005, 146, 223–235. [Google Scholar] [CrossRef]
- Galarza, J.A.; Carreras-Carbonell, J.; Macpherson, E.; Pascual, M.; Roques, S.; Turner, G.F.; Rico, C. The Influence of Oceanographic Fronts and Early-Life-History Traits on Connectivity among Littoral Fish Species. Proc. Natl. Acad. Sci. USA 2009, 106, 1473–1478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boissin, E.; Stöhr, S.; Chenuil, A. Did Vicariance and Adaptation Drive Cryptic Speciation and Evolution of Brooding in Ophioderma longicauda (Echinodermata: Ophiuroidea), a Common Atlanto-Mediterranean Ophiuroid? Mol. Ecol. 2011, 20, 4737–4755. [Google Scholar] [CrossRef] [PubMed]
- Claremont, M.; Reid, D.G.; Williams, S.T. Evolution of Corallivory in the Gastropod Genus Drupella. Coral Reefs 2011, 30, 977–990. [Google Scholar] [CrossRef]
- Barco, A.; Houart, R.; Bonomolo, G.; Crocetta, F.; Oliverio, M. Molecular Data Reveal Cryptic Lineages within the Northeastern Atlantic and Mediterranean Small Mussel Drills of the Ocinebrina edwardsii Complex (Mollusca: Gastropoda: Muricidae). Zool. J. Linn. Soc-Lond. 2013, 169, 389–407. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, T.; Valdés, Á. The Model Organism Hermissenda crassicornis (Gastropoda: Heterobranchia) Is a Species Complex. PLoS ONE 2016, 11, e0154265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furfaro, G.; Mariottini, P. A New Dondice Marcus Er. 1958 (Gastropoda: Nudibranchia) from the Mediterranean Sea Reveals Interesting Insights into the Phylogenetic History of a Group of Facelinidae Taxa. Zootaxa 2020, 4731, 1–22. [Google Scholar] [CrossRef]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Deptartment of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 2002. [Google Scholar]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 SnRNA DNA Sequences and Arthropod Molecular Evolution. Aust. J. Zool. 1999, 46, 419–437. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castresana, J. Selection of Conserved Blocks from Multiple Alignments for Their Use in Phylogenetic Analysis. Mol. Biol. Evol. 2000, 17, 540–552. [Google Scholar] [CrossRef] [Green Version]
- Talavera, G.; Castresana, J. Improvement of Phylogenies after Removing Divergent and Ambiguously Aligned Blocks from Protein Sequence Alignments. Syst. Biol. 2007, 56, 564–577. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; BOISSELIER, M.-C.; Cruaud, C.; Holford, M.; Samadi, S. Large-Scale Species Delimitation Method for Hyperdiverse Groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA Barcoding and Taxonomy in Diptera: A Tale of High Intraspecific Variability and Low Identification Success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. RaxmlGUI: A Graphical Front-End for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Calvo, M.; Templado, J.; Oliverio, M.; Machordom, A. Hidden Mediterranean Biodiversity: Molecular Evidence for a Cryptic Species Complex within the Reef Building Vermetid Gastropod Dendropoma petraeum (Mollusca: Caenogastropoda). Biol. J. Linn. Soc. 2009, 96, 898–912. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Vitale, F.; Licchelli, C.; Mariottini, P. Two Seas for One Great Diversity: Checklist of the Marine Heterobranchia (Mollusca; Gastropoda) from the Salento Peninsula (South-East Italy). Diversity 2020, 12, 171. [Google Scholar] [CrossRef]
- Salento Sommerso. Available online: https://www.salentosommerso.it/pp_nudibranchia/Doriopsilla areolata/pp_Doriopsilla areolata.php (accessed on 8 March 2022).
- Pruvot-Fol, A. Études Des Nudibranches de La Méditerranée. Arch. Zool. Exp. Gén. 1951, 88, 1–80. [Google Scholar]
- Perrone, A.S. Il Genere Doriopsilla Bergh, 1880 in Mediterraneo: Descrizione Di Doriopsilla Rarispinosa Pruvot-Fol, 1951 (Opisthobranchia: Nudibranchia). Boll. Malac. 1986, 22, 97–112. [Google Scholar]
- Cattaneo-Vietti, R.; Chemello, R.; Giannuzzi-Savelli, R. Atlas of Mediterranean Nudibranchs; La Conchiglia: Rome, Italy, 1990. [Google Scholar]
- Siokou-Frangou, I.; Christaki, U.; Mazzocchi, M.G.; Montresor, M.; Ribera d’Alcalá, M.; Vaqué, D.; Zingone, A. Plankton in the Open Mediterranean Sea: A Review. Biogeosciences 2010, 7, 1543–1586. [Google Scholar] [CrossRef] [Green Version]
- Elhmaidi, D.; Nefzi, H.; Carton, X.; Lili, T. Particle Dispersion in the Western Mediterranean Basin. Open Oceanogr. J. 2015, 4, 137–143. [Google Scholar] [CrossRef] [Green Version]
- Pinardi, N.; Masetti, E. Variability of the Large Scale General Circulation of the Mediterranean Sea from Observations and Modelling: A Review. Palaeogeogr. Palaeoclimatol. Palaeoecol. 2000, 158, 153–173. [Google Scholar] [CrossRef]
- Poulain, P.-M.; Menna, M.; Mauri, E. Surface Geostrophic Circulation of the Mediterranean Sea Derived from Drifter and Satellite Altimeter Data. J. Phys. Oceanogr. 2012, 42, 973–990. [Google Scholar] [CrossRef]
- Valdés, Á.; Alexander, J.; Crocetta, F.; Yokeş, M.B.; Giacobbe, S.; Poursanidis, D.; Zenetos, A.; Lucas Cervera, J.; Caballer, M.; Galil, B.S.; et al. The Origin and Dispersal Pathway of the Spotted Sea Hare Aplysia dactylomela (Mollusca: Opisthobranchia) in the Mediterranean Sea. Aquat. Invasions 2013, 8, 427–436. [Google Scholar] [CrossRef]
- Kyle, C.J.; Boulding, E.G. Comparative Population Genetic Structure of Marine Gastropods (Littorina Spp.) with and without Pelagic Larval Dispersal. Mar. Biol. 2000, 137, 835–845. [Google Scholar] [CrossRef]
- Blanc, P.-L. The Opening of the Plio-Quaternary Gibraltar Strait: Assessing the Size of a Cataclysm. Geodin. Acta 2002, 15, 303–317. [Google Scholar] [CrossRef]
- Zenetos, A.; Gofas, S.; Velarque, M.; Çinar, M.E.; García Raso, J.E.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien Species in the Mediterranean Sea by 2010. A Contribution to the Application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial Distribution. Mediterr. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef] [Green Version]
- Maggio, T.; Lo Brutto, S.; Garoia, F.; Tinti, F.; Arculeo, M. Microsatellite Analysis of Red Mullet Mullus barbatus (Perciformes, Mullidae) Reveals the Isolation of the Adriatic Basin in the Mediterranean Sea. ICES J. Mar. Sci. 2009, 66, 1883–1891. [Google Scholar] [CrossRef] [Green Version]
- Ragionieri, L.; Schubart, C.D. Population Genetics, Gene Flow, and Biogeographical Boundaries of Carcinus estuarii (Crustacea: Brachyura: Carcinidae) along the European Mediterranean Coast. Biol. J. Linn. Soc. 2013, 109, 771–790. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Voucher | Locality | Accession Numbers | ||
|---|---|---|---|---|---|
| H3 | 16S | COI | |||
| Doriopsilla albopunctata | LACM:DISCO 11426 | White Point, California, USA (E-PAC) | - | - | MK550636 |
| CPIC 00909 | Long Beach, California, USA (E-PAC) | - | KR002428 | KR002480 | |
| CPIC 00915 | Long Beach, California, USA (E-PAC) | KR002524 | KR002429 | KR002481 | |
| CPIC 00916 | Long Beach, California, USA (E-PAC) | - | KR002430 | KR002482 | |
| CPIC 00917 | Long Beach, California, USA (E-PAC) | KR002525 | - | KR002483 | |
| CPIC 00930 | Malibu, California, USA (E-PAC) | KR002527 | KR002431 | KR002485 | |
| CPIC 00932 | Malibu, California, USA (E-PAC) | KR002528 | KR002432 | KR002486 | |
| CPIC 01254 | Shell Beach, California, USA (E-PAC) | KR002535 | KR002440 | KR002494 | |
| CPIC 01255 | Shell Beach, California, USA (E-PAC) | KR002536 | KR002441 | KR002495 | |
| CPIC 01084 | Mission Bay, California, USA (E-PAC) | - | KR002442 | KR002496 | |
| CPIC 01083 | Redondo Beach, California, USA (E-PAC) | - | KR002443 | KR002497 | |
| CPIC 00918 | Long Beach, California, USA (E-PAC) | KR002526 | - | KR002484 | |
| CPIC 01239 | Carpinteria, California, USA (E-PAC) | KR002534 | KR002439 | KR002493 | |
| CPIC 00987 | Carpinteria, California, USA (E-PAC) | KR002529 | KR002434 | KR002488 | |
| CPIC 00986 | Carpinteria, California, USA (E-PAC) | - | KR002433 | KR002487 | |
| LACM:3420 | Carpinteria, California, USA (E-PAC) | KR002530 | KR002435 | KR002489 | |
| - | Newport Beach, California, USA (E-PAC) | KR002531 | KR002436 | KR002490 | |
| - | Newport Beach, California, USA (E-PAC) | KR002532 | KR002437 | KR002491 | |
| - | Newport Beach, California, USA (E-PAC) | KR002533 | KR002438 | KR002492 | |
| Doriopsilla areolata | RM3_109 | Hvar, Croatia (MED) | ON209460 | ON229526 | ON211997 |
| RM3_1169 | Portopiccolo, Sistiana, Trieste, Italy (MED) | ON209466 | ON229532 | ON211996 | |
| As D. areolata in GB | MNCN:15.05/23766 | Girona, Catalonia, Spain (MED) | KC171040 | KC171023 | - |
| MNCN:15.05/23782 | Las Palmas, Gran Canaria, Spain (E-ATL) | KC171037 | KC171026 | - | |
| LACM:2001-10.3 | Cadiz, Andalusia, Spain (E-ATL) | KC171035 | KC171024 | - | |
| LACM:2001-10.4 | Cadiz, Andalusia, Spain (E-ATL) | KC171036 | KC171025 | - | |
| MNCN:15.05/23784 | Angola (E-ATL) | KC171039 | KC171033 | - | |
| MNCN:15.05/23789 | Angola (E-ATL) | KC171038 | KC171031 | - | |
| Doriopsilla bertschi | CPIC 00976 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002551 | - | KR002515 |
| CPIC 01058 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002552 | KR002462 | KR002517 | |
| CPIC 01059 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | - | KR002463 | KR002518 | |
| LACM:140785 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002561 | KR002471 | KR002519 | |
| LACM:3421 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002553 | KR002464 | - | |
| Doriopsilla davebehrensi | CPIC 01038 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002564 | KR002475 | KR002520 |
| LACM:3419 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002565 | KR002476 | KR002521 | |
| LACM:76-5.6 | Bahia de los Angeles, Baja California, Mexico (E-PAC) | KR002566 | KR002478 | - | |
| - | Newport Beach, California, USA (E-PAC) | - | KR002477 | KR002522 | |
| Doriopsilla fulva | CPIC 00933 | Malibu, California, USA (E-PAC) | KR002537 | KR002444 | KR002498 |
| CPIC 00934 | Malibu, California, USA (E-PAC) | KR002538 | KR002445 | KR002499 | |
| CPIC 00936 | Malibu, California, USA (E-PAC) | KR002539 | KR002446 | KR002500 | |
| CPIC 00937 | Malibu, California, USA (E-PAC) | - | KR002447 | KR002501 | |
| CPIC 01240 | Mendocino, California, USA (E-PAC) | KR002541 | KR002449 | KR002503 | |
| CPIC 01022 | Palos Verdes, California, USA (E-PAC) | KR002540 | KR002448 | KR002502 | |
| Doriopsilla gemela | CPIC 00923 | Malibu, California, USA (E-PAC) | KR002542 | KR002450 | - |
| CPIC 00924 | Malibu, California, USA (E-PAC) | KR002543 | KR002451 | KR002504 | |
| CPIC 00931 | Malibu, California, USA (E-PAC) | - | KR002452 | KR002505 | |
| CPIC 00938 | Malibu, California, USA (E-PAC) | KR002544 | KR002453 | KR002506 | |
| CPIC 00939 | Malibu, California, USA (E-PAC) | KR002545 | KR002454 | KR002507 | |
| CPIC 00978 | Carpinteria, California, USA (E-PAC) | KR002546 | KR002455 | KR002508 | |
| CPIC 00979 | Carpinteria, California, USA (E-PAC) | KR002547 | KR002456 | KR002509 | |
| CPIC 00980 | Carpinteria, California, USA (E-PAC) | - | KR002457 | KR002510 | |
| CPIC 00981 | Carpinteria, California, USA (E-PAC) | - | KR002458 | KR002511 | |
| CPIC 00982 | Carpinteria, California, USA (E-PAC) | KR002548 | KR002459 | KR002512 | |
| CPIC 00983 | Carpinteria, California, USA (E-PAC) | KR002549 | KR002460 | KR002513 | |
| CPIC 00984 | Carpinteria, California, USA (E-PAC) | KR002550 | KR002461 | KR002514 | |
| Doriopsilla janaina | CPIC 00590 | Peru (E-PAC) | KC171034 | KC171022 | - |
| Doriopsilla miniata | CAS:IZ:176370 | South Africa (E-ATL) | KC171043 | KC171030 | - |
| CAS:IZ:176418 | Western Cape, South Africa (E-ATL) | KC171041 | KC171028 | - | |
| CAS:IZ:176933 | Western Cape, South Africa (E-ATL) | KC171042 | KC171029 | - | |
| D. pelseneeri | RM3_177 | Tarifa, Andalusia, Spain (E-ATL) | ON209459 | ON229525 | ON211995 |
| As D. areolata in GB | - | Cadiz, Andalusia, Spain (E-ATL) | - | AJ225186 | AJ223262 |
| - | Berlengas, Portugal (E-ATL) | - | KT820536 | KT833266 | |
| - | Berlengas, Portugal (E-ATL) | - | - | KT833267 | |
| D. rarispinosa | RM3_768 | Tavolara Island, Sardinia, Italy (MED) | ON209461 | ON229527 | ON211998 |
| RM3_770 | Tavolara Island, Sardinia, Italy (MED) | ON209462 | ON229528 | ON211999 | |
| RM3_771 | Tavolara Island, Sardinia, Italy (MED) | ON209463 | ON229529 | ON212000 | |
| RM3_772 | Tavolara Island, Sardinia, Italy (MED) | ON209464 | ON229530 | ON212001 | |
| RM3_498 | Kerkennah, Tunisia (MED) | ON209465 | ON229531 | ON212002 | |
| RM3_1278 | Golfo di Olbia, Sardinia, Italy (MED) | ON209467 | ON229533 | ON212003 | |
| RM3_1280 | Golfo di Olbia, Sardinia, Italy (MED) | ON209468 | ON229534 | ON212004 | |
| RM3_1282 | Golfo di Olbia, Sardinia, Italy (MED) | ON209469 | ON229535 | ON212005 | |
| RM3_1286 | Golfo di Olbia, Sardinia, Italy (MED) | ON209470 | ON229536 | ON212006 | |
| RM3_2187 | Palavas-les-flots, Occitanie, France (MED) | ON209471 | ON229537 | ON212007 | |
| RM3_2188 | Palavas-les-flots, Occitanie, France (MED) | ON209472 | ON229538 | ON212008 | |
| RM3_2189 | Palavas-les-flots, Occitanie, France (MED) | ON209473 | ON229539 | - | |
| RM3_2190 | L’Escala, Girona, Catalonia, Spain (MED) | ON209474 | ON229540 | ON212009 | |
| RM3_2191 | L’Escala, Girona, Catalonia, Spain (MED) | ON209475 | ON229541 | ON212010 | |
| Doriopsilla spaldingi | CPIC 00908 | San Pedro, California, USA (E-PAC) | KR002523 | KR002427 | KR002479 |
| Doriopsilla sp. 1 | RM3_194 | Tarifa, Andalusia, Spain (E-ATL) | ON209458 | ON229524 | ON211994 |
| Felimare tricolor | BAU 20547 | Secche di Tor Paterno MPA, Lazio, Italy (MED) | MK474153 | LN715193 | LN715211 |
| Phyllidia coelestis | CAS:IZ:190982 | Kranket Island, Madang Prov., Papua New Guinea (W-PAC) | - | MF958279 | MF958412 |
| Phco18LS1 | Lembeh, North Sulawesi, Strait Indonesia (IWP) | - | MK852557 | MK911039 | |
| Phyllidia flava | RM3_546 | Giglio Island, Tuscany, Italy (MED) | ON209476 | - | ON212011 |
| 1. | 2. | 3. | 4. | 5. | 6. | 7. | 8. | 9. | 10. | 11. | 12. | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1. D. albopunctata | - | |||||||||||
| 2. D. areolata | 0.18 | - | ||||||||||
| 3. D. bertschi | 0.16 | 0.16 | - | |||||||||
| 4. D. davebehrensi | 0.08 | 0.18 | 0.16 | - | ||||||||
| 5. D. fulva | 0.09 | 0.17 | 0.15 | 0.08 | - | |||||||
| 6. D. gemela | 0.15 | 0.16 | 0.10 | 0.15 | 0.15 | - | ||||||
| 7. D. pelseneeri | 0.17 | 0.03 | 0.16 | 0.18 | 0.16 | 0.17 | - | |||||
| 8. D. rarispinosa | 0.19 | 0.06 | 0.15 | 0.17 | 0.17 | 0.16 | 0.06 | - | ||||
| 9. D. spaldingi | 0.18 | 0.18 | 0.15 | 0.16 | 0.17 | 0.15 | 0.18 | 0.17 | - | |||
| 10. Doriopsilla sp. 1 | 0.21 | 0.09 | 0.17 | 0.20 | 0.20 | 0.16 | 0.09 | 0.09 | 0.18 | - | ||
| 11. Phyllidia spp. | 0.18 | 0.18 | 0.17 | 0.18 | 0.17 | 0.16 | 0.18 | 0.17 | 0.17 | 0.19 | - | |
| 12. Felimare tricolor | 0.22 | 0.18 | 0.19 | 0.22 | 0.21 | 0.19 | 0.17 | 0.17 | 0.19 | 0.19 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furfaro, G.; Schreier, C.; Trainito, E.; Pontes, M.; Madrenas, E.; Girard, P.; Mariottini, P. The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity. Diversity 2022, 14, 297. https://doi.org/10.3390/d14040297
Furfaro G, Schreier C, Trainito E, Pontes M, Madrenas E, Girard P, Mariottini P. The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity. Diversity. 2022; 14(4):297. https://doi.org/10.3390/d14040297
Chicago/Turabian StyleFurfaro, Giulia, Christopher Schreier, Egidio Trainito, Miquel Pontes, Enric Madrenas, Pascal Girard, and Paolo Mariottini. 2022. "The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity" Diversity 14, no. 4: 297. https://doi.org/10.3390/d14040297
APA StyleFurfaro, G., Schreier, C., Trainito, E., Pontes, M., Madrenas, E., Girard, P., & Mariottini, P. (2022). The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity. Diversity, 14(4), 297. https://doi.org/10.3390/d14040297