
New Comparative Data on the Long Bone Microstructure of Large Extant and Extinct Flightless Birds



Abstract
:1. Introduction
2. Materials and Methods
2.1. Biological Sample
2.2. Thin-Section Preparation and Histological Descriptions
2.3. Simple Linear Measurements and Global Compactness
3. Results
3.1. Tarsometatarsi Proportions and Type of Locomotion
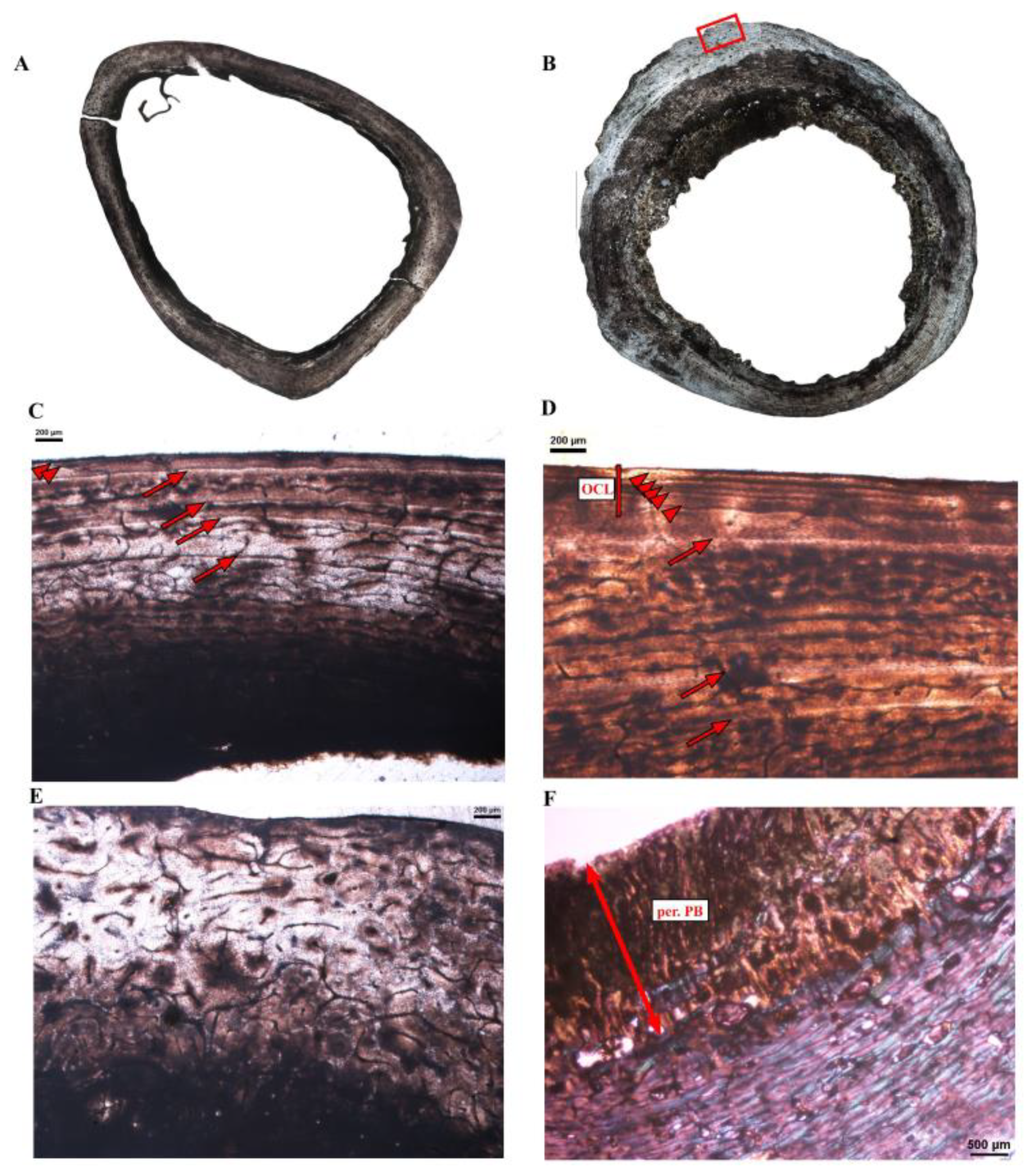
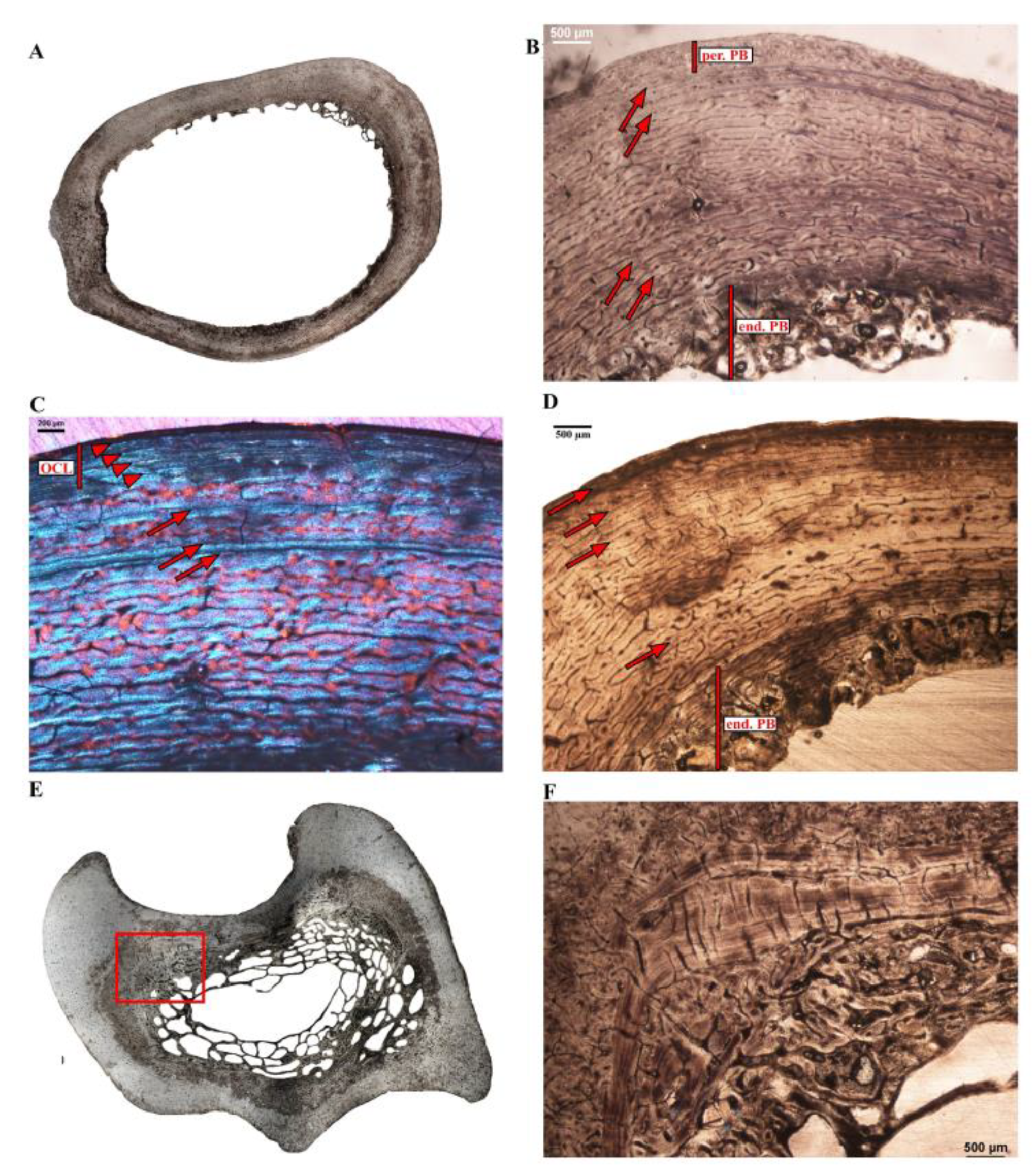
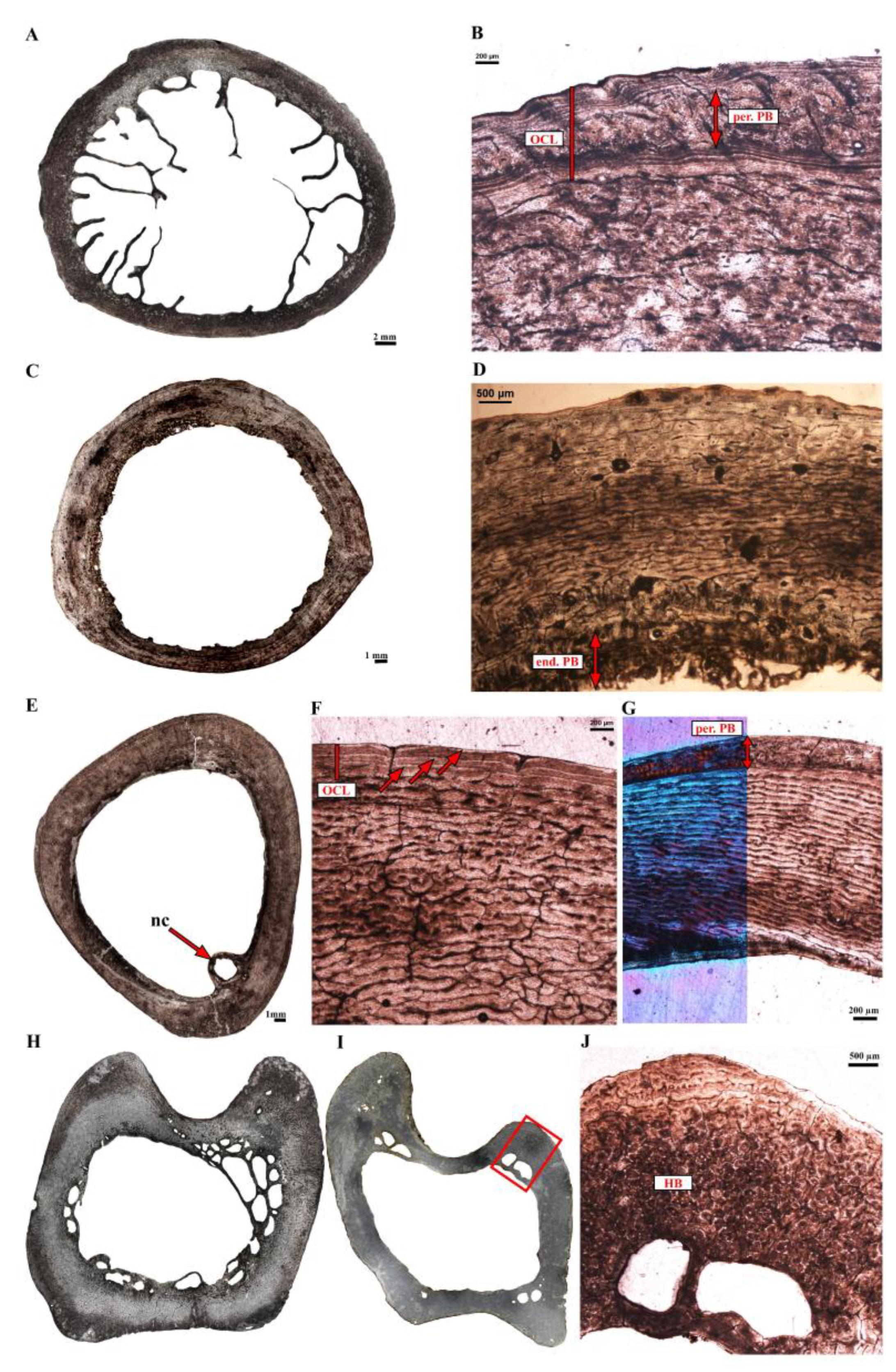
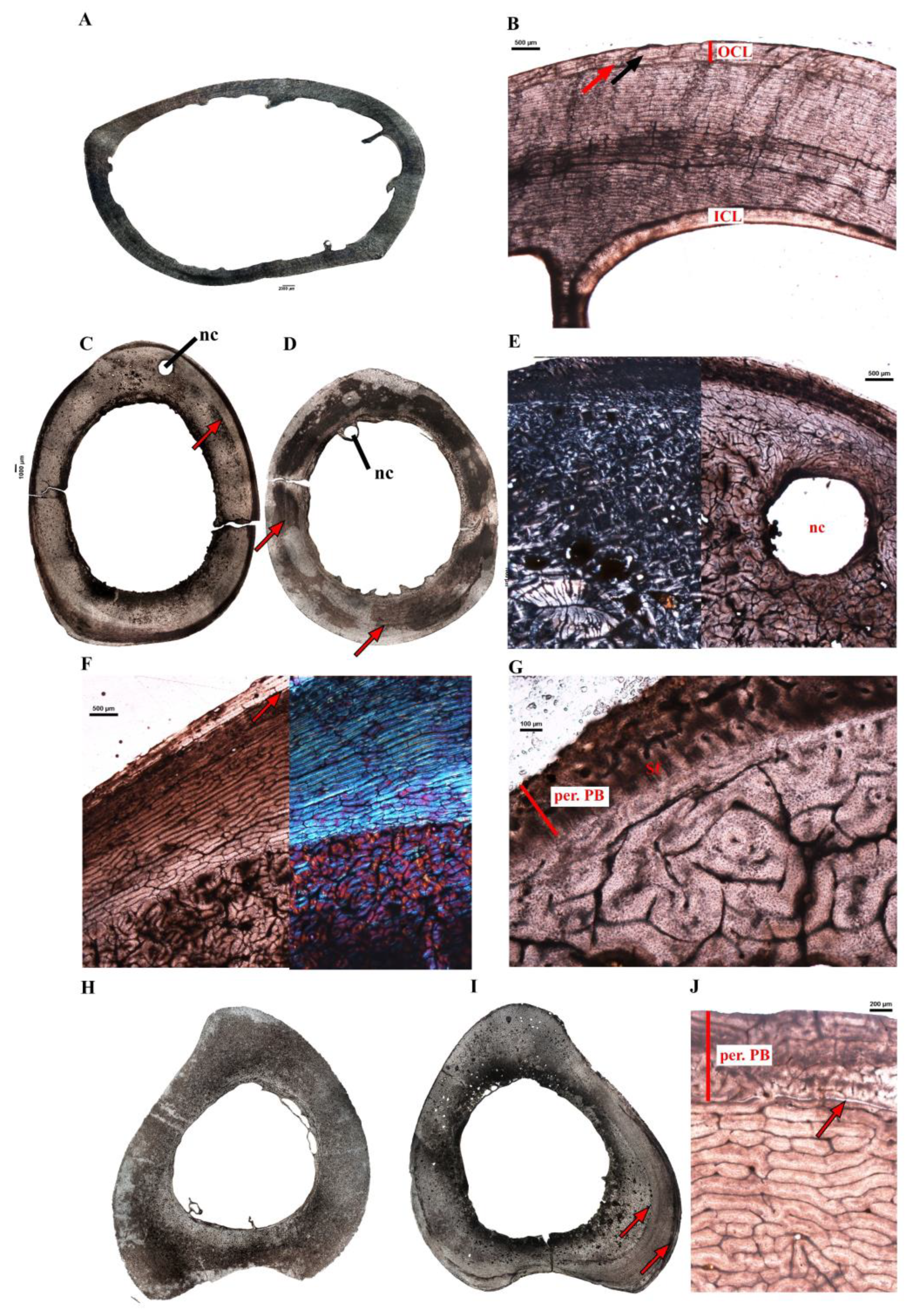
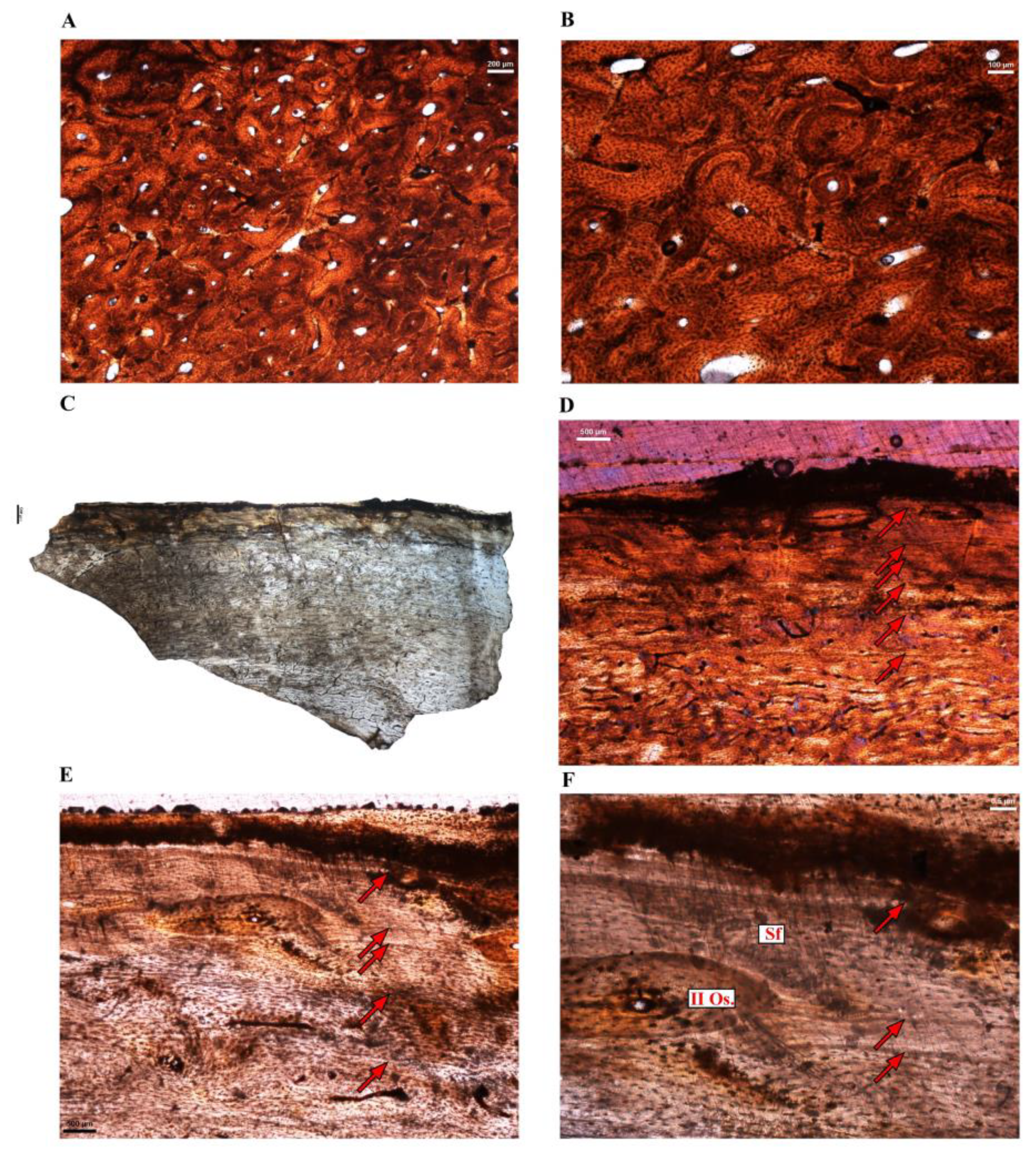
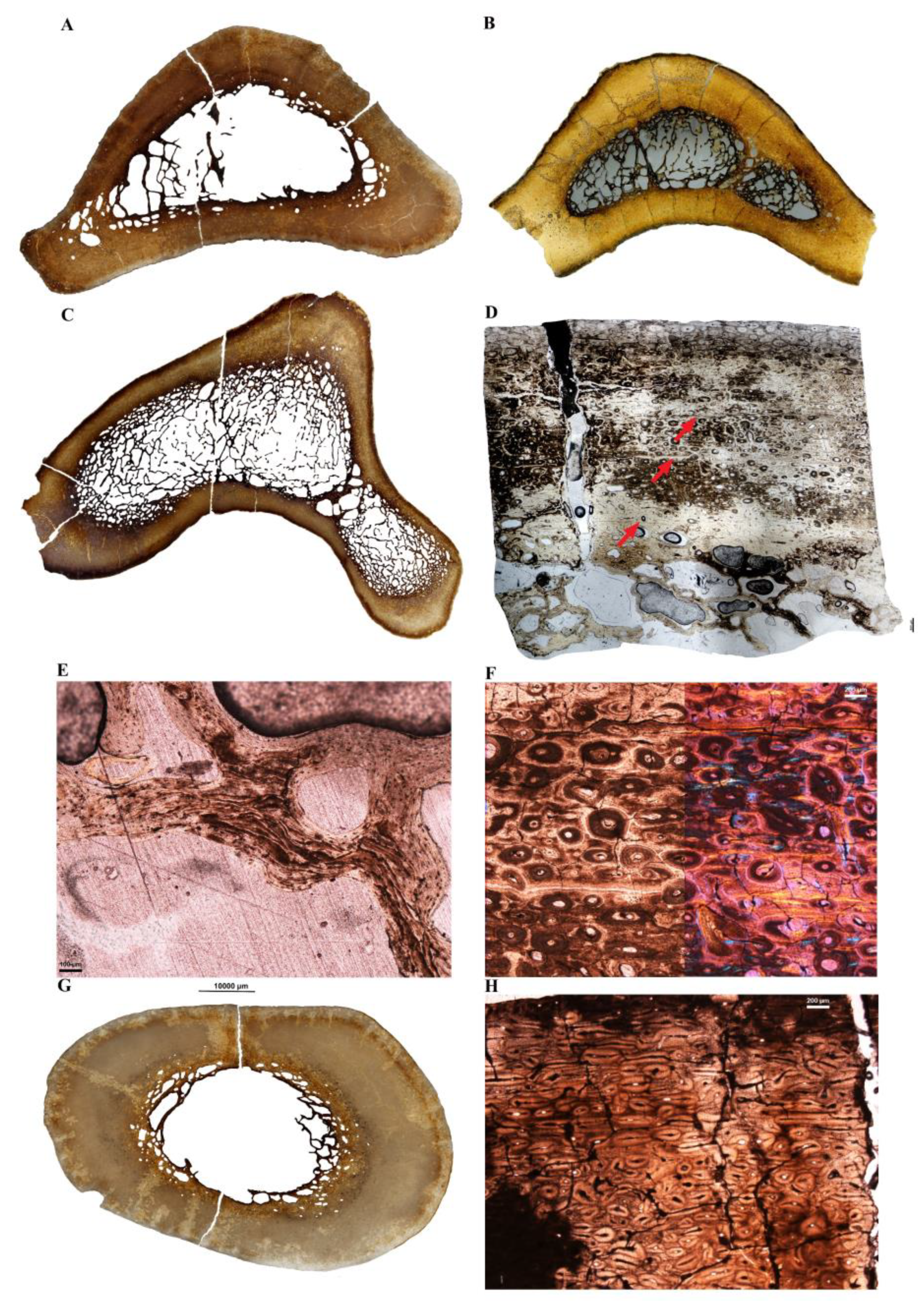
3.2. Microanatomical Observations
3.3. Histological Descriptions
3.3.1. Extant Ratites
3.3.2. Extinct Terrestrial Birds
4. Discussion
4.1. Limb Bone Proportions
4.2. Long Bone Microanatomy and Locomotor Patterns in Large Terrestrial Birds
4.2.1. Interskeletal Element Variability
4.2.2. Differences between Cursorial Ratites and Extinct Terrestrial Birds
4.3. New Data on the Long Bone Histology of Extant Ratites
4.3.1. Interskeletal Element Variability
4.3.2. Growth Marks in the Cortex of Extant Ratites
4.3.3. Comments on OCL Formation and Ontogenetic Status in Birds
4.4. New Data on the Long Bone Histology of Extinct Large Terrestrial Birds
4.4.1. Aepyornithidae
4.4.2. Dinornithidae
4.4.3. Gastornis sp.
4.5. Pathologies Evident in Modern Samples
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abourachid, A.; Renous, S. Bipedal locomotion in ratites (Paleognatiform): Examples of cursorial birds. Ibis 2000, 142, 538–549. [Google Scholar] [CrossRef]
- Coombs, W.P. Theoretical aspects of cursorial adaptations in dinosaurs. Q. Rev. Biol. 1978, 53, 393–418. [Google Scholar] [CrossRef]
- Stein, B.R.; Casinos, A. What is a cursorial mammal? J. Zool. 1997, 242, 185–192. [Google Scholar] [CrossRef]
- Carrano, M.T. What, if anything, is a cursor? Categories versus continua for determining locomotor habit in mammals and dinosaurs. J. Zool. 1999, 247, 29–42. [Google Scholar] [CrossRef]
- Storer, R. Adaptive radiation in birds. In Biology and Comparative Physiology of Birds; Academic Press: New York, NY, USA, 1960; Volume 1, pp. 15–55. [Google Scholar]
- Alexander, R.; Maloiy, G.M.O.; Njau, R.; Jayes, A.S. Mechanics of running of the ostrich (Struthio camelus). J. Zool. 1979, 187, 169–178. [Google Scholar] [CrossRef]
- Picasso, M.B.J. Postnatal ontogeny of the locomotor skeleton of a cursorial bird: Greater rhea. J. Zool. 2012, 286, 303–311. [Google Scholar] [CrossRef]
- Gilbert, M.M.; Snively, E.; Cotton, J. The tarsometatarsus of the ostrich Struthio camelus: Anatomy, bone densities, and structural mechanics. PLoS ONE 2016, 11, e0149708. [Google Scholar] [CrossRef] [Green Version]
- Angst, D.; Buffetaut, E.; Lecuyer, C.; Amiot, R. A new method for estimating locomotion type in large ground birds. Palaeontology 2015, 59, 217–223. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D. The giant flightless bird Gargantuavis philoinos from the Late Cretaceous of southwestern Europe: A review. In Cretaceous Period: Biotic Diversity and Biogeography; Khosla, A., Lucas, S.G., Eds.; NMMNH&S Bulletin: Albuquerque, NM, USA, 2016; Volume 71, pp. 45–50. [Google Scholar]
- Gregory, W.K. Notes on the principles of quadrupedal locomotion and on the mechanism of the limbs in hoofed animals. Ann. N. Y. Acad. Sci. 1912, 22, 267–294. [Google Scholar] [CrossRef]
- Smith, J.M.; Savage, R.J.G. Some locomotory adaptations in mammals. Zool. J. Linn. Soc. 1956, 42, 603–622. [Google Scholar] [CrossRef]
- Murray, P.F.; Vickers-Rich, P. Magnificent Mihirungs: The Colossal Flightless Birds of the Australian Dreamtime (Life of the Past); Indiana University Press: Bloomington, IN, USA, 2004; 416p. [Google Scholar]
- Buffetaut, E.; Angst, D. Stratigraphic distribution of large flightless birds in the Palaeogene of Europe and its palaeobiological and palaeogeographical implications. Earth Sci. Rev. 2014, 138, 394–408. [Google Scholar] [CrossRef]
- Handley, W.D.; Chinsamy, A.; Yates, A.M.; Worthy, T.H. Sexual dimorphism in the late Miocene mihirung Dromornis stirtoni (Aves: Dromornithidae) from the Alcoota Local Fauna of central Australia. J. Vertebr. Paleontol. 2016, 36, e1180298. [Google Scholar] [CrossRef]
- Worthy, T.H.; Handley, W.D.; Archer, M.; Hand, S.J. The extinct flightless mihirungs (Aves, Dromornithidae): Cranial anatomy, a new species, and assessment of Oligo-Miocene lineage diversity. J. Vertebr. Paleontol. 2016, 36, e1031345. [Google Scholar] [CrossRef]
- Worthy, T.H.; Degrange, F.J.; Handley, W.D.; Lee, M.S. The evolution of giant flightless birds and novel phylogenetic relationships for extinct fowl (Aves, Galloanseres). R. Soc. Open Sci. 2017, 4, 170975. [Google Scholar] [CrossRef] [Green Version]
- Chinsamy-Turan, A.; Worthy, T.H.; Handley, W. Growth strategies linked to prevailing environmental conditions in Australian giant flightless mihirung birds (Aves: Dromornithidae). J. Vertebr. Palaeontol. 2019, 79. [Google Scholar]
- Mitchell, K.J.; Llamas, B.; Soubrier, J.; Rawlence, N.J.; Worthy, T.H.; Wood, J.; Lee, M.S.Y.; Cooper, A. Ancient DNA reveals elephant birds and kiwi are sister taxa and clarifies ratite bird evolution. Science 2014, 344, 898–900. [Google Scholar] [CrossRef] [Green Version]
- Martin, L.D. The status of the late Paleocene birds Gastornis and Remiornis. Nat. Hist. Mus. Los Angeles Cty. Sci. Ser. 1992, 36, 97–108. [Google Scholar]
- Canoville, A.; De Buffrénil, V.; Laurin, M. Bone Microanatomy and Lifestyle in Tetrapods. In Vertebrate Skeletal Histology and Paleohistology; De Buffrénil, V., De Ricqlès, A.J., Zylberberg, L., Padian, K., Eds.; CRC Press: Boca Raton, FL, USA, 2021; pp. 724–743. [Google Scholar]
- Currey, J.D.; Alexander, R. The thickness of the walls of tubular bones. J. Zool. 1985, 206, 453–468. [Google Scholar] [CrossRef]
- Canoville, A.; Laurin, M. Evolution of humeral microanatomy and lifestyle in amniotes, and some comments on palaeobiological inferences. Biol. J. Linn. Soc. 2010, 100, 384–406. [Google Scholar] [CrossRef]
- Quemeneur, S.; De Buffrénil, V.; Laurin, M. Microanatomy of the amniote femur and inference of lifestyle in limbed vertebrates. Biol. J. Linn. Soc. 2013, 109, 644–655. [Google Scholar] [CrossRef] [Green Version]
- Houssaye, A.; Waskow, K.; Hayashi, S.; Cornette, R.; Lee, A.H.; Hutchinson, J.R. Biomechanical evolution of solid bones in large animals: A microanatomical investigation. Biol. J. Linn. Soc. 2016, 117, 350–371. [Google Scholar] [CrossRef] [Green Version]
- Legendre, L.J.; Bourdon, E.; Scofield, R.P.; Tennyson, A.J.; Lamrous, H.; Ricqlès, A.; Cubo, J. Bone histology, phylogeny, and palaeognathous birds (Aves: Palaeognathae). Biol. J. Linn. Soc. 2014, 112, 688–700. [Google Scholar] [CrossRef] [Green Version]
- Chinsamy, A.; Elzanowski, A. Bone histology: Evolution of growth pattern in birds. Nature 2001, 412, 402–403. [Google Scholar] [CrossRef] [PubMed]
- Chinsamy, A. Histological perspectives on growth in the birds Struthio camelius and Sagittarius serpentarius. Cour. Forsch.-Inst. Senckenberg 1995, 181, 317–323. [Google Scholar]
- Chinsamy, A.; Chiappe, L.M.; Dodson, P. Mesozoic avian bone microstructure: Physiological implications. Paleobiology 1995, 21, 561–574. [Google Scholar] [CrossRef]
- De Ricqlès, A.; Padian, K.; Horner, J.R. The bone histology of basal birds in phylogenetic and ontogenetic perspectives. In New Perspective on the Origin and Evolution of Birds, Proceedings of the International Symposium in Honor of John, H. Ostrom, New Haven, USA, 13–14 February 1999; Gauthier, J., Gall, L.F., Eds.; Yale University Press: New Haven, CT, USA, 2001; pp. 411–426. [Google Scholar]
- Chinsamy-Turan, A. The Microstructure of Dinosaur Bone: Deciphering Biology with Fine-Scale Techniques; Johns Hopkins University Press: Baltimore, MD, USA, 2005; 216p. [Google Scholar]
- Chinsamy, A.; Buffetaut, E.; Canoville, A.; Angst, D. Insight into the growth dynamics and systematic affinities of the Late Cretaceous Gargantuavis from bone microstructure. Naturwissenschaften 2014, 101, 447–452. [Google Scholar] [CrossRef]
- Chinsamy, A.; Angst, D.; Canoville, A.; Göhlich, U.B. Bone histology yields insights into the biology of the extinct elephant birds (Aepyornithidae) from Madagascar. Biol. J. Linn. Soc. 2020, 130, 268–295. [Google Scholar] [CrossRef]
- Atterholt, J.; Poust, A.W.; Erickson, G.M.; O’Connor, J.K. Intraskeletal osteohistovariability reveals complex growth strategies in a Late Cretaceous enantiornithine. Front. Earth Sci. 2021, 9, 118. [Google Scholar] [CrossRef]
- Bourdon, E.; Castanet, J.; De Ricqlès, A.; Scofield, P.; Tennyson, A.; Lamrous, H.; Cubo, J. Bone growth marks reveal protracted growth in New Zealand kiwi (Aves, Apterygidae). Biol. Lett. 2009, 5, 639–642. [Google Scholar] [CrossRef]
- Van Soest, R.W.M.; Van Utrecht, W.L. The layered structure of bones of birds as a possible indication of age. Bijdr Dierkd. 1971, 41, 61–66. [Google Scholar] [CrossRef] [Green Version]
- Ponton, F.; Elżanowski, A.; Castanet, J.; Chinsamy, A.; De Margerie, E.; De Ricqlès, A.; Cubo, J. Variation of the outer circumferential layer in the limb bones of birds. Acta Ornithol. 2004, 39, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Horner, J.R.; De Ricqlès, A.; Padian, K. Long bone histology of the hadrosaurid dinosaur Maiasaura peeblesorum: Growth dynamics and physiology based on an ontogenetic series of skeletal elements. J. Vertebr. Paleontol. 2000, 20, 115–129. [Google Scholar] [CrossRef]
- Padian, K.; Lamm, E.T. Bone Histology of Fossil Tetrapods: Advancing Methods, Analysis, and Interpretation; University of California Press: Oakland, CA, USA, 2013; 298p. [Google Scholar]
- Angst, D.; Chinsamy, A.; Steel, L.; Hume, J.P. Bone histology sheds new light on the ecology of the dodo (Raphus cucullatus, Aves, Columbiformes). Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foote, J.S. A Contribution to the Comparative Histology of the Femur; Smithsonian Contributions to Knowledge: Washington, DC, USA, 1916; 242p. [Google Scholar]
- Houde, P. Histological evidence for the systematic position of Hesperornis (Odontornithes: Hesperornithiformes). Auk 1987, 104, 125–129. [Google Scholar] [CrossRef]
- Castanet, J.; Rogers, K.C.; Cubo, J.; Jacques-Boisard, J. Periosteal bone growth rates in extant ratites (ostriche and emu). Implications for assessing growth in dinosaurs. Comptes Rendus de l’Académie des Sciences-Series III-Sci. de la Vie 2000, 323, 543–550. [Google Scholar] [CrossRef]
- Padian, K.; De Ricqlès, A.; Horner, J.R. Dinosaurian growth rates and bird origins. Nature 2001, 412, 405–408. [Google Scholar] [CrossRef]
- Turvey, S.T.; Green, O.R.; Holdaway, R.N. Cortical growth marks reveal extended juvenile development in New Zealand moa. Nature 2005, 435, 940–943. [Google Scholar] [CrossRef]
- Amprino, R.; Godina, G. La struttura delle ossa nei vertebrati: Ricerche comparative negli anfibi e negli amnioti. Comment. Pont. Acad. Sei. 1947, 11, 329–464. [Google Scholar]
- De Ricqlès, A.; Bourdon, E.; Legendre, L.J.; Cubo, J. Preliminary assessment of bone histology in the extinct elephant bird Aepyornis (Aves, Palaeognathae) from Madagascar. Comptes Rendus Palevol 2016, 15, 205–216. [Google Scholar] [CrossRef]
- Chinsamy, A.; Angst, D.; Canoville, A.; Göhlich, U. Bone histology and biology of the giant insular extinct bird, Aepyornis maximus. In Abstract Book of the 9th International Meeting of the Society of Avian Paleontology and Evolution; Palaeontological Association: Diamante, Argentina, 2016. [Google Scholar]
- Chinsamy, A.; Worthy, T.H. Histovariability and Palaeobiological Implications of the Bone Histology of the Dromornithid, Genyornis newtoni. Diversity 2021, 13, 219. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D. A femur of the Late Cretaceous giant bird Gargantuavis from Cruzy (southern France) and its systematic implications. Palaeovertebrata 2019, 42, e3. [Google Scholar] [CrossRef]
- Buffetaut, E.; Angst, D. Gargantuavis is an insular basal ornithurine: A comment on Mayr et al., 2020, ‘A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic’. Cretac. Res. 2020, 112, 104438. [Google Scholar] [CrossRef]
- Mayr, G.; Codrea, V.; Solomon, A.; Bordeianu, M.; Smith, T. A well-preserved pelvis from the Maastrichtian of Romania suggests that the enigmatic Gargantuavis is neither an ornithurine bird nor an insular endemic. Cretac. Res. 2020, 106, 104271. [Google Scholar] [CrossRef]
- Steel, L. Bone histology and skeletal pathology of two recently-extinct flightless pigeons: Raphus cucullatus and Pezophaps solitarius. J. Vertebr. Paleontol. 2009, 29, 185. [Google Scholar]
- Starck, J.M.; Chinsamy, A. Bone microstructure and developmental plasticity in birds and other dinosaurs. J. Morphol. 2002, 254, 232–246. [Google Scholar] [CrossRef]
- Buffetaut, E. The giant bird Gastornis in Asia: A revision of Zhongyuanus xichuanensis Hou, 1980, from the Early Eocene of China. Paleontol. J. 2013, 47, 1302–1307. [Google Scholar] [CrossRef]
- Canoville, A.; de Buffrénil, V. Ontogenetic development and intraspecific variability of bone microstructure in the king penguin Aptenodytes patagonicus: Considerations for paleoecological inferences in Sphenisciformes. Anat. Rec. 2016, 299, 270. [Google Scholar]
- Barrat, A. Quelques aspects de la biologie et de l’écologie du manchot royal (Aptenodytes patagonicus) des îles Crozet. CNFRA 1976, 40, 9–52. [Google Scholar]
- Cherel, Y.; Le Maho, Y. Five months of fasting in king penguin chicks: Body mass loss and fuel metabolism. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1985, 249, 387–392. [Google Scholar] [CrossRef]
- Buffetaut, E. New remains of the giant bird Gastornis from the Upper Paleocene of the eastern Paris Basin and the relationships between Gastornis and Diatryma. Neues Jahrbuch für Geologie und Paläontologie-Monatshefte 1997, 3, 179–190. [Google Scholar] [CrossRef]
- Hansford, J.P.; Turvey, S.T. Unexpected diversity within the extinct elephant birds (Aves: Aepyornithidae) and a new identity for the world’s largest bird. R. Soc. Open Sci. 2018, 5, 181295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stein, K.; Sander, M. Histological core drilling: A less destructive method for studying bone histology. In Methods in Fossil Preparation: Proceedings of the First Annual Fossil Preparation and Collections Symposium; Brown, M.A., Kane, J.F., Parker, W.G., Eds.; Petrified Forest: Petrified Forest National Park, AZ, USA, 2009; pp. 69–80. [Google Scholar]
- Redelstorff, R.; Sander, P.M. Long and girdle bone histology of Stegosaurus: Implications for growth and life history. J. Vertebr. Paleontol. 2009, 4, 1087–1099. [Google Scholar] [CrossRef]
- Redelstorff, R.; Hübner, T.R.; Chinsamy, A.; Sander, P.M. Bone histology of the stegosaur Kentrosaurus aethiopicus (Ornithischia: Thyreophora) from the Upper Jurassic of Tanzania. Anat. Rec. 2013, 296, 933–952. [Google Scholar] [CrossRef] [PubMed]
- Canoville, A.; Chinsamy, A. Bone microstructure of pareiasaurs (Parareptilia) from the Karoo Basin, South Africa: Implications for growth strategies and lifestyle habits. Anat. Rec. 2017, 300, 1039–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinsamy, A.; Raath, M.A. Preparation of fossil bone for histological examination. Palaeont. Afr. 1992, 29, 39–44. [Google Scholar]
- Francillon-Vieillot, H.; De Buffrénil, V.; Castanet, J.; Géraudie, J.; Meunier, F.J.; Sire, J.Y.; Zylberberg, L.; De Ricqlés, A. Skeletal biomineralization: Patterns, processes and evolutionary trends. In Microstructure and Mineralization of Vertebrate Skeletal Tissues; Van Nostrand Reinhold: New York, NY, USA, 1990; pp. 471–548. [Google Scholar]
- Girondot, M.; Laurin, M. Bone profiler: A tool to quantify, model, and statistically compare bone-section compactness profiles. J. Vertebr. Paleontol. 2003, 23, 458–461. [Google Scholar] [CrossRef]
- Chinsamy, A.; Tumarkin-Deratzian, A. Pathologic bone tissues in a turkey vulture and a nonavian dinosaur: Implications for interpreting endosteal bone and radial fibrolamellar bone in fossil dinosaurs. Anat. Rec. 2009, 292, 1478–1484. [Google Scholar] [CrossRef]
- Watanabe, J. Ontogeny of surface texture of limb bones in modern aquatic birds and applicability of textural ageing. Anat. Rec. 2018, 301, 1026–1045. [Google Scholar] [CrossRef] [Green Version]
- Heck, C.T.; Woodward, H.N. Intraskeletal bone growth patterns in the North Island Brown Kiwi (Apteryx mantelli): Growth mark discrepancy and implications for extinct taxa. J. Anat. 2021, 239, 1075–1095. [Google Scholar] [CrossRef]
- Phillips, M.J.; Gibb, G.C.; Crimp, E.A.; Penny, D. Tinamous and moa flock together: Mitochondrial genome sequence analysis reveals independent losses of flight among ratites. Syst. Biol. 2010, 59, 90–107. [Google Scholar] [CrossRef] [Green Version]
- Baker, A.J.; Haddrath, O.; McPherson, J.D.; Cloutier, A. Genomic support for a moa–tinamou clade and adaptive morphological convergence in flightless ratites. Mol. Biol. Evol. 2014, 31, 1686–1696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinsamy, A.; Marugán-Lobón, J.; Serrano, F.J.; Chiappe, L.M. Life history traits and biology of the basal pygostylian, Confuciusornis sanctus. Anat. Rec. 2020, 303, 949–962. [Google Scholar] [CrossRef] [PubMed]
- Woodward, H.N.; Tremaine, K.; Williams, S.A.; Zanno, L.E.; Horner, J.R.; Myhrvold, N. Growing up Tyrannosaurus rex: Osteohistology refutes the pygmy “Nanotyrannus” and supports ontogenetic niche partitioning in juvenile Tyrannosaurus. Sci. Adv. 2020, 6, eaax6250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enlow, D.H.; Brown, S.O. A comparative histological study of fossil and recent bone tissues. Part II. Tex. J. Sci. 1957, 9, 186–204. [Google Scholar]
- McFarlin, S.C.; Terranova, C.J.; Zihlman, A.L.; Enlow, D.H.; Bromage, T.G. Regional variability in secondary remodeling within long bone cortices of catarrhine primates: The influence of bone growth history. J. Anat. 2008, 213, 308–324. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | Sample Number | Bone | MNHN Collection Number | Sample Type | Lg (mm) | Dm (mm) | Lg/Dm | Cm (mm) |
|---|---|---|---|---|---|---|---|---|
| Extant palaeognaths | ||||||||
| Casuarius casuarius | Cc-fm-1 | Femur (l) | 1923-879 SS-S.B.S2.M1.C1 | CT | 224 | - | - | 87 |
| Casuarius casuarius | Cc-fm-2 | Femur (r) | 1923-879 SS-S.B.S2.M1.C1 | CT | - | - | - | - |
| Casuarius casuarius | Cc-tb-1 | Tibiotarsus (r) | 1923-877 SS-S.B.S2.M1.C1 | CT | 389 | - | - | 78 |
| Casuarius casuarius | Cc-tb-2 | Tibiotarsus (l) | 1923-877 SS-S.B.S2.M1.C1 | CT | 386 | - | - | 78 |
| Casuarius casuarius | Cc-tm-1 | Tarsometatarsus (l) | 1923-878 SS-S.B.S2.M1.C1 | CT | 295 | 22.8 | 12.94 | 76 |
| Casuarius casuarius | Cc-tm-2 | Tarsometatarsus (l) | 1923-878 | CT | 290 | 23.2 | 12.5 | 74 |
| Dromaius novaehollandiae | Dn-fm-1 | Femur (r) | 1923-900 SS-S.B.S2.M1.C23 | CT | 221 * | - | - | 95 * |
| Dromaius novaehollandiae | Dn-fm-2 | Femur (l) | 1923-897 SS-S.B.S2.M1.C23 | CT | 234 | - | - | 99 |
| Dromaius novaehollandiae | Dn-tb-1 | Tibiotarsus (l) | 1923-899 SS-S.B.S2.M1.C23 | CT | 456 | - | - | 91 |
| Dromaius novaehollandiae | Dn-tb-2 | Tibiotarsus (l) | 1923-899 SS-S.B.S2.M1.C23 | CT | 454 | - | - | 87 |
| Dromaius novaehollandiae | Dn-tm-1 | Tarsometatarsus (l) | 1923-898 SS-S.B.S2.M1.C23 | CT | 387 | 17.92 | 21.60 | 76 |
| Dromaius novaehollandiae | Dn-tm-2 | Tarsometatarsus (r) | 1923-898 SS-S.B.S2.M1.C23 | CT | 400 | 19.28 | 20.75 | 84 |
| Rhea americana | Ra-fm-1 | Femur (l) | 1920-116 | CT | - | - | - | - |
| Rhea americana | Ra-fm-2 | Femur (l) | - | CT | 186 | - | - | 60 |
| Rhea americana | Ra-tb-1 | Tibiotarsus (r) | - | CT | 286 | - | - | 55 |
| Rhea americana | Ra-tb-2 | Tibiotarsus (l) | 1920-116 | CT | 354 | - | - | 73 |
| Rhea americana | Ra-tm-1 | Tarsometatarsus (l) | 1920-116 | CT | 316 | 15.64 | 20.20 | 58 |
| Rhea americana | Ra-tm-2 | Tarsometatarsus (l) | - | CT | 272 | 13.15 | 20.68 | 48 |
| Struthio camelus | Sc-fm-1 | Femur (l) | A4751 SS-S.B.S2.M1.C21 | CT | 301 * | - | - | 140 * |
| Struthio camelus | Sc-fm-2 | Femur (l) | - | CT | 327 | - | - | 140.5 |
| Struthio camelus | Sc-tb-1 | Tibiotarsus (l) | - | CT | 597 | - | - | 109 |
| Struthio camelus | Sc-tb-2 | Tibiotarsus (r) | A-5-100 SS-S.B.S2.M1.C21 | CT | 470 | - | - | 97 |
| Struthio camelus | Sc-tm-1 | Tarsometatarsus (r) | - | CT | 514 | 32.95 | 15.60 | 97 |
| Struthio camelus | Sc-tm-2 | Tarsometatarsus (l) | - | CT | 460 | 28.90 | 15.92 | 85 |
| Extinct palaeognaths | - | |||||||
| Vorombe titan | Ae-fm-1 | Femur (r) | 1937-62 | BC | 360 | - | - | - |
| Aepyornis maximus | Ae-fm-2 | Femur (l) | MAD367 1906-17 | CT | 380 | - | - | 244 |
| Aepyornis maximus | Ae-fm-3 | Femur (r) | MAD 365 | CT | 350 | - | - | 244 |
| Vorombe titan | Ae-tb-1 | Tibiotarsus | 1910-12 | CT | - | - | - | 190 |
| Aepyornithidae indet. | Ae-tb-2 | Tibiotarsus pe (l) | - | CT | - | - | - | - |
| Aepyornithidae indet. | Ae-tb-3 | Tibiotarsus pe (r) | - | CT | - | - | - | - |
| Aepyornithidae indet. | Ae-tb-4 | Tibiotarsus (l) | 1937-62 | BC | 612 | - | - | - |
| Vorombe titan | Ae-tb-5 | Tibiotarsus (r) | 1937-62 | BC | 645 | - | - | - |
| Vorombe titan | Ae-tb-6 | Tibiotarsus (l) | 1937-62 | BC | 670 | - | - | - |
| Aepyornithidae indet. | Ae-tb-7 | Tibiotarsus (r) | 1937-62 | BC | 615 | - | - | - |
| Vorombe titan | Ae-tb-8 | Tibiotarsus (l) | 1937-62 | BC | 760 | - | - | - |
| Aepyornithidae indet. | Ae-tm-1 | Tarsometatarsus (l) | 1906-16 Mr Belo | CT | 430 | 81.8 | 5.26 | 190 |
| Aepyornithidae indet. | Ae-tm-2 | Tarsometatarsus (l) | 1937-62 | BC | 481 | 88.03 | 5.46 | - |
| Dinornis sp. | Di-fm-1 | Femur (l) | - | CT | 315 | - | - | 182 |
| Dinornis sp. | Di-tb-1 | Tibiotarsus (l) | - | CT | - | - | - | 160 ** |
| Dinornithidae indet. | Di-tb-3 | Tibiotarsus (l) | - | CT | - | - | - | - |
| Dinornithidae indet. | Di-tb-4 | Tibiotarsus (?) | - | CT | - | - | - | - |
| Dinornis sp. | Di-tm-1 | Tarsometatarsus (l) | - | CT | 235 | 42 | 5.60 | 107 |
| Extinct neognaths | ||||||||
| Gastornis sp. | Ga-fm-1 | Fem/tibiotarus (?) | Mu12734 | Frag. | - | - | - | - |
| Gastornis sp. | Ga-fm-2 | Femur (r) | BR12419 | BC | - | - | - | - |
| Gastornis sp. | Ga-fm-3 | Femur (r) | BR12420 | Frag. | - | - | - | - |
| Gastornis sp. | Ga-tb-1 | Tibiotarsus (?) | - | Frag. | - | - | - | - |
| Gastornis sp. | Ga-tb-2 | Tibiotarsus (?) | CRL2457 | BC | - | - | - | - |
| Species | Locomotion Type | Sampling Number | MD (mm) | Comp. | Bone Trabeculae |
|---|---|---|---|---|---|
| Casuarius casuarius | Cursorial 1 | Cc-fm-1 | 29.0 | 0.395 | NO |
| Dromaius novaehollandiae | Cursorial 1 | Dn-fm-2 | 32.7 | 0.389 | YES |
| Dn-tb-2 | 28.5 | 0.500 | NO | ||
| Dn-tm-1 | - | 0.655 | YES | ||
| Dn-tm-2 | - | 0.560 | NO | ||
| Rhea americana | Cursorial 1 | Ra-fm-1 | - | 0.656 | NO |
| Ra-fm-2 | 22.6 | 0.404 | NO | ||
| Ra-tb-1 | 17.0 | 0.459 | NO | ||
| Ra-tb-2 | 21.6 | 0.644 | NO | ||
| Ra-tm-1 | 20.3 | 0.826 | NO | ||
| Ra-tm-2 | 16.5 | 0.618 | NO | ||
| Struthio camelus | Cursorial 1 | Sc-fm-1 | 46.3 | 0.392 | NO |
| Sc-fm-2 | 55.1 | 0.289 | YES (broken) | ||
| Sc-tb-1 | 37.2 | 0.593 | NO | ||
| Sc-tm-2 | 28.6 | 0.702 | NO | ||
| Aepyornithidae | Graviportal 2,3 | Ae-fm-2 | 96.5 | 0.512 | YES |
| Ae-fm-3 | 82.4 | 0.386 | YES | ||
| Ae-tb-1 | 71.6 | 0.706 | YES (broken) | ||
| Ae-tm-1 | 76.7 | 0.785 | YES (broken) | ||
| Dinornithidae | Graviportal 2 | Di-fm-1 | 55.8 | 0.741 | NO |
| Di-tb-3 | 68.5 | 0.733 | YES (broken) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Canoville, A.; Chinsamy, A.; Angst, D. New Comparative Data on the Long Bone Microstructure of Large Extant and Extinct Flightless Birds. Diversity 2022, 14, 298. https://doi.org/10.3390/d14040298
Canoville A, Chinsamy A, Angst D. New Comparative Data on the Long Bone Microstructure of Large Extant and Extinct Flightless Birds. Diversity. 2022; 14(4):298. https://doi.org/10.3390/d14040298
Chicago/Turabian StyleCanoville, Aurore, Anusuya Chinsamy, and Delphine Angst. 2022. "New Comparative Data on the Long Bone Microstructure of Large Extant and Extinct Flightless Birds" Diversity 14, no. 4: 298. https://doi.org/10.3390/d14040298
APA StyleCanoville, A., Chinsamy, A., & Angst, D. (2022). New Comparative Data on the Long Bone Microstructure of Large Extant and Extinct Flightless Birds. Diversity, 14(4), 298. https://doi.org/10.3390/d14040298