
Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Species and Distribution Data
2.2. Study Site and Treatments
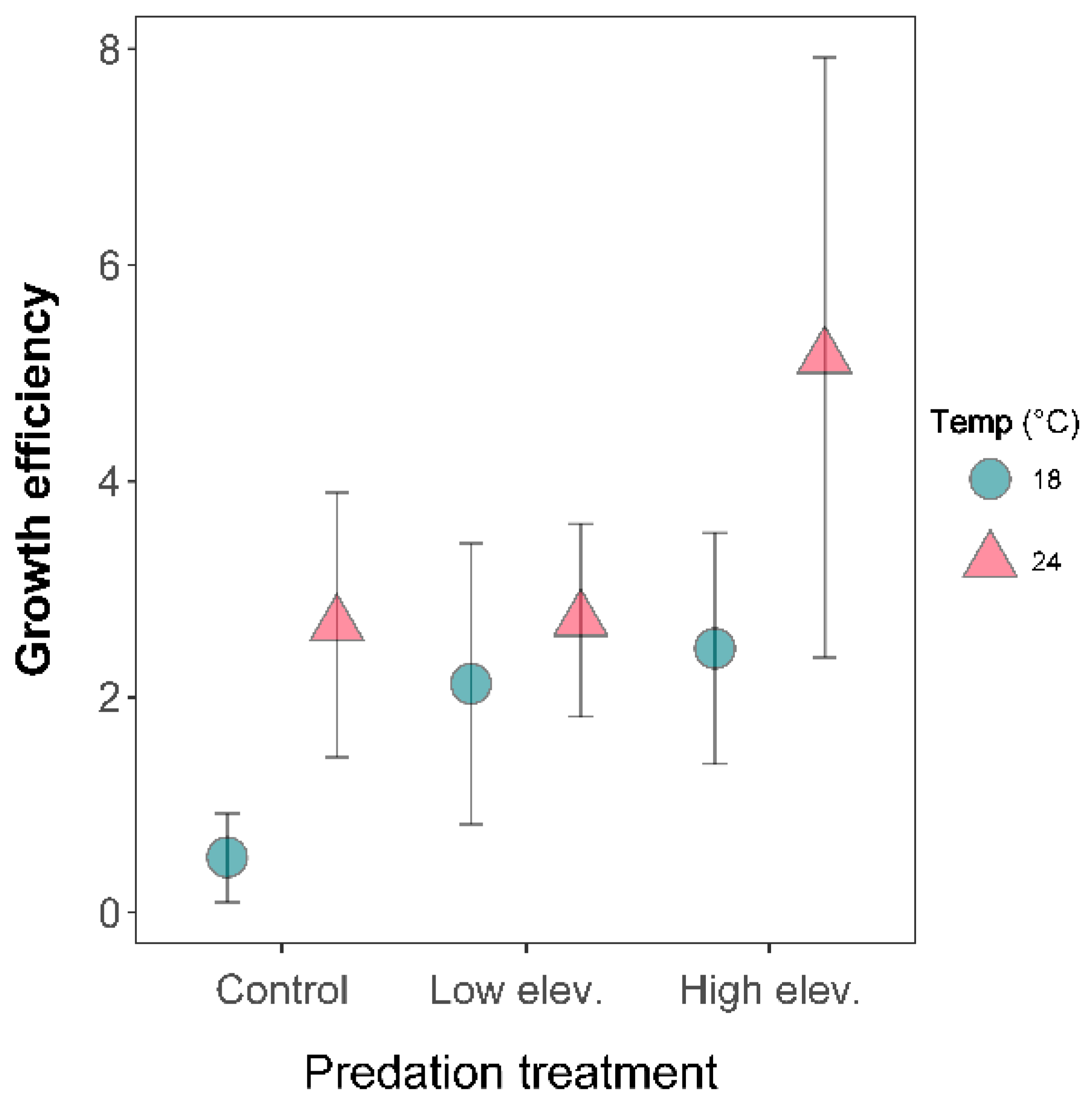
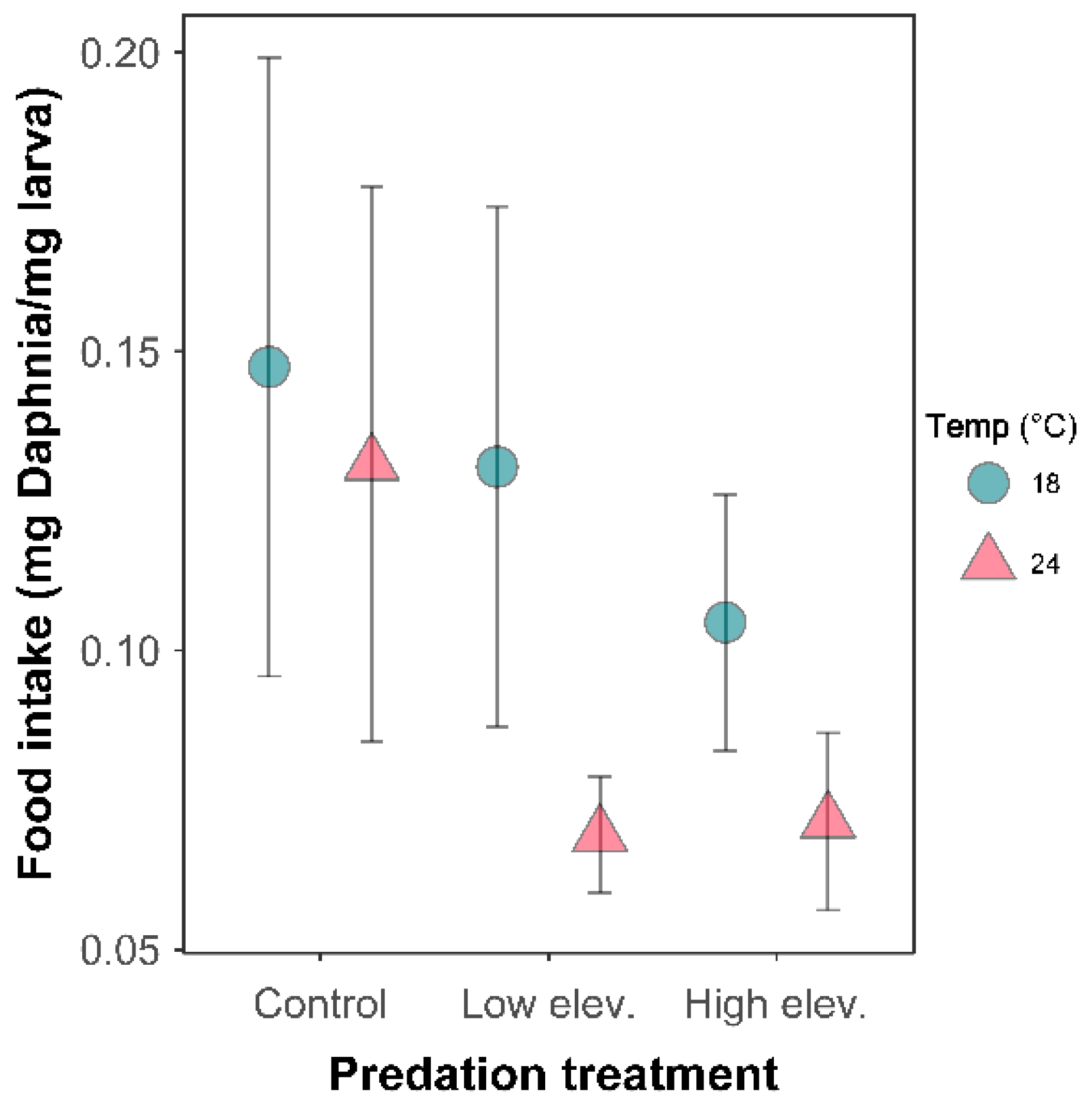
2.3. Growth Efficiency and Food Intake
2.4. Morphological Defense
2.5. Statistical Analyses
3. Results
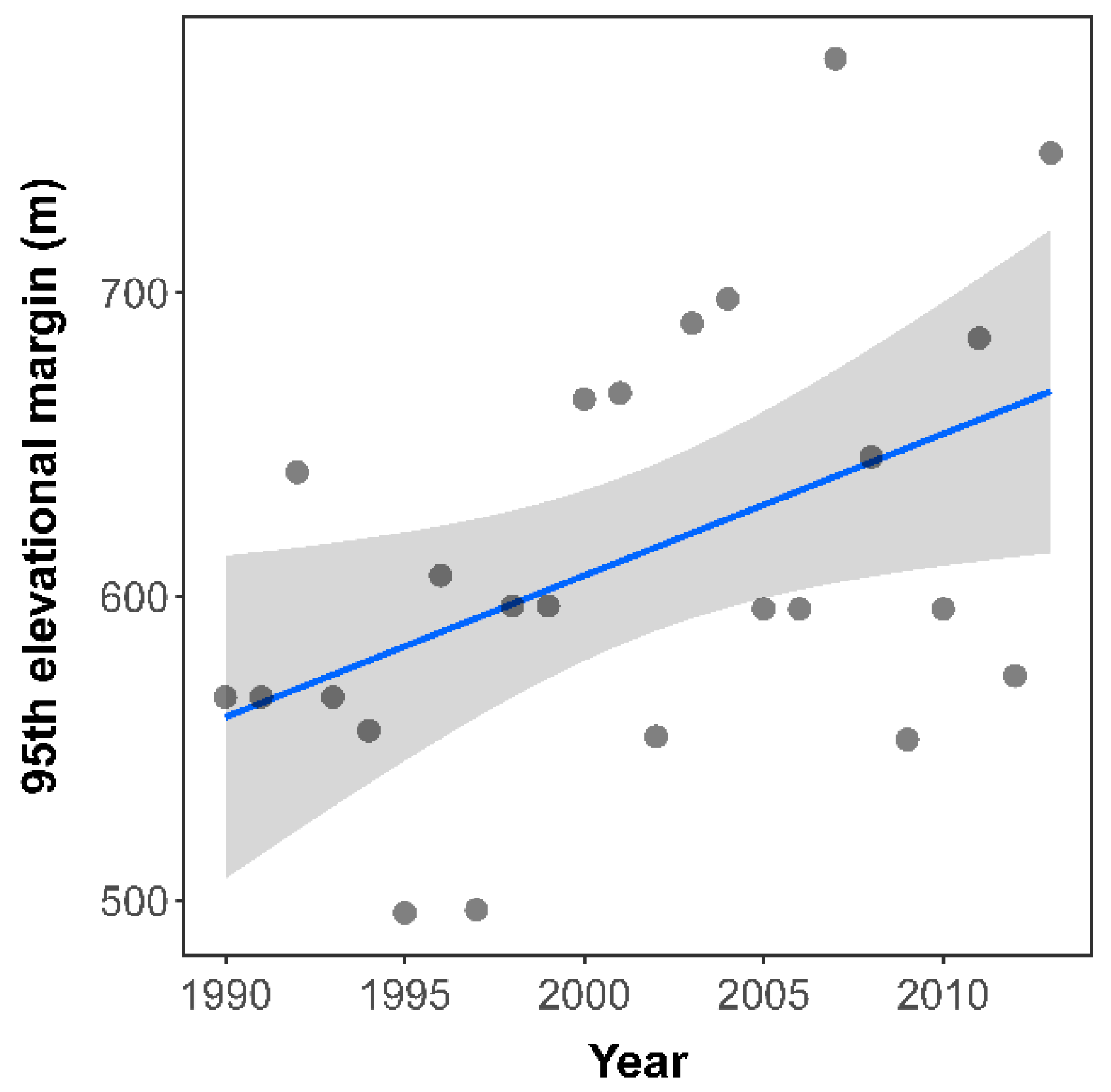
3.1. Elevational Range Shift
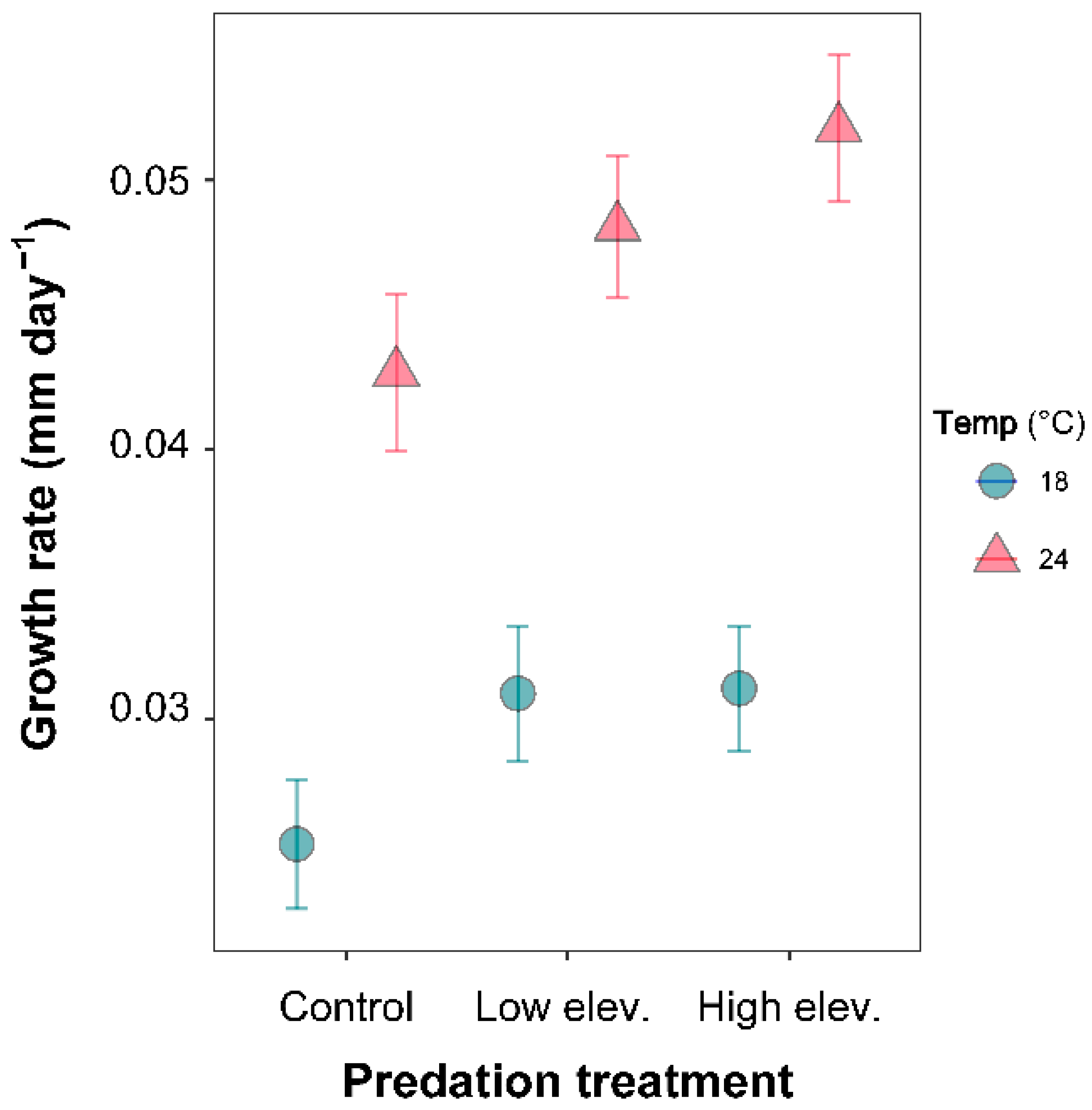
3.2. Larval Growth
3.3. Behavioral Response
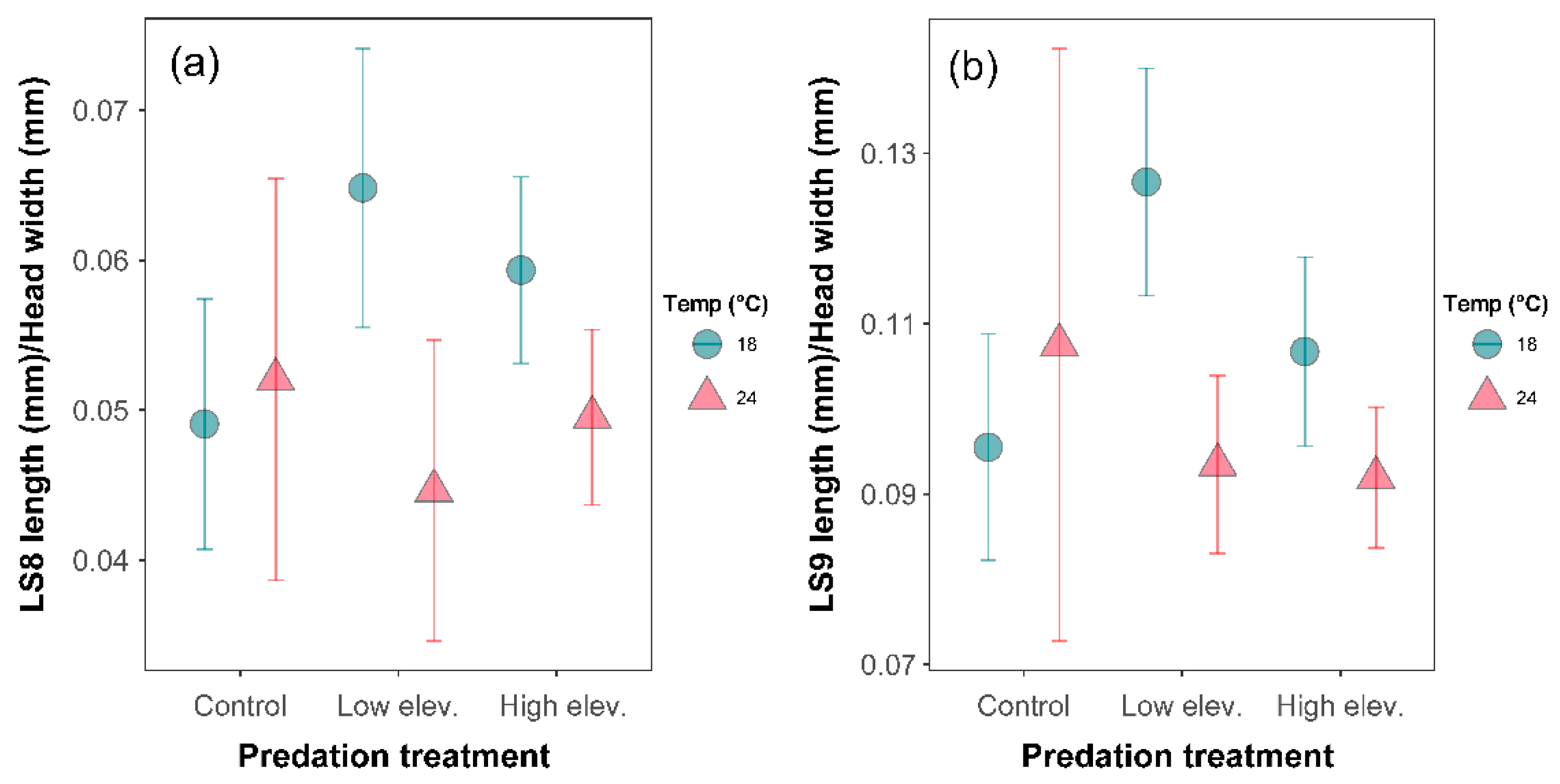
3.4. Morphological Response
4. Discussion
4.1. Range Shift
4.2. Temperature Effects
4.3. Predation Effects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sih, A. Predators and prey life style. In Predation: Direct and Indirect Impacts on Aquatic Communities; Kerfoot, W.C., Sih, A., Eds.; University Press of New England: Hanover, UK, 1987. [Google Scholar]
- Werner, E.E.; Anholt, B.R. Ecological consequences of the trade-off between growth and mortality rates mediated by foraging activity. Am. Nat. 1993, 142, 242–272. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.; Dingemanse, N.; Hankison, S.; Langenhof, M.; Rollins, K. Early exposure to nonlethal predation risk by size-selective predators increases somatic growth and decreases size at adulthood in threespined sticklebacks. J. Evol. Biol. 2011, 24, 943–953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Block, M.; Pauwels, K.; Van Den Broeck, M.; De Meester, L.; Stoks, R. Local genetic adaptation generates latitude-specific effects of warming on predator–prey interactions. Glob. Chang. Biol. 2013, 19, 689–696. [Google Scholar] [CrossRef] [PubMed]
- Stoks, R.; Swillen, I.; De Block, M. Behaviour and physiology shape the growth accelerations associated with predation risk, high temperatures and southern latitudes in Ischnura damselfly larvae. J. Anim. Ecol. 2012, 81, 1034–1040. [Google Scholar] [CrossRef]
- Richardson, J.M. A comparative study of phenotypic traits related to resource utilization in anuran communities. Evol. Ecol. 2002, 16, 101–122. [Google Scholar] [CrossRef]
- Stoks, R.; McPeek, M.; Mitchell, J. Evolution of prey behavior in response to changes in predation regime: Damselflies in fish and dragonfly lakes. Evolution 2003, 57, 574–585. [Google Scholar] [CrossRef]
- Conover, D.O.; Schultz, E.T. Phenotypic similarity and the evolutionary significance of countergradient variation. Trends Ecol. Evol. 1995, 10, 248–252. [Google Scholar] [CrossRef]
- Wellborn, G.A.; Skelly, D.K.; Werner, E.E. Mechanisms creating community structure across a freshwater habitat gradient. Annu. Rev. Ecol. Evol. Syst. 1996, 27, 337–363. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Ryrholm, N.; Stefanescu, C.; Hill, J.K.; Thomas, C.D.; Descimon, H.; Huntley, B.; Kaila, L.; Kullberg, J.; Tammaru, T. Poleward shifts in geographical ranges of butterfly species associated with regional warming. Nature 1999, 399, 579. [Google Scholar] [CrossRef]
- Hassall, C.; Thompson, D.J. Accounting for recorder effort in the detection of range shifts from historical data. Methods Ecol. Evol. 2010, 1, 343–350. [Google Scholar] [CrossRef]
- Lima, S.L.; Dill, L.M. Behavioral decisions made under the risk of predation: A review and prospectus. Can. J. Zool. 1990, 68, 619–640. [Google Scholar] [CrossRef]
- Antoł, A.; Sniegula, S. Damselfly eggs alter their development rate in the presence of an invasive alien cue but not a native predator cue. Ecol. Evol. 2021, 11, 9361–9369. [Google Scholar] [CrossRef] [PubMed]
- Blanckenhorn, W.; Demont, M. Bergmann and converse Bergmann latitudinal clines in arthropods: Two ends of a continuum? Integr. Comp. Biol. 2004, 44, 413–424. [Google Scholar] [CrossRef] [Green Version]
- Stoks, R.; Block, M.D.; McPeek, M.A. Physiological costs of compensatory growth in a damselfly. Ecology 2006, 87, 1566–1574. [Google Scholar] [CrossRef]
- Stoks, R.; Block, M.D.; Van De Meutter, F.; Johansson, F. Predation cost of rapid growth: Behavioural coupling and physiological decoupling. J. Anim. Ecol. 2005, 74, 708–715. [Google Scholar] [CrossRef]
- McPeek, M.A. The growth/predation risk trade-off: So what is the mechanism? Am. Nat. 2004, 163, E88–E111. [Google Scholar] [CrossRef]
- McPeek, M.A.; Grace, M.; Richardson, J.M. Physiological and behavioral responses to predators shape the growth/predation risk trade-off in damselflies. Ecology 2001, 82, 1535–1545. [Google Scholar] [CrossRef]
- Thaler, J.S.; McArt, S.H.; Kaplan, I. Compensatory mechanisms for ameliorating the fundamental trade-off between predator avoidance and foraging. Proc. Natl. Acad. Sci. USA 2012, 109, 12075–12080. [Google Scholar] [CrossRef] [Green Version]
- Mikolajewski, D.J.; Rolff, J. Benefits of morphological defence demonstrated by direct manipulation in larval dragonflies. Evol. Ecol. Res. 2004, 6, 619–626. [Google Scholar]
- Mikolajewski, D.J.; Johansson, F. Morphological and behavioral defenses in dragonfly larvae: Trait compensation and cospecialization. Behav. Ecol. 2004, 15, 614–620. [Google Scholar] [CrossRef]
- Kishida, O.; Trussell, G.C.; Mougi, A.; Nishimura, K. Evolutionary ecology of inducible morphological plasticity in predator–prey interaction: Toward the practical links with population ecology. Popul. Ecol. 2010, 52, 37–46. [Google Scholar] [CrossRef] [Green Version]
- Dmitriew, C.; Rowe, L. Resource limitation, predation risk and compensatory growth in a damselfly. Oecologia 2005, 142, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Angert, A.L.; Crozier, L.G.; Rissler, L.J.; Gilman, S.E.; Tewksbury, J.J.; Chunco, A.J. Do species’ traits predict recent shifts at expanding range edges? Ecol. Lett. 2011, 14, 677–689. [Google Scholar] [CrossRef] [PubMed]
- Hickling, R.; Roy, D.B.; Hill, J.K.; Thomas, C.D. A northward shift of range margins in British Odonata. Glob. Chang. Biol. 2005, 11, 502–506. [Google Scholar] [CrossRef]
- Wildermuth, H.; Gonseth, Y.; Maibach, A. Die Libellen der Schweiz. Fauna Helv. 2005, 12, 398. [Google Scholar]
- Khelifa, R.; Zebsa, R.; Kahalerras, A.; Mahdjoub, H. Clutch size and egg production in Orthetrum nitidinerve Selys, 1841 (Anisoptera: Libellulidae): Effect of body size and age. Int. J. Odonatol. 2012, 15, 51–58. [Google Scholar] [CrossRef]
- Khelifa, R. Spatiotemporal Pattern of Phenology across Geographic Gradients in Insects. Ph.D. Thesis, University of Zurich, Zurich, Switzerland, 2017. [Google Scholar]
- Thompson, D.J. Towards a realistic predator-prey model: The effect of temperature on the functional response and life history of larvae of the damselfly, Ischnura elegans. J. Anim. Ecol. 1978, 47, 757–767. [Google Scholar] [CrossRef]
- Corbet, P.S. Dragonflies: Behaviour and Ecology of Odonata; Harley Books: Colchester, UK, 1999. [Google Scholar]
- R Development Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Lenth, R. Least-Squares Means: The R package (lsmeans). J Stat. Softw. 2016, 69, 1–33. [Google Scholar] [CrossRef] [Green Version]
- Angilletta, M.J. Thermal Adaptation: A Theoretical and Empirical Synthesis; Oxford University Press: Oxford, UK, 2009. [Google Scholar]
- Suhling, F.; Suhling, I.; Richter, O. Temperature response of growth of larval dragonflies—An overview. Int. J. Odonatol. 2015, 18, 15–30. [Google Scholar] [CrossRef]
- Van Doorslaer, W.; Stoks, R. Growth rate plasticity to temperature in two damselfly species differing in latitude: Contributions of behaviour and physiology. Oikos 2005, 111, 599–605. [Google Scholar] [CrossRef]
- Karl, I.; Fischer, K. Why get big in the cold? Towards a solution to a life-history puzzle. Oecologia 2008, 155, 215–225. [Google Scholar] [CrossRef]
- De Block, M.; Stoks, R. Fitness effects from egg to reproduction: Bridging the life history transition. Ecology 2005, 86, 185–197. [Google Scholar] [CrossRef]
- Hunt, R.J.; Swift, M. Predation by larval damselflies on cladocerans. J. Freshw. Ecol. 2010, 25, 345–351. [Google Scholar] [CrossRef]
- Raczyński, M.; Stoks, R.; Johansson, F.; Sniegula, S. Size-mediated priority effects are trait-dependent and consistent across latitudes in a damselfly. Oikos 2021, 130, 1535–1547. [Google Scholar] [CrossRef]
- Traill, L.W.; Lim, M.L.; Sodhi, N.S.; Bradshaw, C.J. Mechanisms driving change: Altered species interactions and ecosystem function through global warming. J. Anim. Ecol. 2010, 79, 937–947. [Google Scholar] [CrossRef]
- Yang, L.H.; Rudolf, V. Phenology, ontogeny and the effects of climate change on the timing of species interactions. Ecol. Lett. 2010, 13, 1–10. [Google Scholar] [CrossRef]
- Altwegg, R. Predator-induced life-history plasticity under time constraints in pool frogs. Ecology 2002, 83, 2542–2551. [Google Scholar] [CrossRef]
- Slos, S.; Stoks, R. Behavioural correlations may cause partial support for the risk allocation hypothesis in damselfly larvae. Ethology 2006, 112, 143–151. [Google Scholar] [CrossRef]
- Wohlfahrt, B.; Mikolajewski, D.J.; Joop, G.; Vamosi, S.M. Ontogenetic changes in the association between antipredator responses and growth variables. Ecol. Entomol. 2007, 32, 567–574. [Google Scholar] [CrossRef]
- Mikolajewski, D.J.; Brodin, T.; Johansson, F.; Joop, G. Phenotypic plasticity in gender specific life-history: Effects of food availability and predation. Oikos 2005, 110, 91–100. [Google Scholar] [CrossRef]
- Johansson, F.; Andersson, J. Scared fish get lazy, and lazy fish get fat. J. Anim. Ecol. 2009, 78, 772–777. [Google Scholar] [CrossRef] [PubMed]
- Benard, M.F. Predator-induced phenotypic plasticity in organisms with complex life histories. Annu. Rev. Ecol. Evol. Syst. 2004, 35, 651–673. [Google Scholar] [CrossRef]
- Abrams, P.A.; Rowe, L. The effects of predation on the age and size of maturity of prey. Evolution 1996, 50, 1052–1061. [Google Scholar] [CrossRef] [PubMed]
- Higginson, A.; Ruxton, G. Adaptive changes in size and age at metamorphosis can qualitatively vary with predator type and available defenses. Ecology 2010, 91, 2756–2768. [Google Scholar] [CrossRef] [PubMed]
- Corbet, P.S.; Suhling, F.; Soendgerath, D. Voltinism of Odonata: A review. Int. J. Odonatol. 2006, 9, 1–44. [Google Scholar] [CrossRef]
- Johansson, F.; Stoks, R.; Rowe, L.; De Block, M. Life history plasticity in a damselfly: Effects of combined time and biotic constraints. Ecology 2001, 82, 1857–1869. [Google Scholar] [CrossRef]
- Pierce, C. Predator avoidance, microhabitat shift, and risk-sensitive foraging in larval dragonflies. Oecologia 1988, 77, 81–90. [Google Scholar] [CrossRef]
- Sih, A.; Bell, A.; Johnson, J.C. Behavioral syndromes: An ecological and evolutionary overview. Trends Ecol. Evol. 2004, 19, 372–378. [Google Scholar] [CrossRef] [Green Version]
- Dmitriew, C.M. The evolution of growth trajectories: What limits growth rate? Biol. Rev. 2011, 86, 97–116. [Google Scholar] [CrossRef]
- Metcalfe, N.B.; Monaghan, P. Growth versus lifespan: Perspectives from evolutionary ecology. Exp. Gerontol. 2003, 38, 935–940. [Google Scholar] [CrossRef]
- Johansson, F. Reaction norms and production costs of predator-induced morphological defences in a larval dragonfly (Leucorrhinia dubia: Odonata). Can. J. Zool. 2002, 80, 944–950. [Google Scholar] [CrossRef]
- Van Buskirk, J. Phenotypic lability and the evolution of predator-induced plasticity in tadpoles. Evolution 2002, 56, 361–370. [Google Scholar] [CrossRef]
- Belden, L.; Wildy, E.; Hatch, A.; Blaustein, A. Juvenile western toads, Bufo boreas, avoid chemical cues of snakes fed juvenile, but not larval, conspecifics. Anim. Behav. 2000, 59, 871–875. [Google Scholar] [CrossRef] [Green Version]
- Kiesecker, J.M.; Chivers, D.P.; Anderson, M.; Blaustein, A.R. Effect of predator diet on life history shifts of red-legged frogs, Rana aurora. J. Chem. Ecol. 2002, 28, 1007–1015. [Google Scholar] [CrossRef]
- Li, D.; Jackson, R.R. Influence of diet-related chemical cues from predators on the hatching of egg-carrying spiders. J. Chem. Ecol. 2005, 31, 333–342. [Google Scholar] [CrossRef]
- Ferland-Raymond, B.; Murray, D.L. Predator diet and prey adaptive responses: Can tadpoles distinguish between predators feeding on congeneric vs. conspecific prey? Can. J. Zool. 2008, 86, 1329–1336. [Google Scholar] [CrossRef]
- Ott, J. The expansion of Crocothemis erythraea (Brullé, 1832) in Germany-an indicator of climatic changes. In Odonata: Biology of Dragonflies; Scientific Publishers: Jodhpur, India, 2007; pp. 201–222. [Google Scholar]
- Korkeamäki, E.; Suhonen, J. Distribution and habitat specialization of species affect local extinction in dragonfly Odonata populations. Ecography 2002, 25, 459–465. [Google Scholar] [CrossRef]
- Trussell, G.C.; Ewanchuk, P.J.; Matassa, C.M. The fear of being eaten reduces energy transfer in a simple food chain. Ecology 2006, 87, 2979–2984. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zebsa, R.; Mahdjoub, H.; Khelifa, R. Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators. Diversity 2022, 14, 302. https://doi.org/10.3390/d14040302
Zebsa R, Mahdjoub H, Khelifa R. Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators. Diversity. 2022; 14(4):302. https://doi.org/10.3390/d14040302
Chicago/Turabian StyleZebsa, Rabah, Hayat Mahdjoub, and Rassim Khelifa. 2022. "Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators" Diversity 14, no. 4: 302. https://doi.org/10.3390/d14040302
APA StyleZebsa, R., Mahdjoub, H., & Khelifa, R. (2022). Similar Response of a Range Expanding Dragonfly to Low- and High-Elevation Predators. Diversity, 14(4), 302. https://doi.org/10.3390/d14040302