1. Introduction
Assessing species diversity has been one of the earliest and still major goals in natural history research, conservation science, and biogeography [
1,
2]. However, detection errors and imperfect detection may compromise the results of species distribution studies and limit effective conservation management [
3,
4]. Among the major issues in species diversity assessment is data quality: How complete are the data, and how big is the error to be considered in terms of imperfect detection [
3], phantom species, and “hidden diversity” [
5,
6]? The advantages of objective (nonpreferential) sampling of the presence and absence of species, of improved species detection, and of cross-validation procedures have been emphasized in numerous studies (e.g., [
7,
8,
9,
10,
11]), but even in standardized monitoring programs imperfect detection of plants is common [
3,
12].
Most large-scale plant inventories such as of islands are based on unsystematically collated data of heterogeneous spatial precision and detection capability by various observers. Between-observer variation and sampling bias are the rule rather than the exception [
3,
13]. Plant species detectability depends on a combination of factors, including species morphology, ecology and rarity, spatial (habitat density and habitat suitability) and temporal variation (season, phenology, and disturbance), observer skills, detection method, and timing and effort of the survey [
3,
4,
14,
15]. Some of these factors and especially observer experience are hard to quantify, and standardization may not always be effective when recording rare species [
13,
16]. Skilled botanists look for species of particular interest in sites where they sense a priori that they may occur.
With its 480 km², Limnos is the 8th largest island in the Aegean Sea and the largest of the North Aegean islands. Somewhat surprisingly, it ranked merely 20th in terms of species numbers among the Aegean islands [
17], and only the most recent survey [
18] prompted an upgrade to island number 12 in species richness. In terms of floristic knowledge, Strid [
17] classified Limnos among “the more poorly explored” islands “with perhaps 20–25% [species] to be added”. The rather uniform geology and in contrast to other Aegean islands the low relief (the highest summits of the island reach only just above 400 m) in addition to the scarceness of forests and cliff habitats are among the most plausible factors for the relatively low overall species richness of Limnos.
The botanical exploration of Limnos started with the renowned Austrian botanist K.H. Rechinger [
19,
20]. Since then, various workers have contributed to the knowledge of the island’s flora, with different aims and backgrounds, e.g., [
21] (vegetation survey), [
22] (trees and shrubs), [
23] (orchids), [
24] (entire flora), [
25] (crops and crop wild relatives), and [
26] (summer and ruderal flora), as well as several minor contributions published by [
27,
28,
29,
30,
31,
32,
33] and others, summing up the species number to 817 taxa [
17]. Our own records between 2016 and 2021 provided further 231 species, an addition of 28% [
18]. Such erratic history of research over a whole century is not untypical in regional floristics in Europe, but it renders it difficult to assess the research intensity and the degree of floristic coverage.
In view of the miscellaneous approaches by former collectors, our intention at the beginning was not inventorying the plant species of Limnos, and our approach was as preferential as the sampling before. But the number of newly found species was surprisingly high, which is why we wanted to know if patterns can be revealed in the set of newly found species.
Specifically, our aims are to explore (a) whether there are differences in traits between the previously known taxa and the newly detected ones, (b) which species are prone to disregard, and (c) which characteristics make plant species particularly neglectable in a Mediterranean island setting. We further related our findings to the question of the extent to which skills and observer experience and sampling effort contribute to the new detections.
2. Materials and Methods
The present study is based on historical and recent floristic surveys for the island of Limnos—historical data included in the previous floristic milestones by [
17,
24] and new data as described in the floristic update by [
18]. Most historical data reported as new in the inventory by [
24] were collected between 1989 and 1997 during 24 days of field work (2 days in March, 3 in April, 12 in May, 3 in June, and 4 days in November) by four researchers. The new data were collected by the authors in 2016 and 2018–2021 in spring (21 days in April and 40 days in May) and autumn (4 days at the end of September and 18 days in October). The authors formed two independent groups, somewhat in a complementary way, one focused on the vegetation of the predominant agropastoral landscapes (Bergmeier, Meyer, and Panitsa) and the other searched for plants as a leisure activity in special places and habitats (Krause and Ristow). A stratified sampling strategy (e.g., [
11]) covering the entire island was not attempted and was beyond our scope.
We created a floristic catalogue including the names of all plant taxa registered for the island of Limnos, as well as life forms, size (height), flower color, reproduction period (flowering and/or fruiting), perceptibility, rarity (abundance on the island of Limnos), status (native on Limnos or non-native, the latter mostly neophytes), and broad habitat preferences. Of the 231 newly reported taxa, 14 were considered nonestablished (compared to 24 such taxa in the former inventories). They were not included in the analyses, since we believe that casuals follow different patterns than established species. New casuals comprise less than 2% of the flora, but more than 1/3 of the total nonestablished flora. Established non-native taxa were included in the database to be analyzed. Twenty-one taxa (c. 2%) were not mentioned but noted in another name or taxon concept. Mere taxonomic specifications of aggregates or subspecies were disregarded. Taxonomy, life form, chorology, status categories, and habitat preferences followed Dimopoulos et al. [
34,
35,
36].
We used the following main
Life forms for the floristic analysis: phanerophytes (
P), chamaephytes (
Ch), hemicryptophytes (
H), geophytes (
G), therophytes (
T), and hydrophytes (
Aq). The
Size (height) of the plant taxa was classified as
vs.: very small < 0.05 m,
s: small 0.5–0.15 m,
m: medium 0.15–0.5 m,
l: large 0.5–1.8 m,
vl: very large > 1.8 m.
Flower colors were grouped as blue, greenish-unobtrusive (incl. brown-black-grey), pink-purple (incl. violet), red, white (incl. cream), and yellow (incl. orange). The
Reproduction (flowering) period was recognized as:
w-winter to early spring,
s-mid-late spring and
a-summer and autumn.
Perceptibility highlighted several plant groups, which are commonly neglected or undercollected in the field, such as “thistles” (prickly plants not easy to handle), graminoids (species with grass-like appearance, mainly Cyperaceae, Juncaceae, and Poaceae), yellow-flowering Asteraceae (this and the previous group with often concealed characters), hydrophytes (highly variable and sometimes difficult to access), and shrub/tree (incl. woody lianas) (commonly encountered only vegetatively).
Rarity–the abundance on the island of Limnos was classified, based on dot density [
17] and chiefly on own collection and observation data, as:
5–Abundant, taxa common in numerous sites of the island,
4–Scattered, taxa widely scattered in many sites or sometimes abundant,
3–Sparse, taxa constantly sparse or locally gregarious in several sites,
2–Rare, taxa sparse or locally gregarious but restricted to a few sites and
1–Very rare, taxa rare in only 1–2 sites of the island.
Habitat preferences were grouped as:
A-Freshwater habitats (including seasonally inundated),
C-Cliffs, rocks, walls, ravines, boulders,
G-Temperate and submediterranean grasslands,
M-Coastal habitats,
P-Xeric Mediterranean phrygana and grasslands,
R-Agricultural and ruderal habitats,
W-Woodlands and scrub [
35].
To test the similarities and differences between the two data subsets—the previously already known plant taxa and the newly recorded ones—we calculated the Sørensen dissimilarity index (β
SOR) formulated as:
where
Si is the total number of taxa in site
i,
ST is the total number of taxa in all sites considered together, and
bij,
bji are the number of taxa exclusive to sites
i and
j, respectively, when compared in pairs [
37].
3. Results
The update of the phytodiversity of Limnos revealed a high number of newly detected taxa not previously known to occur on the island [
18]. While most of the newly recorded taxa were found in spring (73.5% in April and May), a considerable proportion was collected during autumn (26.5% in September and October) (
Figure 1). About 10% of the newly recorded plant species are established neophytes (as opposed to 6% neophytes among the previously known species) of which 50% were detected during autumn fieldwork.
More than half of all taxa (53%) belong to seven plant families: Poaceae, Asteraceae, Fabaceae, Caryophyllaceae, Brassicaceae, Apiaceae, and Lamiaceae. These families are represented by 67.5% among the newly detected taxa and by 50.5% among the formerly known ones. Seven plant families (Rutaceae, Sapindaceae, Cymodoceaceae, Haloragaceae, Molluginaceae, Posidoniaceae, and Zosteraceae), each with one species, were represented only among the new records but not in the previously known flora.
The Convolvulaceae, Chenopodiaceae, Malvaceae, Poaceae, Boraginaceae, Polygonaceae, Ranunculaceae, and Rubiaceae occurred with higher proportions among the newly detected taxa than among the previously known flora, in contrast to other species-rich plant families such as Juncaceae, Orchidaceae, Euphorbiaceae, Geraniaceae, Brassicaceae, Fabaceae, Veronicaceae, Caryophyllaceae, Lamiaceae, Asteraceae, and Cyperaceae, which were underrepresented among the newly detected species (
Figure 2).
The newly detected taxa belong to 164 genera of which 56 were not formerly known for the flora of Limnos. The newly found genera include 13 genera of Poaceae, 7 of Asteraceae, 7 of Apiaceae, 3 of Chenopodiaceae, 2 genera each of Polygonaceae, Malvaceae, Brassicaceae, Caryophyllaceae, and Ranunculaceae; and 13 genera of 13 other families.
Therophytes predominate in the flora of Limnos with 55.8% (of which 12.5% were newly detected) followed by hemicryptophytes 20.5% (4.1% newly detected). Although newly found hydrophytes were few, they added almost 46% to this life form proportion (
Figure 3).
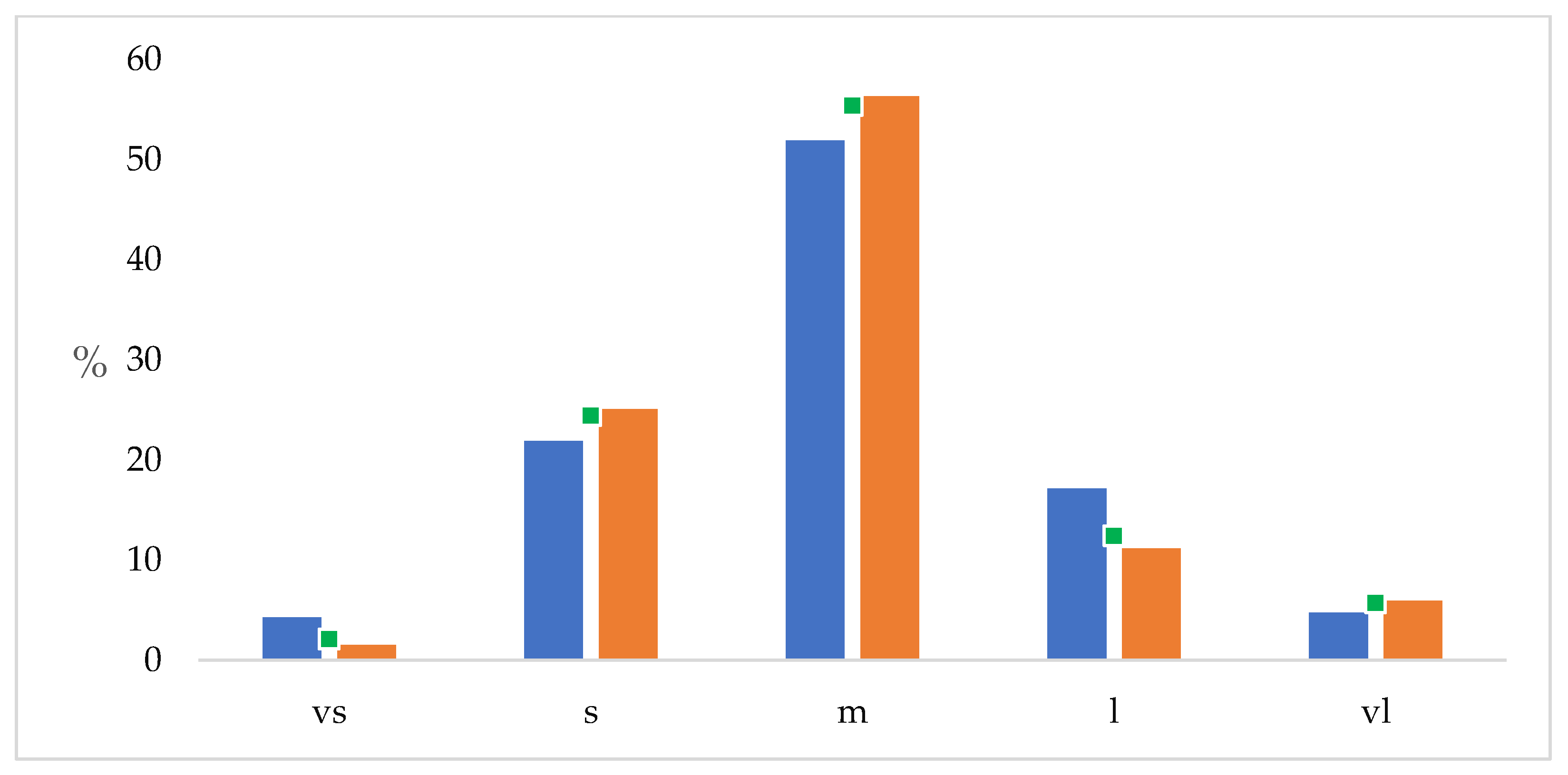
Newly detected taxa comprised mainly medium-sized plants (52,5%), followed by small (22.5%), large (16.7%), very large (4.9%), and very small taxa (3.4%) (
Figure 4). However, the newly detected taxa of large-sized plants were found to represent 27.9% of all the large-sized ones, and the newly detected taxa of very small-sized plants represent 36.8% of all very small-sized ones, both have distinctly higher proportions than among the earlier known taxa.
Concerning the trends of the proportions of plant size in different life form categories, newly detected taxa include small and medium-sized hydrophytes and higher percentages of very large geophytes and hemicryptophytes and also higher percentages of very small and large therophytes than former ones (
Supplementary Figure S1).
Plants with greenish-unobtrusive flowers dominate in the flora (34%), followed by pink-purple-(24.8%) and yellow-flowered plants (21.7%). Plant taxa with greenish-unobtrusive flowers presented the higher proportion among the newly detected species (42.8%) followed by those with yellow flowers (20.9%), (
Figure 5).
Graminoids and hydrophytes were represented by higher proportions among the newly detected taxa than among the previously known taxa. Yellow-flowering Asteraceae had rather similar proportions (
Figure 6a). Newly detected graminoids represented 29.8% of all graminoids, newly detected water plants 60% of all water plants, newly detected yellow-flowering Asteraceae 24.4% of all yellow-flowering Asteraceae, and newly detected thistles 16.6% of all thistles registered.
Most plants of Limnos (86,4%) reproduce during spring (s) or spring and early summer (s-a), and the pattern was similar for newly detected taxa (82.4%) and for previously known taxa (87.4%) (
Figure 6b). Plants reproducing during summer and autumn represent 15% of the newly detected taxa, a much higher proportion than for all registered taxa (10.1%) and for the previously known ones (8.8%).
Rarity (abundance) estimates of the plant taxa of Limnos showed that more than 78% of the newly detected taxa were classified as very rare or rare on the island, and there were no abundant taxa among them (
Figure 7). While the proportions of previously known taxa (and of the entire flora) form a normal curve, the newly detected taxa follow a degressive order with highest proportions for very rare and rare species. Newly detected hydrophytes, sparse hemicryptophytes, scattered geophytes, and very rare, rare, sparse, and scattered therophytes were represented among the newly detected taxa with proportions higher than those among the whole flora (
Supplementary Figure S2).
Plants of freshwater and coastal habitats were found with higher proportions among the newly detected species than among the previously known species (
Figure 8). Plants of agricultural and ruderal habitats as well as those of grasslands had generally high and similar proportions both among the newly detected and the previously known taxa.
Wild plants of arable fields which were the focus of our fieldwork represent 10.7% of the newly detected taxa and belong to 13 families and 21 genera. No less than 63.5% of them are rare or very rare on the island. Most of them are spring flowering and therophytes (86.4%).
The value of the Sørensen dissimilarity index (β
SOR) when comparing newly detected and formerly known taxa of Limnos is 0.434. The values of β
SOR for families and genera are rather low (0.12 and 0.18, respectively), while values of β
SOR for very rare taxa (0.53), very large-sized taxa (0.58), large-sized taxa (0.54), hydrophytes (0.46), therophytes (0.46), and hemicryptophytes (0.47) were found to be high (
Supplementary Figure S3).
4. Discussion
Our results show that there are differences in traits between the species previously known on the island and the newly detected ones. Some species groups appear to be more prone to observation and others to oversight. Overrepresented among the previously overlooked species are certain plant families, very small-sized but also large-sized plants, species with unobtrusive flowers, such as grasses, seagrasses, and other hydrophytes, autumnal species, plants of coastal and freshwater habitats, to a lesser extent also ruderal and arable habitats, and in general (very) rare species. The relatively high proportion of plants with inconspicuous flowers, very small size, and human-made (ruderal and arable) habitats among the newly detected species confirms [
38] who detected an esthetic bias among researchers. However, our results suggest not only a lack of esthetic plant traits (such as flower color or size) to be more common among newly detected taxa but also plants of unattractive (to many observers) or allegedly trivial habitats. Moreover, apart from an esthetic (plant-related) and ecological (habitat-related) bias, a perceptibility bias is detectable in that even among large plants a considerable proportion had been neglected. Perceptibility, in turn, appears to be a syndrome of various factors including reproduction seasonality, plant morphology, and specimen handiness. Such shortcomings in floristic investigations are sometimes generalized as observer bias but, as they may be of significance in biogeography, ecology, macroecology, and conservation studies and interpretation, specifications are needed. Species traits and rarity, habitat characteristics, phenology, spatial and temporal variation together with survey characteristics as the timing of survey are of high importance for species detection [
12,
14] and for the conservation of plant and phylogenetic diversity (among others, [
15,
38]). It is essential that such morphological, ecological and phenological information is provided in as much detail as possible in taxonomic identification work and flora (currently in common use in our study are chiefly the regional flora of Davis et al. [
39], Strid [
17], and Kleinsteuber et al. [
40]). Among the influencing background factors of esthetic, ecological, and perceptibility biases are taxonomic skills, floristic mapping experience [
41], sampling effort, an evolutionary adaptation of human cognition, cultural and learned preferences of researchers, and practical constraints in the field [
38].
4.1. Taxonomic Skills and Experience
The newly found taxa cover a wide range of 163 genera and 60 plant families including bindweed (Convolvulaceae), mallow (Malvaceae), chenopod (Chenopodiaceae), grass (Poaceae), and knotweed (Polygonaceae) families. The chenopod and grass families are notorious for being difficult to recognize or identify (small character traits not easy to see), which may result in a certain observer disinclination. Other plant families are less species-rich, and a few previously undercollected genera such as
Cuscuta, Heliotropium, Ranunculus sect.
Batrachium, and
Galium might change the overall picture of representation of these families. On the other hand, the families of Orchidaceae and Juncaceae, which are taxonomically and floristically difficult but are barely represented among the new records, show the relevance of taxonomic specialism in floristic surveys. Orchids and rushes were studied on the island by B. Biel and S. Snogerup, respectively [
23,
24], two well-known specialists of these groups. However, all in all, taxonomic skills and specialism, while all-important in plant inventorying, appear to be exceeded in relevance by rarity effects: most of the newly detected species were classified as rare or very rare, thus spotted either by chance or as a result of high sampling effort.
Experience in floristic surveying is the filter to reduce the effort of sampling. As a rule of thumb, the larger the study area, the more important experience appears to be–mainly due to the increasing number of sampling options. An experienced botanist filters the sampling options and prefers to sample representatively considering first of all habitat diversity and habitat rarity.
Different species groups and habitats may need different sampling strategies. Incomplete detection especially of (very) small plant species was to be expected, but in Poaceae, both (very) large and very small grasses were found disproportionately represented among the newly detected species suggesting that sampling experience may be more important than plant size to overcome the perceptibility bias, at least in some generally undercollected groups. In Chenopodiaceae, newly detected species were commonly of small size, and here it is about overcoming both the esthetic and the ecological bias.
Our figures show a certain overrepresentation among the newly found species of agricultural and ruderal habitats as well as of neophytes. Such seemingly trivial sites (and mostly “unesthetic” species) are commonly under recorded, especially in areas with a lot of “natural” vegetation. This coincides also with the overrepresentation of species of late (summer and autumn) reproduction. Neophytes such as Dysphania multifida and Phyla canescens may be the only real newcomers in the flora of the island, which may have been introduced and established in the last 20 or so years. But they contribute only very little to the newly found species.
Equally, coastal habitats require more thorough examination beyond a few representative sites, chiefly due to high local variation (beta diversity), especially on the island of Limnos that has the highest coastline–area ratio of all major Aegean islands. In contrast, permanent or seasonal freshwater habitats are rare and small-scale on all Aegean islands. The habitat is characterized by plants uncommon in the Mediterranean [
42,
43], fairly low within-site (alpha) but considerable turnover (beta) diversity (own data). Therefore, it requires spatial overrepresentative sampling in comparison to large-scale habitats.
Flower color is an obvious trait for recognition. It does not come as a surprise but confirms the esthetic bias [
38] that a major part of the newly found species has unobtrusive flowers, such as Poaceae.
With the itemization of plant groups with reduced “perceptibility”, we hypothesized that certain groups are more commonly neglected or undercollected than others. Our hypotheses were found to be confirmed in graminoids (yet only Poaceae, the other two major families Juncaceae and Cyperaceae were well sampled before) and water plants but not obviously in yellow Asteraceae, thistles, and shrubs/trees. The latter group of woody plants was extensively studied before [
22].
4.2. Effort of Sampling
Floristic inventories require a harmonic combination of taxonomic skills, knowledge of the studied area and the related floristic literature, well-developed systematic awareness, and an appropriate sampling design [
44]. It may seem self-explanatory that a higher sampling effort results in a higher amount of detected species diversity. The very high amount of sampling time and timing representativity are surely crucial for the new survey. Our results show that the autumn was insufficiently covered in the previous surveys. The need for intensive exploration of Mediterranean island flora during autumn and early spring and the importance of “all-season” floristic investigation was also emphasized by [
15]. Almost all newly found species with an abundance class higher than rare were found in the first half of the survey [
18], which highlights the importance of increased sampling effort. However, even later in the survey period, several new rare or very rare species were found (
Figure 1), including species of plant geographical importance (e.g., new to the North Aegean islands). We may conclude that a certain number of rare species still awaits detection, unveiling our own inventory deficits and blind spots.
From a methodological point of view, it depends on the research or monitoring objective and on the resources in time and money whether the additional diversity that can be expected is worth the extra effort. In our study, it appears to be worthwhile: as much as 20% of the newly found taxa were classified as not rare (but as scattered or scarce); the vast majority (78%) were classified as very rare or rare on the island, independent of phenology and sampling period. Generally, the rarer a species, the more important the sampling effort is. If the research objective is landscape-scale, where chiefly large-scale habitats are to be characterized by dominant or gregarious plants, then the sampling effort may be much lower than in diversity-based research, e.g., in the field of nature conservation science, where also small-scale habitats and within-habitat variation is to be considered. In the latter case, yet with limited resources and time budgets, a compromise in sampling effort must be made to which our findings and considerations regarding taxonomic skills and experience, species traits and perceptibility, habitat frequency, and variation may contribute. Although our factor analysis casts merely a single island’s flashlight on some species that are more likely overlooked than others, it has become clear that a combination of multiple factors applies in large-scale surveys controlling detection and nondetection of species. Detection probability in unstandardized sampling such as commonly on Mediterranean islands is largely a function of botanist skills, experience, and sampling effort. Nondetection due to esthetic, ecological, and perceptibility observer biases may result in uneven taxonomic, growth form, season, and habitat coverage of species inventories.
In-depth biodiversity studies in medium- to large-sized study areas, such as on species richness patterns, species-habitat relations, and conservation status assessments, can hardly be successful if not underpinned by sufficient research time and geographical, ecological, and taxonomic experience. Floristic research should be focused on certain (regionally specific) taxa or species groups, in order to systematically improve existing piecemeal collated data. Moreover, rare target species may not be detected or only randomly detected by unsystematic sampling. Concerning specific plant traits and fine-grained species–habitat relationships, further studies are wanted.



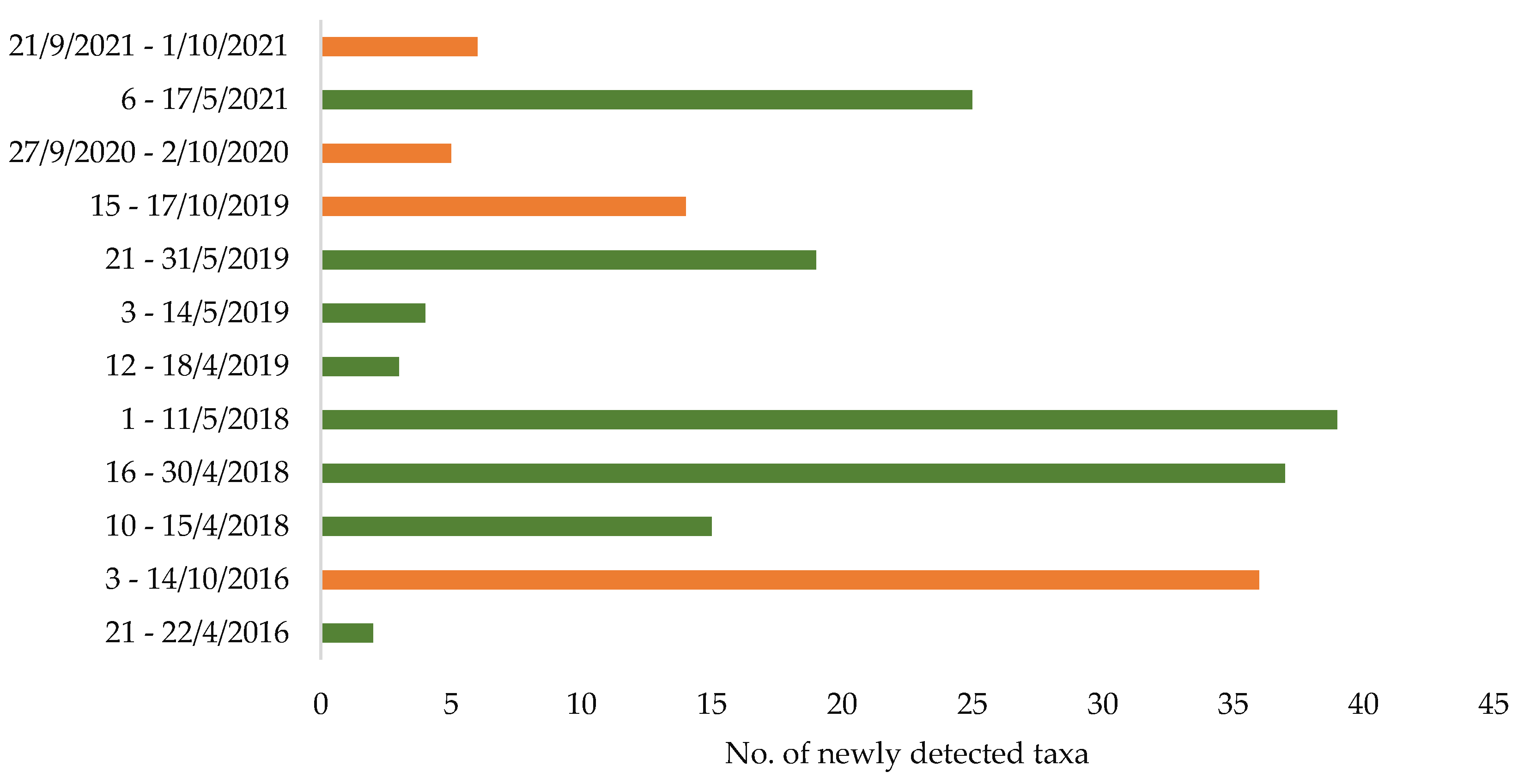{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
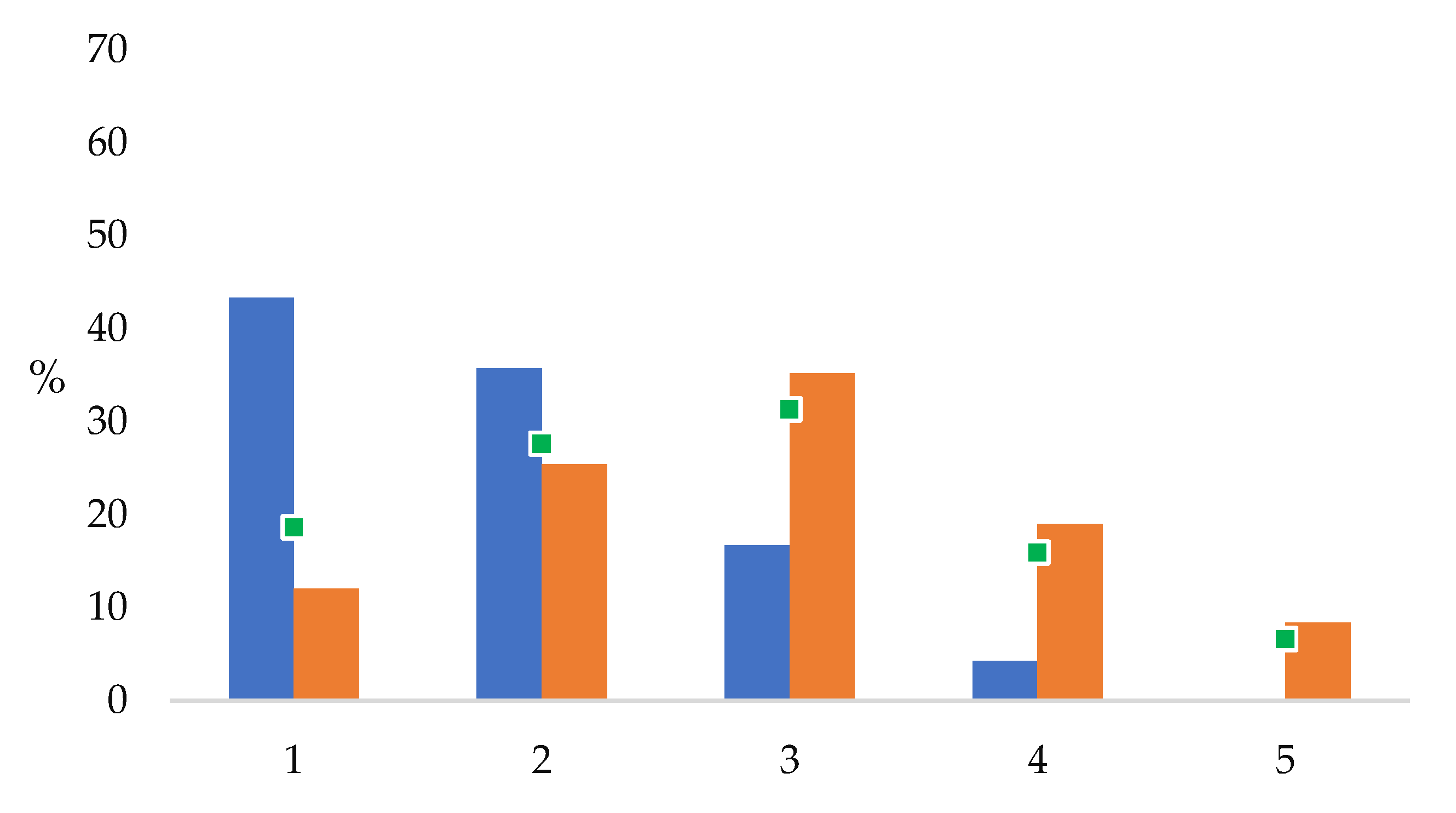{kind=link}
{kind=link}







