
Genetic Structure and Diversity of Native Tench (Tinca tinca L. 1758) Populations in Hungary—Establishment of Basic Knowledge Base for a Breeding Program


 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling and DNA Extraction, Microsatellite Analysis
2.2. mtDNA Cytb Sequence Amplification and Analysis
2.3. PCR RFLP Analysis
2.4. Statistical Analysis
3. Results
3.1. Microsatellite Analysis
3.1.1. Genetic Diversity and Population Size
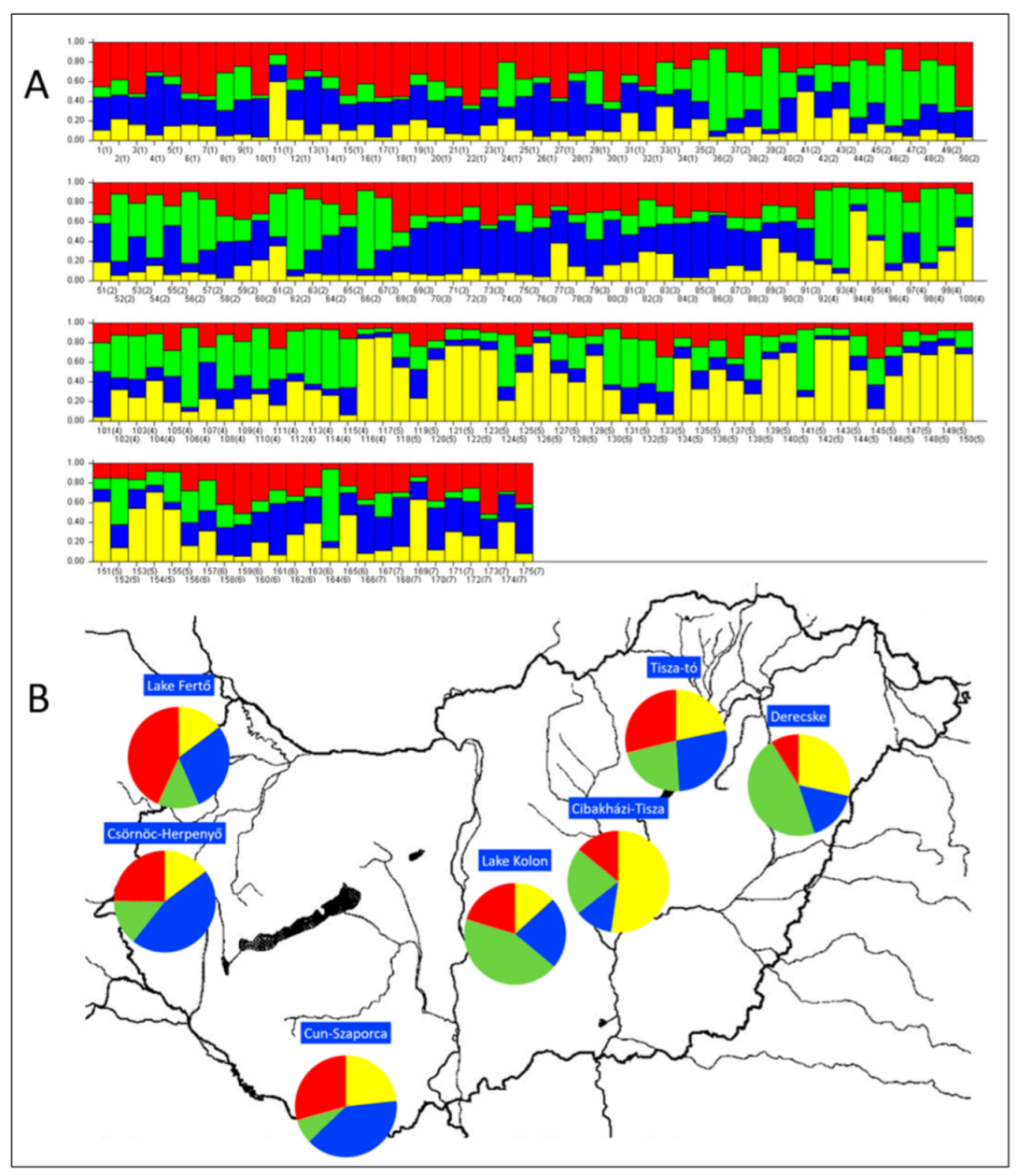
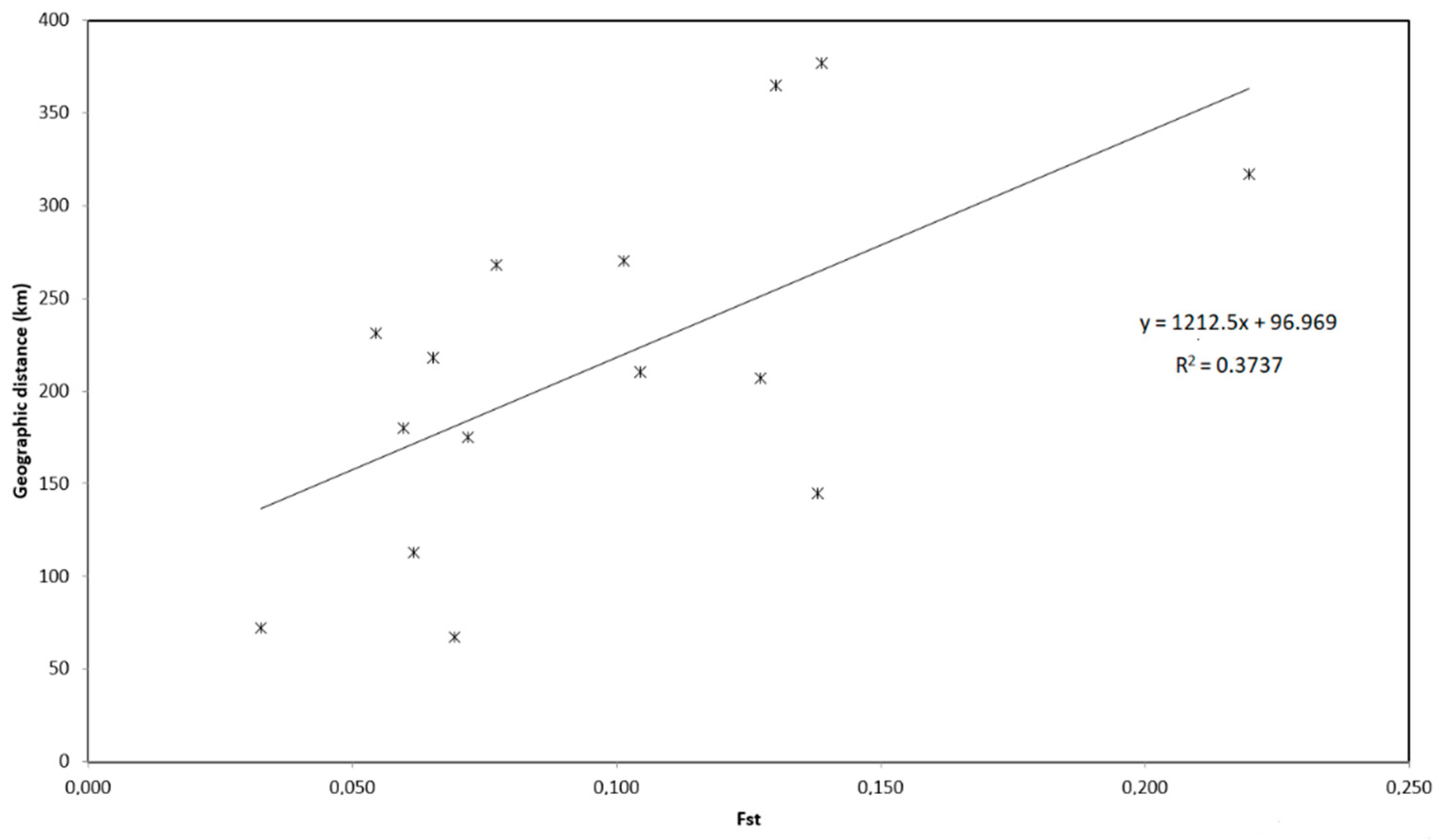
3.1.2. Genetic Structure
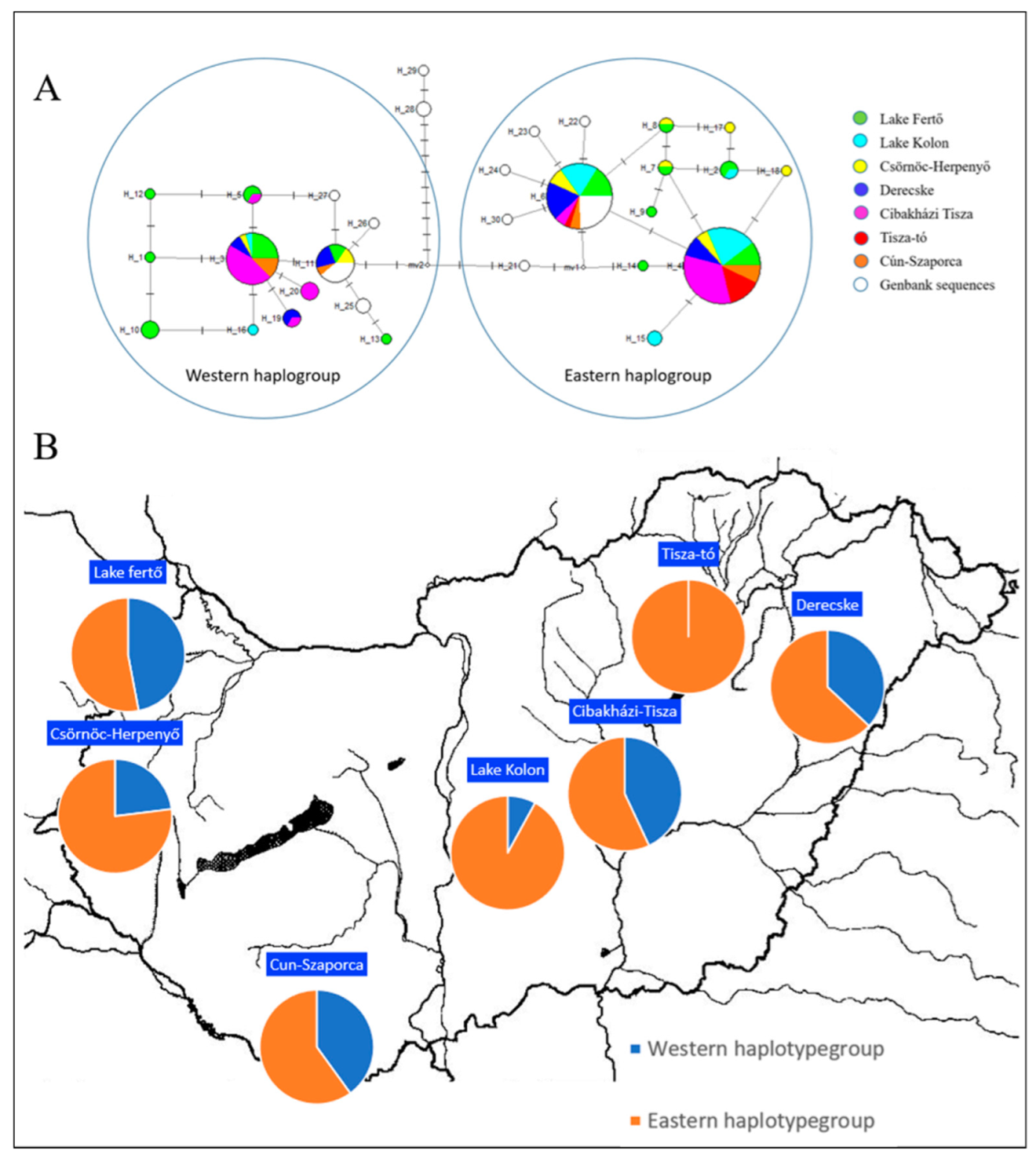
3.2. Mihtocondrial DNA Analysis
3.3. PCR-RFLP Analysis of Act and Rps7 Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Brylinska, M.; Brylinski, E.; Bninska, M. Tinca tinca (Linnaeus, 1758). In The Freshwater Fishes of Europe, 1st ed.; Banarescu, P., Ed.; Aula Verlag GmbH: Wiebelsheim, Germany, 1999; Volume 5, pp. 229–302. [Google Scholar]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Publications Kottelat: Cornol, Switzerland; Freyhof: Berlin, Germany, 2007; p. 646. [Google Scholar]
- Steffens, W. The Tench [Tinca tinca L.], a neglected pond fish species. Pol. Arch. Hydrobiol. 1995, 42, 161–180. [Google Scholar]
- Celada, J.D.; Aguilera, A.; Carral, J.M.; Sáez-Royuela, M.; Melendre, P.M.; Pérez, J.R. Effects of stocking density on survival and growth of juvenile Tench (Tinca Tinca L.). Aquac. Int. 2007, 15, 461–465. [Google Scholar] [CrossRef]
- Gela, D.; Flajšhans, M.; Kocour, M.; Rodina, M.; Linhart, O. Tench (Tinca tinca) broodstock management in breeding station under conditions of pond culture: A review. Aquac. Int. 2006, 14, 195–203. [Google Scholar] [CrossRef]
- Kujawa, R.; Kucharczyk, D.; Mamcarz, A.; Żarski, D.; Targońska, K. Artificial spawning of common Tench Tinca tinca (Linnaeus, 1758), obtained from wild and domestic stocks. Aquac. Int. 2011, 19, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Moruzi, I.; Pishchenko, Y.; Kropachev, D. Results of the first experiment on domestication of the Tench (Tinca tinca, L., 1758) in ponds of the novosibirsk region. In Proceedings of the Sustainable Use, Protection Of Animal World And Forest Management in the Context of Climate Change, Chișinău, Moldova, 12–13 October 2016; pp. 213–214. [Google Scholar]
- Wang, J.; Min, W.; Guan, M.; Gong, L.; Ren, J.; Huang, Z.; Zheng, H.; Zhang, J.; Liu, H.; Han, Y. Tench farming in China: Present status and future prospects. Aquac. Int. 2006, 14, 205–208. [Google Scholar] [CrossRef]
- Wolnicki, J.; Myszkowski, L.; Korwin-Kossakowski, M.; Kamiński, R.; Stanny, L.A. Effects of different diets on juvenile Tench, Tinca tinca (L.) reared under controlled conditions. Aquac. Int. 2006, 14, 89–98. [Google Scholar] [CrossRef]
- Wolnicki, J.; Kamiński, R.; Sikorska, J. Combined effects of water temperature and daily food availability period on the growth and survival of Tench (Tinca tinca) larvae. Aquacult. Res. 2017, 48, 3809–3816. [Google Scholar] [CrossRef]
- Billard, R.; Flajshans, M. The current state of research and culture of Tench, Tinca tinca Linnaeus [1758]. Concluding remarks on the workshop. Pol. Arch. Hydrobiol. 1995, 42, 219–225. [Google Scholar]
- Ćirković, M.; Ljubojević, D.; Đorđević, V.; Novakov, N.; Petronijević, R.; Matekalo-Sverak, V.; Trbović, D. The breed effect on productivity and meat nutrient compsition of fish. Kafkas Univ. Vet. Fak. Derg. 2012, 18, 75–780. [Google Scholar]
- Lo Presti, R.; Kohlmann, K.; Kersten, P.; Gasco, L.; Di Stasio, L. Tinca Gobba Dorata del Pianalto di Poirino: Genetic characterization by microsatellite markers. Ital. J. Anim. Sci. 2010, 9, 4. [Google Scholar] [CrossRef] [Green Version]
- Adámek, Z.I.; Sukop, P.; Rendón, M.; Kouřil, J. Food competition between 2+ Tench (Tinca tinca L.), Common Carp (Cyprinus carpio L.) and Bigmouth buffalo (Ictiobus cyprinellus Val.) in pond polyculture: Food competition between Tench, Carp and Bigmouth Buffalo. J. Appl. Ichthyol. 2003, 19, 165–169. [Google Scholar] [CrossRef]
- Grozev, G.K.; Hubenova-Siderova, T.; Paskaleva, E. Natural feeding of Tench (Tinca tinca L.) at polyculture rearing in carp ponds. Bulg. J. Agric. Sci. 2000, 6, 209–214. [Google Scholar]
- Dulski, T.; Kozłowski, K.; Ciesielski, S. Habitat and seasonality shape the structure of Tench (Tinca tinca L.) gut microbiome. Sci. Rep. 2020, 10, 4460. [Google Scholar] [CrossRef] [PubMed]
- Pantazis, P. Culture potential of the cyprinid Tinca tinca Linnaeus 1758 (Tench) in Greece. In Proceedings of the 4th International Symposium on Hydrobiology and Fisheries, Volos, Greece, 9–11 June 2011. [Google Scholar]
- Kamiński, R.; Sikorska, J.; Wolnicki, J. Diet and water temperature affect growth and body deformities in juvenile Tench Tinca tinca (L.) reared under controlled conditions. Aquacult. Res. 2016, 48, 1327–1337. [Google Scholar] [CrossRef]
- Wachirachaikarn, A.; Sutthakiet, O.; Senanan, W.; Na-Nakorn, U. Development of the new microsatellite multiplex PCR panel and genetic variation of farmed snakeskin gourami, Trichopodus pectoralis. Aquac. Int. 2020, 28, 751–765. [Google Scholar] [CrossRef]
- Eknath, A.E.; Tayamen, M.M.; Palada-de Vera, M.S.; Danting, J.C.; Reyes, R.A.; Dionisio, E.E.; Capili, J.B.; Bolivar, H.L.; Abella, T.A.; Circa, A.V.; et al. Genetic improvements of farmed tilapias: The growth performance of eight strains of Oreochromis niloticus tested in different farm environments. Aquaculture 1993, 111, 171–188. [Google Scholar] [CrossRef]
- Eknath, A.E.; Acosta, B.O. Genetic Improvement of Farmed Tilapias (GIFT) Project: Final Report; International Center for Living Aquatic Resources Management: Makaty City, Philippines, 1998. [Google Scholar]
- Askari, G.H.; Shabani, A.; Kolangi Miandare, H. Application of molecular markers in fisheries and aquaculture. Sci. J. Anim. Sci. 2013, 2, 82–88. [Google Scholar]
- Chauhan, T.; Kumar, R. Molecular markers and their applications in fisheries and aquaculture. Adv. Biosci. Biotechnol. 2010, 1, 281–291. [Google Scholar] [CrossRef] [Green Version]
- Lujić, J.; Kohlmann, K.; Kersten, P.; Marinović, Z.; Ćirković, M.; Simić, V. Phylogeographic identification of Tench Tinca tinca (L., 1758) (Actinopterygii: Cyprinidae) from the Northern Balkans and adjacent regions and its implications for conservation. Zool. Stud. 2017, 56, 1–9. [Google Scholar] [CrossRef]
- Mastrochirico Filho, V.A.; Freitas, M.V.; Ariede, R.B.; Lira, L.V.; Mendes, N.J.; Hashimoto, D.T. Genetic applications in the conservation of Neotropical freshwater fish. In Biological Resources of Water; IntechOpen: Rijeka, Croatia, 2018; pp. 249–284. [Google Scholar]
- Hernandez, C.; Bougas, B.; Perreault-Payette, A.; Simard, A.; Côté, G.; Bernatchez, L. 60 specific eDNA qPCR assays to detect invasive, threatened, and exploited freshwater vertebrates and invertebrates in Eastern Canada. Environ. DNA 2020, 2, 373–386. [Google Scholar] [CrossRef]
- Kohlmann, K.; Kersten, P.; Panicz, R.; Memiş, D.; Flajšhans, M. Genetic variability and differentiation of wild and cultured tench populations inferred from microsatellite loci. Rev. Fish Biol. Fish. 2010, 20, 279–288. [Google Scholar] [CrossRef]
- Kohlmann, K.; Kersten, P.; Flajšhans, M. Comparison of microsatellite variability in wild and cultured Tench (Tinca tinca). Aquaculture 2007, 272, S147–S151. [Google Scholar] [CrossRef]
- Lajbner, Z.; Kotlik, P. PCR-RFLP assays to distinguish the Western and Eastern phylogroups in wild and cultured Tench Tinca tinca. Mol. Ecol. Resour. 2011, 11, 374–377. [Google Scholar] [CrossRef] [PubMed]
- Lo Presti, R.; Kohlmann, K.; Kersten, P.; Gasco, L.; Lisa, C.; Di Stasio, L. Genetic variability in Tench (Tinca Tinca L.) as revealed by PCR-RFLP analysis of mitochondrial DNA. Ital. J. Anim. Sci. 2021, 11, e19. [Google Scholar] [CrossRef]
- Lo Presti, R.; Kohlmann, K.; Kersten, P.; Lisa, C.; Di Stasio, L. Sequence variability at the mitochondrial ND1, ND6, Cyt b and D-Loop segments in Tench (Tinca tinca L.). J. Appl. Ichthyol. 2014, 30, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Kumar, G.; Langa, J.; Montes, I.; Conklin, D.; Kocour, M.; Kohlmann, K.; Estonba, A. A novel transcriptome-derived SNPs array for Tench (Tinca tinca L.). PLoS ONE 2019, 14, e0213992. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic consequences of climatic oscillations in the quaternary. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [Green Version]
- Lajbner, Z.; Kohlmann, K.; Linhart, O.; Kotlík, P. Lack of reproductive isolation between the Western and Eastern phylogroups of the Tench. Rev. Fish Biol. Fish. 2010, 20, 289–300. [Google Scholar] [CrossRef]
- Karaiskou, N.; Gkagkavouzis, K.; Minoudi, S.; Botskaris, D.; Markou, K.; Kalafatakis, S.; Antonopoulou, E.; Triantafyllidis, A. Genetic structure and divergence of Tinca tinca European populations. J. Fish Biol. 2020, 97, 930–934. [Google Scholar] [CrossRef]
- Lajbner, Z.; Linhart, O.; Kotlík, P. Human-aided dispersal has altered but not erased the phylogeography of the Tench. Evol. Appl. 2011, 4, 545–561. [Google Scholar] [CrossRef]
- Kohlmann, K.; Kersten, P. Microsatellite loci in Tench: Isolation and variability in a test population. Aquac. Int. 2006, 14, 3–7. [Google Scholar] [CrossRef]
- Crooijmans, R.P.M.A.; Poel, J.V.D.; Groenen, M.A.M.; Bierbooms, V.A.F.; Komen, J. Microsatellite markers in Common Carp (Cyprinus carpio L.). Anim. Genet. 1997, 28, 129–134. [Google Scholar] [CrossRef]
- Baerwald, M.R.; May, B. Characterization of microsatellite loci for five members of the minnow family Cyprinidae found in the Sacramento-San Joaquin delta and its tributaries. Mol. Ecol. Notes 2004, 4, 385–390. [Google Scholar] [CrossRef]
- Machordom, A.; Doadrio, I. Evidence of a cenozoic betic–kabilian connection based on freshwater fish phylogeography (Luciobarbus, Cyprinidae). Mol. Phylogenet. Evol. 2001, 18, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Touriya, A.; Rami, M.; Cattaneo-Berrebi, G.; Ibanez, C.; Augros, S.; Boissin, E.; Dakkak, A.; Berrebi, P. Primers for EPIC amplification of intron sequences for fish and other vertebrate population genetic studies. Biotechniques 2003, 35, 676–682. [Google Scholar] [CrossRef]
- Chow, S.; Hazama, K. Universal PCR primers for S7 ribosomal protein gene introns in fish. Mol. Ecol. 1998, 7, 1255–1256. [Google Scholar]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-checker: Software for identifying and correcting gen-otyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Rousset, F. Genepop’007: A complete re-implementation of the genepop software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef] [PubMed]
- Guo, S.W.; Thompson, E.A. Performing the exact test of Hardy-Weinberg proportion for multiple alleles. Biometrics 1992, 48, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Weir, B.S. Genetic Data Analysis; Sinauer Associates, Inc.: Sunderland, MA, USA, 1996; pp. 166–167. [Google Scholar]
- Cavalli-Sforza, L.L.; Edwards, A.W. Phylogenetic analysis. Models and estimation procedures. Am. J. Hum. Genet. 1967, 19, 233–257. [Google Scholar]
- Chapuis, M.P.; Estoup, A. Microsatellite null alleles and estimation of population differentiation. Mol. Biol. Evol. 2007, 24, 621–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Do, C.; Waples, R.S.; Peel, D.; Macbeth, G.M.; Tillett, B.J.; Ovenden, J.R. NeEstimator v2: Re-implementation of software for the estimation of contemporary effective population size (Ne) from genetic data. Mol. Ecol. Resour. 2013, 14, 209–214. [Google Scholar] [CrossRef]
- Piry, S.; Luikart, G.; Cornuet, J.M. Computer note. BOTTLENECK: A computer program for detecting recent reductions in the effective size using allele frequency data. J. Hered. 1999, 90, 502–503. [Google Scholar] [CrossRef]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multil ocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Falush, D.; Stephens, M.; Pritchard, J.K. Inference of population structure using multilocus genotype data: Linked loci and correlated allele frequencies. Genetics 2003, 164, 1567–1587. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; VonHoldt, B.M. Structure Harvester: A website and program for visualizing STRUCTURE output and imple-menting the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software structure: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Zhang, Z.; Schwartz, S.; Wagner, L.; Miller, W. A greedy algorithm for aligning DNA sequences. J. Comput. Biol. 2000, 7, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Lajbner, Z.; Linhart, O.; Kotlik, P. Molecular phylogeography of the Tench Tinca tinca (Linnaeus,1758). In Proceedings of the 12th European Congress of Ichthyology, Cavtat (Dubrovnik), Croatia, 9–13 September 2007; p. 35. [Google Scholar]
- Molnár, T.; Benedek, I.; Kovács, B.; Zsolnai, A.; Lehoczky, I. Genetic consequences of pond production of a pikeperch (Sander lucioperca L.) stock with natural origin: The effects of changed selection pressure and reduced population size. PeerJ 2020, 8, e8745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponzoni, R.W.; Khaw, H.L.; Nguyen, N.H.; Hamzah, A. Inbreeding and effective population size in the Malaysian nucleus of the GIFT strain of Nile tilapia (Oreochromis niloticus). Aquaculture 2010, 302, 42–48. [Google Scholar] [CrossRef]
- Clavero, M. Beyond one bone: Interdisciplinarity to assess nativeness of the Tench (Tinca tinca) in Spain. Aquat. Conserv. 2019, 29, 1863–1869. [Google Scholar] [CrossRef]
- Morissette, O.; Lecomte, F.; Vachon, N.; Drouin, A.; Sirois, P. Quantifying migratory capacity and dispersal of the invasive Tench (Tinca tinca) in the St. Lawrence River using otolith chemistry. Can. J. Fish. Aquat. Sci. 2021, 78, 1628–1638. [Google Scholar] [CrossRef]
- Bernos, T.A.; Avlijaš, S.; Hill, J.; Morissette, O.; Ricciardi, A.; Mandrak, N.E.; Jeffries, K.M. Spatiotemporal changes in genetic diversity and structure of a recent fish invasion in eastern North America. bioRxiv 2022. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
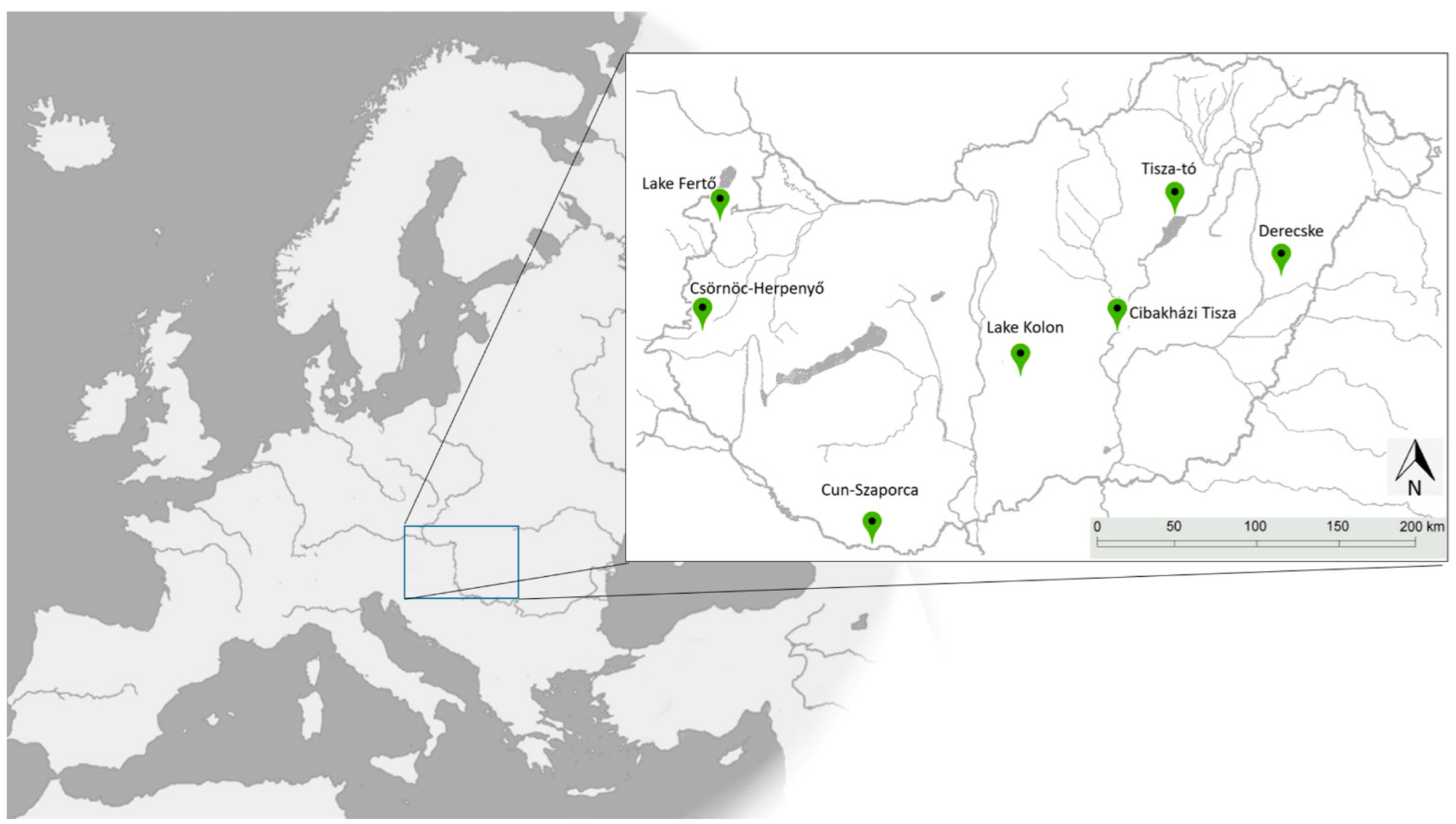
| Location | Label | GPS Coordinates | Region | Sample Size | |
|---|---|---|---|---|---|
| Derecske | d | N 47°20′20.48″ | E 21°34′25.13″ | eastern | 25 |
| Lake Tisza | t | N 47°39′31.10″ | E 20°41′54.25″ | eastern | 10 |
| Cibakházi Tisza | cht | N 46°59′03.53″ | E 20°10′21.77″ | eastern | 39 |
| Lake Kolon | k | N 46°45′35.05″ | E 19°20′26.80″ | central | 32 |
| Cun-Szaporca | csz | N 45°46′55.10″ | E 18°06′09.31″ | central | 10 |
| Csörnöc-Herpenyő | csh | N 46°59′37.83″ | E 16°36′55.11″ | western | 25 |
| Lake Fertő | f | N 47°38′08.30″ | E 16°44′39.28″ | western | 34 |
| Lake Fertő | Lake Kolon | Csörnöc-Herpenyő | Derecske | Cibakházi Tisza | Lake Tisza | Cun-Szaporca | |
|---|---|---|---|---|---|---|---|
| Na | 4.16 ± 2.24 | 3.83 ± 1.69 | 3.16 ± 1.40 | 2.41 ± 1.16 | 3.25 ± 1.42 | 2.91 ± 1.16 | 2.83 ± 1.19 |
| Neff | 2.22 ± 0.78 a | 1.81 ± 0.57 ab | 1.92 ± 0.61 ab | 1.38 ± 0.29 b | 1.80 ± 0.63 ab | 1.92 ± 0.50 ab | 1.92 ± 0.69 ab |
| Ho | 0.43 ± 0.20 b | 0.32 ± 0.16 ab | 0.44 ± 0.22 b | 0.17 ± 0.13 a | 0.37 ± 0.20 ab | 0.36 ± 0.18 ab | 0.47 ± 0.31 b |
| uHe | 0.49 ± 0.18 a | 0.39 ± 0.20 ab | 0.43 ± 0.19 ab | 0.25 ± 0.16 b | 0.40 ± 0.18 ab | 0.47 ± 0.17 ab | 0.43 ± 0.25 ab |
| Fis | 0.11 ± 0.19 ab | 0.15 ± 0.18 ab | −0.03 ± 0.15 a | 0.28 ± 0.31 b | 0.04 ± 0.19 ab | 0.16 ± 0.30 ab | −0.12 ± 0.32 a |
| AR | 3.27 ± 1.36 | 2.74 ± 1.02 | 2.59 ± 1.05 | 2.02 ± 0.69 | 2.50 ± 0.93 | 2.78 ± 1.03 | 2.72 ± 1.11 |
| ARp | 0.40 ± 0.42 a | 0.12 ± 0.22 ab | 0.16 ± 0.21 ab | 0.02 ± 0.06 b | 0.12 ± 0.24 ab | 0.12 ± 0.31 ab | 0.10 ± 0.29 ab |
| Fst/Genetic Distance | Lake Fertő | Lake Kolon | Csörnöc-Herpenyő | Derecske | Cibakházi Tisza | Tisza-tó | Cun-Szaporca |
|---|---|---|---|---|---|---|---|
| Lake Fertő | - | 0.228 | 0.209 | 0.299 | 0.256 | 0.243 | 0.286 |
| Lake Kolon | 0.065 | - | 0.246 | 0.232 | 0.237 | 0.195 | 0.291 |
| Csörnöc-Herpenyő | 0.032 | 0.104 | - | 0.255 | 0.236 | 0.245 | 0.250 |
| Derecske | 0.130 | 0.059 | 0.138 | - | 0.227 | 0.271 | 0.332 |
| Cibakházi Tisza | 0.077 | 0.069 | 0.101 | 0.061 | - | 0.232 | 0.266 |
| Tisza-tó | 0.008 | 0.029 | 0.043 | 0.120 | 0.042 | - | 0.258 |
| Cun-Szaporca | 0.054 | 0.138 | 0.071 | 0.219 | 0.127 | 0.051 | - |
| Stock | Nh | S | Eta | Hd (Mean ± SD) | Pi (Mean ± SD) |
|---|---|---|---|---|---|
| Lake Fertő | 14 | 11 | 12 | 0.909 ± 0.026 | 0.00728 ± 0.00035 |
| Lake Kolon | 5 | 9 | 9 | 0.556 ± 0.130 | 0.00290 ± 0.00123 |
| Csörnöc-Herpenyő | 9 | 9 | 9 | 0.813 ± 0.081 | 0.00456 ± 0.00111 |
| Derecske | 5 | 8 | 8 | 0.789 ± 0.057 | 0.00597 ± 0.00076 |
| Cibakházi Tisza | 6 | 10 | 10 | 0.655 ± 0.060 | 0.00552 ± 0.00037 |
| Tisza-tó | 2 | 1 | 1 | 0.222 ± 0.166 | 0.00036 ± 0.00027 |
| Cun-Szaporca | 4 | 7 | 7 | 0.778 ± 0.091 | 0.00596 ± 0.00094 |
| Cytb | ||
|---|---|---|
| Western (%) | Eastern (%) | |
| Derecske | 37 | 63 |
| Lake Tisza | 0 | 100 |
| Cibakházi Tisza | 43 | 57 |
| Lake Kolon | 8 | 92 |
| Cun-Szaporca | 40 | 60 |
| Csörnöc-Herpenyő | 23 | 77 |
| Lake Fertő | 47 | 53 |
| Average | 28 | 72 |
| Act (Eco52l) | Rps7 (Ndel) | |||||
|---|---|---|---|---|---|---|
| W (%) | E (%) | WE-Het (%) | W (%) | E (%) | WE-Het (%) | |
| Derecske | 8 | 36 | 56 | 4 | 56 | 40 |
| Lake Tisza | 10 | 0 | 90 | 60 | 30 | 10 |
| Cibakházi Tisza | 50 | 16 | 34 | 37 | 3 | 60 |
| Lake Kolon | 16 | 31 | 53 | 12 | 47 | 41 |
| Cun-Szaporca | 60 | 0 | 40 | 60 | 0 | 40 |
| Csörnöc-Herpenyő | 54 | 13 | 33 | 12 | 32 | 56 |
| Lake Fertő | 32 | 24 | 44 | 44 | 15 | 41 |
| avarage | 33 | 17 | 50 | 33 | 26 | 41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Al Fatle, F.A.; Meleg, E.E.; Sallai, Z.; Szabó, G.; Várkonyi, E.; Urbányi, B.; Kovács, B.; Molnár, T.; Lehoczky, I. Genetic Structure and Diversity of Native Tench (Tinca tinca L. 1758) Populations in Hungary—Establishment of Basic Knowledge Base for a Breeding Program. Diversity 2022, 14, 336. https://doi.org/10.3390/d14050336
Al Fatle FA, Meleg EE, Sallai Z, Szabó G, Várkonyi E, Urbányi B, Kovács B, Molnár T, Lehoczky I. Genetic Structure and Diversity of Native Tench (Tinca tinca L. 1758) Populations in Hungary—Establishment of Basic Knowledge Base for a Breeding Program. Diversity. 2022; 14(5):336. https://doi.org/10.3390/d14050336
Chicago/Turabian StyleAl Fatle, Fatema Ali, Erika Edviné Meleg, Zoltán Sallai, Gergely Szabó, Eszter Várkonyi, Béla Urbányi, Balázs Kovács, Tamás Molnár, and István Lehoczky. 2022. "Genetic Structure and Diversity of Native Tench (Tinca tinca L. 1758) Populations in Hungary—Establishment of Basic Knowledge Base for a Breeding Program" Diversity 14, no. 5: 336. https://doi.org/10.3390/d14050336
APA StyleAl Fatle, F. A., Meleg, E. E., Sallai, Z., Szabó, G., Várkonyi, E., Urbányi, B., Kovács, B., Molnár, T., & Lehoczky, I. (2022). Genetic Structure and Diversity of Native Tench (Tinca tinca L. 1758) Populations in Hungary—Establishment of Basic Knowledge Base for a Breeding Program. Diversity, 14(5), 336. https://doi.org/10.3390/d14050336