
Effect of Aquatic Vegetation Restoration after Removal of Culture Purse Seine on Phytoplankton Community Structure in Caizi Lakes


Abstract
:1. Introduction
2. Materials and Methods
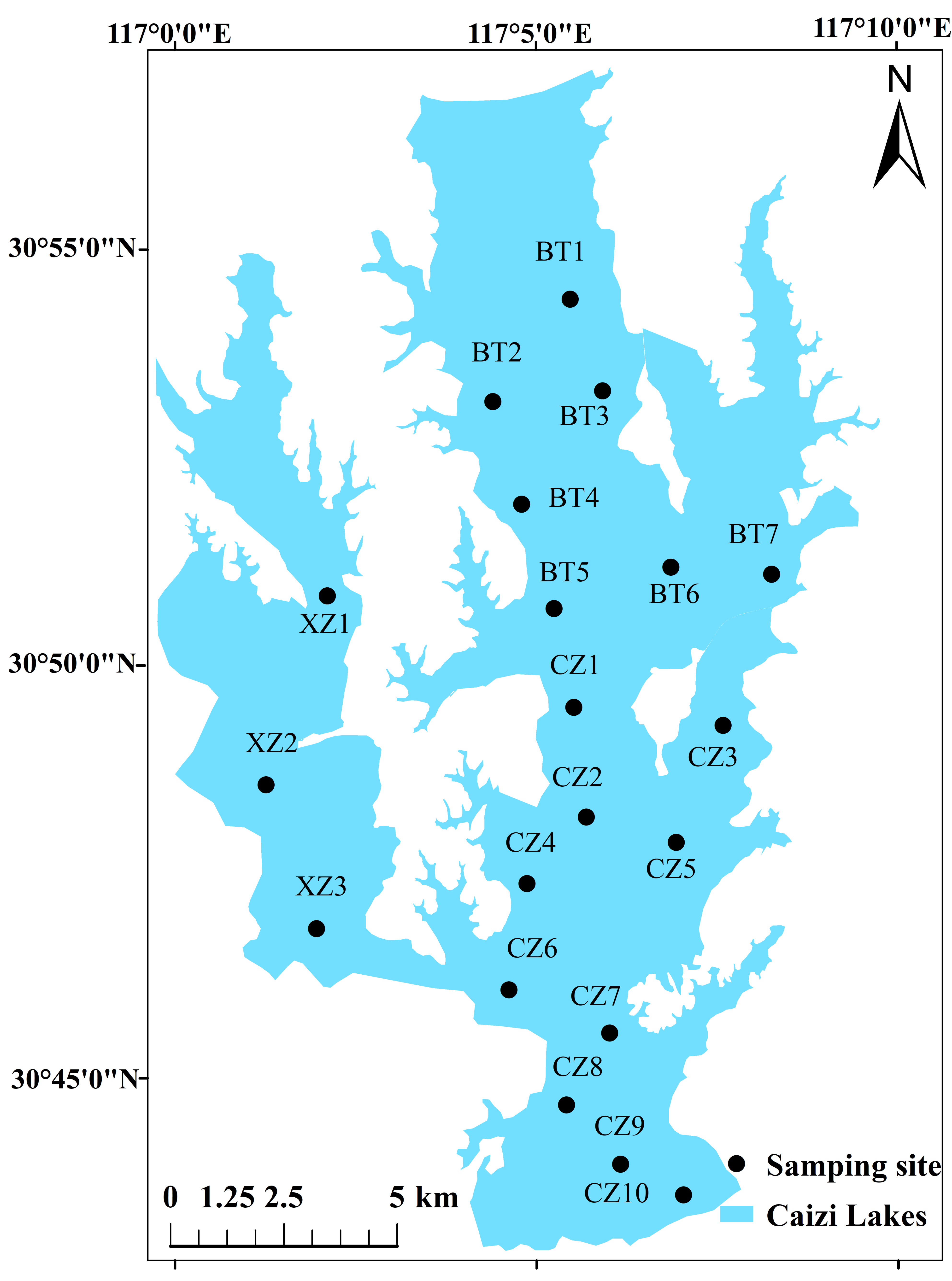
2.1. Description of Study Area
2.2. Sampling Point Setting and Sampling Time
2.3. Data Collection
2.4. Data Processing
3. Results
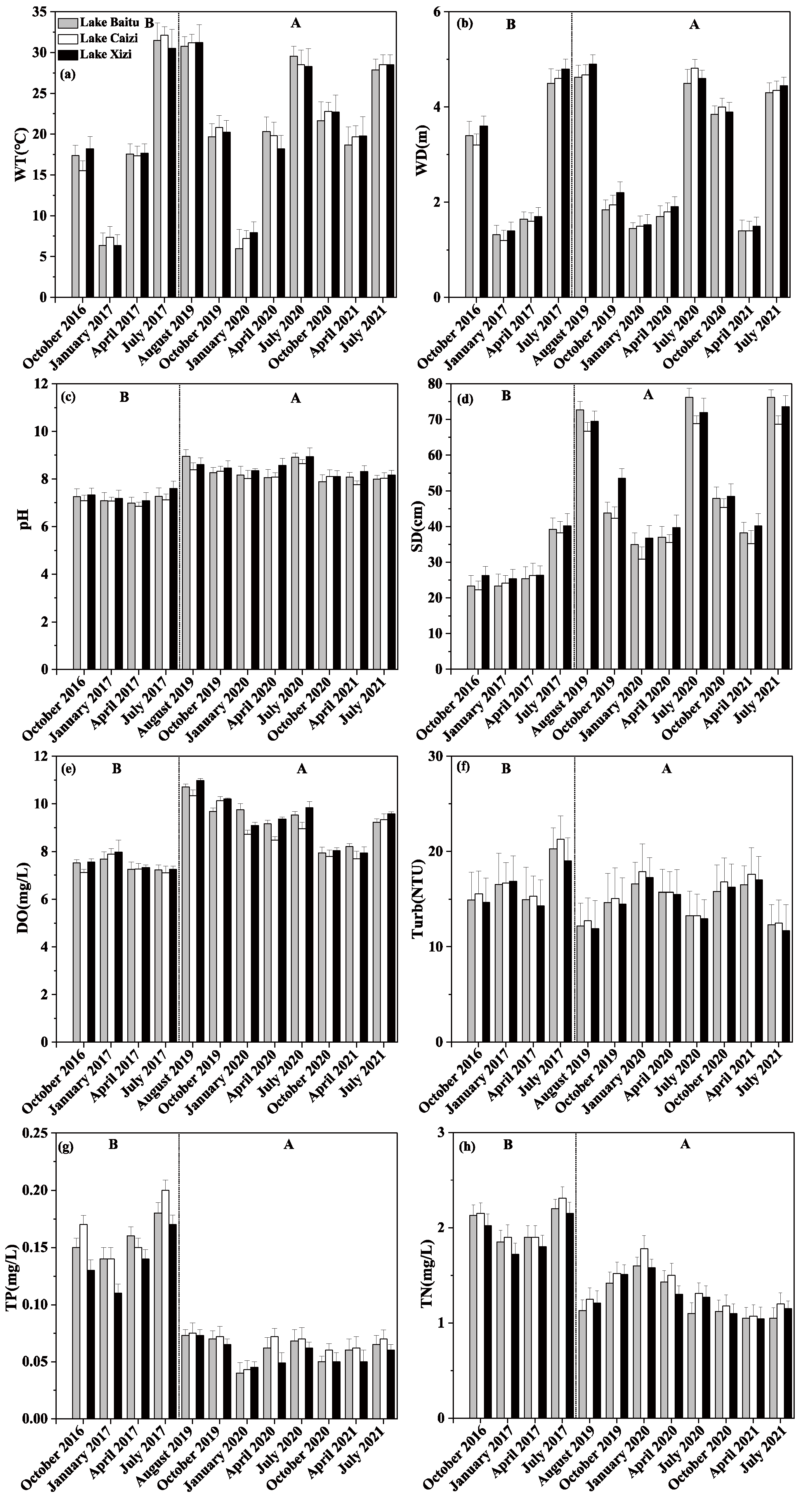
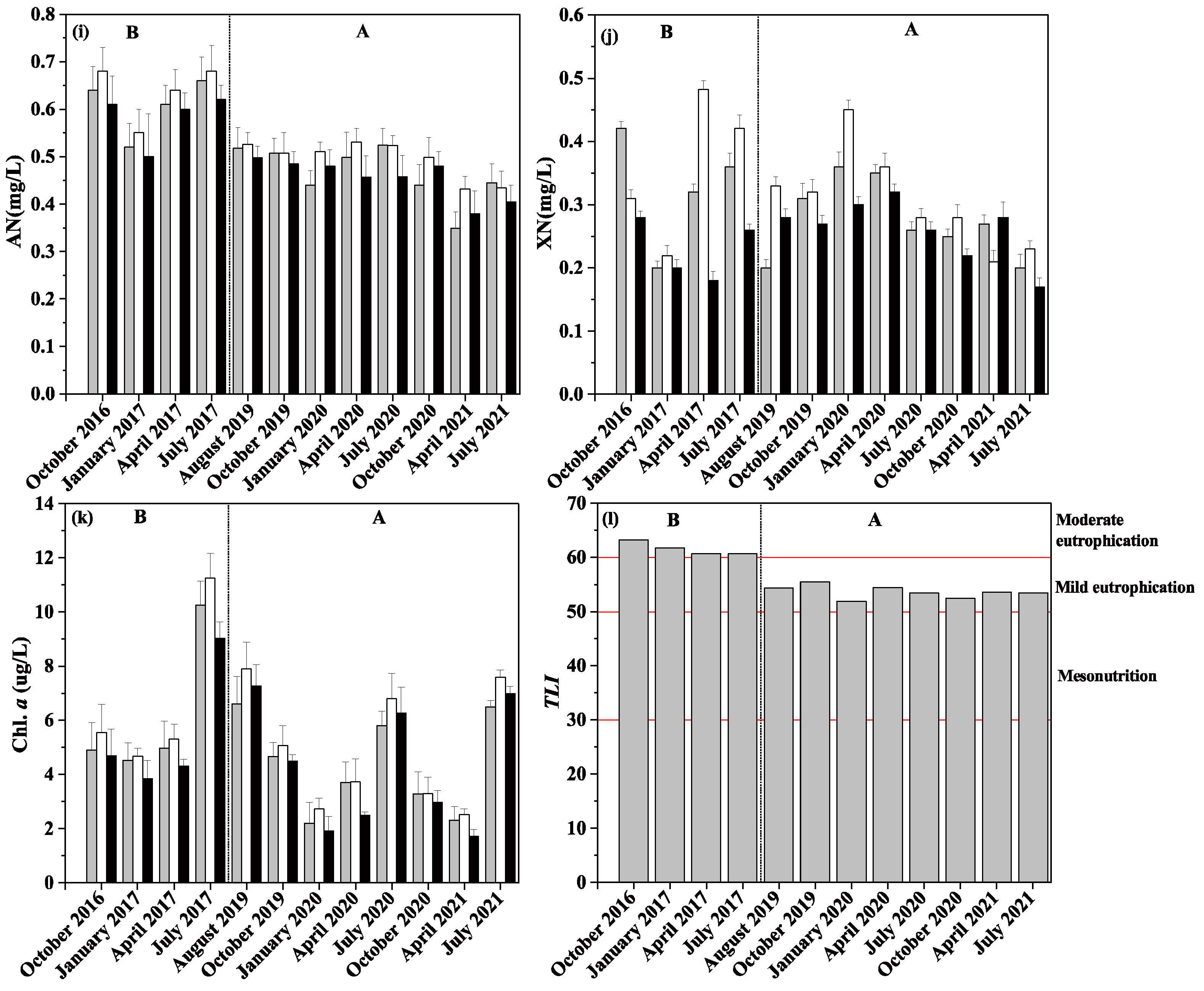
3.1. Physical and Chemical Indicators of Water Body
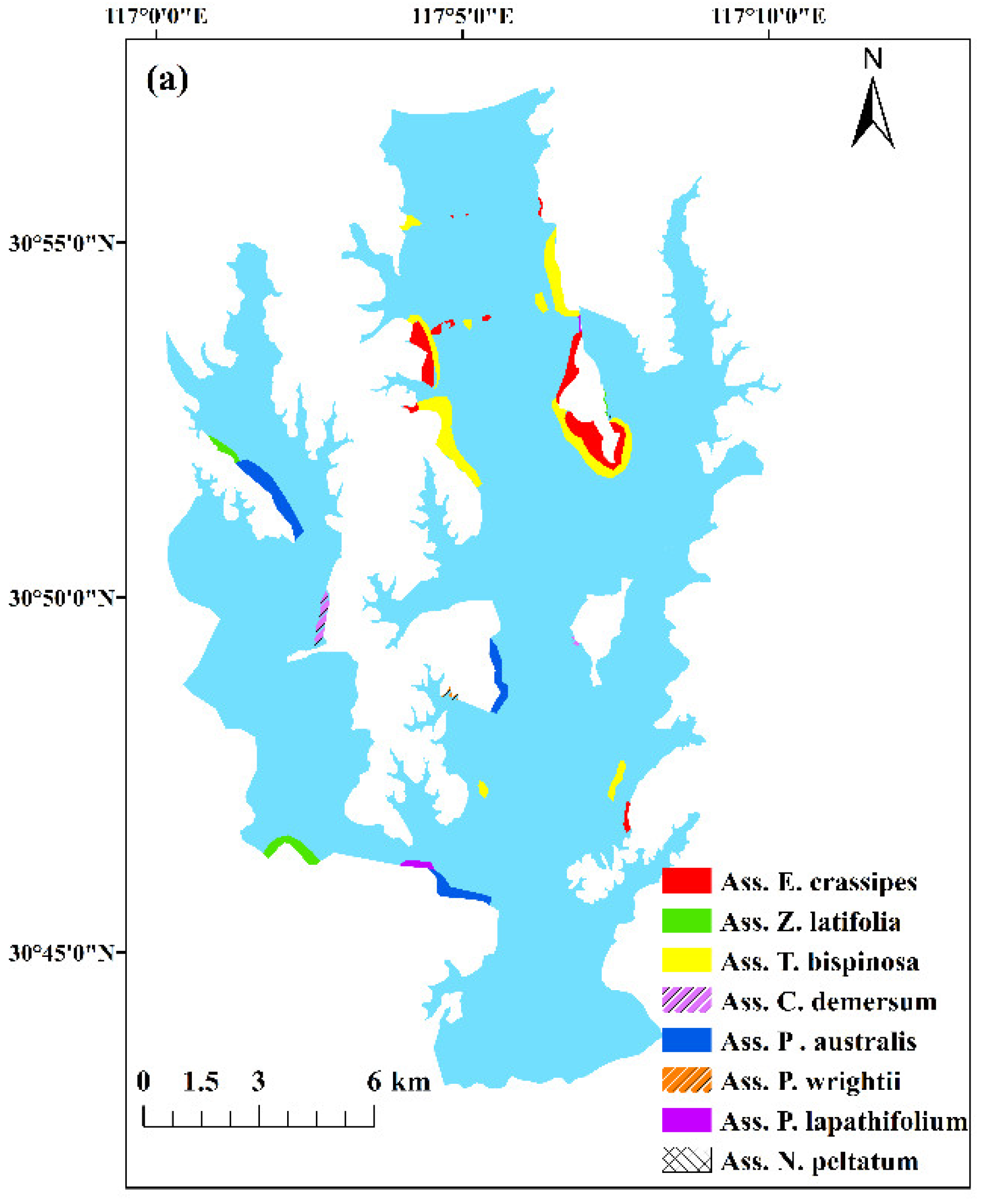
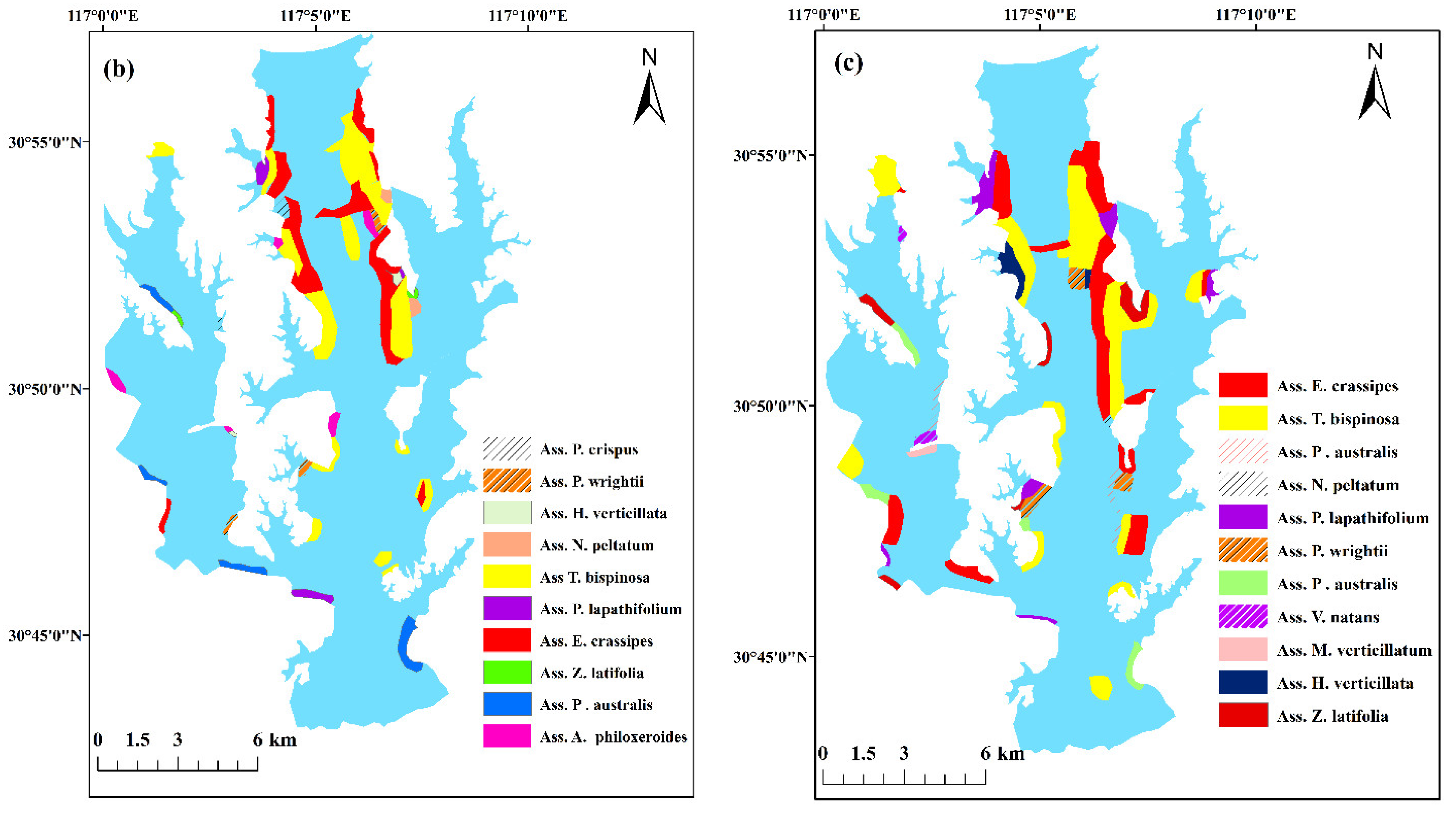
3.2. Distribution of Aquatic Vegetation
3.3. Temporal and Spatial Dynamics of Phytoplankton Cell Density, Biomass and Diversity Index
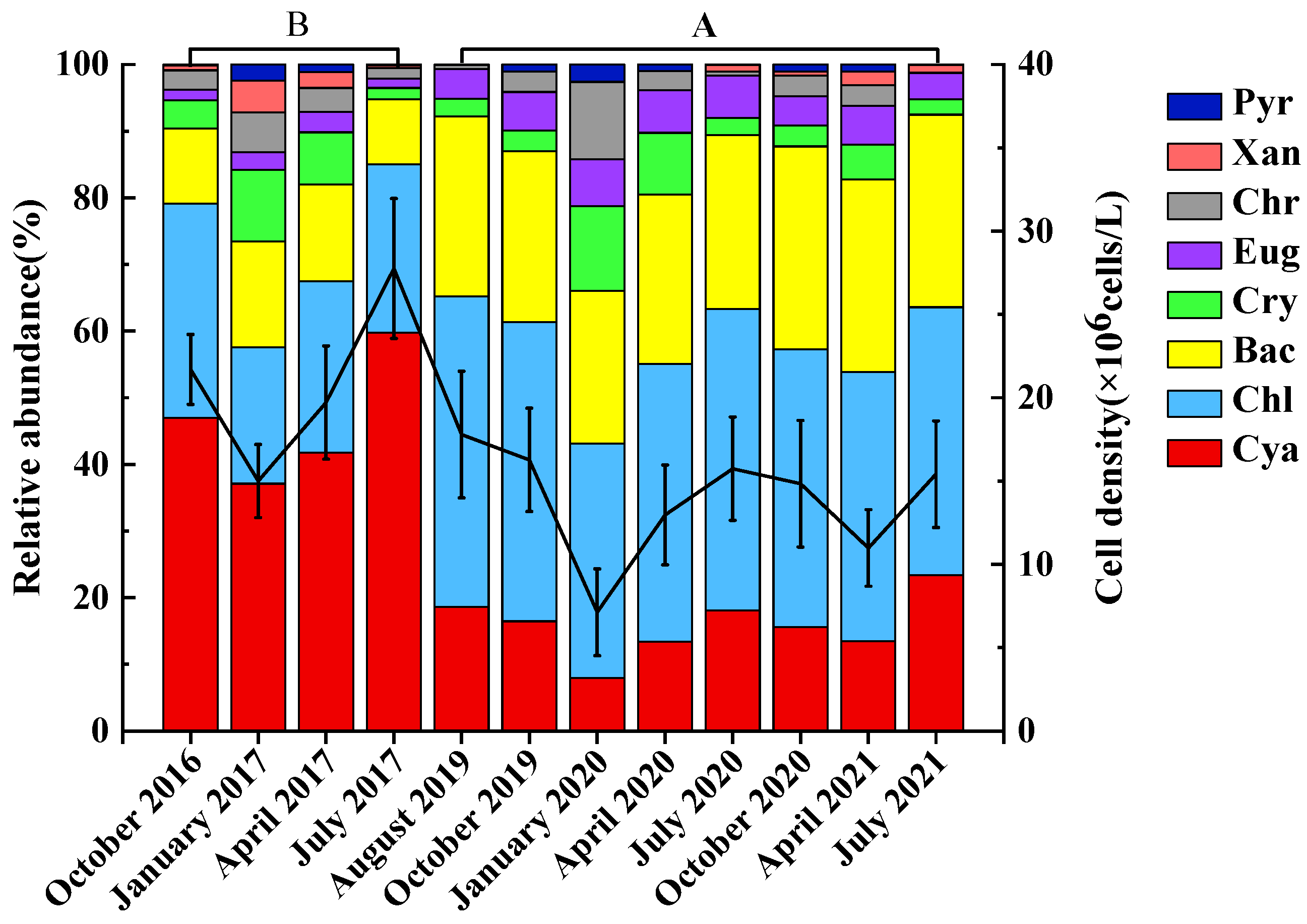
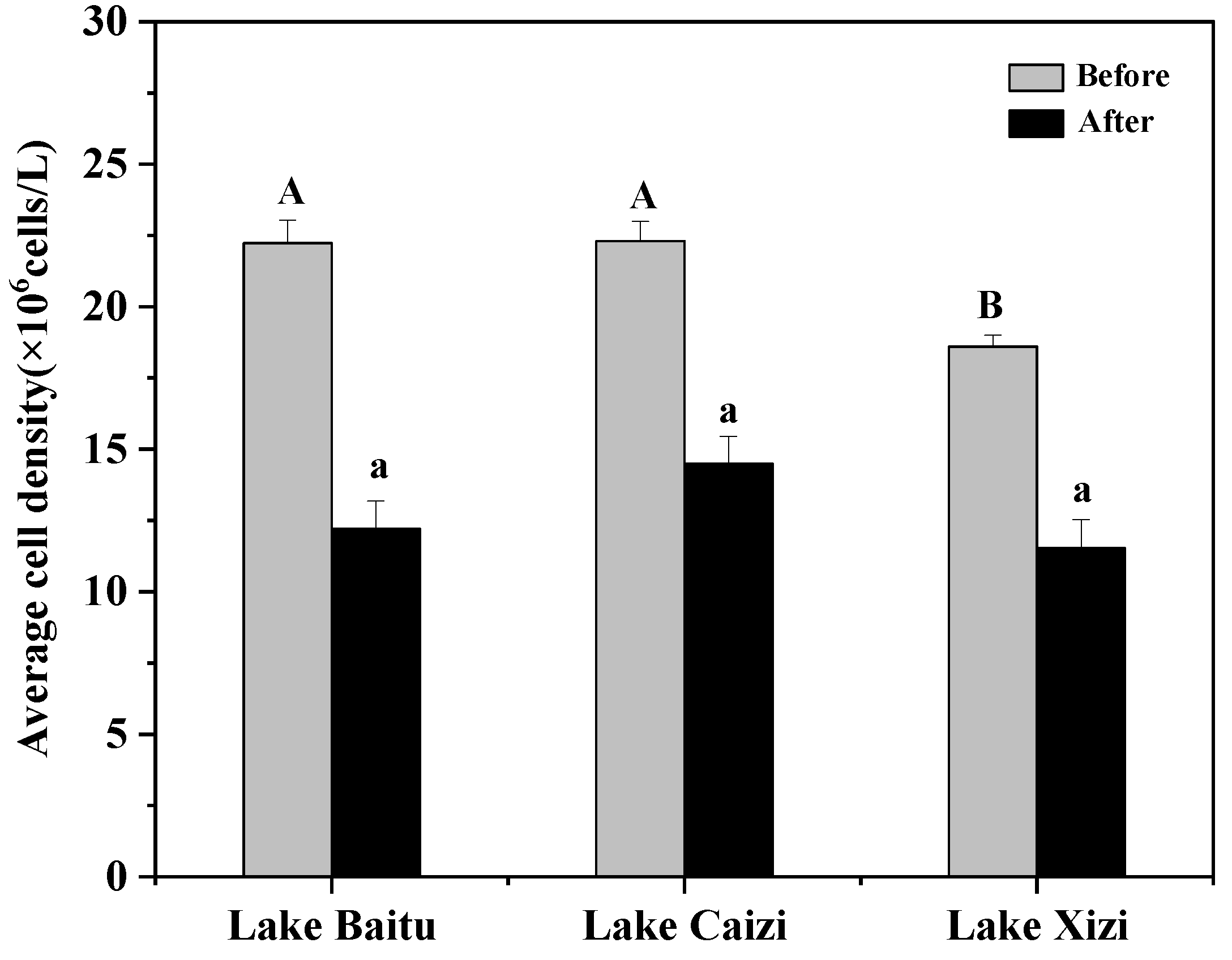
3.3.1. Temporal and Spatial Variation of Phytoplankton Cell Density
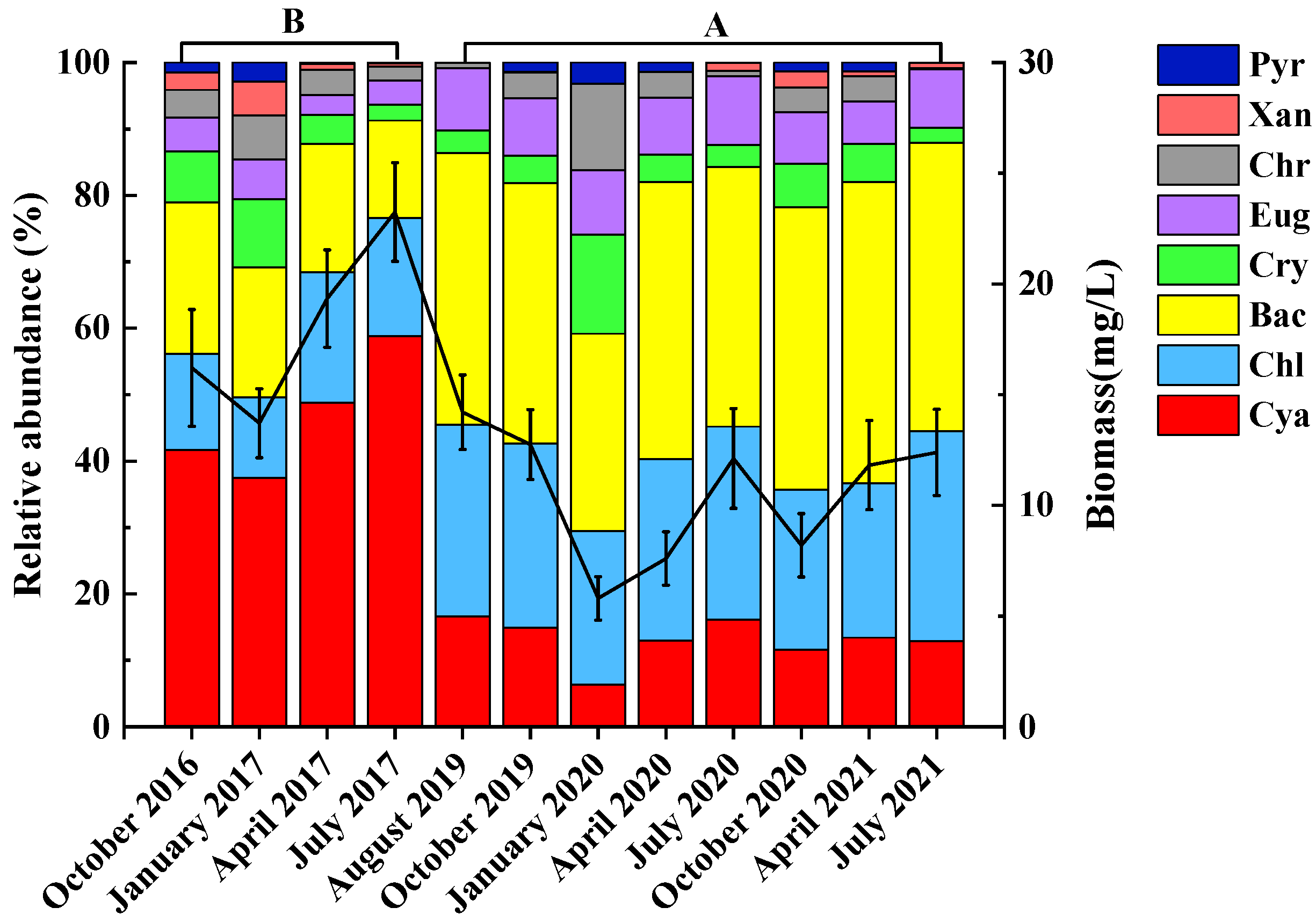
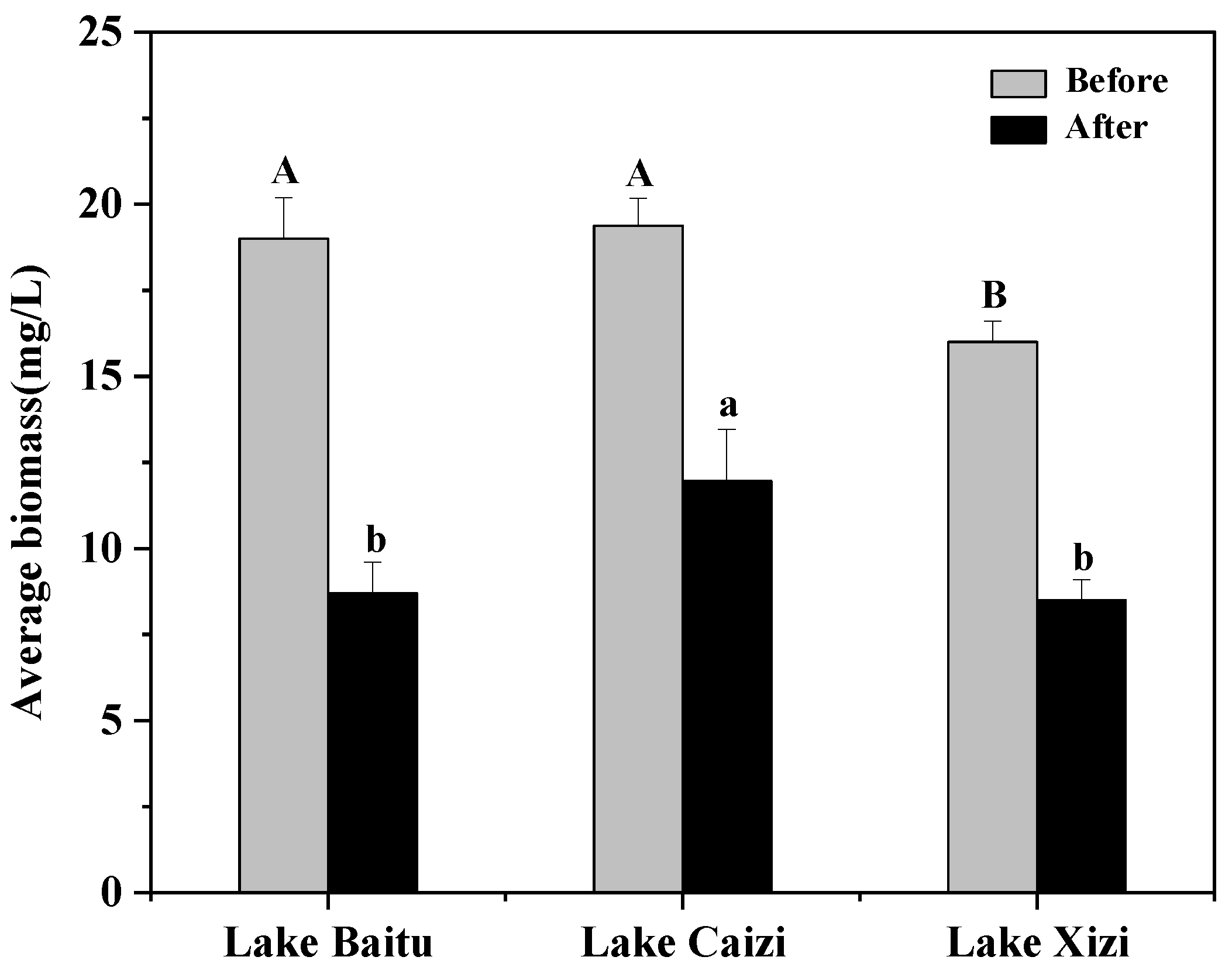
3.3.2. Temporal and Spatial Variation of Phytoplankton Biomass
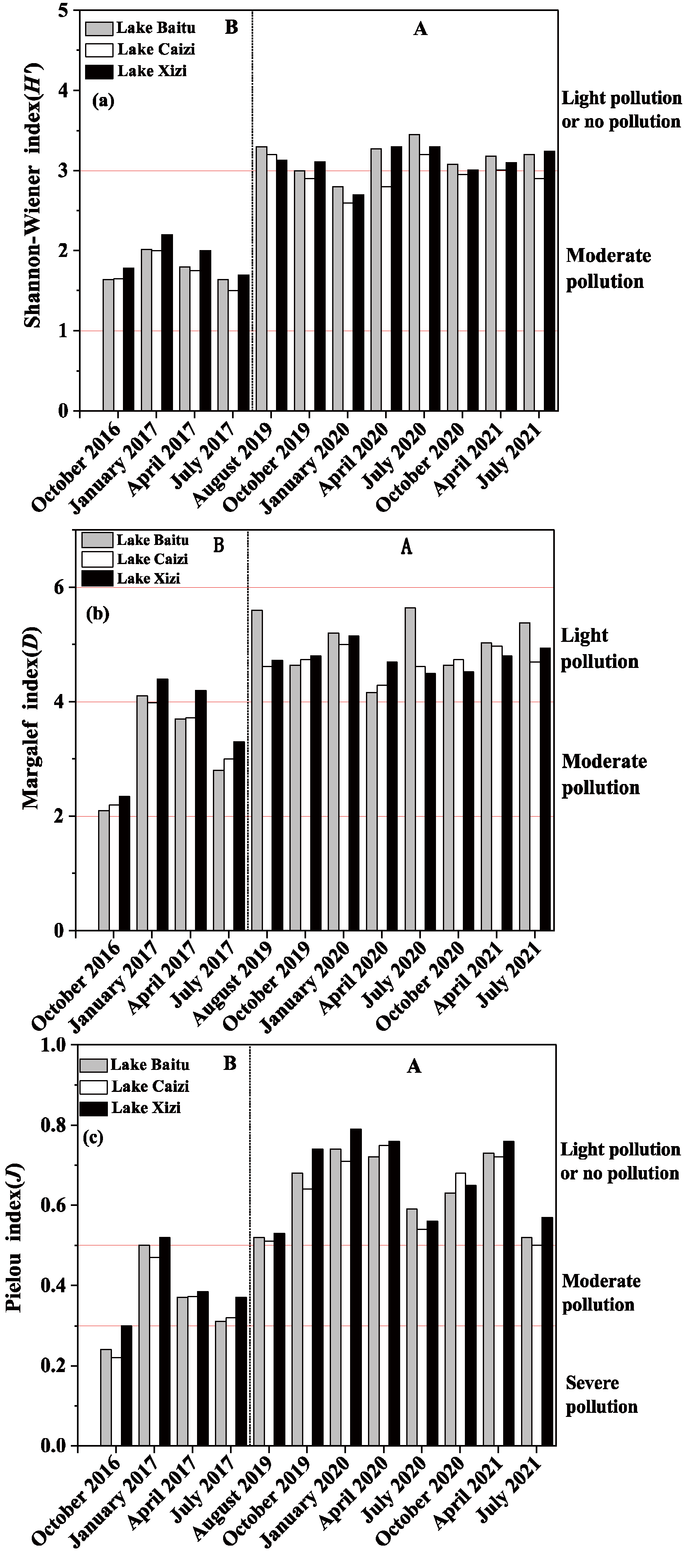
3.3.3. Temporal and Spatial Variation of Phytoplankton Diversity
3.4. Dominant Species of Phytoplankton
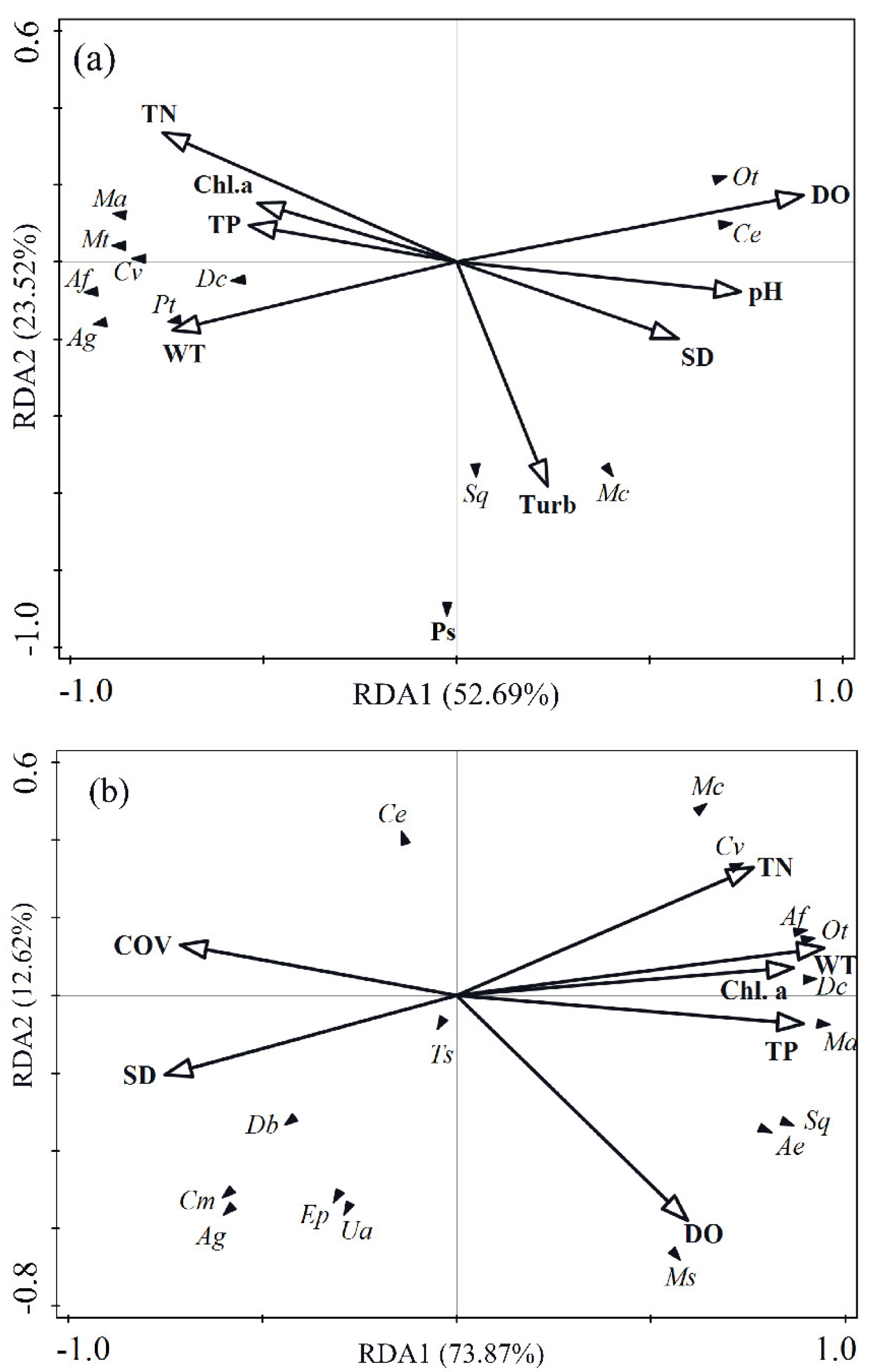
3.5. Relationship between Phytoplankton and Environmental Factors
4. Discussion
4.1. Restoration of the Aquatic Vegetation and Its Impact on the Water Environment
4.2. Effects of Aquatic Vegetation Restoration on the Cell Density, Biomass and Diversity of Phytoplankton
4.3. Effects of Aquatic Vegetation Restoration on Phytoplankton Species Change
4.4. Mechanism of Formation of Alternative States of the Lake
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Liu, X.; Wang, H. Estimation of minimum area requirement of river-connected lakes for fish diversity conservation in the Yangtze River floodplain. Divers. Distrib. 2010, 16, 932–940. [Google Scholar] [CrossRef]
- Song, X.X.; Cai, X.B.; Wang, Z.; Li, E.H.; Wang, X.L. Community change of dominant submerged macrophyte in Lake Honghu since 1950s. J. Lake Sci. 2016, 28, 859–867. [Google Scholar]
- Xie, Q.; Qian, L.; Liu, S.; Wang, Y.; Zhang, Y.; Wang, D. Assessment of long-term effects from cage culture practices on heavy metal accumulation in sediment and fish. Ecotoxicol. Environ. Saf. 2020, 194, 110433. [Google Scholar] [CrossRef] [PubMed]
- Ban, X.; Yu, C.; Wei, K. Analysis of lnfluence of Enclosure Aquaculture on Water Quality of Honghu Lake. Environ. Sci. Technol. 2010, 33, 125–129. [Google Scholar] [CrossRef]
- Yang, J.Z.C.; Luo, J.H.; Lu, L.R.; Sun, Z.; Cao, Z.G.; Zeng, Q.F.; Mao, Z.G. Changes in aquatic vegetation communities based on satellite images before and after pen aquaculture removal in East Lake Taihu. J. Lake Sci. 2021, 33, 507–517. [Google Scholar]
- Wang, Y.T.; Zhou, Z.Z.; Wan, X.; Zhang, Y.X. Effect of fast restoration of the aquatic vegetation on phytoplankton community after removal of purse seine culture in Huayanghe Lakes. Sci. Total Environ. 2021, 768, 144024. [Google Scholar] [CrossRef]
- Dhote, S.; Dixit, S. Water quality improvement through macrophytes—A review. Environ. Monit. Assess. 2009, 152, 149–153. [Google Scholar] [CrossRef]
- Rousseaux, C.S.; Gregg, W.W. Interannual variation in phytoplankton primary production at a global scale. Remote Sens. 2014, 6, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Santonja, M.; Le, R.B.; Thiebaut, G. Seasonal dependence and functional implications of macrophyte–phytoplankton allelopathic interaction. Freshw. Biol. 2018, 63, 1161–1172. [Google Scholar] [CrossRef]
- Mulderij, G.; Van Nes, E.H.; Van Donk, E. Macrophyte–phytoplankton interactions: The relative importance of allelopathy versus other factors. Ecol. Modell. 2007, 204, 85–92. [Google Scholar] [CrossRef]
- Felpeto, A.B.; Roy, S.; Vasconcelos, V.M. Allelopathy prevents competitive exclusion and promotes phytoplankton biodiversity. Oikos 2018, 127, 85–98. [Google Scholar] [CrossRef] [Green Version]
- Janssen, A.B.; Hilt, S.; Kosten, S.; de Klein, J.J.; Paerl, H.W.; Van de Waal, D.B. Shifting states, shifting services: Linking regime shifts to changes in ecosystem services of shallow lakes. Freshw. Biol. 2021, 66, 1–12. [Google Scholar] [CrossRef]
- Tamire, G.; Mengistou, S.; Degefe, G. Potential allelopathic impact of Potamogeton schweinfurthii on phytoplankton in Lake Ziway, Ethiopia. Inland Waters 2016, 6, 336–342. [Google Scholar] [CrossRef]
- Havens, K.E.; Jin, K.R.; Rodusky, A.J.; Sharfstein, B.; Brady, M.A.; East, T.L.; Iricanin, N.; James, R.T.; Harwell, M.C.; Steinman, A.D. Hurricane effects on a shallow lake ecosystem and its response to a controlled manipulation of water level. Sci. World J. 2001, 1, 44–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chia, A.M.; Iortsuun, D.N.; Stephen, B.J.; Ayobamire, A.E.; Ladan, Z. Phytoplankton responses to changes in macrophyte density in a tropical artificial pond in Zaria, Nigeria. Afr. J. Aquat. Sci. 2011, 36, 35–46. [Google Scholar] [CrossRef]
- Wang, X.Y.; Jiang, B.; Tian, Z.F.; Cai, J.Z.; Lin, G.J. Impact of water level changes in Lake Caizi (Anhui Province) on main wetland types and wintering bird habitat during wintering period. J. Lake Sci. 2018, 30, 1636–1645. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.; Xu, N.; Liu, B.; Zhou, L.; Wang, J.; Wang, C.; Dai, B.G.; Xiong, W. Metal concentrations and risk assessment in water, sediment and economic fish species with various habitat preferences and trophic guilds from Lake Caizi, Southeast China. Ecotoxicol. Environ. Saf. 2018, 157, 1–8. [Google Scholar] [CrossRef]
- Cheng, B.; Jiang, B.; Li, H.Q. Design and realization of Caizi Lake wetland ecological database management system. Water Resour. Prot. 2020, 36, 46–52. [Google Scholar]
- Guo, W.L.; Zhou, Z.Z.; Chen, J.W.; Zheng, X.D.; Ye, X.X. Effects of extreme flooding on aquatic vegetation cover in Shengjin Lake, China. Hydrol. Processes 2022, 36, e14459. [Google Scholar] [CrossRef]
- SEPA. State EPA of China, Monitoring and Analysis Methods for Water and Wastewater, 4th ed.; China Environmental Science Press: Beijing, China, 2002; pp. 223–281. [Google Scholar]
- Hu, H.J.; Wei, Y.X. Freshwater Algae in China: System, Classification and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Shannon, C.E.; Weaver, W. The Mathematical Theory of Communication; University of Illinois Press: Champaign, IL, USA, 1949. [Google Scholar]
- Pielou, E.C. An Introduction to Mathematical Ecology; John Wiley & Sons: Hoboken, NJ, USA, 1969. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Wang, M.C.; Liu, X.Q.; Zhang, J.H. Evaluate method and classification standard on lake eutrophication. Environ. Monit. 2002, 18, 47–49. [Google Scholar] [CrossRef]
- Tan, F.X.; Luo, J.B.; Gong, S.S.; Zhou, W.B.; Xiang, M.M.; Meng, J.X.; Chai, Y. Effects of removal of the breeding seine on aquatic plant diversity in Yuanxinhu area of Chana lake. Hubei Agric. Sci. 2019, 58, 92–96. [Google Scholar] [CrossRef]
- Zhong, J.X.; Lin, X.T.; Xu, Z.N.; Jie, X.Y. The top-down effect of fish stocking on the freshwater environment (A review). J. Jinan Univ. 2001, 22, 131–136. [Google Scholar]
- Madsen, J.D.; Chambers, P.A.; James, W.F.; Koch, E.W.; Westlake, D.F. The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia 2001, 444, 71–84. [Google Scholar] [CrossRef]
- Ferreira, T.F.; Crossetti, L.O.; Marques, D.M.M.; Cardoso, L.; Fragoso Jr, C.R.; van Nes, E.H. The structuring role of submerged macrophytes in a large subtropical shallow lake: Clear effects on water chemistry and phytoplankton structure community along a vegetated-pelagic gradient. Limnologica 2018, 69, 142–154. [Google Scholar] [CrossRef]
- Preiner, S.; Dai, Y.; Pucher, M.; Reitsema, R.E.; Schoelynck, J.; Meire, P.; Hein, T. Effects of macrophytes on ecosystem metabolism and net nutrient uptake in a groundwater fed lowland river. Sci. Total Environ. 2020, 721, 137620. [Google Scholar] [CrossRef]
- Madsen, T.V.; Cedergreen, N. Sources of nutrients to rooted submerged macrophytes growing in a nutrient-rich stream. Freshw. Biol. 2002, 47, 283–291. [Google Scholar] [CrossRef]
- Riis, T.; Tank, J.L.; Reisinger, A.J.; Aubenau, A.; Roche, K.R.; Levi, P.S.; Baattrup-Pedersen, A.; Alnoee, A.B.; Bolster, D. Riverine macrophytes control seasonal nutrient uptake via both physical and biological pathways. Freshw. Biol. 2020, 65, 178–192. [Google Scholar] [CrossRef]
- Wang, W.H.; Wang, Y.; Sun, L.Q.; Zheng, Y.C.; Zhao, J.C. Research and application status of ecological floating bed in eutrophic landscape water restoration. Sci. Total Environ. 2020, 704, 135434. [Google Scholar] [CrossRef]
- Yin, X.; Lu, J.; Wang, Y.; Liu, G.; Hua, Y.; Wan, X.; Zhao, J.; Zhu, D. The abundance of nirS-type denitrifiers and anammox bacteria in rhizospheres was affected by the organic acids secreted from roots of submerged macrophytes. Chemosphere 2020, 240, 124903. [Google Scholar] [CrossRef]
- Bicudo, D.C.; Fonseca, B.M.; Bini, L.M.; Crossetti, L.O.; Bicudo, C.; Araújo de Jesus, T. Undesirable side-effects of water hyacinth control in a shallow tropical reservoir. Freshw. Boil. 2007, 52, 1120–1133. [Google Scholar] [CrossRef]
- Liu, C.Q.; Yang, J.; Ma, X.L.; Liu, L.S.; Wang, Y.; Shu, J.M. Ecological Restoration Using Trapa bispinosa and Nelumbo nucifera on Eutrophic Water Body in Baiyangdian Lake. Wetl. Sci. 2013, 11, 510–514. [Google Scholar] [CrossRef]
- Shen, W.; Huang, X.Q.; Luo, X.; Zhou, K.X.; Ling, H.; Xie, B.W. Comparative Study of Purification Capacity in Aquaculture Water Between Lythrum salicaria and Eichhornia crassipes. J. Neijiang Norm. Univ. 2017, 32, 77–81. [Google Scholar] [CrossRef]
- Bian, G.G. Allelopathy mechanism of floating plant on algae and the application. Acta Hydrobiol. Sin. 2012, 36, 978–982. [Google Scholar] [CrossRef]
- Nurminen, L.; Horpilla, J.; Tallberg, P. Seasonal development of the cladoceran assemblage in a turbid lake: The role of emergent macrophytes. Arch. Hydrobiol. 2001, 151, 127–140. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Huang, L.; Wang, Y.; Li, L.; Li, G.; Xiao, B.; Song, L. Reducing the phytoplankton biomass to promote the growth of submerged macrophytes by introducing artificial aquatic plants in shallow eutrophic waters. Water 2019, 11, 1370. [Google Scholar] [CrossRef] [Green Version]
- Kang, L.J.; Xu, H.; Zou, W.; Zhu, G.W.; Zhu, M.Y.; Ji, P.F.; Chen, J. Influence of Potamogeton crispus on Lake Water Environment and Phytoplankton Community Structure. Environ. Sci. 2020, 41, 4053–4061. [Google Scholar] [CrossRef]
- Wu, F.Q.; Liu, T.M.; Wang, Z.T.; Wang, Y.H.; He, S.Z. Effects of Eichhornia crassipes Growth on Aquatic Plant in Dianchi Lake. J. Anhui Agric. Sci. 2011, 39, 9167–9168. [Google Scholar] [CrossRef]
- Zhou, Q.; Han, S.Q.; Yan, S.H.; Song, W.; Liu, G.F. Impacts of Eichhornia crassipes (Mart.) Solms Stress on the Growth Characteristics, Microcystins and Nutrients Release of Microcystis aeruginosa. Environ. Sci. 2014, 35, 597–604. [Google Scholar] [CrossRef]
- Song, Y.Z.; Zhu, G.W.; Qin, B.Q. Applicability analysis of aquatic macrophytes on controlling nitrogen and phosphorus from water in the Kangshan Bay demonstration area of Lake Taihu. J. Lake Sci. 2013, 25, 259–265. [Google Scholar] [CrossRef] [Green Version]
- Reynolds, C.S. The Ecology of Phytoplankton; Cambridge University Press: Cambridge, UK, 2006. [Google Scholar] [CrossRef]
- Søndergaard, M.; Moss, B. Impact of submerged macrophytes on phytoplankton in shallow freshwater lakes. In The Structuring Role of Submerged Macrophytes in Lakes; Springer: New York, NY, USA, 1998; pp. 115–132. [Google Scholar]
- Morris, K.; Bailey, P.C.; Boon, P.I.; Hughes, L. Effects of plant harvesting and nutrient enrichment on phytoplankton community structure in a shallow urban lake. Hydrobiologia 2006, 571, 77–91. [Google Scholar] [CrossRef]
- Scheffer, M.; Hosper, S.H.; Meijer, M.L.; Moss, B.; Jeppesen, E. Alternative equilibria in shallow lakes. Trends Ecol. Evol. 1993, 8, 275–279. [Google Scholar] [CrossRef]
- Kéfi, S.; Holmgren, M.; Scheffer, M. When can positive interactions cause alternative stable states in ecosystems? Funct. Ecol. 2016, 30, 88–97. [Google Scholar] [CrossRef]
- Ibelings, B.W.; Portielje, R.; Lammens, E.H.; Noordhuis, R.; Van den Berg, M.S.; Joosse, W.; Meijer, M.L. Resilience of alternative stable states during the recovery of shallow lakes from eutrophication: Lake Veluwe as a case study. Ecosystems 2007, 10, 4–16. [Google Scholar] [CrossRef] [Green Version]
- Van Nes, E.H.; Scheffer, M.; Van den Berg, M.S.; Coops, H. Dominance of charophytes in eutrophic shallow lakes—When should we expect it to be an alternative stable state? Aquat. Bot. 2002, 72, 275–296. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| WT (°C) | WD (m) | pH | SD (cm) | DO (mg/L) | TP (mg/L) | TN (mg/L) | NH4+-N (mg/L) | NO3−-N (mg/L) | Turb (NTU) | CODMn (mg/L) | Chl. a (ug/L) | ||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before | October 2016 | 16.9 ± 1.09 | 3.4 ± 0.16 | 7.23 ± 0.1 | 24.36 ± 1.63 | 7.34 ± 0.18 | 0.15 ± 0.02 | 2.1 ± 0.06 | 0.64 ± 0.03 | 0.34 ± 0.06 | 15.05 ± 0.37 | 12.31 ± 0.74 | 5.13 ± 0.36 |
| January 2017 | 6.89 ± 0.41 | 1.3 ± 0.08 | 7.12 ± 0.05 | 24.79 ± 0.47 | 7.93 ± 0.03 | 0.13 ± 0.01 | 1.82 ± 0.08 | 0.52 ± 0.02 | 0.21 ± 0.01 | 16.68 ± 0.13 | 11.45 ± 0.42 | 4.27 ± 0.34 | |
| April 2017 | 17.52 ± 0.13 | 1.65 ± 0.04 | 6.98 ± 0.1 | 26.36 ± 0.02 | 7.29 ± 0.02 | 0.15 ± 0.01 | 1.87 ± 0.05 | 0.62 ± 0.02 | 0.33 ± 0.12 | 14.87 ± 0.42 | 12.02 ± 0.34 | 4.82 ± 0.41 | |
| July 2017 | 31.34 ± 0.65 | 4.7 ± 0.08 | 7.34 ± 0.2 | 39.25 ± 0.82 | 7.18 ± 0.06 | 0.18 ± 0.01 | 2.22 ± 0.07 | 0.65 ± 0.02 | 0.35 ± 0.07 | 20.18 ± 0.91 | 10.69 ± 0.41 | 10.15 ± 0.91 | |
| After | August 2019 | 31.23 ± 0.22 | 4.79 ± 0.09 | 8.65 ± 0.23 | 68.12 ± 1.13 | 10.67 ± 0.26 | 0.07 ± 0.00 | 1.23 ± 0.02 | 0.51 ± 0.01 | 0.27 ± 0.05 | 12.29 ± 0.34 | 9.49 ± 1.00 | 7.59 ± 0.26 |
| October 2019 | 20.54 ± 0.24 | 2.08 ± 0.1 | 8.36 ± 0.08 | 47.94 ± 4.58 | 10.17 ± 0.02 | 0.07 ± 0.00 | 1.48 ± 0.04 | 0.5 ± 0.01 | 0.3 ± 0.02 | 14.75 ± 0.25 | 10.68 ± 0.56 | 4.79 ± 0.24 | |
| January 2020 | 7.59 ± 0.3 | 1.52 ± 0.01 | 8.18 ± 0.14 | 33.85 ± 2.41 | 8.91 ± 0.15 | 0.04 ± 0.00 | 1.65 ± 0.1 | 0.48 ± 0.03 | 0.37 ± 0.06 | 17.26 ± 0.53 | 10.68 ± 0.43 | 2.33 ± 0.33 | |
| April 2020 | 19.03 ± 0.65 | 1.86 ± 0.04 | 8.24 ± 0.24 | 37.7 ± 1.72 | 8.92 ± 0.36 | 0.06 ± 0.01 | 1.38 ± 0.09 | 0.5 ± 0.03 | 0.34 ± 0.02 | 15.65 ± 0.11 | 10.21 ± 0.64 | 3.12 ± 0.51 | |
| July 2020 | 28.42 ± 0.1 | 4.71 ± 0.09 | 8.84 ± 0.13 | 70.43 ± 1.28 | 9.4 ± 0.37 | 0.07 ± 0.00 | 1.23 ± 0.09 | 0.5 ± 0.03 | 0.27 ± 0.01 | 13.19 ± 0.14 | 9.45 ± 0.37 | 6.54 ± 0.22 | |
| October 2020 | 22.77 ± 0.03 | 3.95 ± 0.04 | 8.04 ± 0.11 | 46.92 ± 1.29 | 7.91 ± 0.1 | 0.05 ± 0 | 1.13 ± 0.04 | 0.47 ± 0.02 | 0.25 ± 0.02 | 16.29 ± 0.41 | 10.25 ± 0.36 | 3.14 ± 0.13 | |
| April 2021 | 19.75 ± 0.04 | 1.45 ± 0.04 | 8.05 ± 0.23 | 37.72 ± 2.04 | 7.83 ± 0.1 | 0.06 ± 0.01 | 1.03 ± 0.03 | 0.39 ± 0.03 | 0.2 ± 0.02 | 17.03 ± 0.45 | 10.24 ± 0.34 | 2.12 ± 0.33 | |
| July 2021 | 28.53 ± 0.33 | 4.4 ± 0.04 | 8.07 ± 0.08 | 71.19 ± 2.04 | 9.46 ± 0.1 | 0.07 ± 0 | 1.15 ± 0.04 | 0.43 ± 0.02 | 0.24 ± 0.04 | 12.19 ± 0.34 | 9.36 ± 0.41 | 7.3 ± 0.24 |
| Phylum | Dominant Species | Dominance of Phytoplankton Species before Restoration (Y) | Dominance of Phytoplankton Species after Restoration (Y) | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Spring | Summer | Autumn | Winter | Spring | Summer | Autumn | Winter | ||
| Cyanobacteria | M. aeruginosa | 0.06 | 0.19 | 0.18 | 0.05 | 0.03 | 0.02 | ||
| P. tenue | 0.02 | 0.17 | 0.04 | ||||||
| O. tenuis | 0.22 | 0.11 | 0.03 | 0.02 | |||||
| D. circinale | 0.05 | 0.13 | 0.03 | ||||||
| A. flos-aquae | 0.02 | 0.02 | 0.02 | ||||||
| M. tenuissima | 0.02 | 0.02 | |||||||
| A. elachista | 0.02 | ||||||||
| Chlorophyta | M. contortum | 0.04 | 0.03 | 0.02 | |||||
| S. quadricauda | 0.03 | 0.12 | 0.03 | 0.06 | |||||
| C. vulgaris | 0.02 | 0.05 | 0.02 | 0.02 | 0.05 | 0.05 | |||
| M. simplex | 0.02 | 0.03 | 0.02 | ||||||
| Bacillariophyta | A. granulata | 0.02 | 0.12 | 0.03 | 0.16 | 0 | 0.02 | 0.02 | 0.03 |
| U. acus | 0.02 | 0.04 | 0.06 | ||||||
| E. perpusillum | 0.02 | 0.02 | 0.02 | ||||||
| C. meneghiniana | 0.02 | 0.02 | |||||||
| Cryptophyta | C. erosa | 0.02 | 0.03 | 0.04 | |||||
| Euglenophyta | T. superba | 0.02 | |||||||
| Chrysophyta | D. bavaricum | 0.03 | |||||||
| Parameter | Before Aquatic Vegetation Restoration | After Aquatic Vegetation Restoration | ||
|---|---|---|---|---|
| Cell Density | Biomass | Cell Density | Biomass | |
| r | r | r | r | |
| WT | 0.965 ** | 0.936 ** | 0.934 ** | 0.809 * |
| WD | 0.944 ** | 0.697 | 0.723 * | 0.497 |
| SD | 0.801 * | 0.860 ** | 0.751 * | 0.715 * |
| pH | 0.408 | 0.187 | 0.439 | 0.436 |
| DO | −0.484 | −0.583 | 0.387 | 0.423 |
| TP | 0.961 ** | 0.923 ** | 0.891 ** | 0.879 ** |
| TN | 0.874 ** | 0.614 | 0.171 | 0.434 |
| AN | 0.942 ** | 0.769 * | 0.327 | −0.064 |
| XN | 0.884 ** | 0.786 * | −0.426 | −0.678 |
| Turb | 0.543 | 0.54 | −0.740 * | −0.713 * |
| Chl. a | 0.890 ** | 0.871 ** | 0.795 * | 0.706 |
| N/P | −0.496 | −0.780 * | −0.707 | −0.466 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, W.; Liu, Z.; Guo, W.; Zhou, Z. Effect of Aquatic Vegetation Restoration after Removal of Culture Purse Seine on Phytoplankton Community Structure in Caizi Lakes. Diversity 2022, 14, 395. https://doi.org/10.3390/d14050395
Zhao W, Liu Z, Guo W, Zhou Z. Effect of Aquatic Vegetation Restoration after Removal of Culture Purse Seine on Phytoplankton Community Structure in Caizi Lakes. Diversity. 2022; 14(5):395. https://doi.org/10.3390/d14050395
Chicago/Turabian StyleZhao, Wenqian, Zhenzhong Liu, Wenli Guo, and Zhongze Zhou. 2022. "Effect of Aquatic Vegetation Restoration after Removal of Culture Purse Seine on Phytoplankton Community Structure in Caizi Lakes" Diversity 14, no. 5: 395. https://doi.org/10.3390/d14050395
APA StyleZhao, W., Liu, Z., Guo, W., & Zhou, Z. (2022). Effect of Aquatic Vegetation Restoration after Removal of Culture Purse Seine on Phytoplankton Community Structure in Caizi Lakes. Diversity, 14(5), 395. https://doi.org/10.3390/d14050395