
Biogeography and Diversification of the Tropical and Subtropical Asian Genus Gastrochilus (Orchidaceae, Aeridinae)

 ,
, {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Taxon Sampling and Molecular Data
2.2. Phylogenetic Analysis
2.3. Time Estimation
2.4. Biogeographical Analyses
2.5. Ancestral State Reconstruction and Correlates of Diversification
2.6. Diversification Analysis
2.7. Collection of Species Distribution Data and Environmental Variables
2.8. Estimation of Evolutionary Rate in Niche Traits
2.9. Detection of Phylogenetic Signals of Niche Traits
3. Results
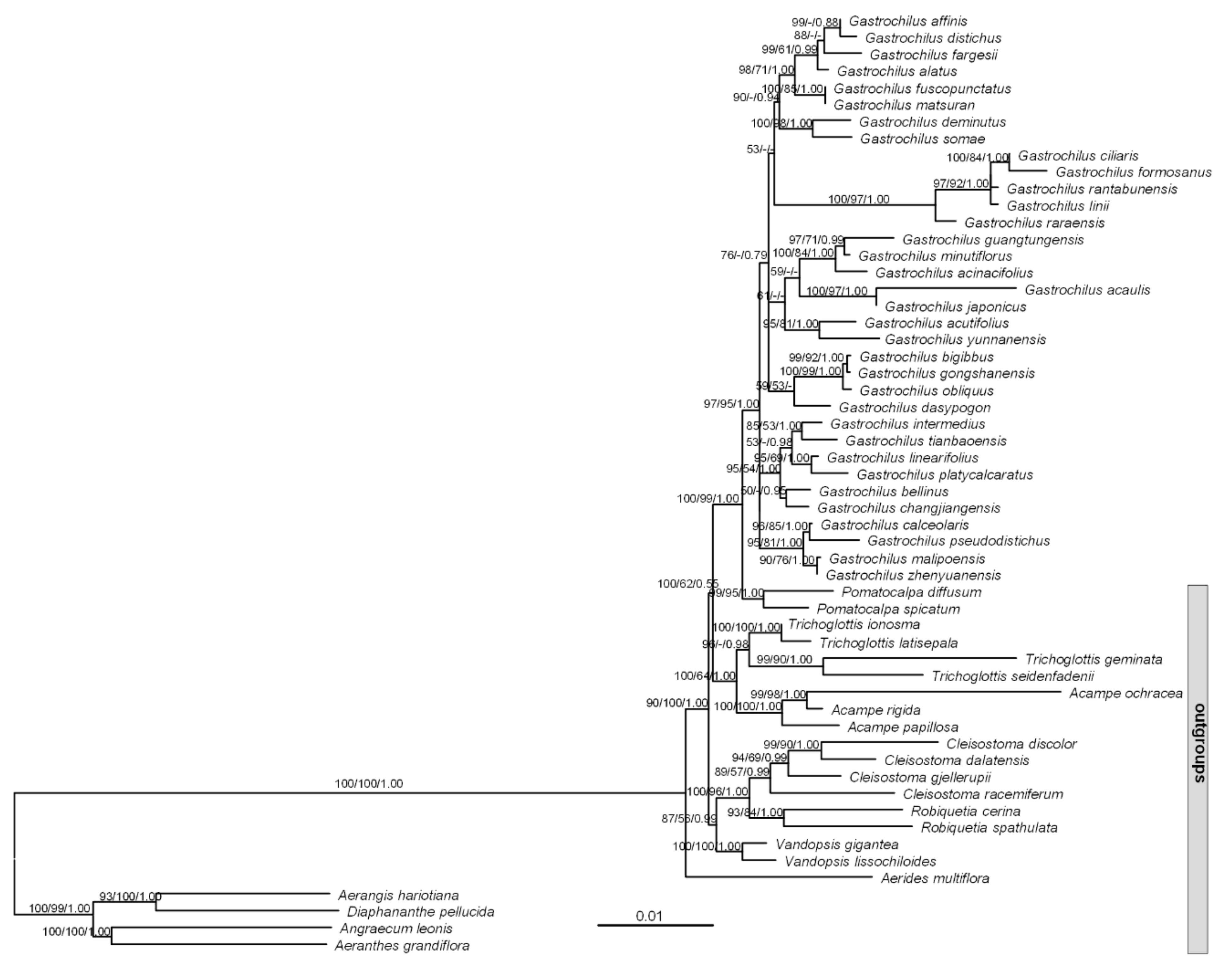
3.1. Phylogenetic Relationships and Divergence Time Estimates within Gastrochilus
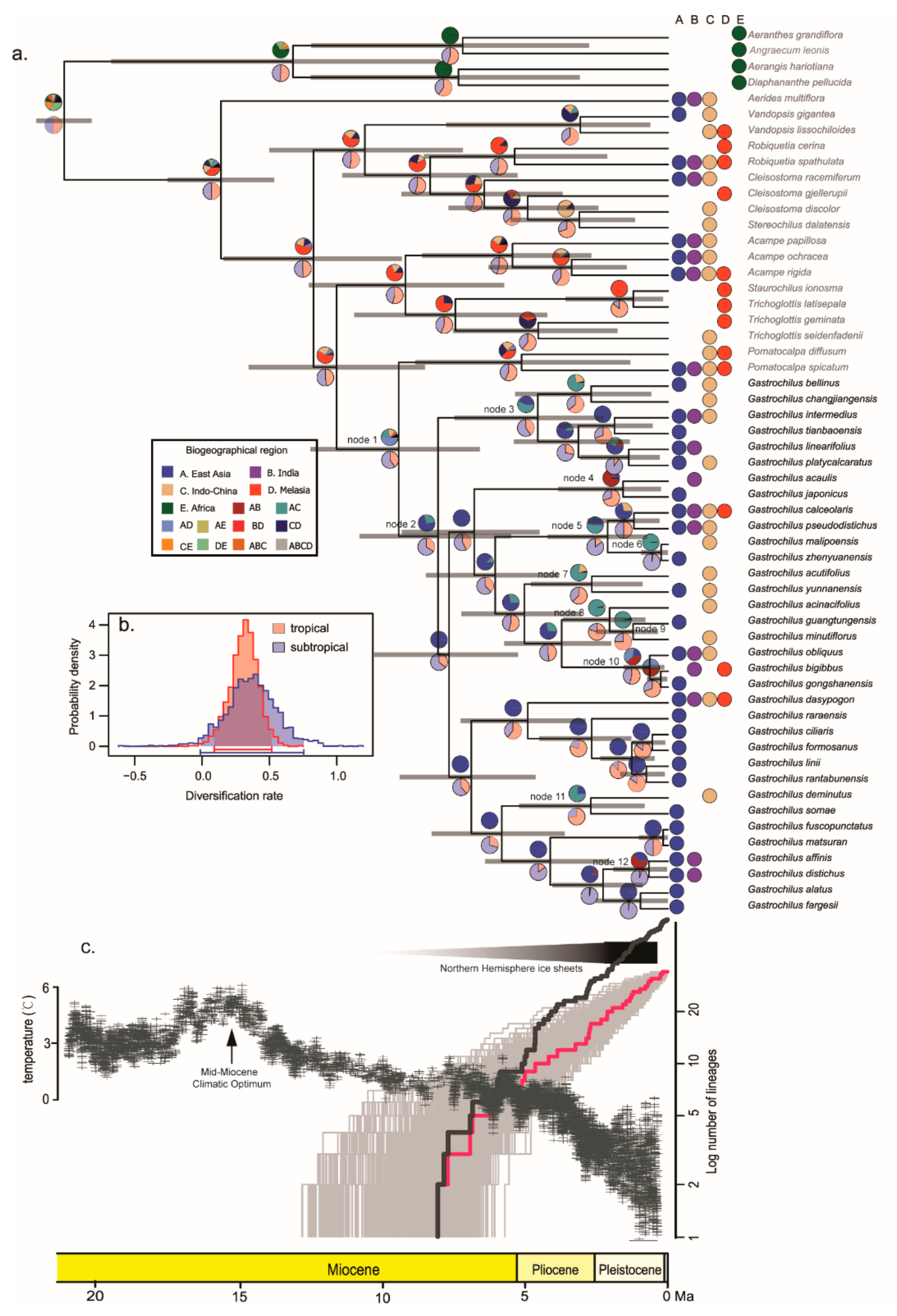
3.2. Ancestral Range Reconstruction
3.3. Diversification of Gastrochilus
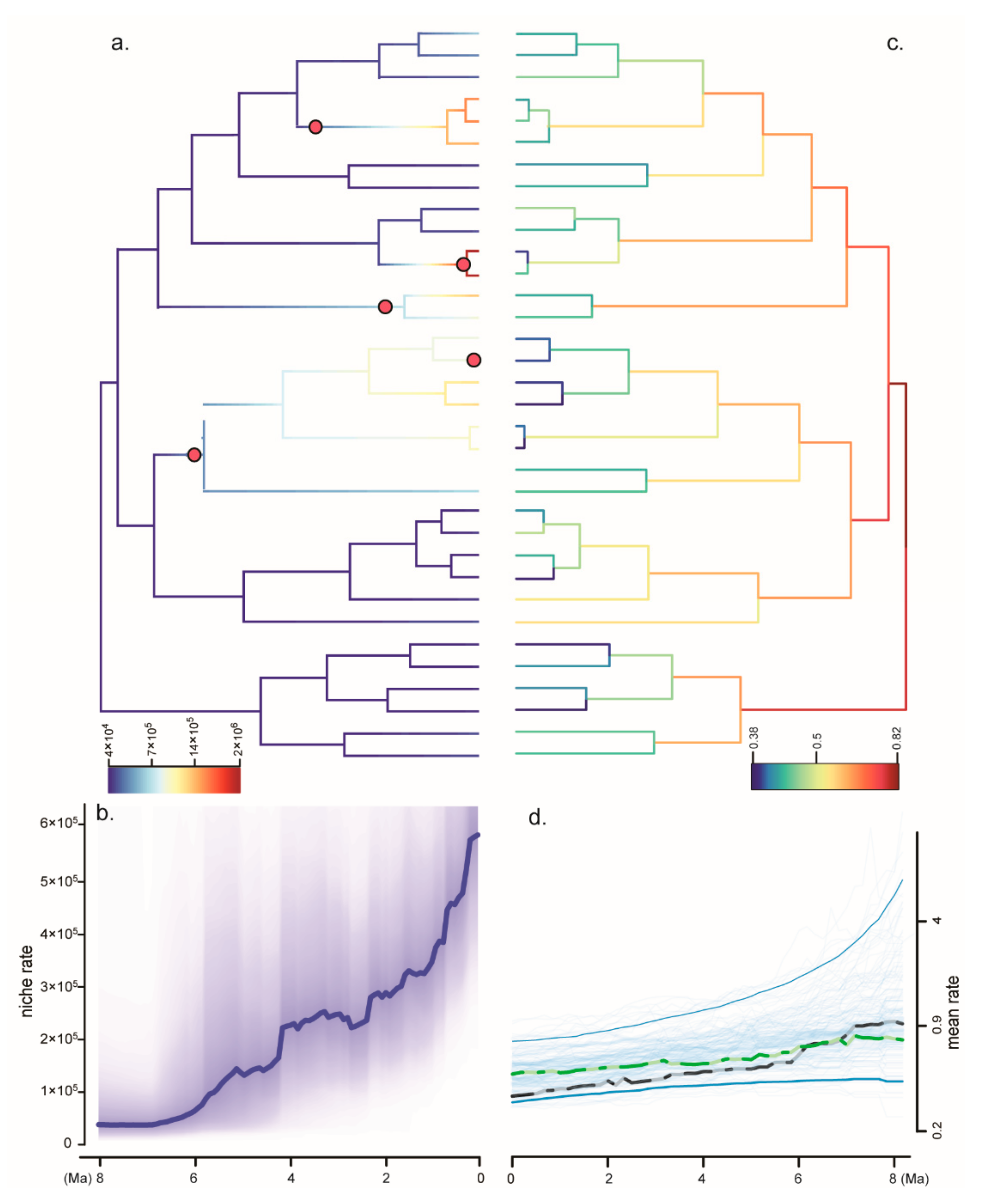
3.4. Niche Evolution and Phylogenetic Signals
4. Discussion
4.1. Temporal and Spatial Mode of Gastrochilus
4.2. Diversification and Niche Evolution of Gastrochilus
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Smith, S.A.; Donoghue, M.J. Combining historical biogeography with niche modeling in the Caprifolium clade of Lonicera (Caprifoliaceae, Dipsacales). Syst. Biol. 2010, 59, 322–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiens, J.; Donoghue, M.J. Historical biogeography, ecology and species richness. Trends Ecol. Evol. 2004, 19, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Donoghue, M.J.; Edwards, E.J. Biome shifts and niche evolution in plants. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 547–572. [Google Scholar] [CrossRef] [Green Version]
- Wiens, J.; Ackerly, D.D.; Allen, A.P.; Anacker, B.L.; Buckley, L.B.; Cornell, H.V.; Damschen, E.I.; Davies, J.T.; Grytnes, A.; Harrison, S.P.; et al. Niche conservatism as an emerging principle in ecology and conservation biology. Ecol. Lett. 2010, 13, 1310–1324. [Google Scholar] [CrossRef]
- Couvreur, T.L.P.; Forest, F.; Baker, W.J. Origin and global diversification patterns of tropical rain forests: Inferences from a complete genus-level phylogeny of palms. BMC Biol. 2011, 9, 44. [Google Scholar] [CrossRef] [Green Version]
- Pyron, R.A.; Costa, G.C.; Patten, M.A.; Burbrink, F.T. Phylogenetic niche conservatism and the evolutionary basis of ecological speciation. Biol. Rev. 2015, 90, 1248–1262. [Google Scholar] [CrossRef]
- Wang, Q.; Wu, S.D.; Su, X.Y.; Zhang, L.J.; Xu, X.T.; Lyu, L.S.; Cai, H.Y.; Shrestha, N.; Liu, Y.P.; Wang, W.; et al. Niche conservatism and elevated diversification shape species diversity in drylands: Evidence from Zygophyllaceae. Proc. R. Soc. B 2018, 285, 20181742. [Google Scholar] [CrossRef] [Green Version]
- Gagnon, E.; Ringlberg, J.J.; Bruneau, A.; Bruneau, A.; Lewis, G.P.; Hughes, C. Global Succulent Biome phylogenetic conservatism across the pantropical Caesalpinia Group (Leguminosae). New Phytol. 2019, 222, 1994–2008. [Google Scholar] [CrossRef]
- Cardillo, M.; Weston, P.H.; Reynolds, Z.K.M.; Olde, P.M.; Mast, A.R.; Lemmon, E.M.; Lemmon, A.R.; Bromham, L. The phylogeny and biogeography of Hakea (Proteaceae) revealed the role of biome shifts in a continental plant radiation. Evolution 2017, 71, 1928–1943. [Google Scholar] [CrossRef]
- Crisp, M.D.; Arroyo, M.T.K.; Cook, L.G.; Gandolfo, M.A.; Jordan, G.J.; McGlone, M.S.; Weston, P.H.; Westoby, M.; Wilf, P.; Linder, H.P. Phylogenetic biome conservation on a global scale. Nature 2009, 458, 754–756. [Google Scholar] [CrossRef]
- Rice, A.; Smarda, P.; Novosolov, M. The global biogeography of polyploid plants. Nat. Ecol. Evol. 2019, 3, 265–273. [Google Scholar] [CrossRef]
- Losos, J.B. Phylogenetic niche conservatism, phylogenetic signal and the relationship between phylogenetic related and ecological similarity among species. Ecol. Lett. 2018, 11, 995–1107. [Google Scholar] [CrossRef]
- Liu, H.C.; Jacquemyn, H.; Chen, W.; Janssens, S.B.; He, X.Y.; Yu, S.; Huang, Y.Q. Niche evolution and historical biogeography of lady slipper orchids in North America and Eurasia. J. Biogeogr. 2020, 48, 2727–2741. [Google Scholar] [CrossRef]
- Rice, N.H.; Martınez-Meyer, E.; Peterson, T. Ecological niche differentiation in the Aphelocoma jays: A phylogenetic perspective. Biol. J. Linn. Soc. 2003, 80, 369–383. [Google Scholar] [CrossRef] [Green Version]
- Pridgeon, A.; Cribb, P.; Chase, M.; Rasmussen, F.N. Genera Orchidacearum: Epidendroideae (Part Three); Oxford University Press: New York, NY, USA, 2014. [Google Scholar]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, S.P.; Hunter, S.J.; Zuluaga, A.; Iles, W.J.D.; Clements, M.A.; Arroyo, M.T.K.; Leebens, M.J.; et al. Orchid phylogenomics and multiple drivers of their extraordinary diversification. Proc. R. Soc. B 2015, 282, 20151553. [Google Scholar] [CrossRef]
- Wen, J. Evolution of eastern Asian and eastern North American disjunct distributions in flowering plants. Annu. Rev. Ecol. Syst. 1999, 30, 421–455. [Google Scholar] [CrossRef]
- Wu, Z.Y.; Wu, S. A proposal for a new floristic kingdom (realm)—The East Asiatic Kingdom, its delineation and characteristics. In Floristic Characteristics and Diversity of East Asian Plants; Zhang, A.L., Wu, S.G., Eds.; Higher Education Press: Beijing, China; Springer Verlag Press: Berlin/Heidelberg, Germany, 1998; pp. 1–42. [Google Scholar]
- Lu, L.M.; Mao, L.F.; Yang, T.; Ye, J.F.; Liu, B.; Li, H.L.; Sun, M.; Miller, J.T.; Mathews, S.; Hu, H.H.; et al. Evolutionary history of the angiosperm flora of China. Nature 2018, 554, 234–238. [Google Scholar] [CrossRef]
- Mittermeier, R.A.; Fonseca, P.; Gil, R.; Hoffmann, M.; Pilgrim, J.; Brooks, T.; Mittermeier, C.G.; Lamoreux, J.; Fonseca, A.B.G. Hotspots Revisited: Earth’s Biologically Richest and Most Endangered Terrestrial Ecoregions; Cermex: Mexico City, Mexico, 2005. [Google Scholar]
- An, Z.S.; Kutzbach, J.E.; Prell, W.L.; Porter, W.L. Evolution of Asian monsoons and phased uplift of the Himalaya-Tibetan plateau since late Miocene times. Nature 2001, 411, 62–66. [Google Scholar]
- Sun, X.J.; Wang, P.X. How old is the Asian monsoon system? Palaeobotanical records from China. Palaeogeogr. Palaeoclimat. Palaeoecol. 2005, 222, 181–222. [Google Scholar] [CrossRef]
- Manish, K.; Pandit, M.K. Geophysical upheavals and evolutionary diversification of plant species in the Himalaya. Peer J. 2018, 6, e5919. [Google Scholar] [CrossRef]
- Pusok, A.E.; Stegman, D.R. The convergence history of India-Eurasia records multiple subduction dynamics processes. Sci. Adv. 2020, 6, eaaz8681. [Google Scholar] [CrossRef] [PubMed]
- Xing, Y.; Ree, R.H. Uplift-driven diversification in the Hengduan Mountains, a temperate biodiversity hotspot. Proc. Natl. Acad. Sci. USA 2017, 114, E3444–E3451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- An, Z.S. Late Cenozoic Climate Change in Asia: Loess, Monsoon and Monsoon-Arid Environment Evolution; Springer: Dordrecht, The Netherlands, 2014. [Google Scholar]
- Ashokan, A.; Xavier, A.; Suksathan, P.; Ardiyani, M.; Leong-Škorničková, J.; Newman, M.; Kress, W.J.; Gowda, V. Himalayan orogeny and monsoon intensification explain species diversification in an endemic ginger (Hedychium: Zingiberaceae) from the Indo-Malayan Realm. Mol. Phylogenet. Evol. 2022, 170, 107440. [Google Scholar] [CrossRef] [PubMed]
- Xiang, X.G.; Mi, X.C.; Zhou, H.L.; Li, J.W.; Chung, S.W.; Li, D.Z.; Huang, W.C.; Jin, W.T.; Li, Z.Y.; Huang, L.Q.; et al. Biogeogrpahical diversification of mainland Asian Dendrobium (Orchidaceae) and its implications for the historical dynamics of evergreen broad-leaved forests. J. Biogeogr. 2016, 43, 1310–1323. [Google Scholar] [CrossRef]
- Yu, X.Q.; Gao, L.M.; Soltis, D.E.; Soltis, P.S.; Yang, J.B.; Fang, L.; Yang, S.X.; Li, D.Z. Insights into the historical assembly of east Asian subtropical evergreen broadleaved forests revealed by the temporal history of the tea family. New Phytol. 2017, 215, 1235–1248. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.H.; Xiang, K.L.; Lian, L.; Peng, H.W.; Erst, S.A.; Xiang, X.G.; Chen, Z.D.; Wang, W. Biogeographic diversification of Mahonia (Berberidaceae): Implications for the origin and evolution of East Asian subtropical evergreen broadleaved forests. Mol. Phylogenet. Evol. 2020, 151, 106910. [Google Scholar] [CrossRef]
- Li, X.Q.; Xiang, X.G.; Zhang, Q.; Jabbour, F.; Ortiz, R.D.C.; Erst, A.S.; Li, Z.Y.; Wang, W. Immigration dynamics of tropical and subtropical Southeast Asian limestone karst floras. Proc. R. Soc. B 2022, 289, 20211308. [Google Scholar] [CrossRef]
- Tsi, Z.H. A preliminary revision of Gastrochilus (Orchidaceae). Guihaia 1996, 16, 123–154. [Google Scholar]
- Yi, S.R.; Huang, Y.; Xiao, B.; Shen, M.L.; Li, J. Gastrochilus brevifimbriatus (Orchidaceae, Aeridinae), a New Species from Chongqing, China. Novon 2010, 20, 113–116. [Google Scholar]
- Kumar, P.; Gale, S.W.; Kocyan, A.; Fischer, G.A.; Averyanov, L.V.; Borosova, R.; Bhattacharjee, A.; Li, J.H.; Pang, K.S. Gastrochilus kadooriei (Orchidaceae), a new species from Hong Kong, with notes on allied taxa in section Microphyllae found in the region. Phytotaxa 2014, 164, 91–103. [Google Scholar] [CrossRef] [Green Version]
- Raskoti, B.B. A new species of Gastrochilus and new records for the orchids of Nepal. Phytotaxa 2015, 233, 179. [Google Scholar] [CrossRef]
- Averyanov, L.V.; Van, C.N.; Truong, V.B.; Maisak, T.; Luu, H.T.; Nguyen, K.S.; Diep, D.Q.; Hoang, T.; Tuan, N.H.; Chu, X.C.; et al. New orchids (Orchidaceae: Cymbidieae and Vandeae) in the Flora of Vietnam. Taiwania 2018, 63, 119–138. [Google Scholar]
- Hsu, T.C.; Hsieh, S.I.; Wu, J.H.; Hung, H.C. Gastrochilus deltoglossus (Orchidaceae: Epidendroideae: Vandeae: Aeridinae), a new species from Taiwan. Taiwania 2018, 63, 360–365. [Google Scholar]
- Rao, W.H.; Liu, Z.J.; Zhang, G.Q.; Chen, X.H.; Huang, J.; Chen, G.Z.; Chen, L.J. A new epiphytic species of Gastrochilus (Orchidaceae: Epidendroideae) from Yunnan, China. Phytotaxa 2019, 413, 296–300. [Google Scholar]
- Wu, X.F.; Ye, D.P.; Pan, B.; Lin, X.Q.; Jiang, H.; Liu, Q. Validation of Gastrochilus prionophyllus (Vandeae, Orchidaceae), a new species from Yunnan province, China. PhytoKeys 2019, 130, 161–169. [Google Scholar] [CrossRef]
- Liu, Q.; Tan, Y.H.; Gao, J.Y. A new species of Gastrochilus (Aeridinae, Vandeae, Orchidaceae) and a new record species from Yunnan, China. Phytotaxa 2016, 282, 66–70. [Google Scholar] [CrossRef]
- Liu, Q.; Gao, J.Y. Gastrochilus dulongjiangensis (Aeridinae, Vandeae, Epidendroideae, Orchidaceae), a new species from Yunnan province, China. Phytotaxa 2018, 340, 293–296. [Google Scholar] [CrossRef]
- Liu, Q.; Kumar, P.; Gao, J.Y. Notes on Gastrochilus gongshanensis (Orchidaceae). Kew Bull. 2019, 74, 21. [Google Scholar] [CrossRef]
- Liu, Q.; Zhou, S.S.; Li, R.; Tan, Y.H.; Zyaw, M.; Xing, X.K.; Gao, J.Y. Notes on the genus Gastrochilus (Orchidaceae) in Myanmar. PhytoKeys 2020, 138, 113–123. [Google Scholar] [CrossRef]
- Ormerod, P.; Kumar, C.S. A new Bhutanese Gastrochilus (Orchidaceae), and another transfer in Indian Cylindrolobus. Harv. Pap. Bot. 2020, 25, 125–127. [Google Scholar] [CrossRef]
- Li, J.W.; Ya, J.D.; Ye, D.P.; Liu, C.; Liu, Q.; Pan, R.; He, Z.X.; Pan, B.; Cai, J.; Lin, D.L.; et al. Taxonomy notes on Vandeae (Orchidaceae) from China: Five new species and two new records. Plant Diver. 2021, 43, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.J. Gastrochilus odoratus (Kudô) J.J. Smith. Bull. Jard. Bot. Buitenzorg 1937, 14, 168. [Google Scholar]
- Chase, M.W.; Cameron, K.M.; Freudenstein, J.V.; Pridgeon, A.M.; Salazar, G.; Berg, C.V.D.; Schuiteman, A. An updated classification of Orchidaceae. Bot. J. Linn. Soc. 2015, 177, 151–174. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.C.; Tsi, Z.H.; Wood, J.J. Gastrochilus D. Don. In Flora of China Vol. 25 (Orchidaceae); Wu, Z.Y., Raven, P.H., Hong, D.Y., Eds.; Science Press: Beijing, China; Missouri Botanical Garden Press: St. Louis, MI, USA, 2009; pp. 491–498. [Google Scholar]
- Liu, Q.; Song, Y.; Jin, X.H.; Gao, J.Y. Phylogenetic relationships of Gastrochilus (Orchidaceae) based on nuclear and plastid DNA data. Bot. J. Linn. Soc. 2019, 189, 228–243. [Google Scholar] [CrossRef]
- Liu, D.K.; Tu, X.D.; Zhao, Z.; Zeng, M.Y.; Zhang, S.; Ma, L.; Zhang, G.Q.; Wang, M.M.; Liu, Z.J.; Lan, S.R.; et al. Plastid phylogenomic data yield new and robust insights into the phylogeny of Cleisostoma-Gastrochilus clades (Orchidaceae, Aeridinae). Mol. Phylogenet. Evol. 2020, 145, 10729. [Google Scholar] [CrossRef] [PubMed]
- Van den Berg, C.; Goldman, D.H.; Freudenstein, J.V.; Pridgeon, A.M.; Cameron, K.M.; Chase, M.W. An overview of the phylogenetic relationships within Epidendroideae inferred from multiple DNA regions and circumscription of Epidendreae and Arethuseae (Orchidaceae). Am. J. Bot. 2005, 92, 613–645. [Google Scholar] [CrossRef]
- Farminhão, J.N.M.; Verlynde, S.; Kaymak, E.; Droissart, V.; Simo-Droissart, M.; Collobert, G.; Martos, F.; Stévart, T. Rapid radiation of angraecoids (Orchidaceae, Angraecinae) in tropical Africa characterised by multiple karyotypic shifts under major environmental instability. Mol. Phylogenet. Evol. 2021, 159, 107105. [Google Scholar] [CrossRef]
- Shaw, J.; Lickey, E.B.; Beck, J.T.; Farmer, S.B.; Liu, W.S.; Miller, J.; Siripun, K.C.; Winder, C.T.; Schilling, E.E.; Small, R.L. The tortoise and the hare II: Relative utility of 21 noncoding chloroplast DNA sequences for phylogenetic analysis. Am. J. Bot. 2005, 92, 142–166. [Google Scholar] [CrossRef] [Green Version]
- Shaw, J.; Lickey, E.B.; Schilling, E.E.; Small, R.L. Comparison of whole chloroplast genome sequences to choose noncoding regions for phylogenetic studies in angiosperms: The tortoise and the hare III. Am. J. Bot. 2007, 94, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl. Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Farris, J.S.; Källersjö, M.; Kluge, A.G.; Bult, C. Testing significance of incongruence. Cladistics 1994, 10, 315–319. [Google Scholar] [CrossRef]
- Wendel, J.; Doyle, J. Phylogenetic incongruence, window into genome history and evolution. In Molecular Systematics of Plants II: DNA Sequencing; Soltis, D.E., Soltis, P.S., Doyle, J.J., Eds.; Kluwer Academic Publishing: Dordrecht, The Netherlands, 1998; pp. 265–296. [Google Scholar]
- Gatesy, J.; O’Grady, P.; Baker, R.H. Corroboration among data sets in simultaneous analysis, hidden support for phylogenetic relationships among higher level Artiodactyl taxa. Cladistics 1999, 15, 271–313. [Google Scholar] [CrossRef]
- Swofford, D.L. PAUP 4.0b10: Phylogenetic analysis using parsimony (and other methods); Sinauer: Sunderland, UK, 2002. [Google Scholar]
- Posada, D.; Crandall, K.A. Modeltest: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J. Tracer v1.5 [online]. Available online: http://beast.bio.ed.ac.uk/Tracer (accessed on 30 November 2009).
- Felsenstein, J. Phylogenies from molecular sequences: Inference and reliability. Ann. Rev. Genet. 1988, 22, 521–565. [Google Scholar] [CrossRef]
- Sorhannus, U.; Bell, C.V. Testing for equality of molecular evolutionary rates: A comparison between a relative-rate test and a likelihood ratio test. Mol. Biol. Evol. 1999, 16, 849–855. [Google Scholar] [CrossRef] [Green Version]
- Bouckaert, R.; Heled, J.; Kühnert, D.; Vaughan, T.; Wu, C.H.; Xie, D.; Suchard, M.A.; Rambaut, A.; Drummond, A.J. BEAST 2: A Software platform for Bayesian evolutionary analysis. PLoS Comput. Biol. 2014, 10, e1003537. [Google Scholar] [CrossRef] [Green Version]
- Givnish, T.J.; Spalink, D.; Ames, M.; Lyon, A.P.; Hunter, S.J.; Zuluaga, A.; Doucette, A.A.; Caro, G.G.; McDaniel, J.; Clements, M.A.; et al. Orchid historical biogeography, diversification, Antarctica and the paradox of orchid dispersal. J. Biogeogr. 2016, 43, 1905–1916. [Google Scholar] [CrossRef]
- Ho, S.Y.W. Calibrating molecular estimates of substitution rates and divergence times in bird. J. Avian Biol. 2007, 38, 409–414. [Google Scholar] [CrossRef]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [Green Version]
- Takhtajan, A. Floristic Regions of the World; University of California Press: Berkeley, CA, USA; Los Angeles, CA, USA, 1986. [Google Scholar]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X.J. RASP (Reconstruct Ancestral State in Phylogenies): A tool for historical biogeography. Mol. Phylogenet. Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef]
- Pagel, M.; Meade, A. BayesTraits: Computer Program and Documentation Version 4.0. 2022. Available online: http://www.evolution.reading.ac.uk/BayesTraitsV4.0.0.html (accessed on 1 February 2022).
- FitzJohn, R.G. Diversitree: Comparative phylogenetic analyses of diversification in R. Mol. Phylogenet. Evol. 2012, 3, 1084–1092. [Google Scholar] [CrossRef]
- Rabosky, D.L. LASER: A maximum likelihood toolkit for detecting temporal shits in diversification rates from molecular phylogenies. Evol. Bioinform. Online 2006, 2, 257–260. [Google Scholar] [CrossRef] [Green Version]
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maliet, O.; Morlon, H. Fast and accurate estimation of species-specific diversification rates using data augmentation. Syst. Biol. 2022, 71, 353–366. [Google Scholar] [CrossRef] [PubMed]
- Revell, L.J. Phytools: An R package for phylogenetic comparative biology (and other things). Mol. Phylogenet. Evol. 2012, 3, 217–223. [Google Scholar] [CrossRef]
- Fick, S.E.; Hijmans, R.J. WorldClim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 2017, 37, 4302–4315. [Google Scholar] [CrossRef]
- Rabosky, D.L.; Grundler, M.; Anderson, C.; Title, P.; Shi, J.J.; Brown, J.W.; Huang, H.; Larson, J.G.; Kembel, S. BAMMtools: An R package for the analysis of evolutionary dynamics on phylogenetic trees. Mol. Phylogenet. Evol. 2014, 5, 701–707. [Google Scholar] [CrossRef]
- Plummer, M.; Best, N.; Cowles, K.; Vines, K. CODA: Convergence diagnosis and output analysis for MCMC. R News 2006, 6, 7–11. [Google Scholar]
- Blomberg, S.P.; Garland, T.; Ives, A.R. Testing for phylogenetic signal in comparative data: Behavioral traits are more labile. Evolution 2003, 57, 717–745. [Google Scholar] [CrossRef]
- Pagel, M. Inferring the historical patterns of biological evolution. Nature 1999, 401, 877–884. [Google Scholar] [CrossRef]
- Zachos, J.C.; Dickens, G.R.; Zeebe, R.E. An early Cenozoic perspective on greenhouse warming and carbon-cycle dynamics. Nature 2008, 451, 279–283. [Google Scholar] [CrossRef]
- Zhang, J.B.; Li, R.Q.; Xiang, X.G.; Manchester, S.R.; Lin, L.; Wang, W.; Wen, J.; Chen, Z.D. Integrated fossil and molecular data reveal the biogeographic diversification of the eastern Asian-eastern North American disjunct hickory genus (Carya Nutt.). PLoS ONE 2013, 8, e70449. [Google Scholar] [CrossRef] [Green Version]
- Kou, Y.X.; Cheng, S.M.; Tian, S.; Li, B.; Fan, D.M.; Chen, Y.J.; Soltis, D.E.; Soltis, P.S.; Zhang, Z.Y. The antiquity of Cyclocarya paliurus (Juglandaceae) provides new insights into the evolution of relict plants in subtropical China since the late Tertiary Miocene. J. Biogeogr. 2016, 43, 351–360. [Google Scholar] [CrossRef]
- Ye, X.Y.; Ma, P.F.; Yang, G.Q.; Guo, C.; Zhang, Y.X.; Chen, Y.M.; Guo, Z.H.; Li, D.Z. Rapid diversification of alpine bamboos associated with the uplift of the Hengduan Mountains. J. Biogeogr. 2019, 46, 2678–2689. [Google Scholar] [CrossRef]
- Filipelli, G.M. Intensification of the Asian monsoon and a chemical weathering event in the late Miocene-early Pliocene: Implications for late Neogene climate change. Geology 1997, 25, 27–30. [Google Scholar] [CrossRef]
- Osborne, C.P. Atmosphere, ecology and evolution: What drove the Miocene expansion of C4 grasslands? J. Ecol. 2008, 96, 35–45. [Google Scholar] [CrossRef] [Green Version]
- Ding, W.N.; Ree, R.H.; Spicer, R.A.; Xing, Y.W. Ancient orogenic and monsoon-driven assembly of the world’s richest temperate alpine flora. Science 2020, 369, 578–581. [Google Scholar] [CrossRef]
- Spicer, R.A.; Yang, J.; Herman, A.B.; Kodrul, T.; Aleksandrova, G.; Maslova, N.; Spicer, T.E.V.; Ding, L.; Xu, Q.; Shukla, A.; et al. Paleogene monsoons across India and South China: Drivers of biotic change. Gondwana Res. 2017, 49, 350–363. [Google Scholar] [CrossRef] [Green Version]
- Chang, Z.; Xiao, J.; Lu, L.; Yao, H. Abrupt shifts in the Indian monsoon during the Pliocene marked by high-resolution terrestrial records from the Yuanmou Basin in southwest China. J. Asian Earth Sci. 2010, 37, 166–175. [Google Scholar] [CrossRef]
- Willeit, M.; Ganopolski, A.; Calov, R.; Brovkin, V. Mid-Pleistocene transition in glacial cycles explained by declining CO2 and regolith removal. Sci. Adv. 2019, 5, eaav7337. [Google Scholar] [CrossRef] [Green Version]
- Hu, A.Q.; Gale, S.W.; Liu, Z.J.; Fischer, G.A.; Saunders, R.M.K. Diversification slowdown in the Cirrhopetalum alliance (Bulbophyllum, Orchidaceae): Insights from the evolutionary dynamics of crassulacean acid metabolism. Front. Plant Sci. 2022, 13, 794171. [Google Scholar] [CrossRef]
- Gamisch, A.; Comes, H.P. Clade-age-dependent diversification under high species turnover shapes species richness disparities among tropical rainforest lineages of Bulbophyllum (Orchidaceae). BMC Evol. Biol. 2019, 19, 93. [Google Scholar] [CrossRef] [Green Version]
- Takamiya, T.; Wongsawad, P.; Sathapattayanon, A.; Tajima, N.; Suzuki, S.; Kitamura, S.; Shioda, N.; Handa, T.; Kitanaka, S.; Lijima, H.; et al. Molecular phylogenetics and character evolution of morphologically diverse groups, Dendrobium section Dendrobium and allies. AoB Plants 2014, 6, plu045. [Google Scholar] [CrossRef] [Green Version]
- Xiang, X.G.; Li, D.Z.; Jin, X.H.; Hu, H.; Zhou, H.L.; Jin, W.T.; Lai, Y.J. Monophyly or paraphyly– the taxonomy of Holcoglossum (Aeridinae: Orchidaceae). PLoS ONE 2012, 7, e52050. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Jin, W.; Zhang, L.; Zhou, P.; Luo, Y.; Zhu, Z.; Xiang, X. Biogeography and Diversification of the Tropical and Subtropical Asian Genus Gastrochilus (Orchidaceae, Aeridinae). Diversity 2022, 14, 396. https://doi.org/10.3390/d14050396
Li Y, Jin W, Zhang L, Zhou P, Luo Y, Zhu Z, Xiang X. Biogeography and Diversification of the Tropical and Subtropical Asian Genus Gastrochilus (Orchidaceae, Aeridinae). Diversity. 2022; 14(5):396. https://doi.org/10.3390/d14050396
Chicago/Turabian StyleLi, Yang, Weitao Jin, Liguo Zhang, Peng Zhou, Yan Luo, Ziwei Zhu, and Xiaoguo Xiang. 2022. "Biogeography and Diversification of the Tropical and Subtropical Asian Genus Gastrochilus (Orchidaceae, Aeridinae)" Diversity 14, no. 5: 396. https://doi.org/10.3390/d14050396
APA StyleLi, Y., Jin, W., Zhang, L., Zhou, P., Luo, Y., Zhu, Z., & Xiang, X. (2022). Biogeography and Diversification of the Tropical and Subtropical Asian Genus Gastrochilus (Orchidaceae, Aeridinae). Diversity, 14(5), 396. https://doi.org/10.3390/d14050396