
Phylogenetic, Microbiome, and Diet Characterisation of Wall Lizards in the Columbretes Archipelago (Spain): Clues for Their Conservation

 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Phylogenetic Analyses
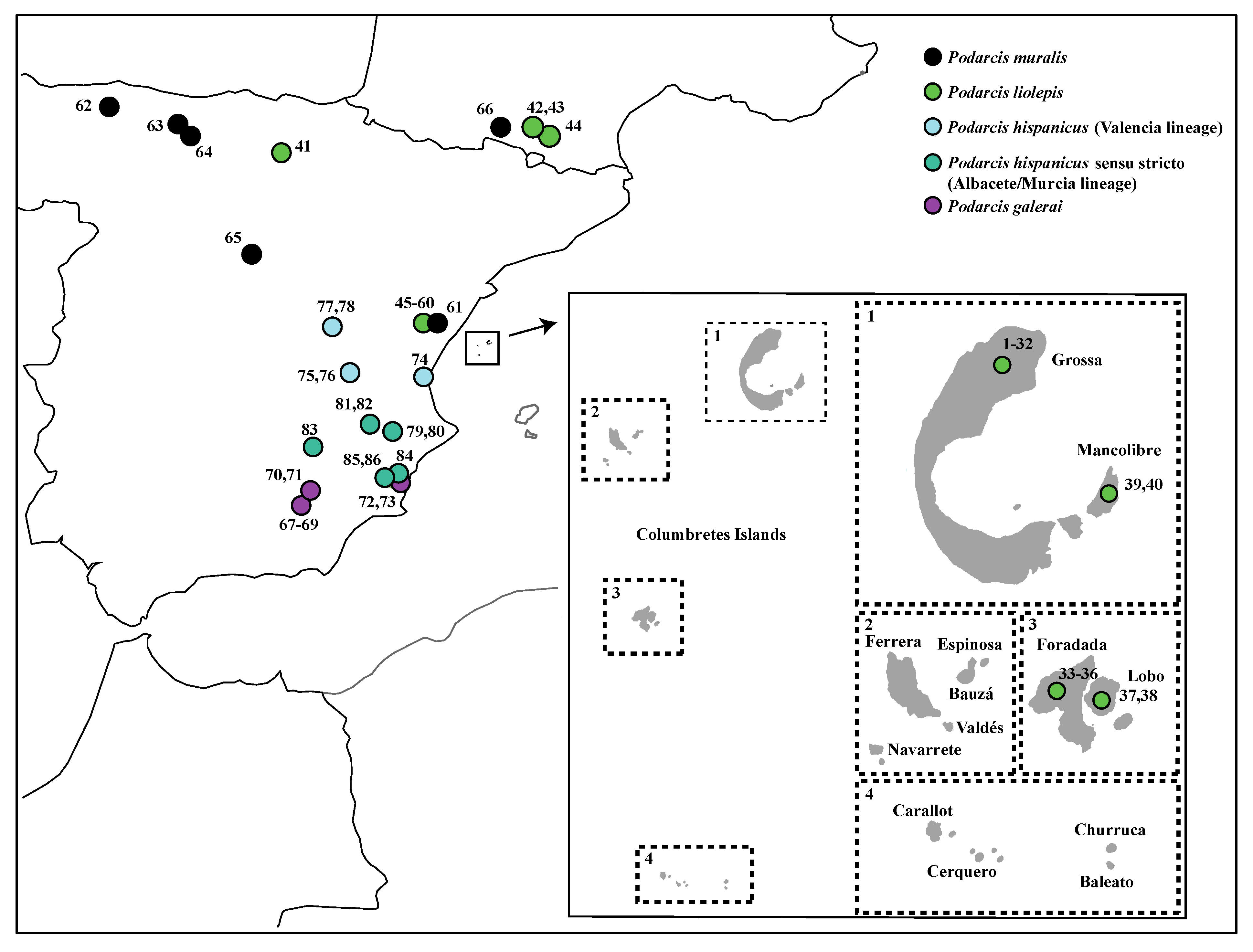
2.1.1. Sampling
2.1.2. DNA Amplification and Sequencing
2.1.3. Divergence and Phylogenetic Analyses
2.2. Metabarcoding Faecal Microbiota and Diet Associations
2.2.1. Sampling
2.2.2. DNA Extraction, DNA-Library Preparation, and Sequencing
2.2.3. Bioinformatic Analyses
3. Results
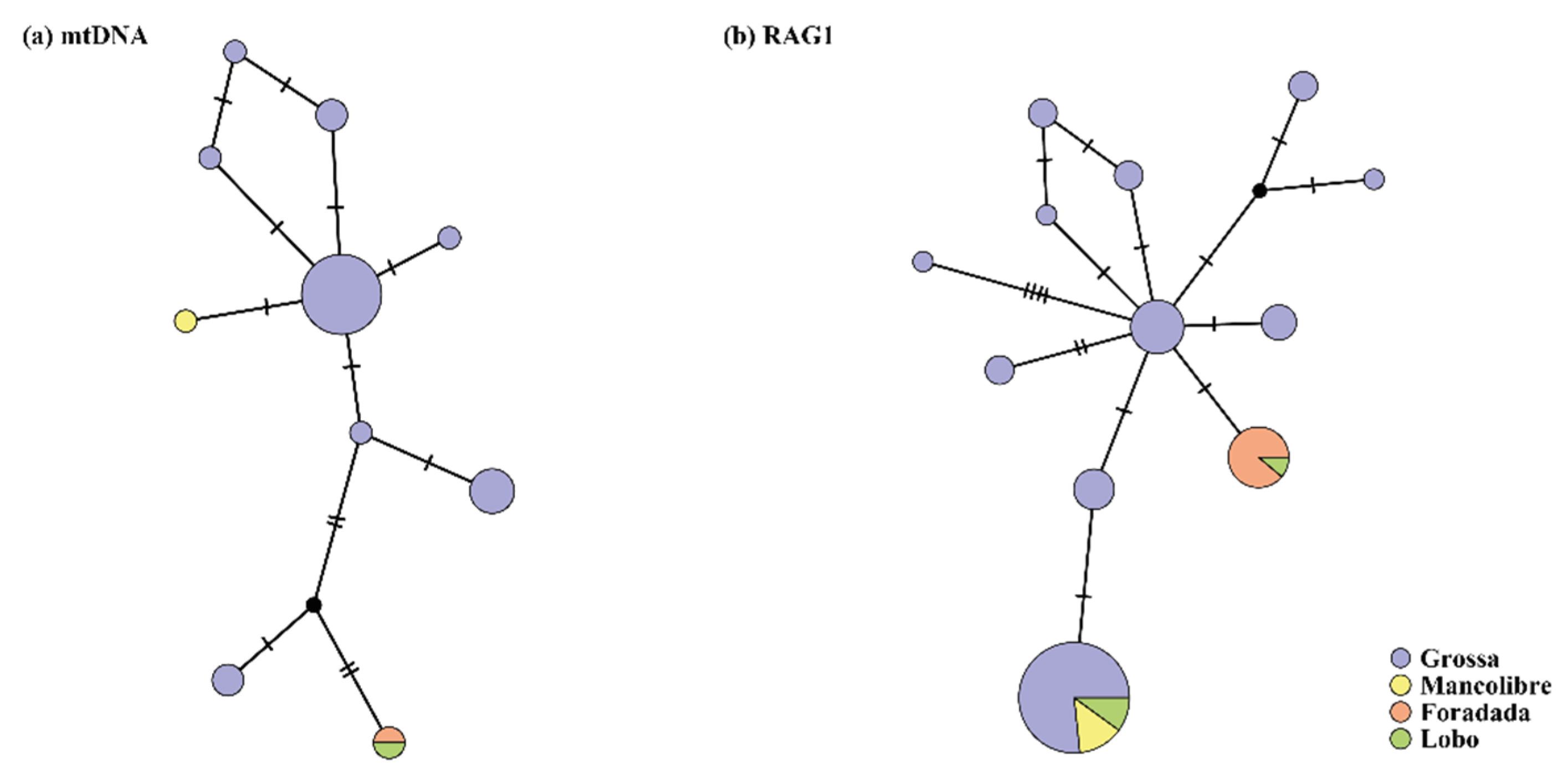
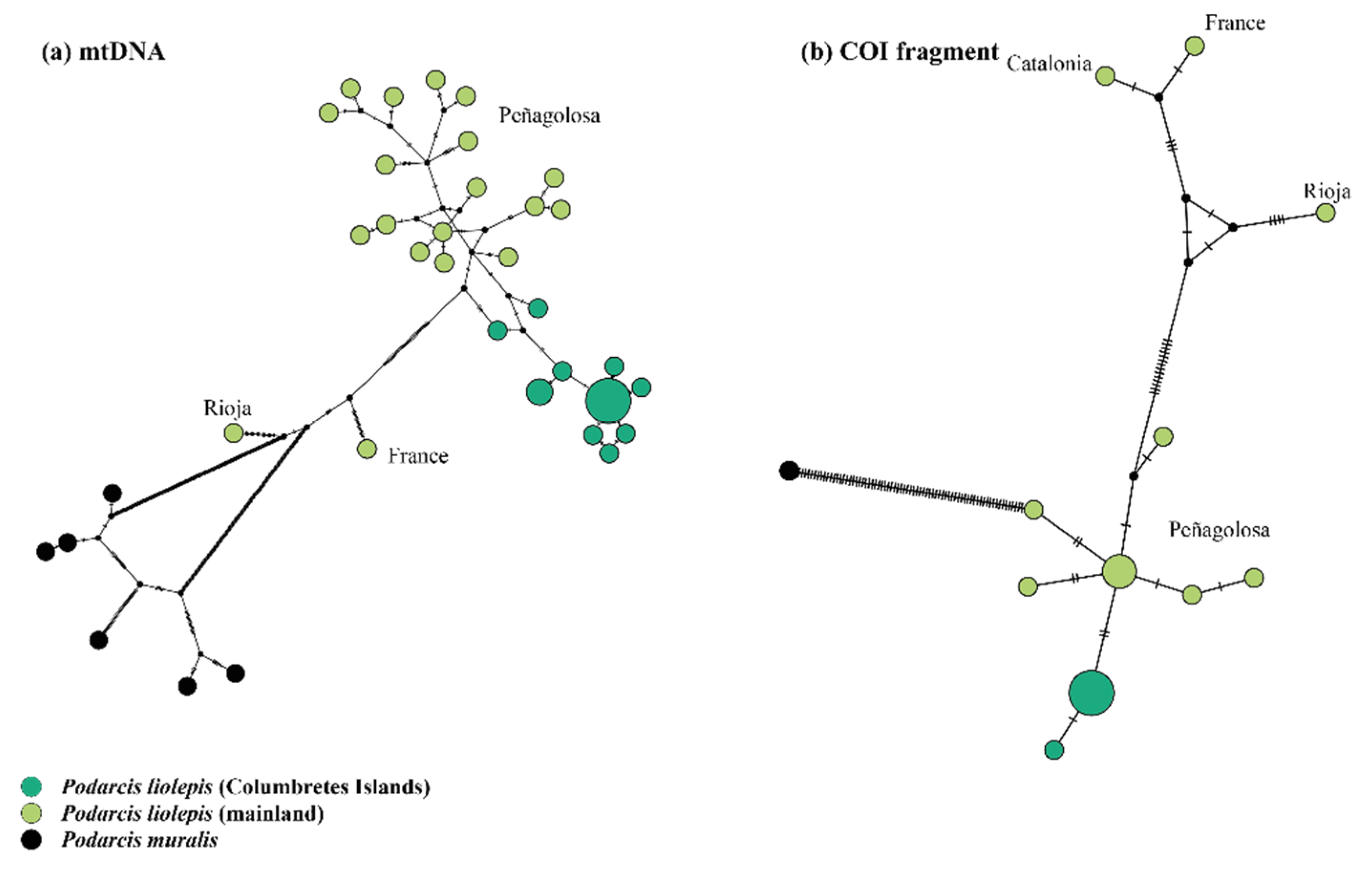
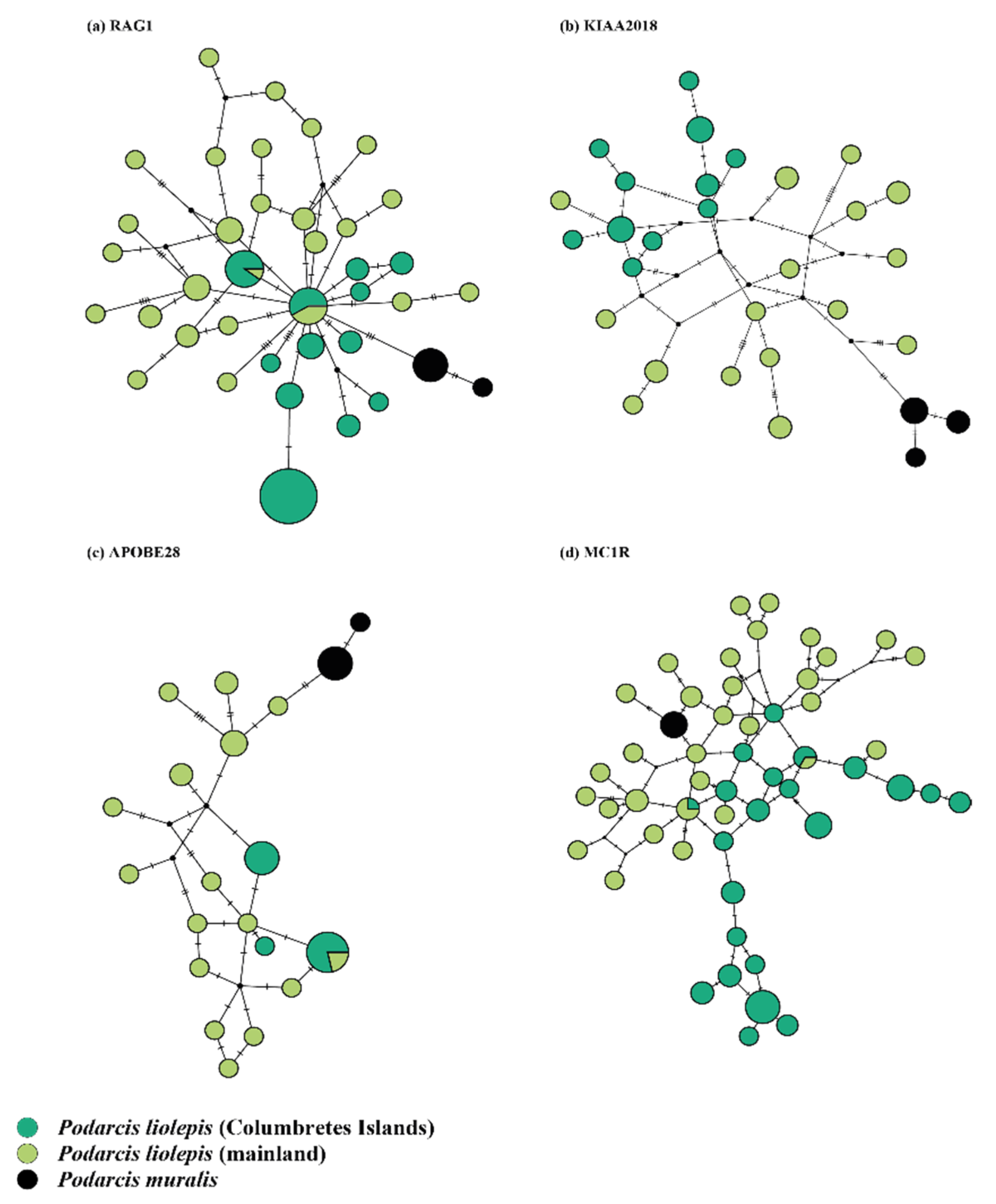
3.1. Genetic Diversity of Columbretes Islands’ Podarcis Based on Mitochondrial and Nuclear Genes
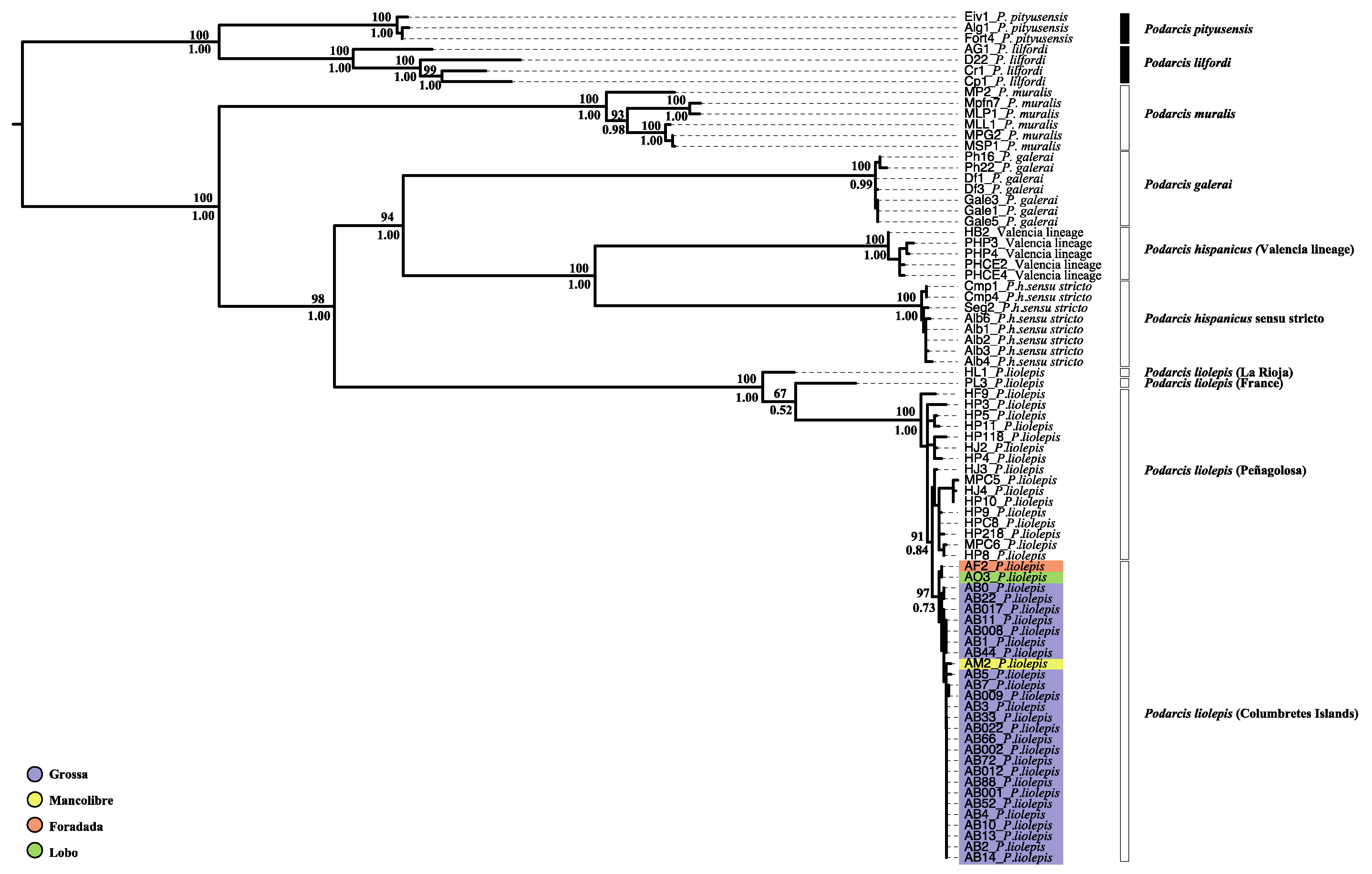
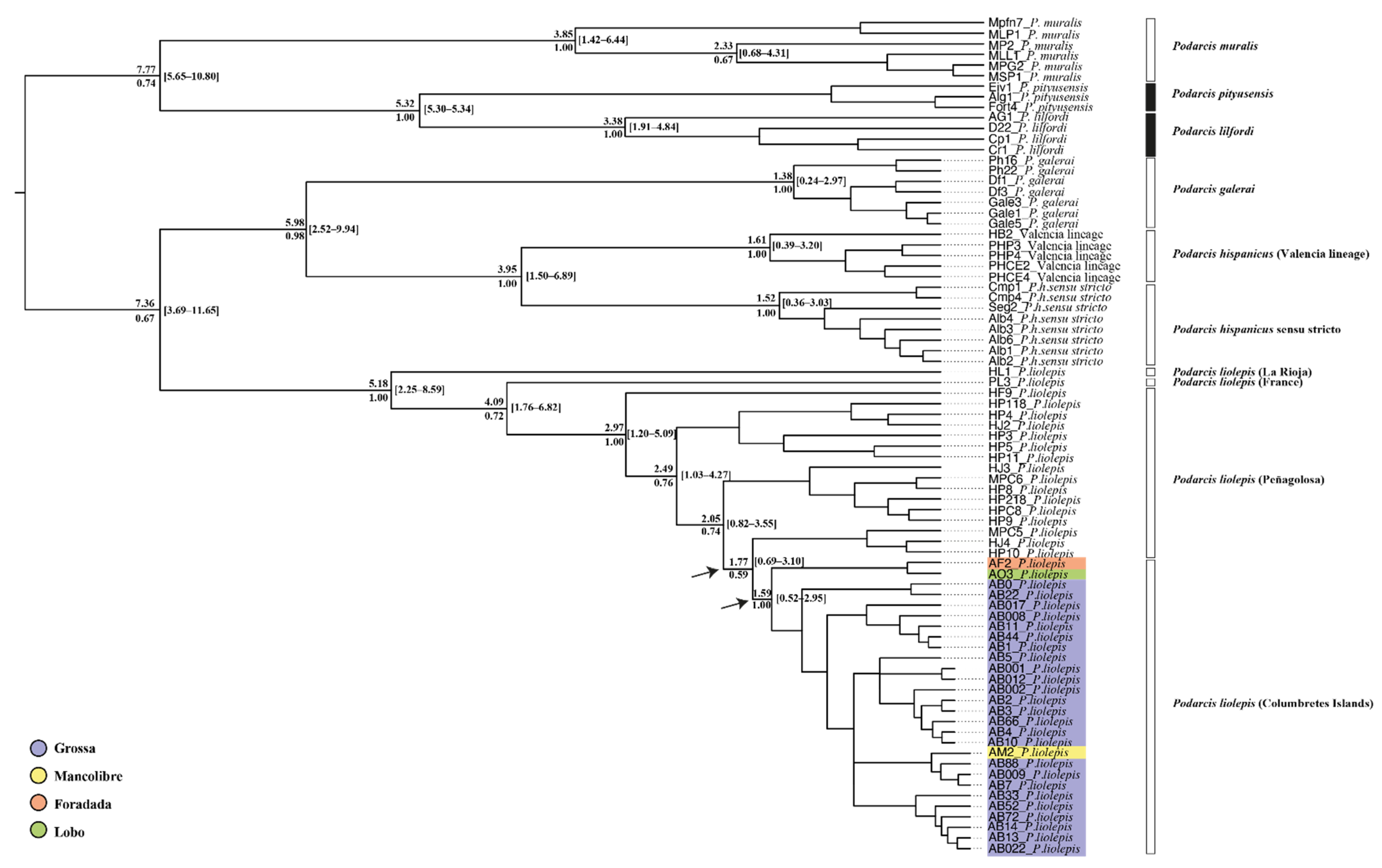
3.2. Phylogenetic Relationships and Possible Origin of Columbretes Islands Wall Lizard
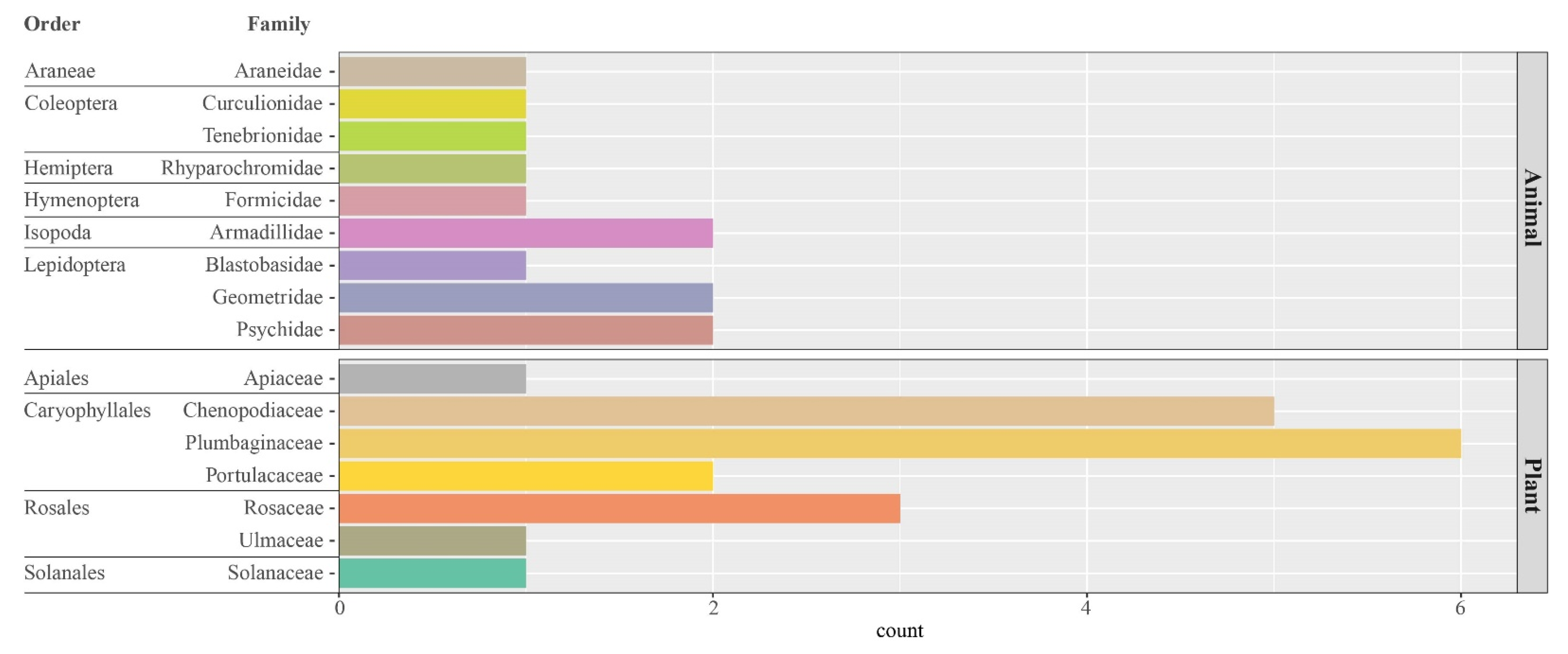
3.3. Molecular Diet Composition
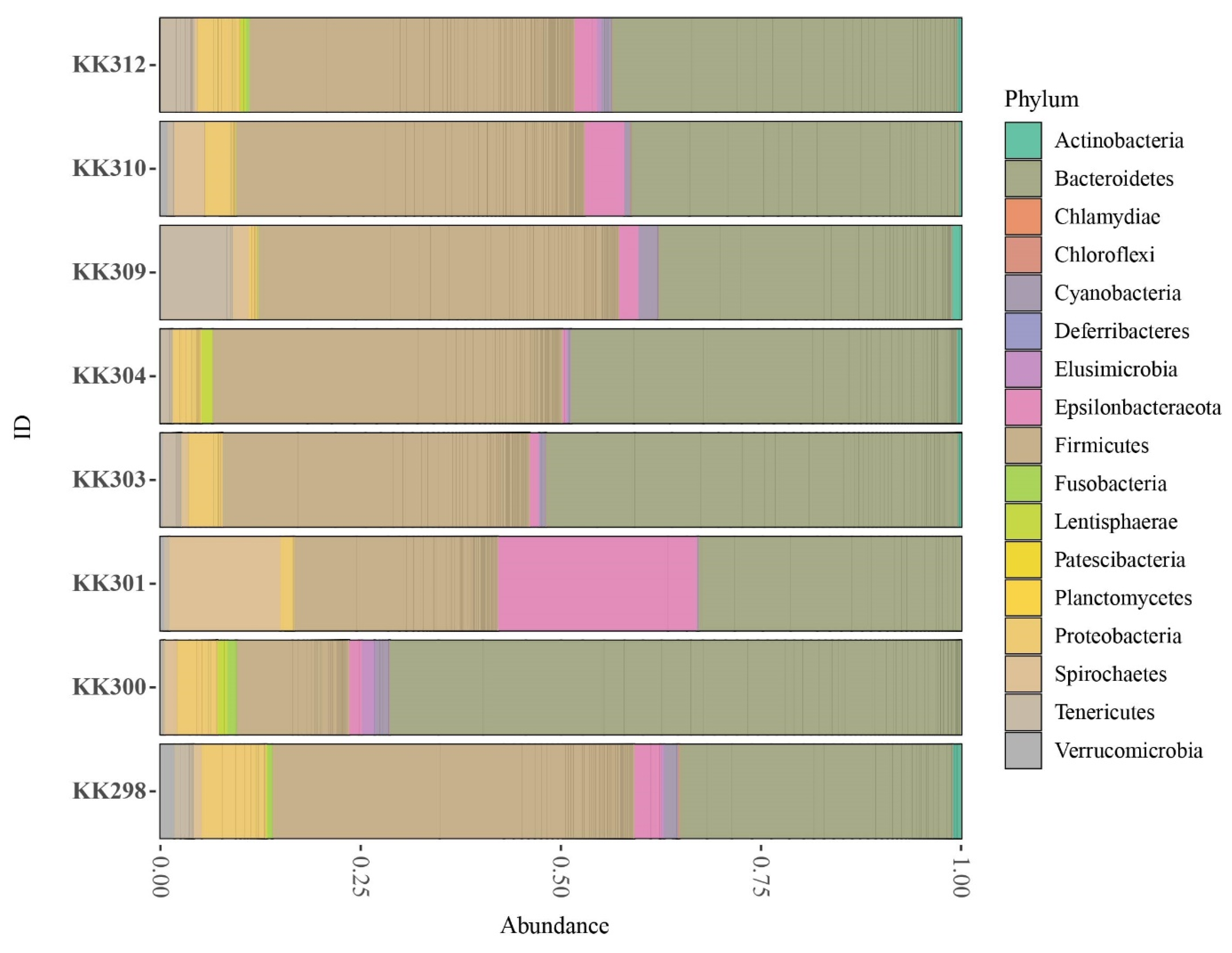
3.4. Microbiota Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Savelli, C. Time–space distribution of magmatic activity in the western Mediterranean and peripheral orogens during the past 30 Ma (a stimulus to geodynamic considerations). J. Geodyn. 2002, 34, 99–126. [Google Scholar] [CrossRef]
- Lustrino, M.; Duggen, S.; Rosenberg, C.L. The Central-Western Mediterranean: Anomalous igneous activity in an anomalous collisional tectonic setting. Earth-Sci. Rev. 2011, 104, 1–40. [Google Scholar] [CrossRef]
- Aparicio, A.; Mitjavila, J.M.; Araña, V.; Villa, I.M. La edad del volcanismo de las islas Columbrete Grande y Alborán (Mediterráneo occidental). Bol. Geol. Min. 1991, 102, 562–570. [Google Scholar]
- Juan, A.; Crespo, M.B.; Cowan, R.S.; Lexer, C.; Fay, M.F. Patterns of variability and gene flow in Medicago citrina, an endangered endemic of islands in the western Mediterranean, as revealed by amplified fragment length polymorphism (AFLP). Mol. Ecol. 2004, 13, 2679–2690. [Google Scholar] [CrossRef] [PubMed]
- Castilla, A.M.; Bauwens, D. Observations on the natural history, present status, and conservation of the insular lizard Podarcis hispanica atrata on the Columbretes archipelago, Spain. Biol. Conserv. 1991, 58, 69–84. [Google Scholar] [CrossRef]
- Castilla, A.M.; Bauwens, D. La Lagartija de las Islas Columbretes; Conselleria de Medi Ambient, Generalitat Valenciana: Valencia, Spain, 1996; 183p. [Google Scholar]
- Castilla, A.M.; Fernández-Pedrosa, V.; Backeljau, T.; González, A.; Latorre, A.; Moya, A. Conservation genetics of insular Podarcis lizards using partial cytochrome b sequences. Mol. Ecol. 1998, 7, 1407–1411. [Google Scholar] [CrossRef] [Green Version]
- Castilla, A.M.; Fernandez-Pedrosa, V.; Harris, D.J.; Gonzalez, A.; Latorre, A.; Moya, A. Mitochondrial DNA divergence suggests that Podarcis hispanica atrata (Squamata: Lacertidae) from the Columbretes Islands merits specific distinction. Copeia 1998, 4, 1037–1040. [Google Scholar] [CrossRef]
- Harris, D.J.; Carranza, S.; Arnold, E.N.; Pinho, C.; Ferrand, N. Complex biogeographical distribution of genetic variation within Podarcis wall lizards across the Strait of Gibraltar. J. Biogeogr. 2002, 29, 1257–1262. [Google Scholar] [CrossRef] [Green Version]
- Harris, D.J.; Sa-Sousa, P. Molecular phylogenetics of Iberian wall lizards (Podarcis): Is Podarcis hispanica a species complex? Mol. Phylogenetics Evol. 2002, 23, 75–81. [Google Scholar] [CrossRef]
- Pinho, C.; Ferrand, N.; Harris, D.J. Reexamination of the Iberian and North African Podarcis (Squamata: Lacertidae) phylogeny based on increased mitochondrial DNA sequencing. Mol. Phylogenetics Evol. 2006, 38, 266–273. [Google Scholar] [CrossRef]
- Pinho, C.; Harris, D.J.; Ferrand, N. Contrasting patterns of population subdivision and historical demography in three western Mediterranean lizard species inferred from mitochondrial DNA variation. Mol. Ecol. 2007, 16, 1191–1205. [Google Scholar] [CrossRef] [PubMed]
- Pinho, C.; Harris, D.J.; Ferrand, N. Non-equilibrium estimates of gene flow inferred from nuclear genealogies suggest that Iberian and North African wall lizards (Podarcis spp.) are an assemblage of incipient species. BMC Evol. Biol. 2008, 8, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renoult, J.P.; Geniez, P.; Bacquet, P.; Guillaume, C.P.; Crochet, P.A. Systematics of the Podarcis hispanicus-complex (Sauria, Lacertidae) II: The valid name of the north-eastern Spanish form. Zootaxa 2010, 2500, 58–68. [Google Scholar] [CrossRef]
- Kaliontzopoulou, A.; Pinho, C.; Harris, D.J.; Carretero, M.A. When cryptic diversity blurs the picture: A cautionary tale from Iberian and North African Podarcis wall lizards. Biol. J. Linn. Soc. 2011, 103, 779–800. [Google Scholar] [CrossRef] [Green Version]
- Renoult, J.P.; Geniez, P.; Bacquet, P.; Benoit, L.; Crochet, P.A. Morphology and nuclear markers reveal extensive mitochondrial introgressions in the Iberian Wall Lizard species complex. Mol. Ecol. 2009, 18, 4298–4315. [Google Scholar] [CrossRef] [PubMed]
- Bassitta, M.; Buades, J.M.; Pérez-Cembranos, A.; Pérez-Mellado, V.; Terrasa, B.; Brown, R.P.; Navarro, P.; Lluch, J.; Ortega, J.; Castro, J.A.; et al. Multilocus and morphological analysis of south-eastern Iberian Wall lizards (Squamata, Podarcis). Zool. Scr. 2020, 49, 668–683. [Google Scholar] [CrossRef]
- Castilla, A.M.; Bauwens, D. Thermal biology, microhabitat selection, and conservation of the insular lizard Podarcis hispanica atrata. Oecologia 1991, 85, 366–374. [Google Scholar] [CrossRef]
- Castilla, A.M. Podarcis atrata. In Atlas y Libro Rojo de los Anfibios y Reptiles de España; Pleguezuelos, J.M., Márquez, R., Lizana, M., Eds.; Dirección General de Conservación de la Naturaleza-Asociación Herpetológica Española: Madrid, Spain, 2002; pp. 238–239. [Google Scholar]
- Castilla, A.M.; Pastor, E.; Pons, G.X. Annual variation in the density of orb-web spiders of the genus Argiope at the Columbretes Islands: Consequences on the endemic lizard Podarcis atrata. Boll. Soc. Hist. Nat. Balear. 2006, 49, 137–143. [Google Scholar]
- Kocher, T.D.; Thomas, W.K.; Meyer, A.; Edwards, S.V.; Pääbo, S.; Villablanca, F.X.; Wilson, A.C. Dynamics of mitochondrial DNA evolution in animals: Amplification and sequencing with conserved primers. Proc. Natl. Acad. Sci. USA 1989, 86, 6196–6200. [Google Scholar] [CrossRef] [Green Version]
- Kartzinel, T.R.; Pringle, R.M. Molecular detection of invertebrate prey in vertebrate diets: Trophic ecology of Caribbean island lizards. Mol. Ecol. Resour. 2015, 15, 903–914. [Google Scholar] [CrossRef]
- Alonso, H.; Granadeiro, J.P.; Waap, S.; Xavier, J.; Symondson, W.O.; Ramos, J.A.; Catry, P. An holistic ecological analysis of the diet of Cory’s shearwaters using prey morphological characters and DNA barcoding. Mol. Ecol. 2014, 23, 3719–3733. [Google Scholar] [CrossRef] [PubMed]
- Pianka, E.P.; Vitt, L.J. Lizards, Windows to the Evolution of Diversity; Pianka, E.R., Vitt, L.J., Eds.; University of California Press: Oakland, CA, USA, 2003. [Google Scholar]
- Bieke, V.; Anthony, H.; Raoul, V.D. Interactions between Habitat Use, Behavior and the Trophic Niche of Lacertid Lizards. In Lizard Ecology: The Evolutionary Consequences of Foraging Mode; Reilly, S.M., McBrayer, L.B., Miles, D.B., Eds.; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Diaz, J.A.; Carrascal, L.M. Prey Size and Food Selection of Psammodromus algirus (Lacertidae) in Central Spain. J. Herpetol. 1990, 24, 342–347. [Google Scholar] [CrossRef] [Green Version]
- Polis, G.A.; Hurd, S.D. Linking marine and terrestrial food webs: Allochthonous input from the ocean supports high secondary productivity on small islands and coastal land communities. Am. Nat. 1996, 147, 396–423. [Google Scholar] [CrossRef]
- Barret, K.; Anderson, W.B.; Wait, A.D.; Grismer, L.L.; Polis, G.A.; Rose, M.D. Marine subsidies alter the diet and abundance of insular and coastal lizard populations. Oikos 2005, 109, 145–153. [Google Scholar] [CrossRef]
- Towns, D.R. Ecology of the black shore skink, Leiolopisma suteri (Lacertilia: Scincidae), in boulder beach habitats. N. Z. J. Zool. 1975, 2, 389–407. [Google Scholar] [CrossRef]
- Catenazzi, A.; Carrillo, J.; Donnelly, M.A. Seasonal and geographic eurythermy in a coastal Peruvian lizard. Copeia 2005, 4, 713–723. [Google Scholar] [CrossRef]
- Castilla, A.M.; Jiménez, J.; Lacomba, I.; Alonso Matilla, L.A.; Carretero, J.L.; García-Carrascosa, A.M. Los reptiles de Columbretes. In Islas Columbretes. Contribución al Estudio de su Medio Natural; Conselleria d’Agricultura, Medi Ambient, Canvi Climàtic i Desenvolupament Rural, Eds.; Generalitat Valenciana: Valencia, Spain, 1987; pp. 181–194. [Google Scholar]
- Castilla, A.M.; Herrel, A.; Gosá, A. Marine prey in the diet of Podarcis atrata from the Columbretes Islands. Munibe 2009, 57, 295–298. [Google Scholar]
- Castilla, A.M.; Dunlap, P. Rate of infanticide by adults in a natural population of the insular lizard, Podarcis hispanica atrata. In Abstracts of the 2nd International Symposium on the Lacertids of the Mediterranean Basin; Quinta de Marim: Algarve, Portugal, 1995; p. 12. [Google Scholar]
- Pérez-Cembranos, A.; León, A.; Pérez-Mellado, V. Omnivory of an Insular Lizard: Sources of Variation in the Diet of Podarcis lilfordi (Squamata, Lacertidae). PLoS ONE 2016, 11, e0148947. [Google Scholar] [CrossRef]
- Valentini, A.; Miquel, C.; Nawaz, M.A.; Bellemain, E.; Coissac, E.; Pompanon, F.; Gielly, L.; Cruaud, C.; Nascetti, G.; Wincker, P.; et al. New perspectives in diet analysis based on DNA barcoding and parallel pyrosequencing: The trnL approach. Mol. Ecol. Resour. 2009, 9, 51–60. [Google Scholar] [CrossRef]
- Pompanon, F.; Deagle, B.E.; Symondson, W.O.C.; Brown, D.S.; Jarman, S.N.; Taberlet, P. Who is eating what: Diet assessment using next generation sequencing. Mol. Ecol. 2012, 21, 1931–1950. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef] [PubMed]
- Symondson, W.O.C. Molecular identification of prey in predator diets. Mol. Ecol. 2002, 11, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Jurado-Rivera, J.A.; Vogler, A.P.; Reid, C.A.M.; Petitpierre, E.; Gómez-Zurita, J. DNA barcoding insect-host plant associations. Proc. R. Soc. B Biol. Sci. 2009, 276, 639–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelfattah, A.; Malacrino, A.; Wisniewski, M.; Cacciola, S.O.; Schena, L. Metabarcoding: A powerful tool to investigate microbial communities and shape future plant protection strategies. Biol. Control 2017, 120, 1–10. [Google Scholar] [CrossRef]
- Moeller, A.H.; Sanders, J.G. Roles of the gut microbiota in the adaptive evolution of mammalian species. Philos. Trans. R. Soc. B Biol. Sci. 2020, 375, 20190597. [Google Scholar] [CrossRef]
- Tamura, K.; Brumer, H. Glycan utilization systems in the human gut microbiota: A gold mine for structural discoveries. Curr. Opin. Struct. Biol. 2021, 68, 26–40. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Gerardo, N.M.; Inouye, D.W.; Medina, M.; Xavier, J.B. Animal behavior and the microbiome. Science 2012, 338, 198–199. [Google Scholar] [CrossRef]
- Russell, C.W.; Bouvaine, S.; Newell, P.D.; Douglas, A.E. Shared metabolic pathways in a coevolved insect-bacterial symbiosis. Appl. Environ. Microbiol. 2013, 79, 6117–6123. [Google Scholar] [CrossRef] [Green Version]
- Moeller, A.H.; Caro-Quintero, A.; Mjungu, D.; Georgiev, A.V.; Lonsdorf, E.V.; Muller, M.N.; Pusey, A.E.; Peeters, M.; Hahn, B.H.; Ochman, H. Cospeciation of gut microbiota with hominids. Science 2016, 353, 380–382. [Google Scholar] [CrossRef] [Green Version]
- Youngblut, N.D.; Reischer, G.H.; Walters, W.; Schuster, N.; Walzer, C.; Stalder, G.; Ley, R.E.; Farnleitner, A.H. Host diet and evolutionary history explain different aspects of gut microbiome diversity among vertebrate clades. Nat. Commun. 2019, 10, 2200. [Google Scholar] [CrossRef] [Green Version]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor: New York, NY, USA, 1989. [Google Scholar]
- Terrasa, B.; Pérez-Mellado, V.; Brown, R.P.; Picornell, A.; Castro, J.A.; Ramon, M.M. Foundations for conservation of intraspecific genetic diversity revealed by analysis of phylogeographical structure in the endangered endemic lizard Podarcis lilfordi. Divers. Distrib. 2009, 15, 207–221. [Google Scholar] [CrossRef]
- Rodríguez, V.; Brown, R.P.; Terrasa, B.; Pérez-Mellado, V.; Castro, J.A.; Picornell, A.; Ramon, M.M. Multilocus genetic diversity and historical biogeography of the endemic wall lizard from Ibiza and Formentera, Podarcis pityusensis (Squamata: Lacertidae). Mol. Ecol. 2013, 22, 4829–4841. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez, V.; Brown, R.P.; Terrasa, B.; Pérez-Mellado, V.; Picornell, A.; Castro, J.A.; Ramon, C. Genetic diversity and historical biogeography of the Maltese wall lizard, Podarcis filfolensis (Squamata: Lacertidae). Conserv. Genet. 2014, 15, 295–304. [Google Scholar] [CrossRef]
- Rodríguez, V.; Buades, J.M.; Brown, R.P.; Terrasa, B.; Pérez-Mellado, V.; Corti, C.; Delaugerre, M.; Castro, J.A.; Picornell, A.; Ramon, M.M. Evolutionary history of Podarcis tiliguerta on Corsica and Sardinia. BMC Evol. Biol. 2017, 17, 27. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Buades, J.M.; Rodríguez, V.; Terrasa, B.; Pérez-Mellado, V.; Brown, R.P.; Castro, J.A.; Picornell, A.; Ramon, M.M. Variability of the mc1r Gene in Melanic and Non-Melanic Podarcis lilfordi and Podarcis pityusensis from the Balearic Archipelago. PLoS ONE 2013, 8, e53088. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [Green Version]
- Tajima, F. Statistical method for testing the neutral mutation hypothesis by DNA polymorphism. Genetics 1989, 123, 585–595. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [PubMed]
- Lanfear, R.; Frandsen, P.B.; Wright, A.M.; Senfeld, T.; Calcott, B. PartitionFinder 2: New methods for selecting partitioned models of evolution for molecular and morphological phylogenetic analyses. Mol. Biol. Evol. 2016, 34, 772–773. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Nguyen, M.A.T.; von Haeseler, A. Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 2013, 30, 1188–1195. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the ultrafast bootstrap approximation. Mol. Biol. Evol. 2017, 35, 518–522. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3. 2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar]
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior summarization in Bayesian phylogenetics using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A. FigTree v. 1.4.4. Institute of Evolutionary Biology, University of Edinburgh, Edinburgh. 2014. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 20 June 2019).
- Clement, M.; Posada, D.; Crandall, K.A. TCS: A computer program to estimate gene genealogies. Mol. Ecol. 2000, 9, 1657–1659. [Google Scholar] [CrossRef] [Green Version]
- Clement, M.; Snell, Q.; Walker, P.; Posada, D.; Crandall, K.A. TCS: Estimating gene genealogies. In International Parallel & Distributed Processing Symposium; IEEE Computer Society: Fort Lauderdale, CA, USA, 2002; Volume 1, p. 7. [Google Scholar]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Heled, J.; Drummond, A.J. Bayesian inference of species trees from multilocus data. Mol. Biol. Evol. 2010, 27, 570–580. [Google Scholar] [CrossRef] [Green Version]
- Brown, R.P.; Terrasa, B.; Pérez-Mellado, V.; Castro, J.A.; Hoskisson, P.A.; Picornell, A.; Ramon, M.M. Bayesian estimation of post-Messinian divergence times in Balearic Island lizards. Mol. Phylogenetics Evol. 2008, 48, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Ren, T.; Kahrl, A.F.; Wu, M.; Cox, R.M. Does adaptive radiation of a host lineage promote ecological diversity of its bacterial communities? A test using gut microbiota of Anolis lizards. Mol. Ecol. 2016, 25, 4793–4804. [Google Scholar] [CrossRef] [PubMed]
- Ahasan, M.S.; Waltzek, T.B.; Huerlimann, R.; Ariel, E. Fecal bacterial communities of wild-captured and stranded green turtles (Chelonia mydas) on the Great Barrier Reef. FEMS Microbiol. Ecol. 2017, 93, fix139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandri, C.; Correa, F.; Spiezio, C.; Trevisi, P.; Luise, D.; Modesto, M.; Remy, S.; Muzungaile, M.M.; Checcucci, A.; Zaborra, C.A.; et al. Fecal microbiota characterization of Seychelles giant tortoises (Aldabrachelys gigantea) living in both wild and controlled environments. Front. Microbiol. 2020, 11, 569249. [Google Scholar] [CrossRef] [PubMed]
- Kohl, K.D.; Brun, A.; Magallanes, M.; Brinkerhoff, J.; Laspiur, A.; Acosta, J.C.; Caviedes-Vidal, E.; Bordenstein, S.R. Gut microbial ecology of lizards: Insights into diversity in the wild, effects of captivity, variation across gut regions and transmission. Mol. Ecol. 2017, 26, 1175–1189. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Lauber, C.L.; Walters, W.A.; Berg-Lyons, D.; Lozupone, C.A.; Turnbaugh, P.J.; Fierer, N.; Knight, R. Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc. Natl. Acad. Sci. USA 2011, 108, 4516–4522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A new versatile primer set targeting a short fragment of the mitochondrial COI region for metabarcoding metazoan diversity: Application for characterizing coral reef fish gut contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [Green Version]
- Geller, J.B.; Meyer, C.P.; Parker, M.; Hawk, H. Redesign of PCR primers for mitochondrial Cytochrome c oxidase subunit I for marine invertebrates and application in all-taxa biotic surveys. Mol. Ecol. Resour. 2013, 13, 851–861. [Google Scholar] [CrossRef]
- Mallott, E.K.; Malhi, R.S.; Garber, P.A. Brief communication: High-Throughput sequencing of fecal DNA to Identify Insects Consumed by Wild Weddell’s Saddleback Tamarins (Saguinus weddelli, Cebidae, Primates) in Bolivia. Am. J. Phys. Anthropol. 2015, 156, 474–481. [Google Scholar] [CrossRef]
- Levin, R.A.; Wagner, W.L.; Hoch, P.C.; Nepokroeff, M.; Pires, J.C.; Zimmer, E.A.; Sytsma, K.J. Family-level relationships of Onagraceae based on chloroplast rbcL and ndhF data. Am. J. Bot. 2003, 90, 107–115. [Google Scholar] [CrossRef] [Green Version]
- Kress, W.; Erickson, D.L.; Jones, F.A.; Swenson, N.G.; Perez, R.; Sanjur, O.; Bermingham, E. Plant DNA barcodes and a community phylogeny of a tropical forest dynamics plot in Panama. Proc. Natl. Acad. Sci. USA 2009, 106, 18621–18626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, J.A.; Simpson, B.B. Paraphyly of Tarasa (Malvaceae) and Diverse Origins of the Polyploid Species. Syst. Bot. 2003, 28, 723–737. [Google Scholar]
- Albanese, D.; Fontana, P.; De Filippo, C.; Cavalieri, D.; Donati, C. MICCA: A complete and accurate software for taxonomic profiling of metagenomic data. Sci. Rep. 2015, 5, 9743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Hillis, D.M.; Bull, J.J. An Empirical Test of Bootstrapping as a Method for Assessing Confidence in Phylogenetic Analysis. Syst. Biol. 1993, 42, 182–192. [Google Scholar] [CrossRef]
- da Silva, L.P.; Mata, V.A.; Lopes, P.B.; Pereira, P.; Jarman, S.N.; Lopes, R.J.; Beja, P. Advancing the integration of multi-marker metabarcoding data in dietary analysis of trophic generalists. Mol. Ecol. Resour. 2019, 19, 1420–1432. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335. [Google Scholar] [CrossRef] [Green Version]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Price, M.N.; Dehal, P.S.; Arkin, A.P. FastTree: Computing Large Minimum Evolution Trees with Profiles instead of a Distance Matrix. Mol. Biol. Evol. 2009, 26, 1641–1650. [Google Scholar] [CrossRef]
- McMurdie, P.J.; Holmes, S. Phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Podnar, M.; Haring, E.; Pinsker, W.; Mayer, W. Unusual origin of a nuclear pseudogene in the Italian wall lizard: Intergenomic and interspecific transfer of a large section of the mitochondrial genome in the genus Podarcis (Lacertidae). J. Mol. Evol. 2007, 64, 308–320. [Google Scholar] [CrossRef] [PubMed]
- Palumbi, S.R. Nucleic acids II: The polymerase chain reaction. In Molecular Systematics; Hillis, D.M., Moritz, C., Mable, B.K., Eds.; Sinauer Ass: Sunderland, UK, 1996; pp. 205–247. [Google Scholar]
- Slatkin, M.; Hudson, R.R. Pairwise comparisons of mitochondrial DNA sequences in stable and exponentially growing populations. Genetics 1991, 129, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Emig, C.C.; Geistdoerfer, P. The Mediterranean deep-sea fauna: Historical evolution, bathymetric variations and geographical changes. Carnets Geol. 2004, 2004, 1–10. [Google Scholar] [CrossRef]
- Greene, H.W.; Mossakowski, D.; Roth, G. Dietary and phenotypic diversity in lizards: Why are some organisms specialised? In Environmental Adaptation and Evolution; Mossakowski, D., Roth, G., Eds.; G. Fischer Verlag: Stuttgart, Germany, 1982; pp. 107–128. [Google Scholar]
- Cooper, W.E.; Vitt, L.J. Distribution, extent, and evolution of plant consumption by lizards. J. Zool. 2002, 257, 487–517. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Mellado, V. Podarcis hispanica (Steindachner, 1870). In Reptiles Fauna Ibérica; Salvador, A., Ramos, M.A., Eds.; Museo Nacional de Ciencias Naturales (CSIC): Madrid, Spain, 1998; Volume 10, pp. 258–272. [Google Scholar]
- Escarré, A.; Pericad, J.R. Fauna alicantina. I. Saurios y ofidios. Cuadernos de la fauna alicantina; Publicaciones del Instituto de Estudios Alicantinos, Serie II: Alicante, Spain, 1981; Volume 15, pp. 1–101. [Google Scholar]
- Llorente, G.A.; Llorente, C.; Santos, X.; Ruiz, X. Algunos datos sobre la alimentación de una población de Podarcis hispanica (Steindachner, 1870) del área metropolitana de Barcelona. In Resúmenes del I Congreso Nacional de Herpetología; Asociación Herpetológica Española: Benicàssim, Spain, 1986; p. 67. [Google Scholar]
- Castilla, A.M.; Vanhooydonck, B.; Catenazzi, A. Feeding behaviour of the Columbretes lizard Podarcis atrata, in relation to Isopoda (Crustaceae) species: Ligia italica and Armadillo officinalis. Belg. J. Zool. 2008, 138, 146–148. [Google Scholar]
- Gil, V.; Pinho, C.J.; Aguiar, C.A.S.; Jardim, C.; Rebelo, R.; Vasconcelos, R. Questioning the proverb ‘more haste, less speed’: Classic versus metabarcoding approaches for the diet study of a remote island endemic gecko. PeerJ 2020, 8, e8084. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.; Blaxter, M. Ancient origin of glycosyl hydrolase family 9 cellulase genes. Mol. Biol. Evol. 2005, 22, 1273–1284. [Google Scholar] [CrossRef]
- Biddle, A.; Stewart, L.; Blanchard, J.; Leschine, S. Untangling the genetic basis of fibrolytic specialization by Lachnospiraceae and Ruminococcaceae in diverse gut communities. Diversity 2013, 5, 627–640. [Google Scholar] [CrossRef]
- Baldo, L.; Riera, J.L.; Mitsi, K.; Pretus, J.L. Processes shaping gut microbiota diversity in allopatric populations of the endemic lizard Podarcis lilfordi from Menorcan islets (Balearic Islands). FEMS Microbiol. Ecol. 2018, 94, fix186. [Google Scholar] [CrossRef]
- Bloodgood, J.C.G.; Hernandez, S.M.; Isaiah, A.; Suchodolski, J.S.; Hoopes, L.A.; Thompson, P.M.; Waltzek, T.B.; Norton, T.M. The effect of diet on the gastrointestinal microbiome of juvenile rehabilitating green turtles (Chelonia mydas). PLoS ONE 2020, 15, e0227060. [Google Scholar]
- Kapli, P.; Botoni, D.; Ilgaz, Ç.; Kumlutaş, Y.; Avcı, A.; Rastegar-Pouyani, N.; Fathinia, B.; Lymberakis, P.; Ahmadzadeh, F.; Poulakakis, N. Molecular phylogeny and historical biogeography of the Anatolian lizard Apathya (Squamata, Lacertidae). Mol. Phylogenetics Evol. 2013, 66, 992–1001. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| N (N Phased) | bp | S | h | Hd | π | k | D Tajima (1989) | |
|---|---|---|---|---|---|---|---|---|
| Nuclear genes | ||||||||
| RAG1 | 26 (52) | 939 | 12 | 12 | 0.704 ± 0.045 | 0.00192 ± 0.00015 | 1.802 | −0.572 n.s. |
| MC1R | 12 (24) | 672 | 12 | 9 | 0.768 ± 0.078 | 0.00586 ± 0.00067 | 3.069 | −0.154 n.s. |
| KIAA2018 | 10 (20) | 489 | 3 | 3 | 0.563 ± 0.063 | 0.00230 ± 0.00026 | 1.126 | 0.886 n.s. |
| APOBE28 | 26 (52) | 665 | 10 | 15 | 0.882 ± 0.027 | 0.00540 ± 0.00020 | 3.593 | 1.770 n.s. |
| mtDNA | ||||||||
| CYTB, NADH, CR, 12S | 28 | 2311 | 11 | 10 | 0.770 ± 0.076 | 0.00091 ± 0.00017 | 2.106 | −0.833 n.s. |
| COI | 10 | 657 | 1 | 2 | 0.200 ± 0.015 | 0.00030 ± 0.00023 | 0.200 | −1.111 n.s. |
| Phylum | Class | Order | Family | Genus | Species | Number of Faecal Samples |
|---|---|---|---|---|---|---|
| Arthropoda | ||||||
| Arachnida | Araneae | Araneidae | 1 | |||
| Insecta | Coleoptera | 1 | ||||
| Curculionidae | Trachyphloeus | 1 | ||||
| Tenebrionidae | 1 | |||||
| Hemiptera | Rhyparochromidae | Lamprodema | maurum | 1 | ||
| Hymenoptera | Formicidae | Pheidole | 1 | |||
| Lepidoptera | 1 | |||||
| Blastobasidae | Blastobasis | 1 | ||||
| Geometridae | 2 | |||||
| Psychidae | 2 | |||||
| Malacostraca | Isopoda | 3 | ||||
| Armadillidae | Armadillo | officinalis | 2 | |||
| Streptophyta | ||||||
| Magnoliopsida | Apiales | Apiaceae | Daucus | 1 | ||
| Caryophyllales | Chenopodiaceae | 2 | ||||
| Beta | vulgaris | 1 | ||||
| Patellifolia | 1 | |||||
| Suaeda | 5 | |||||
| Plumbaginaceae | Limonium | 6 | ||||
| Portulacaceae | Portulaca | 2 | ||||
| Rosales | Rosaceae | Prunus | 3 | |||
| Ulmaceae | Ulmus | 1 | ||||
| Solanales | Solanaceae | Lycium | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bassitta, M.; Alemany, I.; Pérez-Mellado, V.; Pérez-Cembranos, A.; Navarro, P.; Lluch, J.; Jurado-Rivera, J.A.; Castro, J.A.; Picornell, A.; Ramon, C. Phylogenetic, Microbiome, and Diet Characterisation of Wall Lizards in the Columbretes Archipelago (Spain): Clues for Their Conservation. Diversity 2022, 14, 408. https://doi.org/10.3390/d14050408
Bassitta M, Alemany I, Pérez-Mellado V, Pérez-Cembranos A, Navarro P, Lluch J, Jurado-Rivera JA, Castro JA, Picornell A, Ramon C. Phylogenetic, Microbiome, and Diet Characterisation of Wall Lizards in the Columbretes Archipelago (Spain): Clues for Their Conservation. Diversity. 2022; 14(5):408. https://doi.org/10.3390/d14050408
Chicago/Turabian StyleBassitta, Marta, Iris Alemany, Valentín Pérez-Mellado, Ana Pérez-Cembranos, Pilar Navarro, Javier Lluch, José A. Jurado-Rivera, José A. Castro, Antònia Picornell, and Cori Ramon. 2022. "Phylogenetic, Microbiome, and Diet Characterisation of Wall Lizards in the Columbretes Archipelago (Spain): Clues for Their Conservation" Diversity 14, no. 5: 408. https://doi.org/10.3390/d14050408
APA StyleBassitta, M., Alemany, I., Pérez-Mellado, V., Pérez-Cembranos, A., Navarro, P., Lluch, J., Jurado-Rivera, J. A., Castro, J. A., Picornell, A., & Ramon, C. (2022). Phylogenetic, Microbiome, and Diet Characterisation of Wall Lizards in the Columbretes Archipelago (Spain): Clues for Their Conservation. Diversity, 14(5), 408. https://doi.org/10.3390/d14050408