
Seasonal Shifts in the Morphological Traits of Bloom-Forming Cyanobacteria in Lake Chaohu, China



Abstract
:1. Introduction
2. Materials and Methods
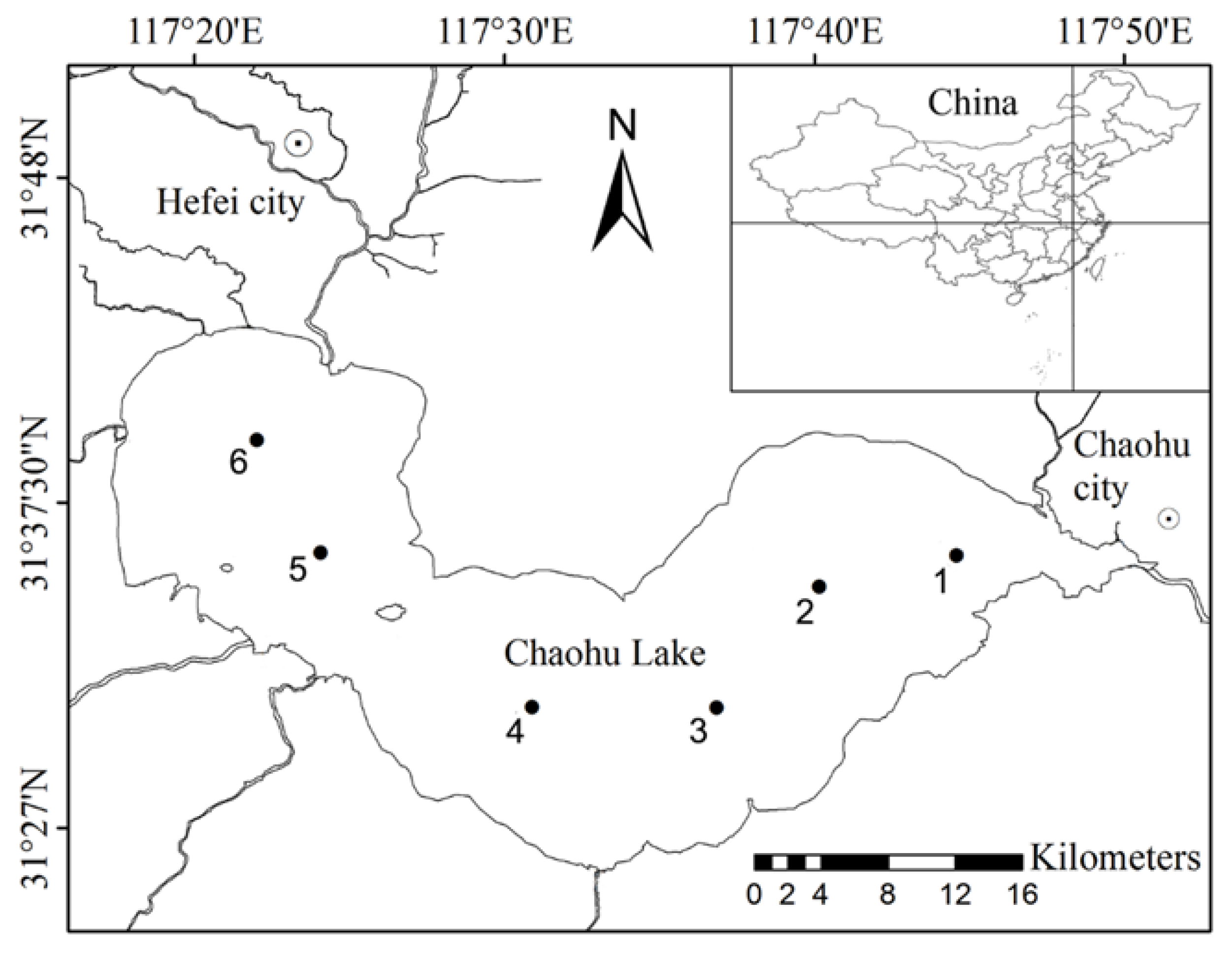
2.1. Study Lake
2.2. Sampling Method
2.3. Statistical Analysis
3. Results
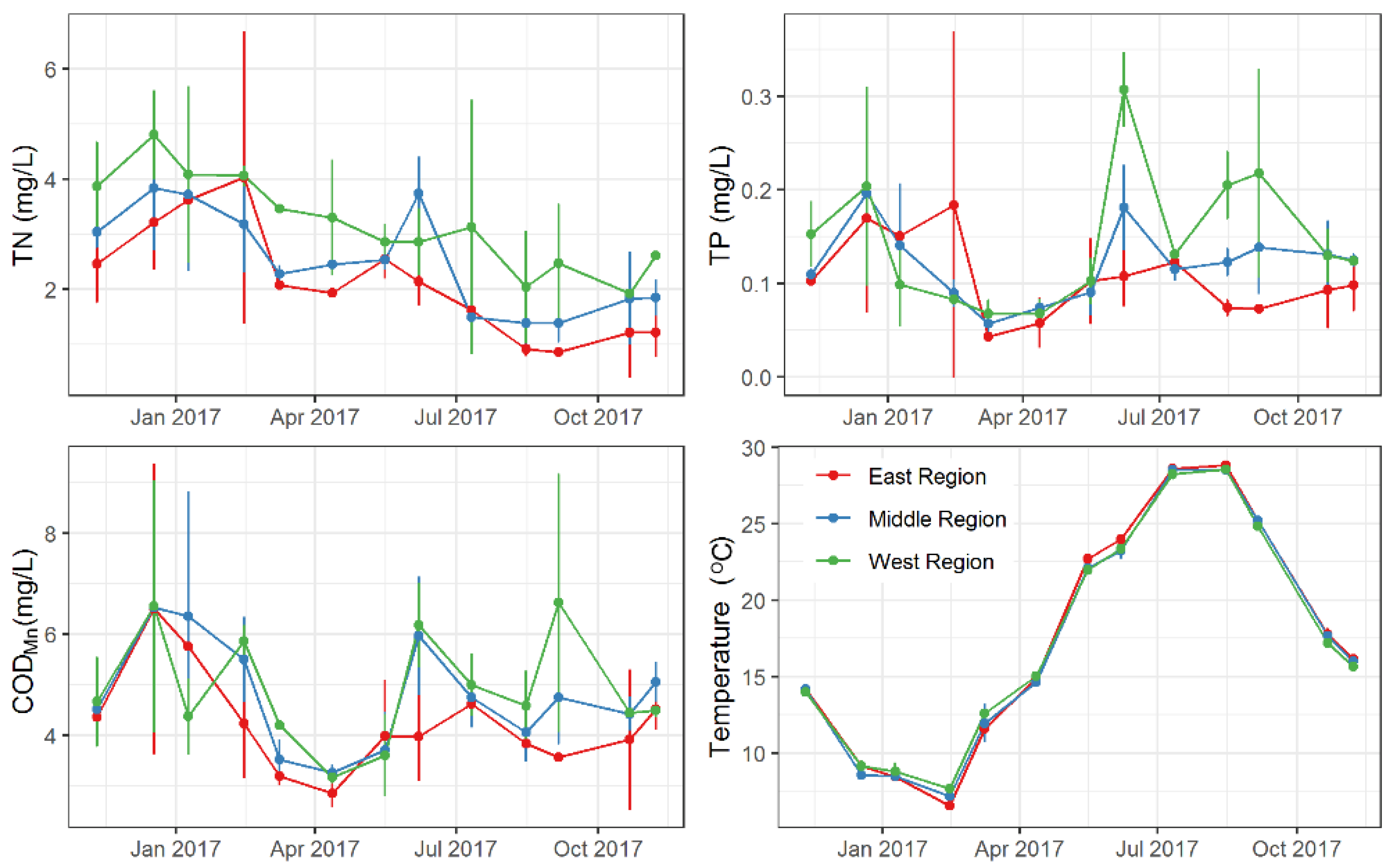
3.1. Monthly Variations in Environmental Factors
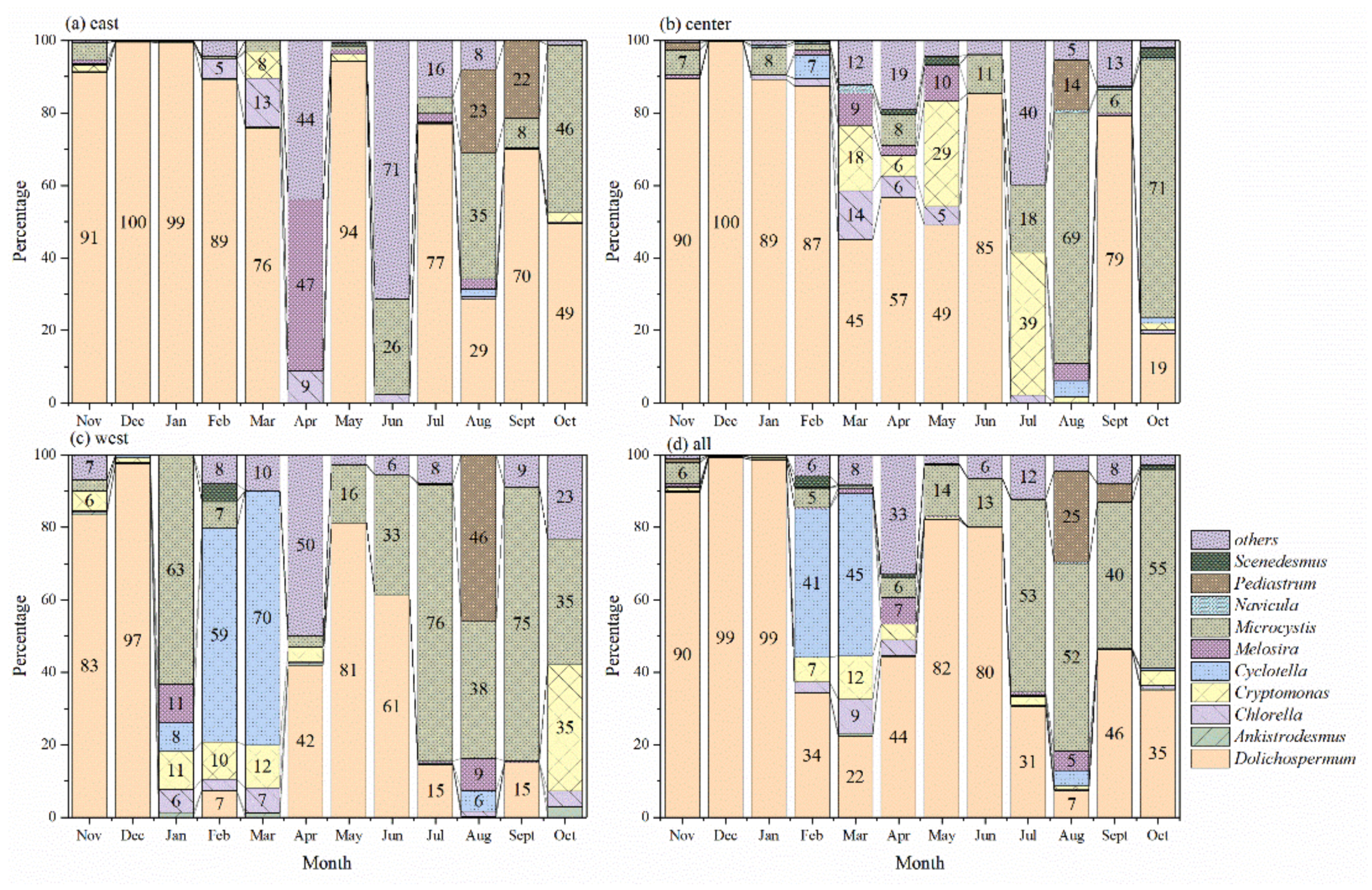
3.2. Seasonal Shift in Phytoplankton Composition
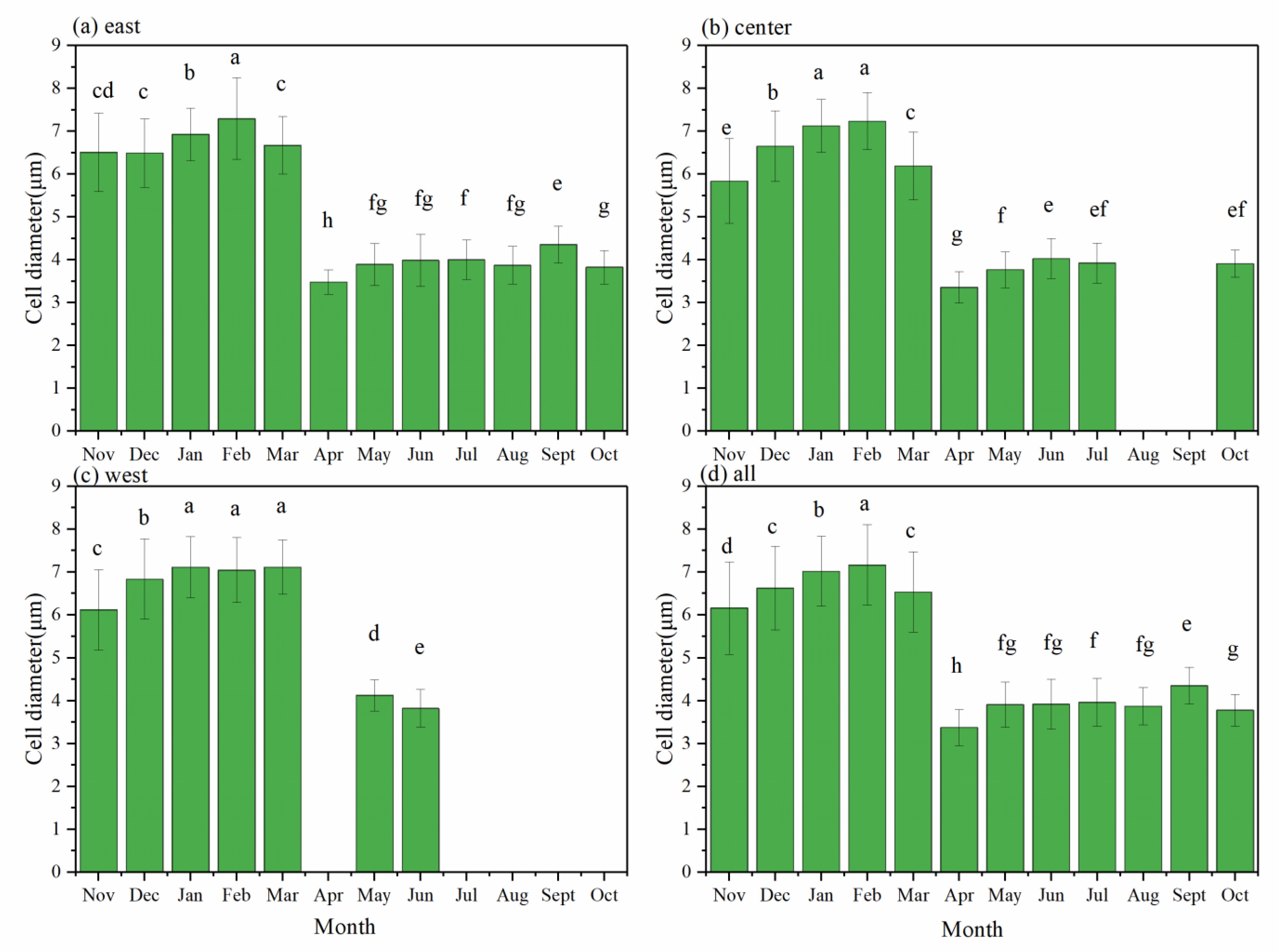
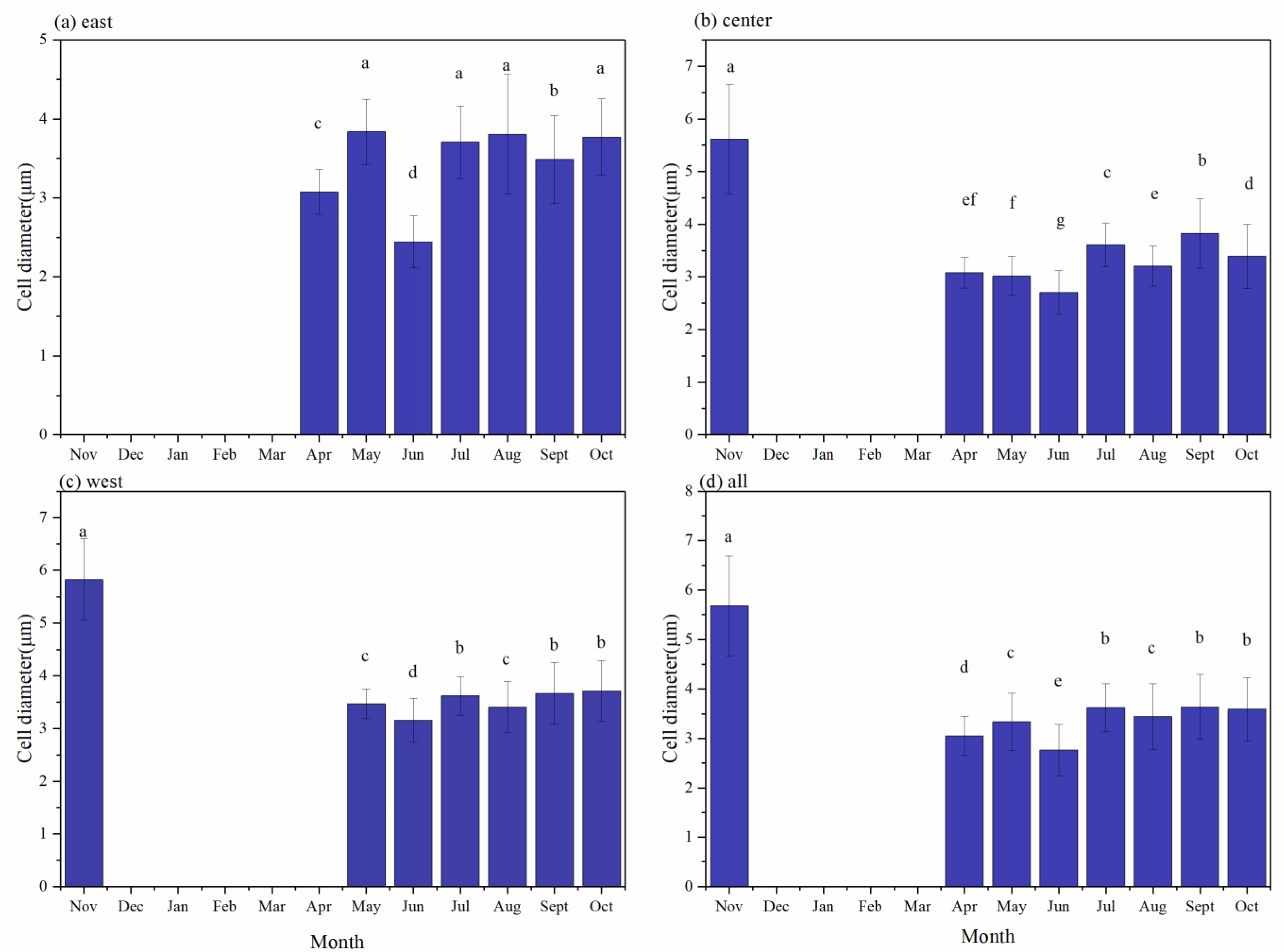
3.3. Cell Diameters in Dolichospermum and Microcystis
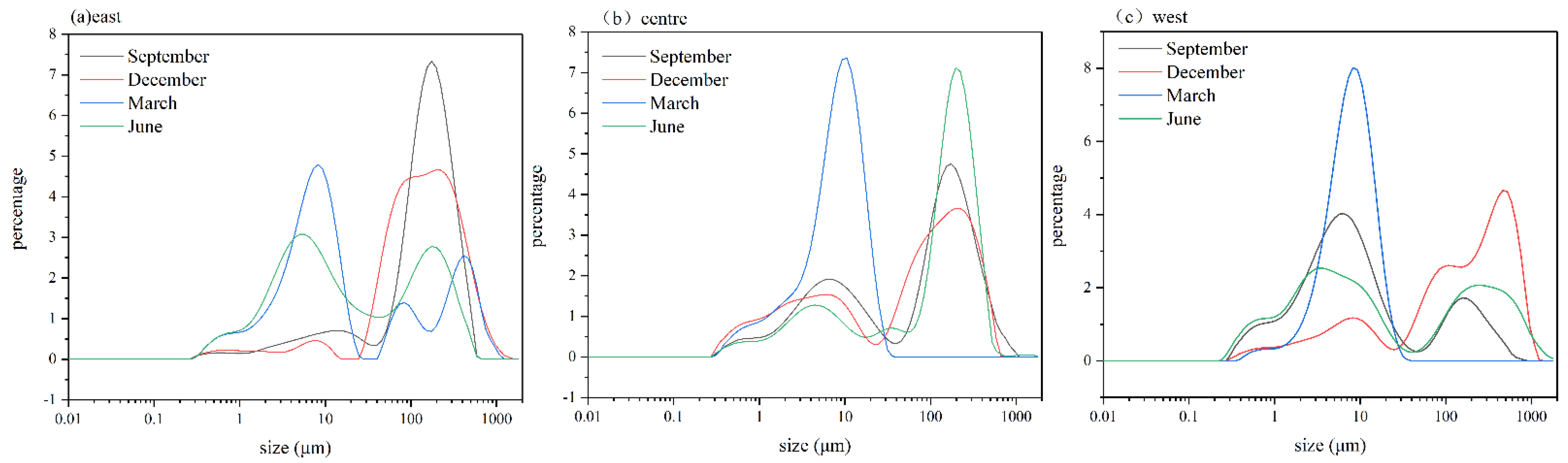
3.4. Colony Size in Different Lake Regions
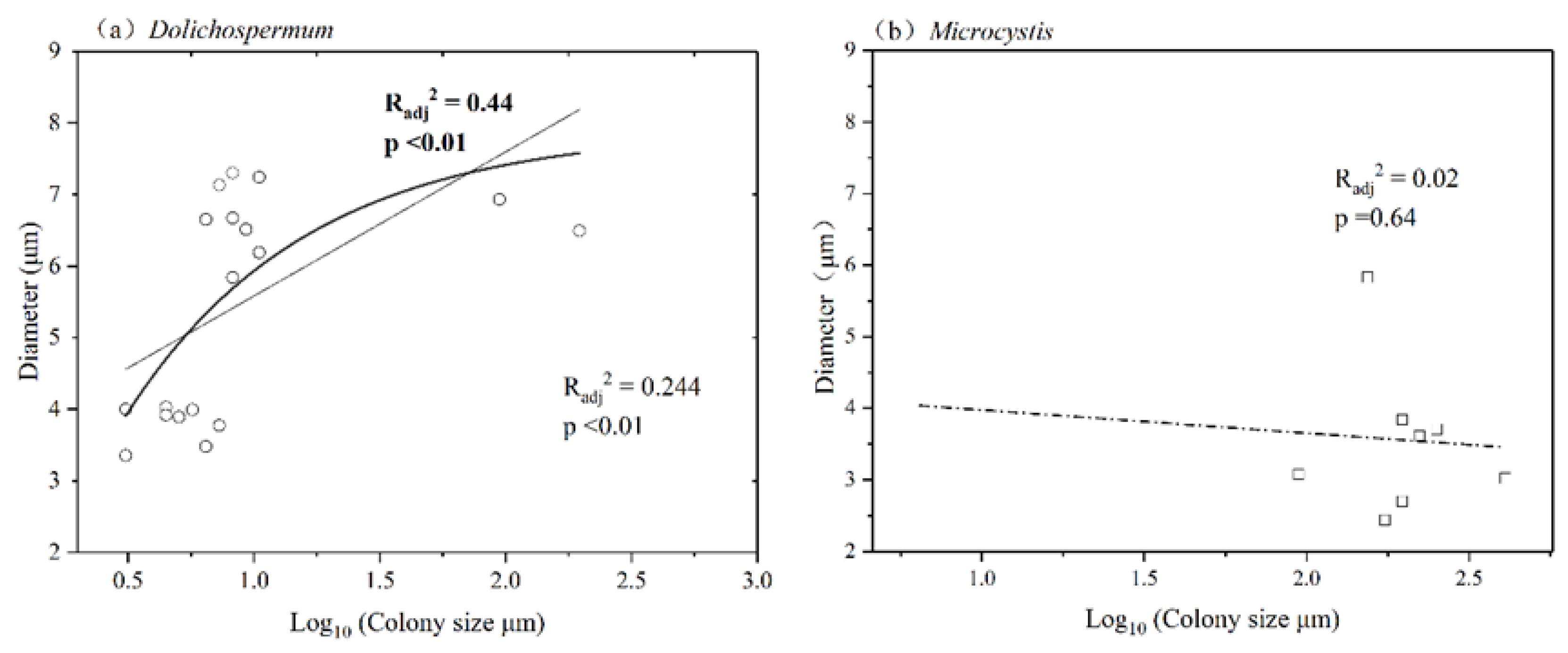
3.5. Corresponding Relationships between Cell Diameter and Colony Size
4. Discussion
4.1. Spatiotemporal Variation in Algal Composition
4.2. The Variation in Colony Size and Cell Diameter
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shan, K.; Song, L.; Chen, W.; Li, L.; Liu, L.; Wu, Y.; Jia, Y.; Zhou, Q.; Peng, L. Analysis of environmental drivers influencing interspecific variations and associations among bloom-forming cyanobacteria in large, shallow eutrophic lakes. Harmful Algae 2019, 84, 84–94. [Google Scholar] [CrossRef] [PubMed]
- Potočár, T.; Pereira, J.A.V.; Brányiková, I.; Barešová, M.; Pivokonský, M.; Brányik, T. Alkaline flocculation of Microcystis aeruginosa induced by calcium and magnesium precipitates. J. Appl. Phycol. 2019, 32, 329–337. [Google Scholar] [CrossRef]
- Jochimsen, M.C.; Kümmerlin, R.; Straile, D. Compensatory dynamics and the stability of phytoplankton biomass during four decades of eutrophication and oligotrophication. Ecol. Lett. 2012, 16, 81–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeppesen, E.; Søndergaard, M.; Jensen, J.P.; Havens, K.E.; Anneville, O.; Carvalho, L.; Coveney, M.F.; Deneke, R.; Dokulil, M.T.; Foy, B. Lake responses to reduced nutrient loading–an analysis of contemporary long-term data from 35 case studies. Freshwat. Biol. 2005, 50, 1747–1771. [Google Scholar] [CrossRef]
- Rigosi, A.; Carey, C.C.; Ibelings, B.W.; Brookes, J.D. The interaction between climate warming and eutrophication to promote cyanobacteria is dependent on trophic state and varies among taxa. Limnol. Oceanogr. 2014, 59, 99–114. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Shi, X.; Yang, Z.; Yu, Y.; Shi, L.; Qin, B. Long-term dynamics and drivers of phytoplankton biomass in eutrophic Lake Taihu. Sci. Total Environ. 2018, 645, 876–886. [Google Scholar] [CrossRef]
- Edwards, K.F.; Litchman, E.; Klausmeier, C.A. Functional traits explain phytoplankton community structure and seasonal dynamics in a marine ecosystem. Ecol. Lett. 2013, 16, 56–63. [Google Scholar] [CrossRef]
- Guan, Y.; Zhang, M.; Yang, Z.; Shi, X.; Zhao, X. Intra-annual variation and correlations of functional traits in Microcystis and Dolichospermum in Lake Chaohu. Ecol. Indic. 2020, 111, 106052. [Google Scholar] [CrossRef]
- Nguyen, M.K.; Moon, J.-Y.; Lee, Y.-C. Loading effects of low doses of magnesium aminoclay on microalgal Microcystis sp. KW growth, macromolecule productions, and cell harvesting. Biomass Bioenergy 2020, 139, 105619. [Google Scholar] [CrossRef]
- Ehrlich, E.; Kath, N.J.; Gaedke, U. The shape of a defense-growth trade-off governs seasonal trait dynamics in natural phytoplankton. ISME J. 2020, 14, 1451–1462. [Google Scholar] [CrossRef]
- Li, R.; Chen, Q.; Zhang, X.; Recknagel, F. Effects of temperature and macronutrients on phytoplankton communities across three largely different lakes identified by a time–space trade-off approach. Ecol. Inform. 2015, 29, 174–181. [Google Scholar] [CrossRef]
- Yang, Z.; Kong, F.X.; Cao, H.S.; Shi, X.L. Observation on colony formation of Microcystis aeruginosa induced by filtered lake water under laboratory conditions. Ann. De Limnol.-Int. J. Limnol. 2009, 41, 169–173. [Google Scholar] [CrossRef]
- Chen, H.; Lurling, M. Calcium promotes formation of large colonies of the cyanobacterium Microcystis by enhancing cell-adhesion. Harmful Algae 2020, 92, 101768. [Google Scholar] [CrossRef]
- Tan, X.; Shu, X.; Duan, Z.; Parajuli, K. Two types of bound extracellular polysaccharides and their roles in shaping the size and tightness of Microcystis colonies. J. Appl. Phycol. 2019, 32, 255–262. [Google Scholar] [CrossRef]
- Li, M.; Zhu, W.; Guo, L.; Hu, J.; Chen, H.; Xiao, M. To increase size or decrease density? Different Microcystis species has different choice to form blooms. Sci. Rep. 2016, 6, 37056. [Google Scholar] [CrossRef] [Green Version]
- Brown, C.R. The ecology and evolution of colony-size variation. Behav. Ecol. Sociobiol. 2016, 70, 1613–1632. [Google Scholar] [CrossRef]
- Brandenburg, K.M.; Sylke, W.; Uwe, J.; Anke, K.; Jacqueline, J.; Bernd, K.; Van, D.; Ian, D. Intraspecific trait variation and trade-offs within and across populations of a toxic dinoflagellate. Ecol. Lett. 2018, 21, 1561–1571. [Google Scholar] [CrossRef] [Green Version]
- Kong, X.; He, Q.; Yang, B.; He, W.; Xu, F.; Janssen, A.B.G.; Kuiper, J.J.; van Gerven, L.P.A.; Qin, N.; Jiang, Y.; et al. Hydrological regulation drives regime shifts: Evidence from paleolimnology and ecosystem modeling of a large shallow Chinese lake. Glob. Chang. Biol. 2017, 23, 737–754. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Yang, Z.; Wei, L.; Yang, W.; Chen, C.; Kong, F. Spatial and seasonal shifts in bloom-forming cyanobacteria in Lake Chaohu: Patterns and driving factors. Phycol. Res. 2016, 64, 44–55. [Google Scholar] [CrossRef]
- Li, X.D.; Chen, Y.H.; Liu, C.; Hong, J.; Deng, H.; Yu, D.J. Eutrophication and related antibiotic resistance of enterococci in the Minjiang River, China. Microb. Ecol. 2020, 80, 1–13. [Google Scholar] [CrossRef]
- Utermöhl, U.F. Zur Vervollkommnung der quantitativen Erfassung des Planktons. Verh. Int. Ver. Limnol. 1958, 5, 567–595. [Google Scholar]
- Hu, H.; Wei, Y. The Freshwater Algae of China: Systematics, Taxonomy and Ecology; Science Press: Beijing, China, 2006. [Google Scholar]
- Li, R.; Watanabe, M.; Watanabe, M.M. Taxonomic studies of planktic species of Anabaena based on morphological characteristics in cultured strains. Hydrobiologia 2000, 438, 117–138. [Google Scholar] [CrossRef]
- Hillebrand, H.; Dürselen, C.D.; Kirschtel, D.; Pollingher, U.; Zohary, T. Biovolume calculation for pelagic and benthic microalgae. J. Phycol. 1999, 35, 403–424. [Google Scholar] [CrossRef]
- Rott, E. Chlorophyll-a-konzentrationen und zellvolumen als parameter der phytoplakntonbiomasse. Ber. Nat.-Med. Ver. Innsbr. 1978, 65, 11–21. [Google Scholar]
- R-C-Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Jin, Z.H.; Tu, C.Q.; Wang, S.H.; Chen, J.Y.; Lu, C.Y.; Huang, W. Phosphorus adsorption characteristics and loss risk in sediments of Lake Bay during the overwinter period of cyanobacteria. Environ. Sci. 2022, 43, 1976–1987. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Yin, J. Composition and influential factors of phytoplankton function groups in Lake Chaohu. J. Lake Sci. 2018, 30, 431–440. [Google Scholar] [CrossRef] [Green Version]
- Hu, M.; Zhang, Y.; Ma, R.; Xue, K.; Cao, Z.; Chu, Q.; Jing, Y. Optimized remote sensing estimation of the lake algal biomass by considering the vertically heterogeneous chlorophyll distribution: Study case in Lake Chaohu of China. Sci. Total Environ. 2021, 771, 144811. [Google Scholar] [CrossRef]
- Wan, L.; Chen, X.; Deng, Q.; Yang, L.; Li, X.; Zhang, J.; Song, C.; Zhou, Y.; Cao, X. Phosphorus strategy in bloom-forming cyanobacteria (Dolichospermum and Microcystis) and its role in their succession. Harmful Algae 2019, 84, 46–55. [Google Scholar] [CrossRef]
- Zhang, M.; Yang, Z.; Yu, Y.; Shi, X. Interannual and Seasonal Shift between Microcystis and Dolichospermum: A 7-Year Investigation in Lake Chaohu, China. Water 2020, 12, 1978. [Google Scholar] [CrossRef]
- Reynolds, C.S. Ecology of Phytoplankton (Ecology, Biodiversity and Conservation); Cambridge University Press: Cambridge, UK, 2006. [Google Scholar]
- Zhu, X.; Li, Q.; Yin, C.; Fang, X.; Xu, X. Role of Spermidine in Overwintering of Cyanobacteria. J. Bacteriol. 2015, 197, 2325–2334. [Google Scholar] [CrossRef] [Green Version]
- Misson, B.; Latour, D. Influence of light, sediment mixing, temperature and duration of the benthic life phase on the benthic recruitment of Microcystis. J. Plankton Res. 2011, 34, 113–119. [Google Scholar] [CrossRef]
- Symes, E.; van Ogtrop, F.F. The effect of pre-industrial and predicted atmospheric CO2 concentrations on the development of diazotrophic and non-diazotrophic cyanobacterium: Dolichospermum circinale and Microcystis aeruginosa. Harmful Algae 2019, 88, 101536. [Google Scholar] [CrossRef] [PubMed]
- Eigemann, F.; Schwartke, M.; Schulz-Vogt, H. Niche separation of Baltic Sea cyanobacteria during bloom events by species interactions and autecological preferences. Harmful Algae 2018, 72, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Ratcliff, W.C.; Fankhauser, J.D.; Rogers, D.W.; Greig, D.; Travisano, M. Origins of multicellular evolvability in snowflake yeast. Nat. Commun. 2015, 6, 6102. [Google Scholar] [CrossRef] [Green Version]
- Yang, Z.; Kong, F. Formation of large colonies: A defense mechanism of Microcystis aeruginosa under continuous grazing pressure by flagellate Ochromonas sp. J. Limnol. 2012, 71, e5. [Google Scholar] [CrossRef] [Green Version]
- Haakonsson, S.; Rodriguez-Gallego, L.; Somma, A.; Bonilla, S. Temperature and precipitation shape the distribution of harmful cyanobacteria in subtropical lotic and lentic ecosystems. Sci. Total Environ. 2017, 609, 1132. [Google Scholar] [CrossRef]
- Ma, J.; Brookes, J.D.; Qin, B.; Paerl, H.W.; Gao, G.; Wu, P.; Zhang, W.; Deng, J.; Zhu, G.; Zhang, Y.; et al. Environmental factors controlling colony formation in blooms of the cyanobacteria Microcystis spp. in Lake Taihu, China. Harmful Algae 2013, 31, 136–142. [Google Scholar] [CrossRef]
- Zhu, W.; Zhou, X.; Chen, H.; Gao, L.; Xiao, M.; Li, M. High nutrient concentration and temperature alleviated formation of large colonies of Microcystis: Evidence from field investigations and laboratory experiments. Water Res. 2016, 101, 167–175. [Google Scholar] [CrossRef]
- Xiao, M.; Li, M.; Reynolds, C.S. Colony formation in the cyanobacterium Microcystis. Biol. Rev. 2018, 93, 1399–1420. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TN (mg/L) | TP (mg/L) | CODMn (mg/L) | pH | SD (cm) | Depth (m) | T (°C) | |
|---|---|---|---|---|---|---|---|
| East | 2.01 ± 1.10 c | 0.10 ± 0.06 c | 4.49 ± 2.29 b | 7.31 ± 0.76 | 12 ± 6.62 | 3.54 ± 0.35 | 17.66 ± 7.84 |
| Central | 2.28 ± 1.03 b | 0.13 ± 0.10 b | 4.67 ± 1.51 b | 7.46 ± 0.83 | 9.75 ± 5.27 | 3.62 ± 0.39 | 17.47 ± 7.79 |
| West | 4.11 ± 3.87 a | 0.27 ± 0.34 a | 6.60 ± 6.99 a | 7.56 ± 0.90 | 13.04 ± 7.77 | 3.25 ± 0.41 | 17.62 ± 7.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, Y.; Zhang, M.; Yang, Z.; Shi, X.; Yu, Y.; Shi, L. Seasonal Shifts in the Morphological Traits of Bloom-Forming Cyanobacteria in Lake Chaohu, China. Diversity 2022, 14, 435. https://doi.org/10.3390/d14060435
Meng Y, Zhang M, Yang Z, Shi X, Yu Y, Shi L. Seasonal Shifts in the Morphological Traits of Bloom-Forming Cyanobacteria in Lake Chaohu, China. Diversity. 2022; 14(6):435. https://doi.org/10.3390/d14060435
Chicago/Turabian StyleMeng, Yangyang, Min Zhang, Zhen Yang, Xiaoli Shi, Yang Yu, and Limei Shi. 2022. "Seasonal Shifts in the Morphological Traits of Bloom-Forming Cyanobacteria in Lake Chaohu, China" Diversity 14, no. 6: 435. https://doi.org/10.3390/d14060435
APA StyleMeng, Y., Zhang, M., Yang, Z., Shi, X., Yu, Y., & Shi, L. (2022). Seasonal Shifts in the Morphological Traits of Bloom-Forming Cyanobacteria in Lake Chaohu, China. Diversity, 14(6), 435. https://doi.org/10.3390/d14060435