
Effects of Sediment Types on the Distribution and Diversity of Plant Communities in the Poyang Lake Wetlands

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
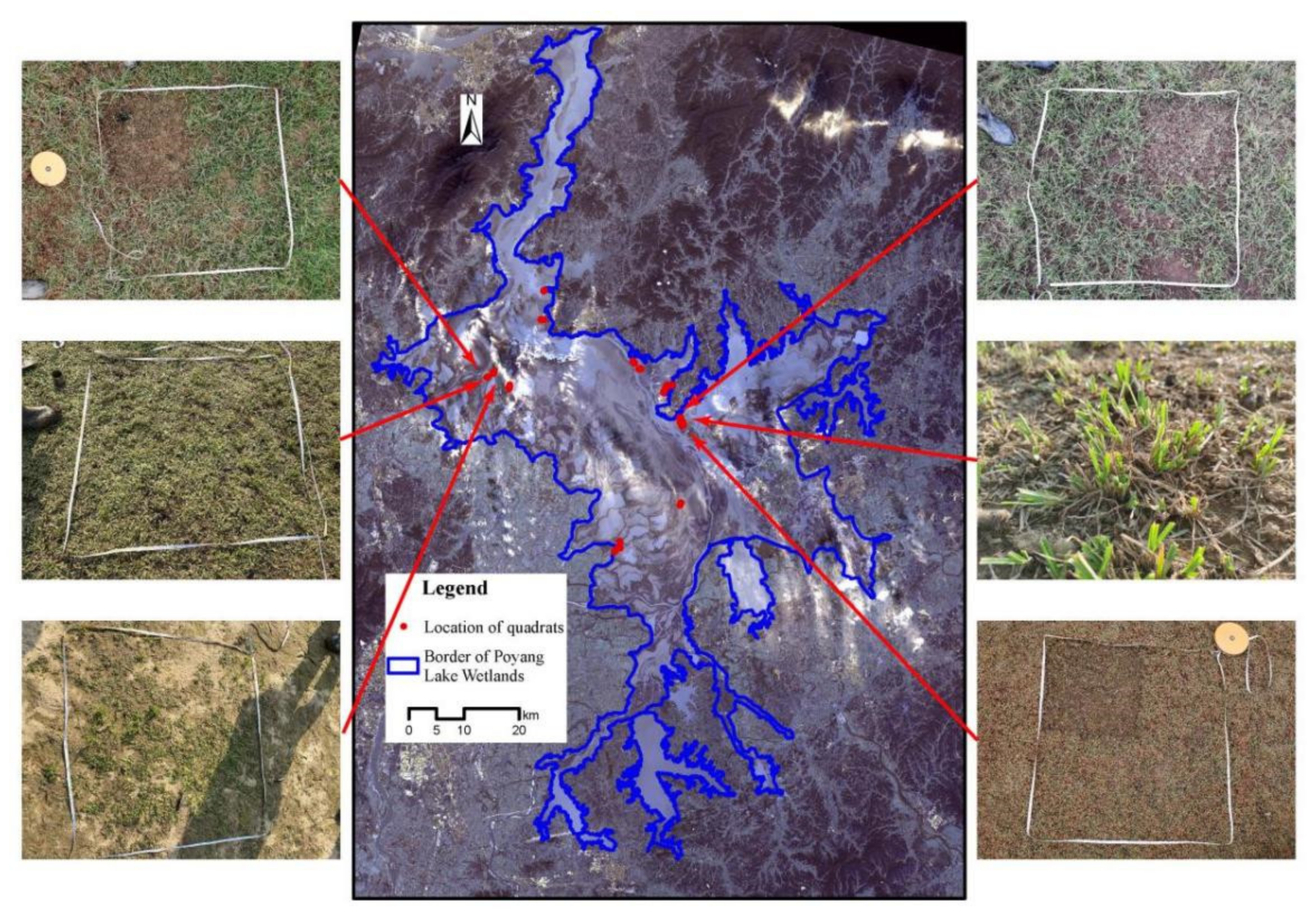
2.1. Study Area
2.2. Field Investigation
2.3. Environmental Data Collection
2.4. Data Analysis
3. Results
3.1. Species Compositions of Plant Communities in the Wetlands
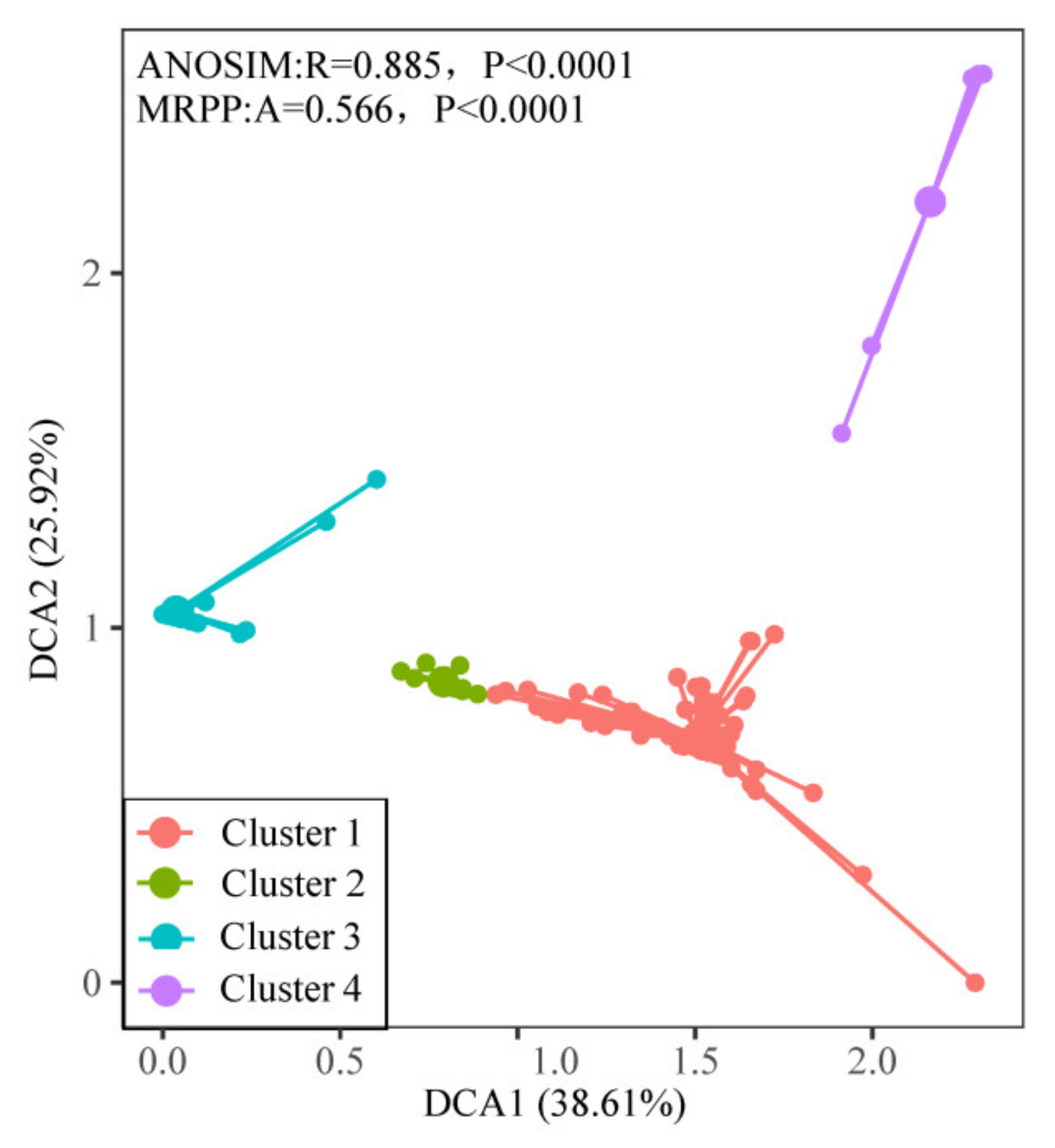
3.2. Quantitative Classification of Plant Communities
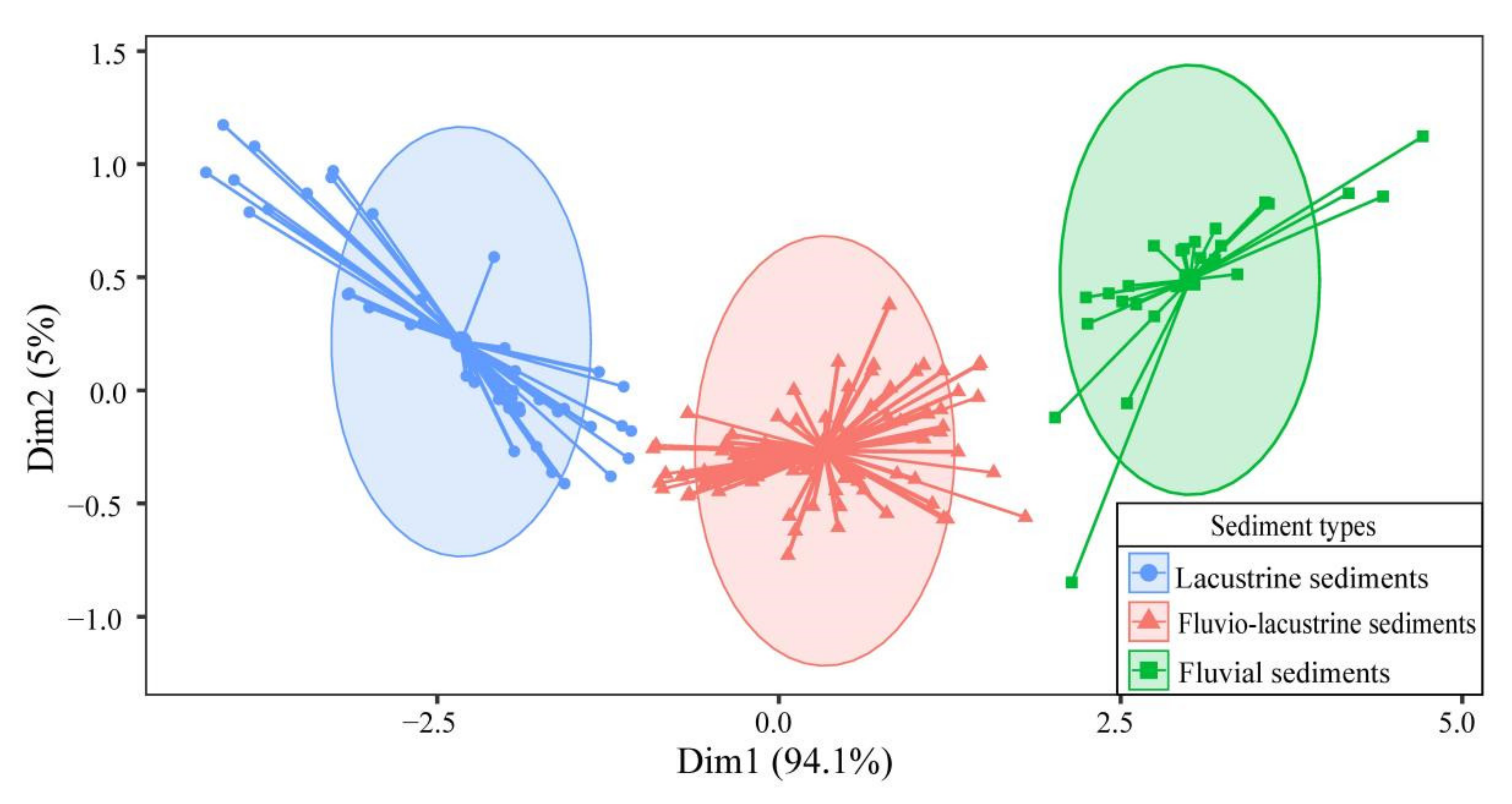
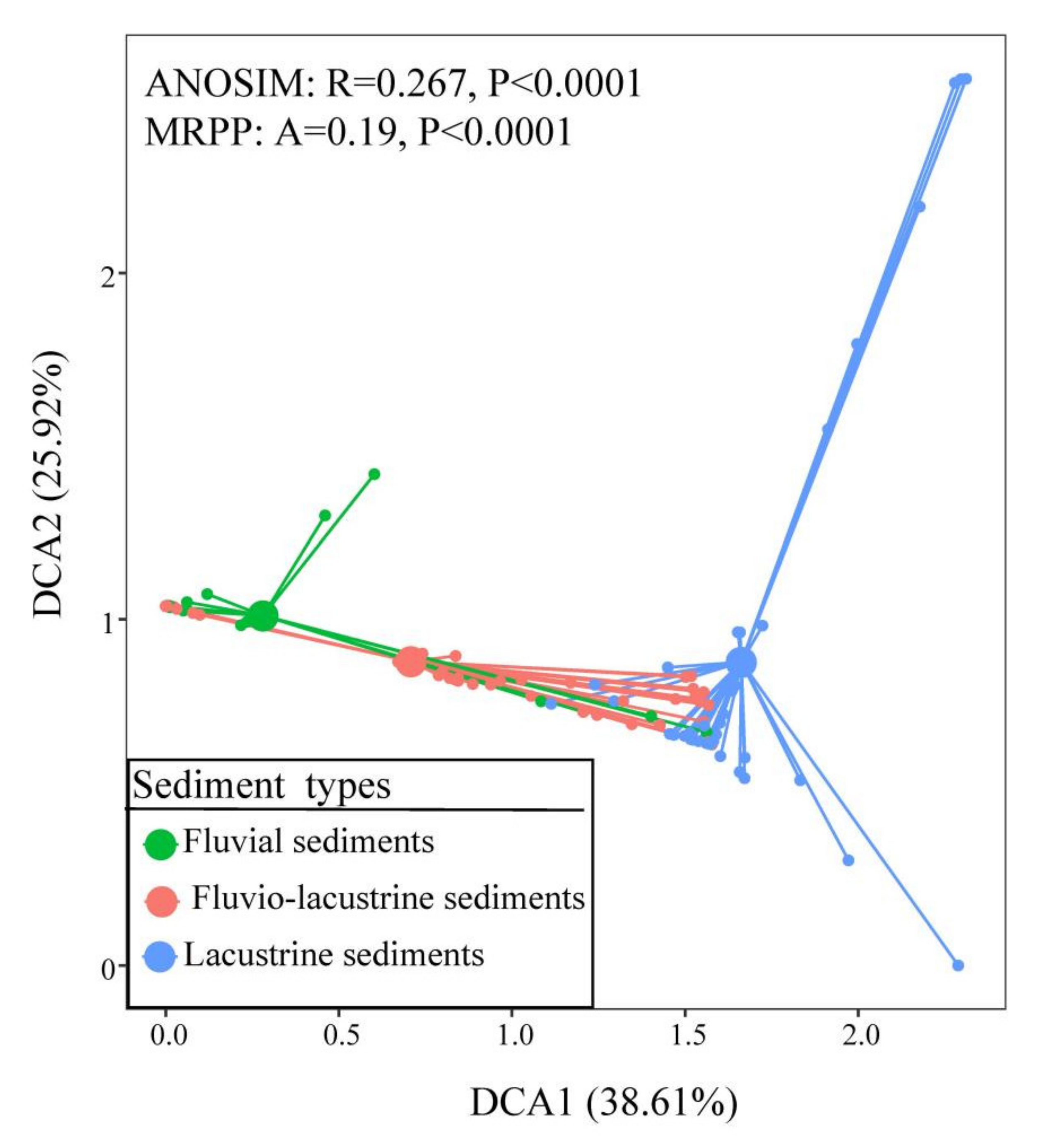
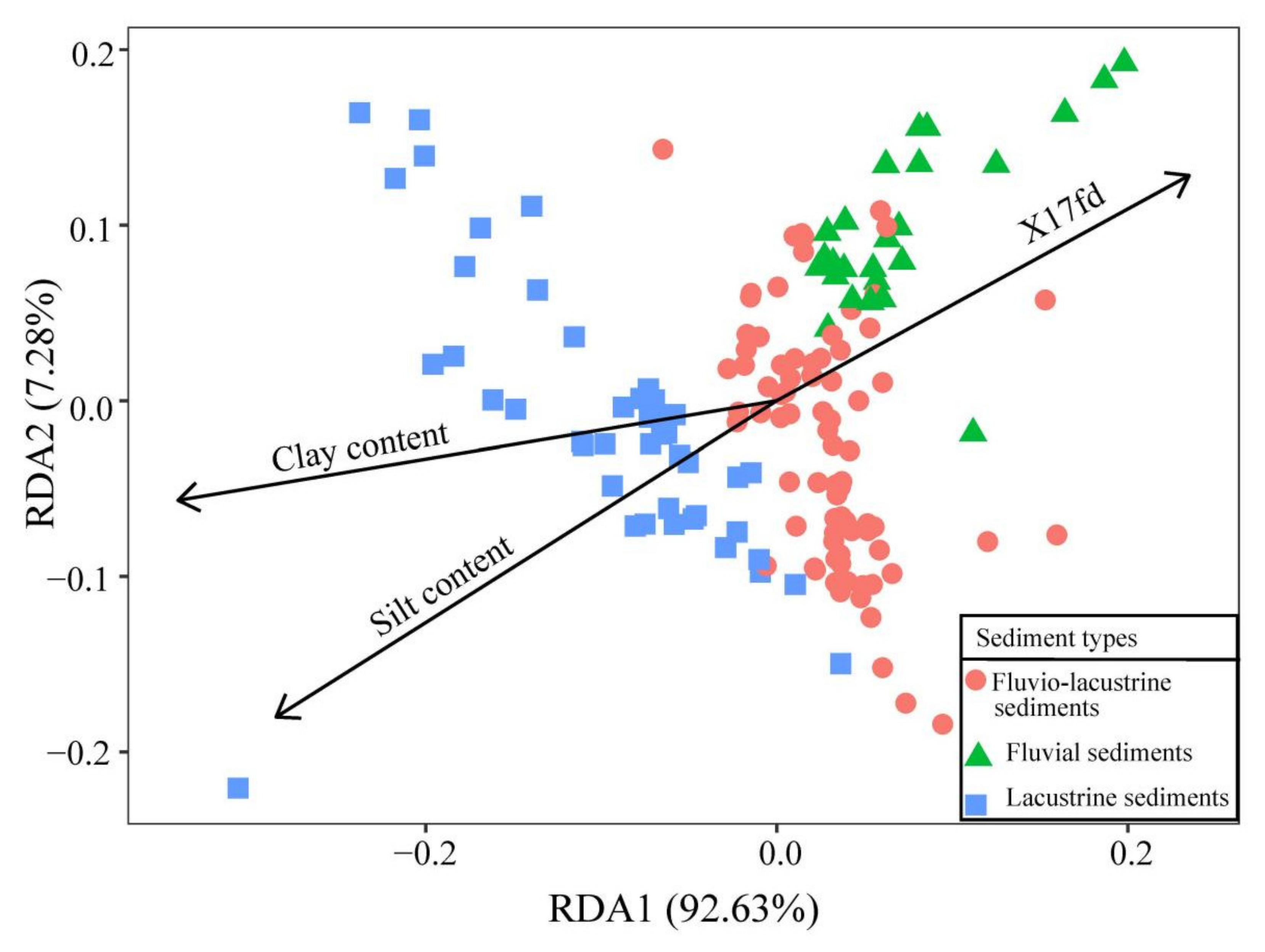
3.3. Sediment Types and Plant Community Distribution
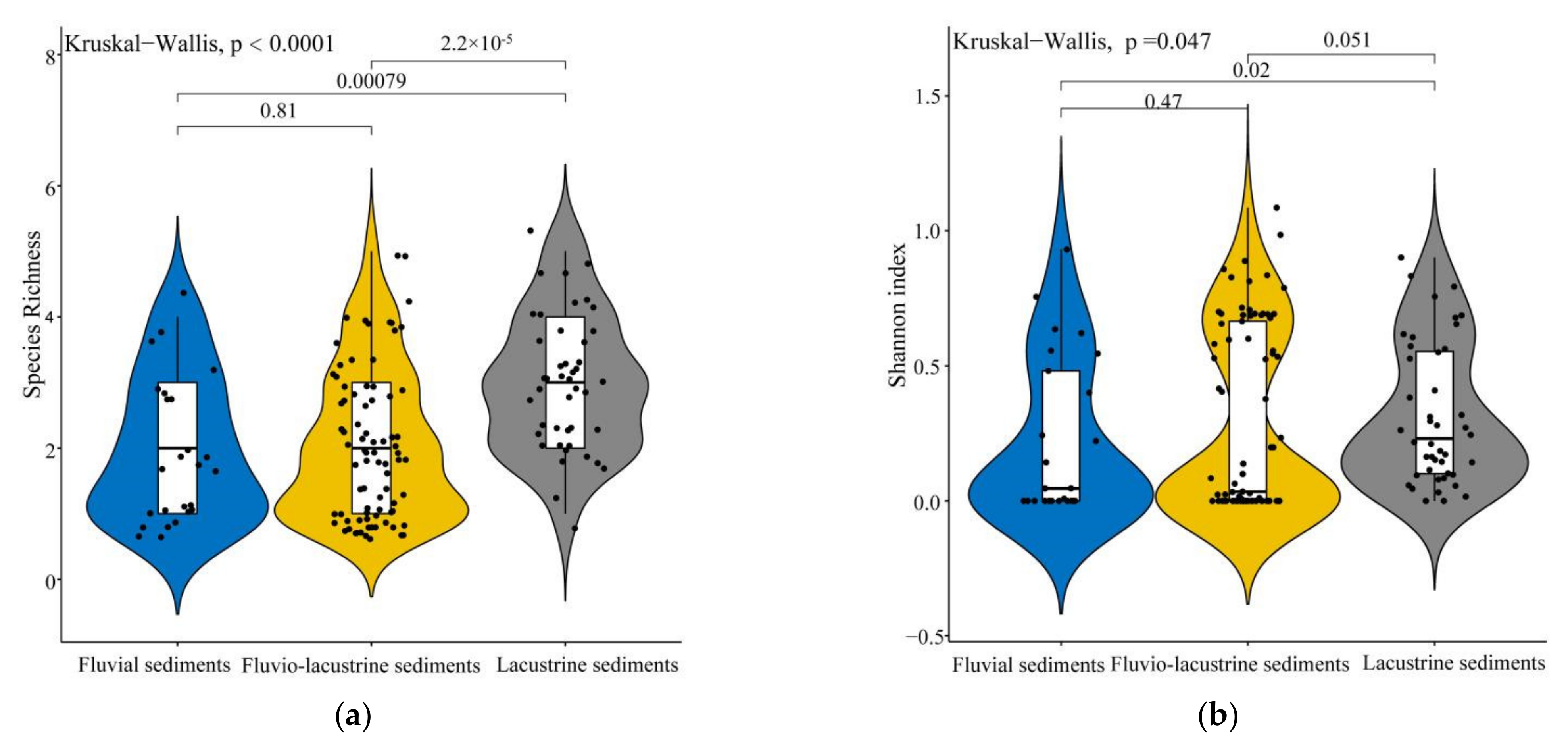
3.4. Comparison of Plant Species Diversity among Three Sediment Types
4. Discussion
4.1. The Distribution of Wetland Plants Differs among Sedimentary Deposition Types along the Beaches of Poyang Lake
4.2. Species Diversity of Wetland Plant Communities Differs among Sediment Types in the Poyang Lake Wetlands
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De Wilde, M.; Puijalon, S.; Bornette, G. Sediment type rules the response of aquatic plant communities to dewatering in wetlands. J. Veg. Sci. 2017, 28, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Huai, W.X.; Li, S.L.; Katul, G.G.; Liu, M.Y.; Yang, Z.H. Flow dynamics and sediment transport in vegetated rivers: A review. J. Hydrodyn. 2021, 33, 400–420. [Google Scholar] [CrossRef]
- Yu, H.; Wang, L.; Liu, C.; Yu, D.; Qu, J. Effects of a spatially heterogeneous nutrient distribution on the growth of clonal wetland plants. BMC Ecol. 2020, 20, 59. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Zheng, S.S.; Wang, P.F.; Hou, J. Interactions between vegetation, water flow and sediment transport: A review. J. Hydrodyn. 2015, 27, 24–37. [Google Scholar] [CrossRef]
- Zheng, P.R.; Li, C.H.; Ye, C.; Wang, H.; Wei, W.W.; Zheng, Y.; Zheng, X.Y. Characteristic and affecting factors of wetland herbs’ distribution in the radiant belt toward land of lake-terrestrial ecotone in Tibet, China. Environ. Sci. Eur. 2022, 34, 14. [Google Scholar] [CrossRef]
- Lo, E.; McGlue, M.; Silva, A.; Bergier, I.; Yeager, K.; Macedo, H.; Swallom, M.; Assine, M. Fluvio-lacustrine sedimentary processes and landforms on the distal Paraguay fluvial megafan (Brazil). Geomorphology 2019, 342, 163–175. [Google Scholar] [CrossRef]
- Fan, H.X.; Xu, L.G.; Wang, X.L.; Jiang, J.H.; Feng, W.J.; You, H.L. Relationship between vegetation community distribution patterns and environmental factors in typical wetlands of Poyang Lake, China. Wetlands 2019, 39, S75–S87. [Google Scholar] [CrossRef]
- Zheng, Z.; Ma, P.F. Changes in above and belowground traits of a rhizome clonal plant explain its predominance under nitrogen addition. Plant Soil 2018, 432, 415–424. [Google Scholar] [CrossRef]
- Guan, B.; Xie, B.H.; Yang, S.S.; Hou, A.X.; Chen, M.; Han, G.X. Effects of five years’ nitrogen deposition on soil properties and plant growth in a salinized reed wetland of the Yellow River Delta. Ecol. Eng. 2019, 136, 160–166. [Google Scholar] [CrossRef]
- Di Luca, G.A.; Maine, M.A.; Mufarrege, M.M.; Hadad, H.R.; Pedro, M.C.; Sanchez, G.C.; Caffaratti, S.E. Phosphorus distribution pattern in sediments of natural and constructed wetlands. Ecol. Eng. 2017, 108, 227–233. [Google Scholar] [CrossRef]
- Liu, Y.; Bachofen, C.; Lou, Y.J.; Ding, Z.; Jiang, M.; Lu, X.G.; Buchmann, N. The effect of temperature changes and K supply on the reproduction and growth of Bolboschoenus planiculmis. J. Plant Ecol. 2021, 14, 337–347. [Google Scholar] [CrossRef]
- Chen, X.S.; Li, X.; Xie, Y.H.; Li, F.; Hou, Z.Y.; Zeng, J. Combined influence of hydrological gradient and edaphic factors on the distribution of macrophyte communities in Dongting Lake wetlands, China. Wetl. Ecol. Manag. 2015, 23, 481–490. [Google Scholar] [CrossRef]
- Ren, G.H.; Deng, B.; Shang, Z.H.; Hou, Y.; Long, R.J. Plant communities and soil variations along a successional gradient in an alpine wetland on the Qinghai-Tibetan Plateau. Ecol. Eng. 2013, 61, 110–116. [Google Scholar] [CrossRef]
- Reza, S.K.; Nayak, D.C.; Chattopadhyay, T.; Mukhopadhyay, S.; Singh, S.K.; Srinivasan, R. Spatial distribution of soil physical properties of alluvial soils: A geostatistical approach. Arch. Agron. Soil Sci. 2016, 62, 972–981. [Google Scholar] [CrossRef]
- Chen, Y.P.; Xia, J.B.; Zhao, X.M.; Zhuge, Y.P. Soil moisture ecological characteristics of typical shrub and grass vegetation on Shell Island in the Yellow River Delta, China. Geoderma 2019, 348, 45–53. [Google Scholar] [CrossRef]
- Weiher, E.; Keddy, P.A. The Assembly of Experimental Wetland Plant Communities. Oikos 1995, 73, 323–335. [Google Scholar] [CrossRef]
- Gao, J.H.; Jia, J.; Kettner, A.J.; Xing, F.; Wang, Y.P.; Xu, X.N.; Yang, Y.; Zou, X.Q.; Gao, S.; Qi, S.; et al. Changes in water and sediment exchange between the Changjiang River and Poyang Lake under natural and anthropogenic conditions, China. Sci. Total Environ. 2014, 481, 542–553. [Google Scholar] [CrossRef]
- Ye, X.; Li, Y.; Li, X.; Zhang, Q. Factors influencing water level changes in China’s largest freshwater lake, Poyang Lake, in the past 50 years. Water Int. 2014, 39, 983–999. [Google Scholar] [CrossRef]
- Dai, Z.J.; Mei, X.F.; Darby, S.E.; Lou, Y.Y.; Li, W.H. Fluvial sediment transfer in the Changjiang (Yangtze) river-estuary depositional system. J. Hydrol. 2018, 566, 719–734. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Wan, R.R.; Yang, G.S.; Wang, X.L.; Xu, L.G.; Li, Y.Y.; Li, B. Impact of seasonal water-level fluctuations on autumn vegetation in Poyang Lake wetland, China. Front. Earth Sci. 2019, 13, 398–409. [Google Scholar] [CrossRef]
- Hu, Z.; Ge, G.; Liu, C.; Chen, F.; Li, S. Structure of Poyang Lake wetland plants ecosystem and influence of lake water level for the structure. Resour. Environ. Yangtze Basin 2010, 19, 597–605. [Google Scholar]
- Fu, H.; Zhong, J.Y.; Yuan, G.X.; Guo, C.J.; Ding, H.J.; Feng, Q.; Fu, Q. A functional-trait approach reveals community diversity and assembly processes responses to flood disturbance in a subtropical wetland. Ecol. Res. 2015, 30, 57–66. [Google Scholar] [CrossRef]
- Qin, T.; Guan, Y.-T.; Zhang, M.-X.; Li, H.-L.; Yu, F.-H. Sediment type and nitrogen deposition affect the relationship between Alternanthera philoxeroides and experimental wetland plant communities. Mar. Freshw. Res. 2018, 69, 811. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, L.-B.; Qu, X.-D.; Kong, W.-J.; Meng, W. Environmental Factors and Community Characteristics of Aquatic Macrophytes in Taizi River Tributaries of Liaoning Province. Plant Sci. J. 2011, 29, 552–560. [Google Scholar] [CrossRef]
- Feng, W.J.; Mariotte, P.; Xu, L.G.; Buttler, A.; Bragazza, L.; Jiang, J.H.; Santonja, M. Seasonal variability of groundwater level effects on the growth of Carex cinerascens in lake wetlands. Ecol. Evol. 2020, 10, 517–526. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.; Ye, X.C.; Werner, A.D.; Li, Y.L.; Yao, J.; Li, X.H.; Xu, C.Y. An investigation of enhanced recessions in Poyang Lake: Comparison of Yangtze River and local catchment impacts. J. Hydrol. 2014, 517, 425–434. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Wan, R.R.; Yang, G.S. Non-stationary water-level fluctuation in China’s Poyang Lake and its interactions with Yangtze River. J. Geogr. Sci. 2015, 25, 274–288. [Google Scholar] [CrossRef] [Green Version]
- Fang, J.; Wang, X.; Shen, Z.; Tang, Z.; He, J.; Yu, D.; Jiang, Y.; Wang, Z.; Zheng, C.; Zhu, J.; et al. Methods and protocols for plant community inventory. Biodivers. Sci. 2009, 17, 533–548. [Google Scholar] [CrossRef]
- Flora of China. Available online: http://www.iplant.cn/foc/ (accessed on 5 March 2021).
- Ye, C. Contrasting Investigation by 137Cs Method and 210Pb Method for the Present Sedimentation Rate of Poyang Lake, Jiangxi. Acta Sedimentol. Sin. 1991, 9, 106. [Google Scholar]
- Barman, U.; Choudhury, R.D. Soil texture classification using multi class support vector machine. Inf. Process. Agric. 2020, 7, 318–332. [Google Scholar] [CrossRef]
- Di Biase, L.; Pace, L.; Mantoni, C.; Fattorini, S. Variations in Plant Richness, Biogeographical Composition, and Life Forms along an Elevational Gradient in a Mediterranean Mountain. Plants 2021, 10, 2090. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Xie, Y.-H.; Chen, X.-S.; Li, F. Adaptation of wetland plants to sedimentation stress: A review. Shengtaixue Zazhi 2011, 30, 155–161. [Google Scholar]
- Murtagh, F.; Legendre, P. Ward’s Hierarchical Agglomerative Clustering Method: Which Algorithms Implement Ward’s Criterion? J. Classif. 2014, 31, 274–295. [Google Scholar] [CrossRef] [Green Version]
- Hill, M.O.; Gauch, H.G. Detrended Correspondence Analysis: An Improved Ordination Technique. Vegetatio 1980, 42, 47–58. [Google Scholar] [CrossRef]
- Shaltout, K.H.; Eid, E.M.; Al-Sodany, Y.M.; Heneidy, S.Z.; Shaltout, S.K.; El-Masry, S.A. Effect of Protection of Mountainous Vegetation against Over-Grazing and Over-Cutting in South Sinai, Egypt. Diversity 2021, 13, 113. [Google Scholar] [CrossRef]
- Han, W.J.; Cao, J.Y.; Liu, J.L.; Jiang, J.; Ni, J. Impacts of nitrogen deposition on terrestrial plant diversity: A meta-analysis in China. J. Plant Ecol. 2019, 12, 1025–1033. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012; Available online: https://www.R-project.org/ (accessed on 8 April 2021).
- Oksanen, J. Vegan Community Ecology Package Version 2.6-2. 2021. Available online: https://CRAN.R-project.org/package=vegan (accessed on 8 April 2021).
- Pavoine, S. Adiv: Analysis of Diversity. R Package Version 2.1.1. 2021. Available online: https://CRAN.R-project.org/package=adiv (accessed on 20 April 2021).
- Giraudoux, P.; Antonietti, J.P.; Beale, C.; Lancelot, R.; Pleydell, D.; Treglia, M. Pgirmess: Spatial Analysis and Data Mining for Field Ecologists. R Package Version 1.7.0. 2021. Available online: https://CRAN.R-project.org-/package=pgirmess (accessed on 22 April 2021).
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer International Publishing AG: Houston, TX, USA, 2016. [Google Scholar]
- Gan, J.; Wan, S.; Li, J.; Tang, C.; Yang, T.; Luozang, Q. Classification of lacustrine soft soil based on sedimentary environment and engineering characteristics—A case study of four typical soft soil in Poyang Lake basin. Geol. Rev. 2019, 65, 983–992. [Google Scholar] [CrossRef]
- Bracken, L.J.; Turnbull, L.; Wainwright, J.; Bogaart, P. Sediment connectivity: A framework for understanding sediment transfer at multiple scales. Earth Surf. Processes Landf. 2015, 40, 177–188. [Google Scholar] [CrossRef] [Green Version]
- Selim, T.; Hesham, M.; Elkiki, M. Effect of sediment transport on flow characteristics in non-prismatic compound channels. Ain Shams Eng. J. 2022, 13, 101771. [Google Scholar] [CrossRef]
- Juma, N.G. Interrelationships between soil structure/texture, soil biota/soil organic matter and crop production. Geoderma 1993, 57, 3–30. [Google Scholar] [CrossRef]
- Smaill, S.J.; Clinton, P.W.; Allen, R.B.; Leckie, A.C.; Davis, M.R. Coarse soil can enhance the availability of nutrients from fine soil. J. Plant Nutr. Soil Sci. 2014, 177, 848–850. [Google Scholar] [CrossRef]
- Mobilian, C.; Craft, C.B. Wetland Soils: Physical and Chemical Properties and Biogeochemical Processes. Ref. Modul. Earth Syst. Environ. Sci. 2021, 3, 157–168. [Google Scholar] [CrossRef]
- Ren, B. Analysis on the Typical Phytocoenoses and Its Principle Factors of Distribution Paterns in East Dongting Lake Wetland. Ph.D. Thesis, Hunan Agricultural University, Changsha, China, 2012. [Google Scholar]
- Zhou, D.M.; Luan, Z.Q.; Guo, X.Y.; Lou, Y.J. Spatial distribution patterns of wetland plants in relation to environmental gradient in the Honghe National Nature Reserve, Northeast China. J. Geogr. Sci. 2012, 22, 57–70. [Google Scholar] [CrossRef]
- Ballantine, K.; Schneider, R.; Groffman, P.; Lehmann, J. Soil Properties and Vegetative Development in Four Restored Freshwater Depressional Wetlands. Soil Sci. Soc. Am. J. 2012, 76, 1482. [Google Scholar] [CrossRef] [Green Version]
- Nyarko, B.K. Wetland river flow interaction in a sedimentary formation of the white Volta basin, Ghana. Earth Sci. Res. 2019, 9, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Feng, W.J.; Santonja, M.; Bragazza, L.; Buttler, A. Shift in plant-soil interactions along a lakeshore hydrological gradient. Sci. Total Environ. 2020, 742, 140254. [Google Scholar] [CrossRef]
- Castelli, R.M.; Chambers, J.C.; Tausch, R.J. Soil-plant relations along a soil-water gradient in great basin riparian meadows. Wetlands 2000, 20, 251–266. [Google Scholar] [CrossRef]
- Mougi, A.; Kondoh, M. Diversity of Interaction Types and Ecological Community Stability. Science 2012, 337, 349–351. [Google Scholar] [CrossRef]
- Ram, J.; Singh, J.S.; Singh, S.P. Plant biomass, species diversity and net primary production in a central Himalayan high altitude grassland. J. Ecol. 1989, 77, 456–468. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the Existence of Three Primary Strategies in Plants and Its Relevance to Ecological and Evolutionary Theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Rodrigues, P.M.S.; Schaefer, C.E.G.R.; de Oliveira Silva, J.; Ferreira Júnior, W.G.; dos Santos, R.M.; Neri, A.V. The influence of soil on vegetation structure and plant diversity in different tropical savannic and forest habitats. J. Plant Ecol. 2018, 11, 226–236. [Google Scholar] [CrossRef]
- Papadopoulos, A. Soil Aggregates, Structure, and Stability. In Encyclopedia of Agrophysics; Gliński, J., Horabik, J., Lipiec, J., Eds.; Springer: Dordrecht, the Netherlands, 2011; pp. 736–740. [Google Scholar]
- Stirzaker, R.J.; Passioura, J.B.; Wilms, Y. Soil structure and plant growth: Impact of bulk density and biopores. Plant Soil 1996, 185, 151–162. [Google Scholar] [CrossRef]
- Li, Y.; Wang, D.; Xin, Z. Spatial distribution of vegetation and soil in aquatic-terrestrial ecotone, Li River. Trans. Chin. Soc. Agric. Eng. 2013, 29, 121–128. [Google Scholar] [CrossRef]
- Limberger, R.; Wickham, S.A. Disturbance and diversity at two spatial scales. Oecologia 2012, 168, 785–795. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S/N | Species | Family | Life Form | Sediment-Tolerant Types |
|---|---|---|---|---|
| 1 | Cardamine lyrata | Brassicaceae | Cryptophyte | Sediment-dependence |
| 2 | Cardamine impatiens | Brassicaceae | Therophyte | Sediment-dependence |
| 3 | Heleocharis valleculosa | Cyperaceae | Cryptophyte | Sediment-dependence |
| 4 | Carex cinerascens | Cyperaceae | Cryptophyte | Sediment-tolerance |
| 5 | Phalaris arundinacea | Gramineae | Cryptophyte | Sediment-tolerance |
| 6 | Potentilla limprichtii | Rosaceae | Chamaephyte | Sediment-tolerance |
| 7 | Polygonum pubescens | Polygonacae | Therophyte | Sediment-tolerance |
| 8 | Polygonum criopolitanum | Polygonacae | Therophyte | Sediment-tolerance |
| 9 | Rumex acetosa | Polygonacae | Chamaephyte | Sediment-tolerance |
| 10 | Rumex acetosella | Polygonacae | Chamaephyte | Sediment-tolerance |
| 11 | Artemisia selengensis | Asteraceae | Cryptophyte | Sediment-dependence |
| 12 | Lapsana apogonoides | Asteraceae | Therophyte | Sediment-sensitivity |
| 13 | Gnaphalium affine | Asteraceae | Therophyte | Sediment-sensitivity |
| 14 | Hemarthria altissima | Gramineae | Hemicryptophyta | Sediment-sensitivity |
| 15 | Kalimeris indica | Asteraceae | Cryptophyte | Sediment-dependence |
| 16 | Ranunculus polii | Ranunculaceae | Therophyte | Sediment-dependence |
| RDA Axes | RDA1 | RDA2 | RDA3 |
|---|---|---|---|
| Silt content | −0.71 | −0.45 | 0.53 |
| Clay content | −0.85 | −0.14 | 0.50 |
| X17fd | 0.58 | 0.32 | 0.74 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Liu, Y.; Liu, Y.; Guo, H.; Chen, G.; Fu, Z.; Fu, Y.; Ge, G. Effects of Sediment Types on the Distribution and Diversity of Plant Communities in the Poyang Lake Wetlands. Diversity 2022, 14, 491. https://doi.org/10.3390/d14060491
Li J, Liu Y, Liu Y, Guo H, Chen G, Fu Z, Fu Y, Ge G. Effects of Sediment Types on the Distribution and Diversity of Plant Communities in the Poyang Lake Wetlands. Diversity. 2022; 14(6):491. https://doi.org/10.3390/d14060491
Chicago/Turabian StyleLi, Jie, Yizhen Liu, Ying Liu, Huicai Guo, Gang Chen, Zhuoting Fu, Yvying Fu, and Gang Ge. 2022. "Effects of Sediment Types on the Distribution and Diversity of Plant Communities in the Poyang Lake Wetlands" Diversity 14, no. 6: 491. https://doi.org/10.3390/d14060491
APA StyleLi, J., Liu, Y., Liu, Y., Guo, H., Chen, G., Fu, Z., Fu, Y., & Ge, G. (2022). Effects of Sediment Types on the Distribution and Diversity of Plant Communities in the Poyang Lake Wetlands. Diversity, 14(6), 491. https://doi.org/10.3390/d14060491