
Assemblages of Meiobenthic and Planktonic Microcrustaceans (Cladocera and Copepoda) from Small Water Bodies of Mountain Subarctic (Putorana Plateau, Middle Siberia)


 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
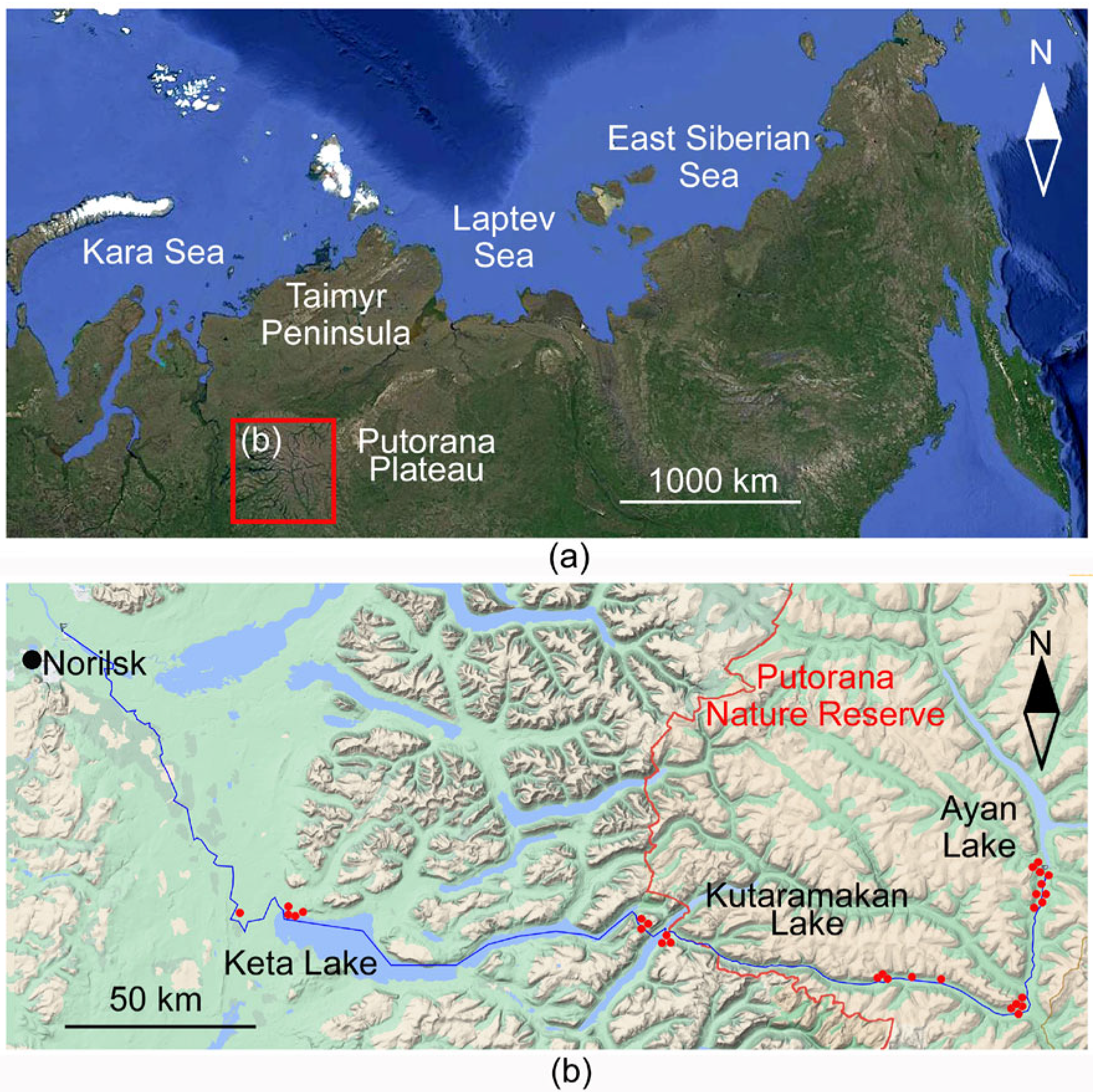
2.1. Studied Area
2.2. Types of Waterbodies
2.3. Sampling
2.4. Literature Data
2.5. Statistical Analysis
3. Results
3.1. Fauna Composition and Species Richness
3.2. Patterns in Species Richness and Assemblage Structure of Meiobenthic and Planktonic Crustacean
3.3. Characteristic and Differentiating Taxons
4. Discussion
4.1. Specificity of the Regional Fauna
4.2. New Records for the Region
4.3. Crustacean Assemblage Structure and Regulating Factors
4.4. Biogeographical Position of the Putoran Plateau Fauna
5. Conclusions
- In the present study, 81 crustacean species were found in the water bodies of the Putorana Plateau: 45 Copepoda, 36 Cladocera. Two species of Cladocera and 21 species for Copepoda are new for the region. The list of known fauna of the region was increased by 22% of the previously known.
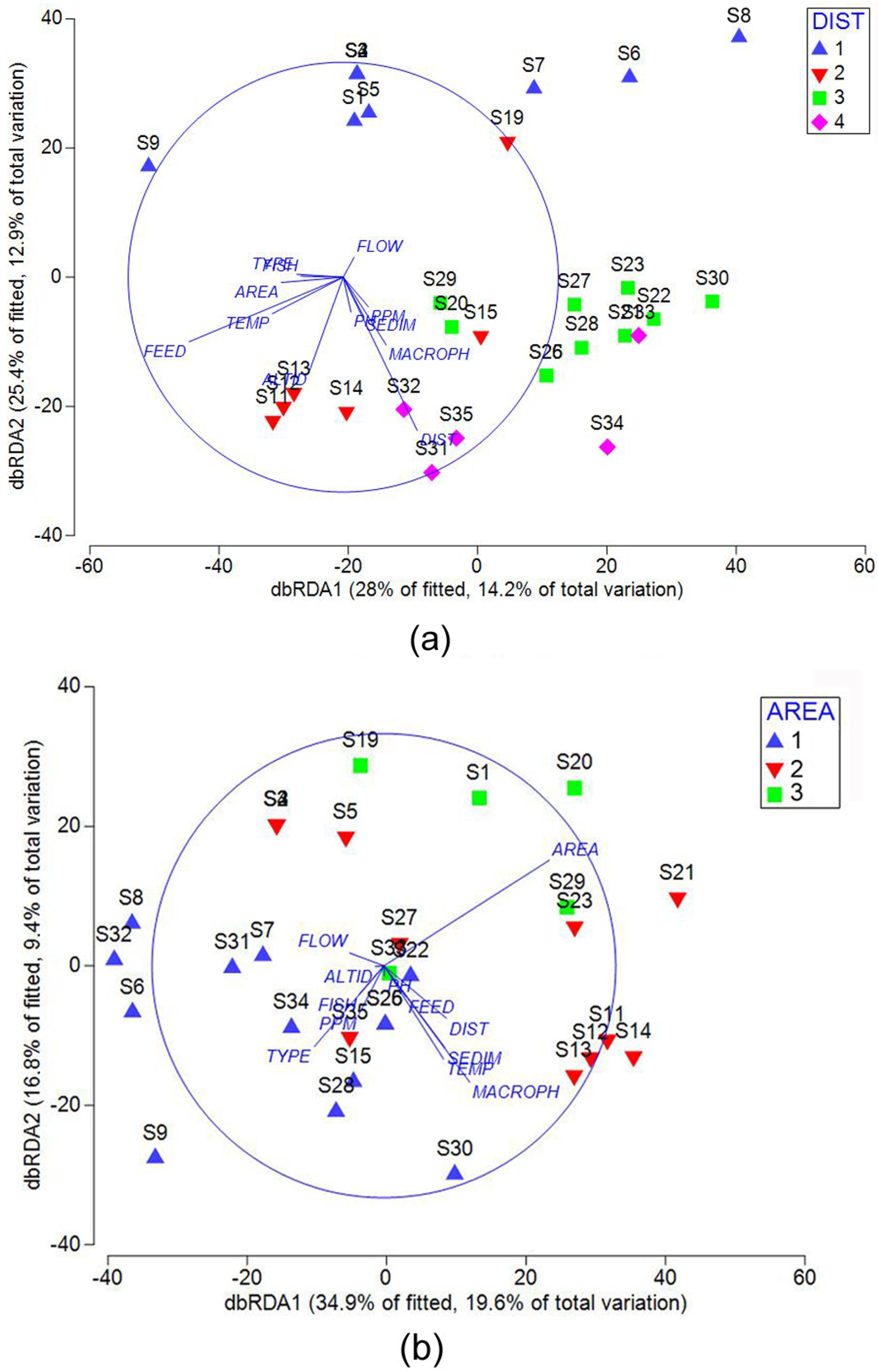
- The species richness of Copepoda (especially the meiobenthic Harpacticoida) is significantly higher in the central part and on the western slopes of the Putorana Plateau than in the foothills. The increase in the species richness of meiofauna in mountainous areas is associated with the high role of eventuality of species introducing into water bodies, which makes it possible to form different species composition and implement a different structure of assemblages in water bodies of the same region.
- Variations in the meiobenthic crustacean assemblage structure are due to the research district, type of water supply and, to a lesser degree, altitude above the sea level. The structure of planktonic crustacean assemblages was generally determined by the area size of the water body and less affected by the macrophytes composition.
- The total microcrustacean fauna of the Putorana Plateau consists of species belonging to different biogeographic faunistic complexes: 18.5% of fauna are cosmopolites; 42.7%—widely Palearctic; 17.7%—Subarctic and Arctic; 8.1%—Holarctic; 2.4%—Eastern Asian–North American; 4.8%—Eastern Asian; 2.4%—West Palearctic; 3.2%—endemics. About 12.8% of the species are relics that survived the last ice age in the reservoirs of the Putorana Plateau.
- The fauna of the northern part of Middle Siberia, in general, and the Putorana Plateau, in particular, is characterized by high species richness and significantly distinguishes from the fauna of both western and eastern regions of the Arctic. The specifics of faunal composition are connected to the climatic features of Middle Siberia and the preservation of Pleistocene fauna on its territory in refugia not subjected to the last glaciation.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon | District (Medial Altitude m a.s.l.) | Faunistic Complex | ||
|---|---|---|---|---|
| Central Part (492 m) | Western Slopes (347 m) | Foothills (84 m) | ||
| Class Branchiopoda | ||||
| Order Anostraca | ||||
| Family Chirocephalidae | ||||
| Polyartemia forcipata Fischer, 1851 | + | ARC (P) | ||
| Subclass Cladocera | ||||
| Order Anomopoda | ||||
| Family Bosminidae | ||||
| Bosmina (Eubosmina) coregoni Baird, 1857 | + | + | wP | |
| B. (Eubosmina) cf. longispina Leydig, 1860 | + | + | + | wP |
| Family Eurycercidae | ||||
| Eurycercus (Eurycercus) cf. lamellatus (O.F. Müller, 1776) | + | + | + | wP |
| * E. (E.) pompholygodes Frey, 1975 | + | + | ARC (P) | |
| Family Chydoridae | ||||
| Acroperus harpae (Baird, 1834) | + | + | + | wP |
| Alona guttata Sars, 1862 | + | + | + | C |
| A. intermedia Sars, 1862 | + | wP | ||
| A. quadrangularis (O.F. Müller, 1785) | + | wP | ||
| Alonella excisa (Fischer, 1854) | + | + | + | wP |
| A. exigua (Lilljeborg, 1901) | + | + | C | |
| A. nana (Baird, 1850) | + | C | ||
| Alonopsis elongata (Sars, 1861) | + | + | ARC (P) | |
| Biapertura affinis (Leydig, 1860) | + | + | + | wP |
| * B. sibirica (Sinev, Karabanov et Kotov, 2020) | + | + | + | wP |
| Chydorus cf. sphaericus (O.F. Müller, 1785) | + | + | + | wP |
| Coronatella rectangula (Sars, 1862) | + | + | + | wP |
| Flavalona costata (Sars, 1862) | + | wP | ||
| Graptoleberis testudinaria (Fischer, 1851) | + | wP | ||
| Pleuroxus trigonellus (O.F. Müller, 1785) | + | + | + | wP |
| P. truncatus (O.F. Müller, 1785) | + | + | + | wP |
| Pseudochydorus globosus (Baird, 1843) | + | + | HOL | |
| Family Ophryoxidae | ||||
| Ophryoxus gracilis Sars, 1862 | + | + | + | wP |
| O. kolymensis Smirnov, 1992 | + | + | + | EA-NA |
| Family Daphnidae | ||||
| Daphnia (Daphnia) cristata Sars, 1862 | + | + | wP | |
| D. (Daphnia) cf. dentifera Forbes, 1893 | + | + | + | EA-NA |
| D. (Daphnia) galeata Sars, 1864 | + | C | ||
| D. (Daphnia) longiremis Sars, 1862 | + | HOL | ||
| D. (Daphnia) cf. longispina O.F. Müller, 1776 | + | + | + | wP |
| D. (Daphnia) pulex Leydig, 1860 | + | + | + | C |
| Ceriodaphnia pulchella Sars, 1862 | + | C | ||
| Scapholeberis mucronata (O.F. Müller, 1776) | + | + | + | wP |
| Simocephalus expinosus (De Geer, 1778) | + | C | ||
| S. vetulus (O.F. Müller, 1776) | + | + | + | wP |
| Order Ctenopoda | ||||
| Family Sididae | ||||
| Sida ortiva Korovchinsky, 1979 | + | EA | ||
| Family Holopediidae | ||||
| Holopedium gibberum Zaddach, 1855 | + | + | + | C |
| Order Onychopoda | ||||
| Family Polyphemidae | ||||
| Polyphemus pediculus (Linnaeus, 1761) | + | + | + | wP |
| Subclass Copepoda | ||||
| Order Calanoida | ||||
| Family Temoridae | ||||
| Heterocope appendiculata Sars, 1863 | + | + | + | ARC (P) |
| H. borealis (Fischer, 1851) | + | ARC (P) | ||
| Family Diaptomidae | ||||
| Arctodiaptomus bacillifer (Koelbel, 1885) | + | wP | ||
| Acanthodiaptomus denticornis (Wierzejski, 1887) | + | + | HOL | |
| A. tibetanus (Daday, 1907) | + | EA | ||
| ** Acanthodiaptomus sp. nov. | + | END | ||
| Mixodiaptomus theeli (Lilljeborg in Guerne & Richard, 1889) | + | wP | ||
| ** Mixodiaptomus sp. nov. | + | END | ||
| Order Cyclopoida | ||||
| Family Cyclopidae | ||||
| * Eucyclops cf. arcanus Alekseev, 1990 | + | + | + | EA |
| E. denticulatus (Graeter, 1903) | + | wP | ||
| E. serrulatus (Fischer, 1851) | + | + | + | C |
| * E. speratus (Lilljeborg, 1901) | + | + | C | |
| Macrocyclops albidus (Jurine, 1820) | + | + | + | C |
| Paracyclops fmbriatus (Fischer, 1853) | + | + | + | C |
| Acanthocyclops capillatus (Sars, 1863) | + | + | + | ARC (C) |
| A. cf. robustus (Sars, 1863) | + | + | ARC (C) | |
| A. venustus s. lat. (Norman & T. Scott, 1906) | + | ARC (P) | ||
| A. vernalis (Fischer, 1853) | + | C | ||
| Cyclops scutifer Sars, 1863 | + | + | HOL | |
| * C. sibiricus Lindberg, 1949 | + | EA-NA | ||
| C. strenuus Fischer, 1851 | + | + | C | |
| * Diacyclops bisetosus (Rehberg, 1880) | + | C | ||
| D. nanus (Sars, 1863) | + | + | ARC (C) | |
| Megacyclops viridis (Jurine, 1820) | + | + | + | C |
| Microcyclops bicolor (Sars G.O., 1863) | + | + | C | |
| Order Harpacticoida | ||||
| Family Canthocamptidae | ||||
| * Attheyella northumbrica trisetosa (Chappuis, 1929) | + | + | + | wP |
| * At. nordenskioldii (Lilljeborg, 1902) | + | + | ARC (P) | |
| * Bryocamptus arcticus (Lilljeborg, 1902) | + | + | ARC (P) | |
| * B. krochini (Borutzky, 1951) | + | ARC (P) | ||
| * B. vejdovskyi (Mrázek, 1893) | + | + | HOL | |
| ** Bryocamptus sp. nov. | + | END | ||
| Canthocamptus glacialis Lilljeborg, 1902 | + | ARC (P) | ||
| * Elaphoidella gracilis (Sars, 1863) | + | WP | ||
| Epactophanes richardi Mrázek, 1893 | + | + | C | |
| * Maraenobiotus brucei (Richard, 1898) | + | ARC (P) | ||
| M. insignipes (Lilljeborg, 1902) | + | + | ARC (P) | |
| * Moraria duthiei (T. Scott & A. Scott, 1896) | + | ARC (P) | ||
| M. mrazeki T. Scott, 1903 | + | + | + | ARC (P) |
| * M. cf. mrazeki | + | - | ||
| ** Moraria sp. nov. | + | + | END | |
| Pesceus schmeili (Mrázek, 1893) | + | + | wP | |
| Family Phyllognathopodidae | ||||
| * Phyllognathopus paludosus Mrázek, 1893 | + | HOL | ||
| Family Parastenocaridae | ||||
| * Parastenocaris brevipes Kessler, 1913 | + | + | WP | |
| Family Ectinosomatidae | ||||
| * Pseudobradya arctica (Olofsson, 1917) | + | ARC (C) | ||
| Family Laophontidae | ||||
| * Onychocamptus mochammed (Blanchard & Richard, 1891) | + | C | ||
References
- Hahne, J.; Melles, M. Climate and vegetation history of the Taymyr Peninsula since Middle Weichselian time–palinologycal evidence from lake sediments. In Land-Ocean Systems in the Siberian Arctic. Dynamics and History; Springer: Berlin/Heidelberg, Germany, 1999; pp. 407–423. [Google Scholar] [CrossRef]
- Mikhailov, N.I. Geomorphological observations in the western part of the Putorana Mountains. Vopr. Geogr. 1947, 3, 7–26. (In Russian) [Google Scholar]
- Kirillov, M.V.; Shub, V.A.; Elizaryeva, M.F. Resources of Surface Water of the USSR; Angaro-Yeniseisky District; Muranova, A.P., Ed.; Gidrometeoizdat: Leningrad, Russia, 1973; Volume 16, p. 723. (In Russian) [Google Scholar]
- Bolshiyanov, D.Y.; Antonov, O.M.; Fedorov, G.B.; Pavlov, M.V. Glaciation of the Putorana Plateau during the last glacial maximum. Izv. RGO 2007, 139, 47–61. (In Russian) [Google Scholar]
- Sarana, V.A. Glaciation of the western part of the Putorana Plateau in the late Pleistocene and Holocene. Vest. Mosk. Uuniv. Geogr. 2017, 1, 73–81. [Google Scholar]
- Romanov, A.A. Introduction. In Vertebrate Fauna of the Putorana Plateau; Romanov, A.A., Ed.; Rosselchosakademia: Moskow, Russia, 2004; pp. 9–14. (In Russian) [Google Scholar]
- Vershinin, N.V. On the origin of the relict fauna in Norilsk Lakes Group. Rep. Acad. Sci. USSR 1960, 135, 753–755. (In Russian) [Google Scholar]
- Gerasimova, I.P. Middle Siberia. Natural Conditions and Natural Resources of the USSR; Nauka: Moscow, Russia, 1964; p. 480. (In Russian) [Google Scholar]
- Belych, F.I. Lake Lama and its fishery use. In The Fishing Importance of the Norilsk Lakes (Melkoje and Lama); Suvorov, E.K., Ed.; Chief Administration of the Northern Sea route: Moscow-Leningrad, Russia, 1940; pp. 73–100. (In Russian) [Google Scholar]
- Gordeeva, L.N. Zooplankton of Norilsk Lakes. Hydrobiological studies on reservoirs of Karelia. Sci. N. Karelian State Pedagog. Inst. 1964, 15, 104–116. (In Russian) [Google Scholar]
- Zadelenov, V.A.; Dubovskaya, O.P.; Bazhina, L.V.; Glushchenko, L.A.; Isaeva, I.G.; Kleush, V.O.; Semenchenko, K.A.; Matasov, V.V.; Shadrin, E.N. New data on biota of some lakes in the western part of the Putorana Plateau. J. Sib. Fed. Univ. Biol. 2017, 10, 87–105. [Google Scholar] [CrossRef]
- Basharova, N.I.; Sheveleva, N.G. The main features of the zooplankton formation in the Angara-Yenisei Reservoirs. Hydrobiol. J. 1993, 29, 9–15. (In Russian) [Google Scholar]
- Sheveleva, N.G.; Shishkin, B.A. Zooplankton. In Hydrochemical and Hydrobiological Studies of the Khantai Reservoir; Tyulina, L.N., Ed.; Nauka: Novosibirsk, Russia, 1986; pp. 81–100. (In Russian) [Google Scholar]
- Aleksiuk, A.P.; Shulga, E.L. Some data on the zooplankton of the Putorana Lakes. In Putorana Lake Province: The Results of Landscape-Limnological Studies of 1968; Parmusin, J.P., Tyulina, L.N., Eds.; Nauka: Novosibirsk, Russia, 1975; pp. 194–199. (In Russian) [Google Scholar]
- Sheveleva, N.G. Zooplankton. In Lakes of the North-West of the Siberian Platform; Tyulina, L.N., Ed.; Nauka: Novosibirsk, Russia, 1981; pp. 123–135. (In Russian) [Google Scholar]
- Dubovskaya, O.P.; Kotov, A.A.; Korovchinsky, N.M.; Smirnov, N.N.; Sinev, A.Y. Zooplankton of lakes in the spurs of the Putorana Plateau and adjacent territories (North of Krasnoyarsk Krai). Cont. Probl. Ecol. 2010, 3, 401–434. [Google Scholar] [CrossRef]
- Fefilova, E.; Dubovskaya, O.; Kononova, O.; Khokhlova, L. A comparative survey of the freshwater copepods of two different regions of the Central Palaearctic: European and Siberian. J. Nat. Hist. 2013, 47, 805–819. [Google Scholar] [CrossRef]
- Fefilova, E.; Dubovskaya, O.; Kononova, O.; Frolova, L.; Abramova, E.; Nigamatzyanova, G. Data on taxa composition of freshwater zooplankton and meiobenthos across Arctic regions of Russia. Data Brief. 2021, 36, 107112. [Google Scholar] [CrossRef]
- Sheveleva, N.G. Diversity of planktonic fauna of Putorana Plateau water bodies. In Bird and Animal Communities of the Putorana Plateau: Studies and Conservation; Romanov, A.A., Ed.; Rosselhozakademia Press: Moscow, Russia, 2006; pp. 239–251. (In Russian) [Google Scholar]
- Aanes, R.; Sæther, B.E.; Smith, F.M.; Cooper, E.J.; Wookey, P.A.; Øritsland, N.A. The Arctic oscillation predicts effects of climate change in two trophic levels in a high arctic ecosystem. Ecol. Lett. 2002, 5, 445–453. [Google Scholar] [CrossRef]
- Zemcova, A.I. Weather types. Putorana lake province. Proc. Limnol. Inst. Sib. Branch USSR Acad. Sci. 1975, 20, 19–31. (In Russian) [Google Scholar]
- Pospelov, I.N.; Pospelova, E.B. Vascular Flora of the World Heritage Site «Putorana Plateau» and Its Buffer Zone («Putoransky» Reserve and its Protected Zone); KMK Scientific Press Ltd.: Moskow, Russia, 2021; p. 184. (In Russian) [Google Scholar]
- Alekseev, V.R.; Tsalolikhin, S.Y. Guide of Freshwater Zooplankton and Zoobenthos of European Russia. Zooplankton, 1; KMK Press: Moscow, Russia, 2010; p. 495. (In Russian) [Google Scholar]
- Borutsky, E.V. Crustaceans Freshwater Harpacticoids. In Fauna of USSR, Crustacea 3; AN USSR: Moscow-Leningrad, Russia, 1952; p. 425. (In Russian) [Google Scholar]
- Borutsky, E.V.; Stepanova, L.A.; Kos, M.S. Key to Identification of Calanoida from Fresh Waters; Nauka: St. Petersburg, Russia, 1991; p. 504. (In Russian) [Google Scholar]
- Dussart, B.H.; Defaye, D. Répertoire mondial des Crustacés Copépodes des Eaux Intérieures. Calanoïdes; CNRS Bordeaux: Paris, France, 1983; p. 224. [Google Scholar]
- Fefilova, E.B. Copepods (Copepoda). Fauna of the European North-East of Russia; KMK Scientific Press: Moscow, Russia, 2015; p. 319. [Google Scholar]
- Krivenkova, I.F.; Pen’kova, O.G.; Makarkina, N.V.; Sheveleva, N.G. Acanthodiaptomus tibetanus (Daday, 1908) (Calanoida, Diaptomidae)—A biogeographic relict in the water bodies of Baikal Rift Zone. IOP Conf. Ser. Earth Environ. Sci. 2022, 962, 012021. [Google Scholar] [CrossRef]
- Korovchinsky, N.M.; Kotov, A.A.; Sinev, A.Y.; Neretina, A.N.; Garibian, P.G. Cladocera (Crustacea) of North Eurasia; MK Scientific Press Ltd.: Moskow, Russia, 2021; Volume 2, p. 544. (In Russian) [Google Scholar]
- Lieder, U. Crustacea: Cladocera/Bosminidae. In Süsswasserfauna von Mitteleuropa; Fischer Verlag: Stuttgart, Germany, 1996; p. 80. [Google Scholar]
- Sinev, A.Y. A key to identifying cladocerans of the genus Alona (Anomopoda, Chydoridae) from the Russian European part and Siberia. Russ. J. Zool. 2002, 81, 926–939. [Google Scholar]
- Smirnov, N.N. Chydoridae of the world fauna. In Fauna SSSR. Rakoobraznye (Crustacea); Smirnov, N.N., Ed.; Nauka: Leningrad, Russia, 1971; p. 529. (In Russian) [Google Scholar]
- Novichkova, A.A.; Azovsky, A.I. Factors affecting regional diversity and distribution of freshwater microcrustaceans (Cladocera, Copepoda) at high latitudes. Pol. Biol. 2017, 40, 185–198. [Google Scholar] [CrossRef]
- Abramova, E.; Vishnyakova, I.; Boike, J.; Abramova, A.; Solovyev, G.; Martynov, F. Structure of freshwater zooplankton communities from tundra waterbodies in the Lena River Delta, Russian Arctic, with a discussion on new records of glacial relict copepods. Pol. Biol. 2017, 40, 1629–1643. [Google Scholar] [CrossRef]
- Chertoprud, E.S.; Novichkova, A.A. Crustaceans in the Meiobenthos and Plankton of the Thermokarst Lakes and Polygonal Ponds in the Lena River Delta (Northern Yakutia, Russia): Species Composition and Factors Regulating Assemblage Structures. Water 2021, 13, 1936. [Google Scholar] [CrossRef]
- Frolova, L.A.; Nigamatzayanova, G.R. Structural and functional characteristics of zooplankton communities of thermokarst lakes of Somoilovskii island (Lena Delta, Sakha Republic (Yakutia)). Uchenye Zap. Kazan. Univ. 2019, 1, 158–172. [Google Scholar] [CrossRef]
- Novichkova, A.A.; Chertoprud, E.S. Fauna of microcrustaceans (Cladocera: Copepoda) of shallow freshwater ecosystems of Wrangel Island (Russian Far East). J. Nat. Hist. 2015, 49, 2955–2968. [Google Scholar] [CrossRef]
- Novichkova, A.A.; Chertoprud, E.S. Cladocera and Copepoda of Shokalsky Island: New data from northwest Siberia. J. Nat. Hist. 2017, 51, 1781–1793. [Google Scholar] [CrossRef]
- Streletskaya, E.A. List of the investigated rotifers, cladocerans and copepods of the water bodies of the Kolyma and Anadyr’ River basins. In Hydrobiological Studies of the Inland Water Bodies in the North-East USSR; Vostok: Vladivostok, Russia, 1975; pp. 32–59. (In Russian) [Google Scholar]
- Streletskaya, E.A. To the question on the systematics of some freshwater crustaceans (Cladocera, Copepoda) of the Kolyma River basin. In Hydrobiological Studies of the Inland Water Bodies in the North-East USSR; Vostok: Vladivostok, Russia, 1975; pp. 60–195. (In Russian) [Google Scholar]
- Streletskaya, E.A. Review of the Fauna of Rotatoria, Cladocera, and Copepoda of the Basin of the Anadyr’ River. Contemp. Prob. Ecol. 2010, 3, 469–480. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R.N. Primer, User Manual/Tutorial, 5th ed.; PRIMER-E Ltd.: Plymouth, UK, 2001; p. 91. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electronoca 2001, 4, 9. [Google Scholar]
- Novikov, A.A.; Abramova, E.N.; Sabirov, R.M. Fauna of the freshwater Harpacticoida (Copepoda) Lena River Delta. Rus. J. Zool. 2021, 100, 264–274. [Google Scholar] [CrossRef]
- Svendsen, J.I.; Alexanderson, H.; Astakhov, V.I.; Demidovd, I.; Dowdeswell, J.A.; Funder, S.; Gataullin, V.; Henriksen, M.; Hjort, C.; Houmark-Nielsen, M.; et al. Late Quaternary ice sheet history of Northern Eurasia. Quat. Sci. Rev. 2004, 23, 1229–1272. [Google Scholar] [CrossRef]
- Sinev, A.Y.; Karabanov, D.P.; Kotov, A.A. A new North Eurasian species of the Alona affinis complex (Cladocera: Chydoridae). Zootaxa 2020, 4767, 115–137. [Google Scholar] [CrossRef]
- Alekseev, V.R.; Defaye, D. Taxonomic differentiation and world geographical distribution of the Eucyclops serrulatus group (Copepoda, Cyclopidae, Eucyclopinae). In Studies on Freshwater Copepoda: A Volume in Honour of Bernard Dussart; Defaye, D., Suarez-Morales, E., Vaupel Klein, J.C., Eds.; BRILL: Leden, The Netherlands, 2011; pp. 41–72. [Google Scholar] [CrossRef]
- Alekseev, V.R. Eucyclops arcanus sp. n. (Copepoda, Cyclopoida) from waterbodies of Bolshezemelskaya Tundra and Prebaikalia. Zool. Zh. 1990, 69, 135–139. (In Russian) [Google Scholar]
- Dodson, S. Species richness of crustacean zooplankton in European lakes of different sizes. SIL Proc. 1922–2010 1991, 24, 1223–1229. [Google Scholar] [CrossRef]
- Kotov, A.A. Faunistic complexes of the Cladocera (Crustacea, Branchiopoda) of Eastern Siberia and Far East of Russia. Biol. Bull. 2016, 43, 970–987. [Google Scholar] [CrossRef]
- Garibian, P.G.; Chertoptud, E.S.; Sinev, A.Y.; Korovchinsky, N.M.; Kotov, A.A. Cladocera and Copepoda (Crustacea: Branchiopoda) of the Lake Bolon and its basin (Far East of Russia). Arthropoda Sel. 2019, 28, 37–63. [Google Scholar] [CrossRef]
- Weather Underground Web Portal. 2022. Available online: https://www.wunderground.com (accessed on 10 March 2022).




| Features | District (Medial Altitude m a.s.l.) | ||
|---|---|---|---|
| Central Part (492 m) | Western Slopes (347 m) | Foothills (84 m) | |
| Species richness of Cladocera | |||
| Anomopoda | 23 | 24 | 28 |
| Ctenopoda | 1 | 1 | 2 |
| Onychopoda | 1 | 1 | 1 |
| Species richness of Copepoda | |||
| Calanoida | 4 | 5 | 2 |
| Cyclopoida | 9 | 16 | 10 |
| Harpacticoida | 11 | 15 | 6 |
| Complex characteristics of fauna | |||
| Cladocera/Copepoda ratio | 1.04 | 0.72 | 1.70 |
| Species number per water body | 17.9 | 17.8 | 22.8 |
| Total species number | 50 | 62 | 49 |
| Meiobenthic crustacean | ||||
|---|---|---|---|---|
| Group | R2 | P | Prop. | Cumul. |
| Marginal Test | ||||
| District | 0.001 | 0.11151 | ||
| Type | 0.733 | 0.026129 | ||
| Feed | 0.002 | 0.083758 | ||
| Area | 0.47 | 0.032965 | ||
| Altitude | 0.003 | 0.092905 | ||
| Flow | 0.175 | 0.047264 | ||
| Temperature | 0.479 | 0.03343 | ||
| pH | 0.367 | 0.038716 | ||
| ppm | 0.022 | 0.06709 | ||
| Sediments | 0.31 | 0.040733 | ||
| Macrophytes | 0.212 | 0.044731 | ||
| Fish | 0.349 | 0.038301 | ||
| Sequential test | ||||
| +DIST | 0.11151 | 0.001 | 0.11151 | 0.11151 |
| +FEED | 0.19577 | 0.004 | 0.084269 | 0.19577 |
| Best Solution | ||||
| SUM | 0.50756 | 0.50756 | ||
| Planktonic crustacean | ||||
| Marginal Test | ||||
| District | 0.286 | 0.040146 | ||
| Type | 0.42 | 0.035657 | ||
| Feed | 0.216 | 0.044911 | ||
| Area | 0.001 | 0.10377 | ||
| Altitude | 0.547 | 0.030024 | ||
| Flow | 0.522 | 0.031115 | ||
| Temperature | 0.055 | 0.060912 | ||
| pH | 0.426 | 0.034961 | ||
| ppm | 0.113 | 0.049079 | ||
| Sediments | 0.229 | 0.042521 | ||
| Macrophytes | 0.024 | 0.075753 | ||
| Fish | 0.64 | 0.028557 | ||
| Sequential test | ||||
| +AREA | 0.10377 | 0.001 | 0.10377 | 0.10377 |
| +MACR | 0.19079 | 0.002 | 0.087018 | 0.19079 |
| Best Solution | ||||
| SUM | 0.56154 | 0.56154 | ||
| Taxon | Average Abundance, % | Contribution to Explained Similarity, % | General Explained Similarity, % |
|---|---|---|---|
| Assemblage 1. Area size0.0001–0.009 km2 | |||
| Heterocope appendiculata | 6.22 | 6.61 | 75.46 |
| Bosmina cf. longispina | 9.16 | 14.40 | |
| Chydorus cf. sphaericus | 11.67 | 20.53 | |
| Daphnia cf. longispina | 8.75 | 13.01 | |
| Pleuroxus truncatus | 7.29 | 6.35 | |
| Polyphemus pediculus | 10.23 | 14.55 | |
| Assemblage 2. Area size0.01–0.05 km2 | |||
| Heterocope appendiculata | 20.80 | 29.49 | 75.4 |
| Acanthodiaptomus tibetanus | 10.42 | 9.53 | |
| Bosmina cf. longispina | 27.77 | 36.39 | |
| Assemblage 3. Area size0.15–7.7 km2 | |||
| Heterocope appendiculata | 42.05 | 81.54 | 81.54 |
| Taxon | Contribution to Explained Difference, % | General Explained Difference, % |
|---|---|---|
| Assemblages 1–2 | ||
| Acanthodiaptomus tibetanus | 6.34 | 73.81 |
| Heterocope appendiculata | 12.32 | |
| Alonella excisa | 4.62 | |
| Bosmina coregoni | 4.42 | |
| Bosmina cf. longispina | 16.32 | |
| Chydorus cf. sphaericus | 6.68 | |
| Daphnia cf. longispina | 5.87 | |
| Pleuroxus truncatus | 5.75 | |
| Polyphemus pediculus | 7.03 | |
| Scapholeberis mucronata | 4.46 | |
| Assemblages 1–3 | ||
| Heterocope appendiculata | 22.57 | 68.7 |
| Cyclops scutifer | 5.31 | |
| Bosmina coregoni | 4.20 | |
| Bosmina cf. longispina | 11.86 | |
| Daphnia cf. longispina | 5.20 | |
| Chydorus cf. sphaericus | 6.26 | |
| Pleuroxus truncatus | 4.04 | |
| Polyphemus pediculus | 5.63 | |
| Scapholeberis mucronata | 3.65 | |
| Assemblages 2–3 | ||
| Heterocope appendiculata | 24.42 | 69.2 |
| Acanthodiaptomus tibetanus | 6.71 | |
| Cyclops scutifer | 6.17 | |
| Alonella excisa | 3.14 | |
| Bosmina cf. longispina | 21.10 | |
| Daphnia cf. longispina | 3.60 | |
| Holopedium gibberum | 4.08 | |
| Taxon | Average Abundance, % | Contribution to Explained Similarity, % | General Explained Similarity, % |
|---|---|---|---|
| Assemblage 1.Central part | |||
| Attheyella northumbrica trisetosa | 26.57 | 51.86 | 76.16 |
| Maraenobiotus insignipes | 24.92 | 24.30 | |
| Assemblage 2.Western slopes: Burgul River valley | |||
| Attheyella northumbrica trisetosa | 14.58 | 20.24 | 50.26 |
| Moraria mrazeki | 9.06 | 9.48 | |
| Phyllognathopus paludosus | 13.97 | 17.52 | |
| Chydorus cf. sphaericus | 10.70 | 12.50 | |
| Assemblage 3.Western slopes: Kutaramakan Lake basin and Irkinda River valley | |||
| Maraenobiotus insignipes | 13.16 | 13.30 | 68.04 |
| Moraria mrazeki | 10.12 | 25.84 | |
| Biapertura sibirica | 8.21 | 13.79 | |
| Chydorus cf. sphaericus | 11.32 | 12.11 | |
| Assemblage 4. Foothills | |||
| Moraria mrazeki | 15.94 | 17.06 | 38.52 |
| Chydorus cf. sphaericus | 16.48 | 21.46 | |
| Taxon | Contribution to Explained Difference, % | General Explained Difference, % |
|---|---|---|
| Assemblages 1–2 | ||
| Attheyella northumbrica trisetosa | 14.62 | 67.01 |
| Bryocamptus sp. nov. | 5.49 | |
| Epactophanes richardi | 4.29 | |
| Maraenobiotus insignipes | 16.01 | |
| Moraria mrazeki | 5.22 | |
| Pesceus schmeili | 7.14 | |
| Phyllognathopus paludosus | 8.05 | |
| Chydorus cf. sphaericus | 6.17 | |
| Assemblages 1–3 | ||
| Attheyella northumbrica trisetosa | 14.46 | 68.01 |
| Bryocamptus arcticus | 3.14 | |
| Bryocamptus sp. nov. | 5.18 | |
| Epactophanes richardi | 5.31 | |
| Maraenobiotus insignipes | 16.07 | |
| Moraria mrazeki | 5.51 | |
| Pesceus schmeili | 7.62 | |
| Biapertura sibirica | 4.47 | |
| Chydorus cf. sphaericus | 6.16 | |
| Assemblages 1–4 | ||
| Attheyella northumbrica trisetosa | 13.87 | 59.83 |
| Bryocamptus sp. nov. | 5.05 | |
| Epactophanes richardi | 3.94 | |
| Maraenobiotus insignipes | 13.22 | |
| Moraria mrazeki | 8.43 | |
| Pesceus schmeili | 6.57 | |
| Chydorus cf. sphaericus | 8.75 | |
| Faunistic Complex | Copepoda | Cladocera | Total Species List | |||
|---|---|---|---|---|---|---|
| Number | % | Number | % | Number | % | |
| Cosmopolites | 13 | 21.0 | 10 | 16.1 | 23 | 18.5 |
| Wide Palearctic | 15 | 24.2 | 38 | 61.3 | 53 | 42.7 |
| Holarctic | 4 | 6.5 | 6 | 9.7 | 10 | 8.1 |
| Subarctic and Arctic | 19 | 30.6 | 3 | 4.8 | 22 | 17.7 |
| East Asian | 3 | 4.8 | 3 | 4.8 | 6 | 4.8 |
| East Asian—North American | 1 | 1.6 | 2 | 3.2 | 3 | 2.4 |
| West Palearctic | 3 | 4.8 | 0 | 0 | 3 | 2.4 |
| Endemic | 4 | 6.5 | 0 | 0 | 4 | 3.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chertoprud, E.S.; Novichkova, A.A.; Novikov, A.A.; Fefilova, E.B.; Vorobjeva, L.V.; Pechenkin, D.S.; Glubokov, A.I. Assemblages of Meiobenthic and Planktonic Microcrustaceans (Cladocera and Copepoda) from Small Water Bodies of Mountain Subarctic (Putorana Plateau, Middle Siberia). Diversity 2022, 14, 492. https://doi.org/10.3390/d14060492
Chertoprud ES, Novichkova AA, Novikov AA, Fefilova EB, Vorobjeva LV, Pechenkin DS, Glubokov AI. Assemblages of Meiobenthic and Planktonic Microcrustaceans (Cladocera and Copepoda) from Small Water Bodies of Mountain Subarctic (Putorana Plateau, Middle Siberia). Diversity. 2022; 14(6):492. https://doi.org/10.3390/d14060492
Chicago/Turabian StyleChertoprud, Elena S., Anna A. Novichkova, Aleksandr A. Novikov, Elena B. Fefilova, Lada V. Vorobjeva, Dmitry S. Pechenkin, and Aleksandr I. Glubokov. 2022. "Assemblages of Meiobenthic and Planktonic Microcrustaceans (Cladocera and Copepoda) from Small Water Bodies of Mountain Subarctic (Putorana Plateau, Middle Siberia)" Diversity 14, no. 6: 492. https://doi.org/10.3390/d14060492
APA StyleChertoprud, E. S., Novichkova, A. A., Novikov, A. A., Fefilova, E. B., Vorobjeva, L. V., Pechenkin, D. S., & Glubokov, A. I. (2022). Assemblages of Meiobenthic and Planktonic Microcrustaceans (Cladocera and Copepoda) from Small Water Bodies of Mountain Subarctic (Putorana Plateau, Middle Siberia). Diversity, 14(6), 492. https://doi.org/10.3390/d14060492