
Comparative Phylogeography of Phoxinus, Delminichthys, Phoxinellus and Telestes in Dinaric Karst: Which Factors Have Influenced Their Current Distributions?


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Samples
2.3. DNA Extraction, Amplification and Sequencing
2.4. Haplotype Networks, Diversity Indices and Pairwise Distances
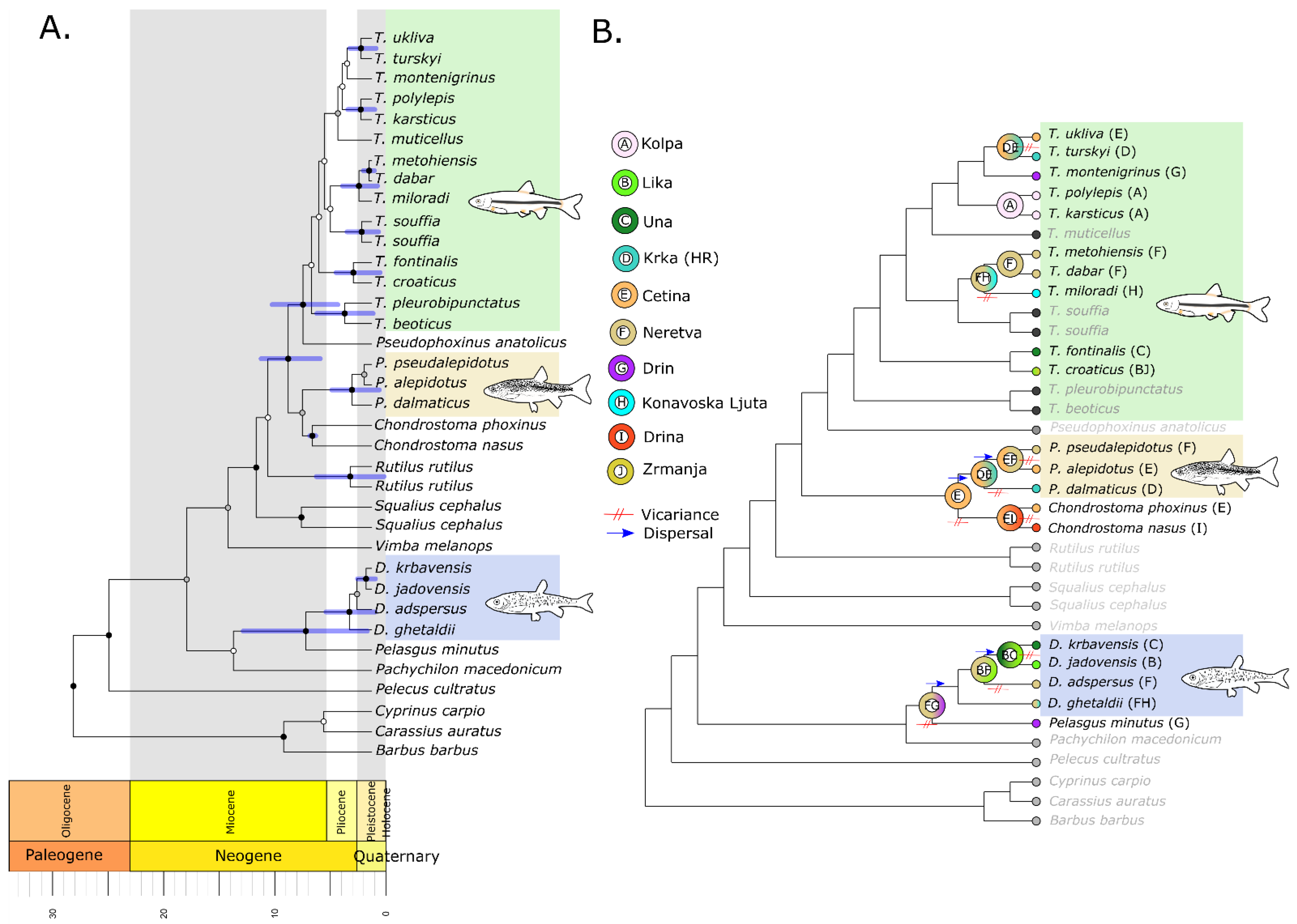
2.5. Phylogenetic Tree Reconstruction and Divergence Time Dating
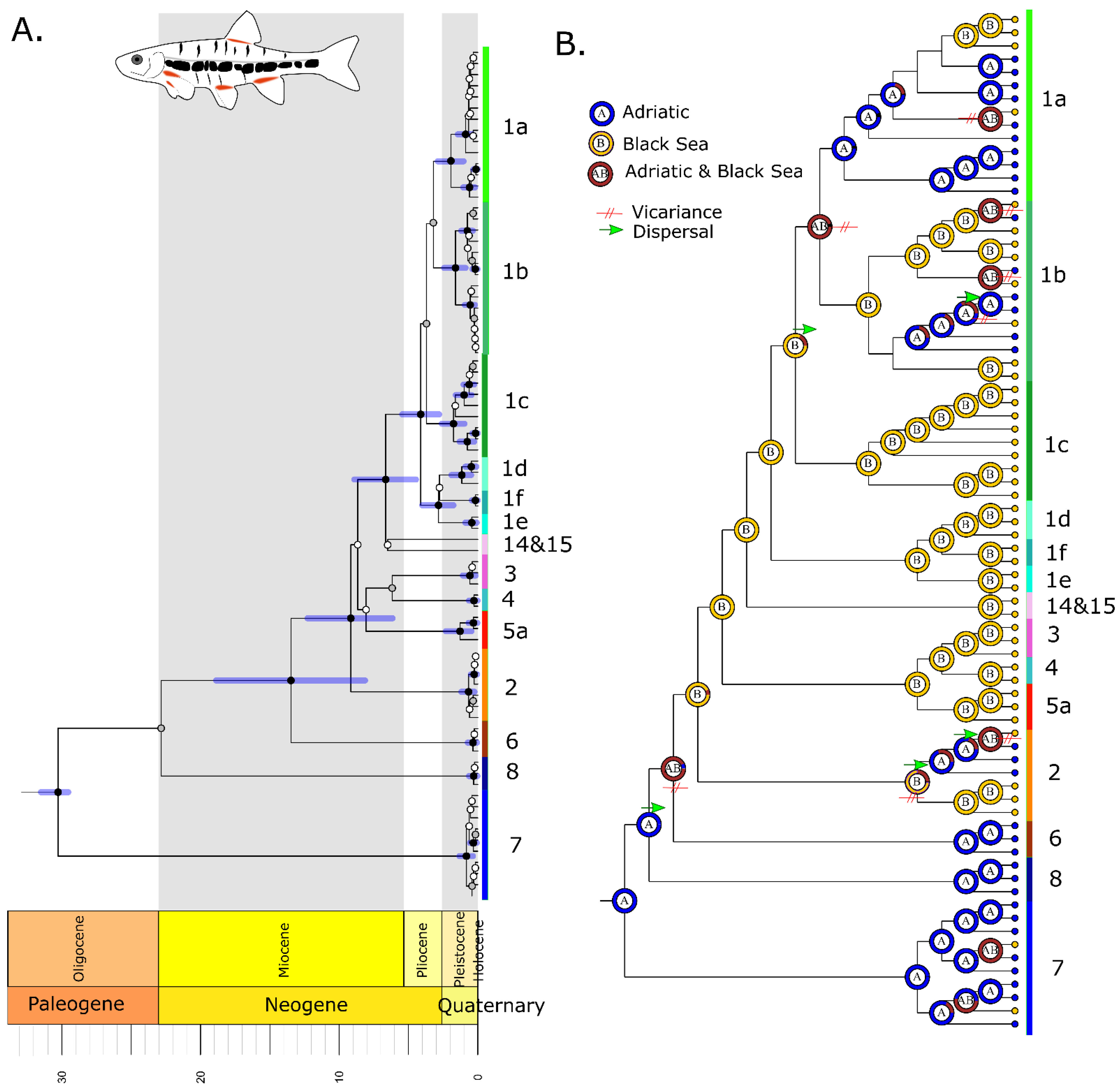
2.6. Ancestral Biogeography
3. Results
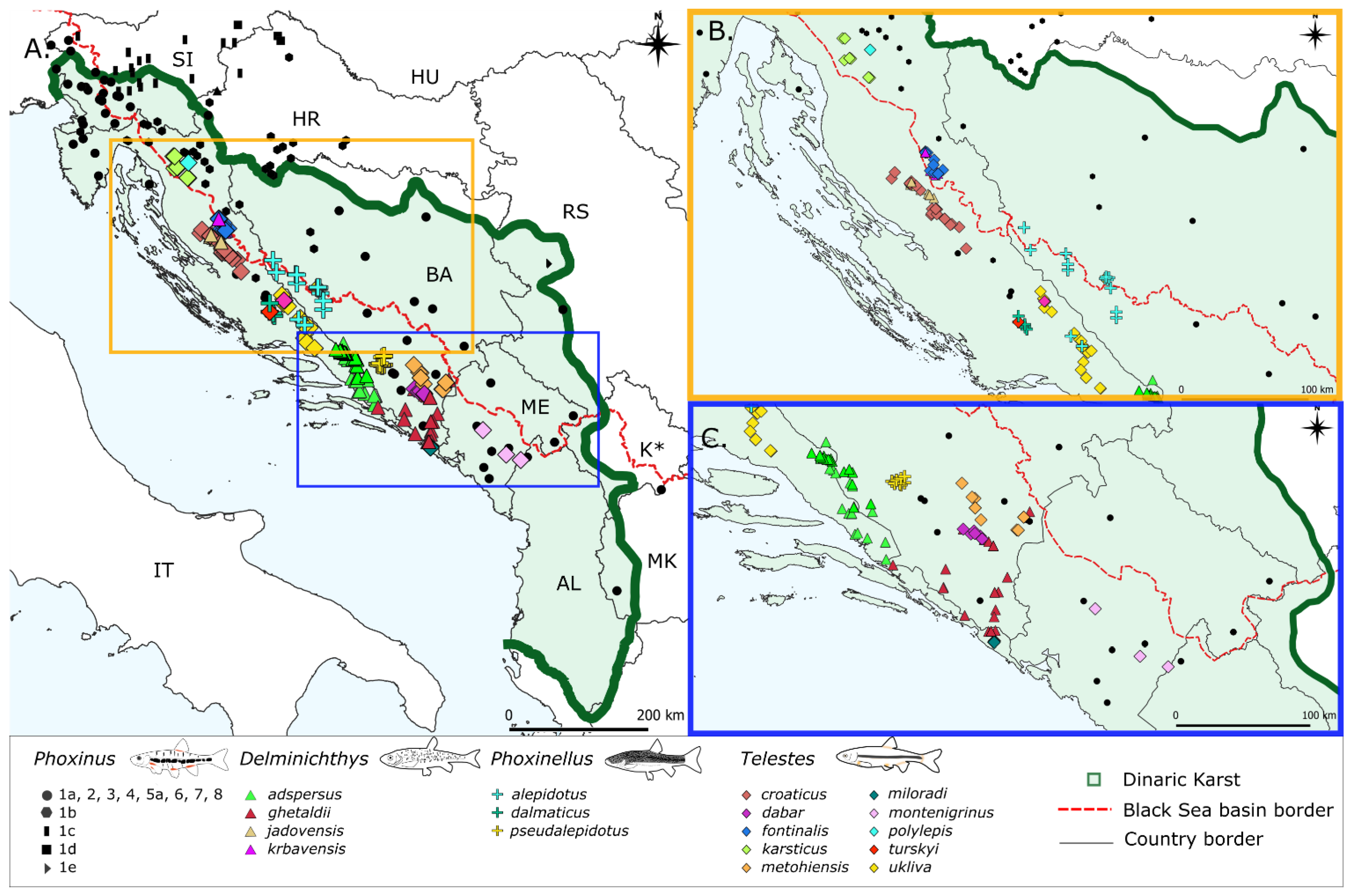
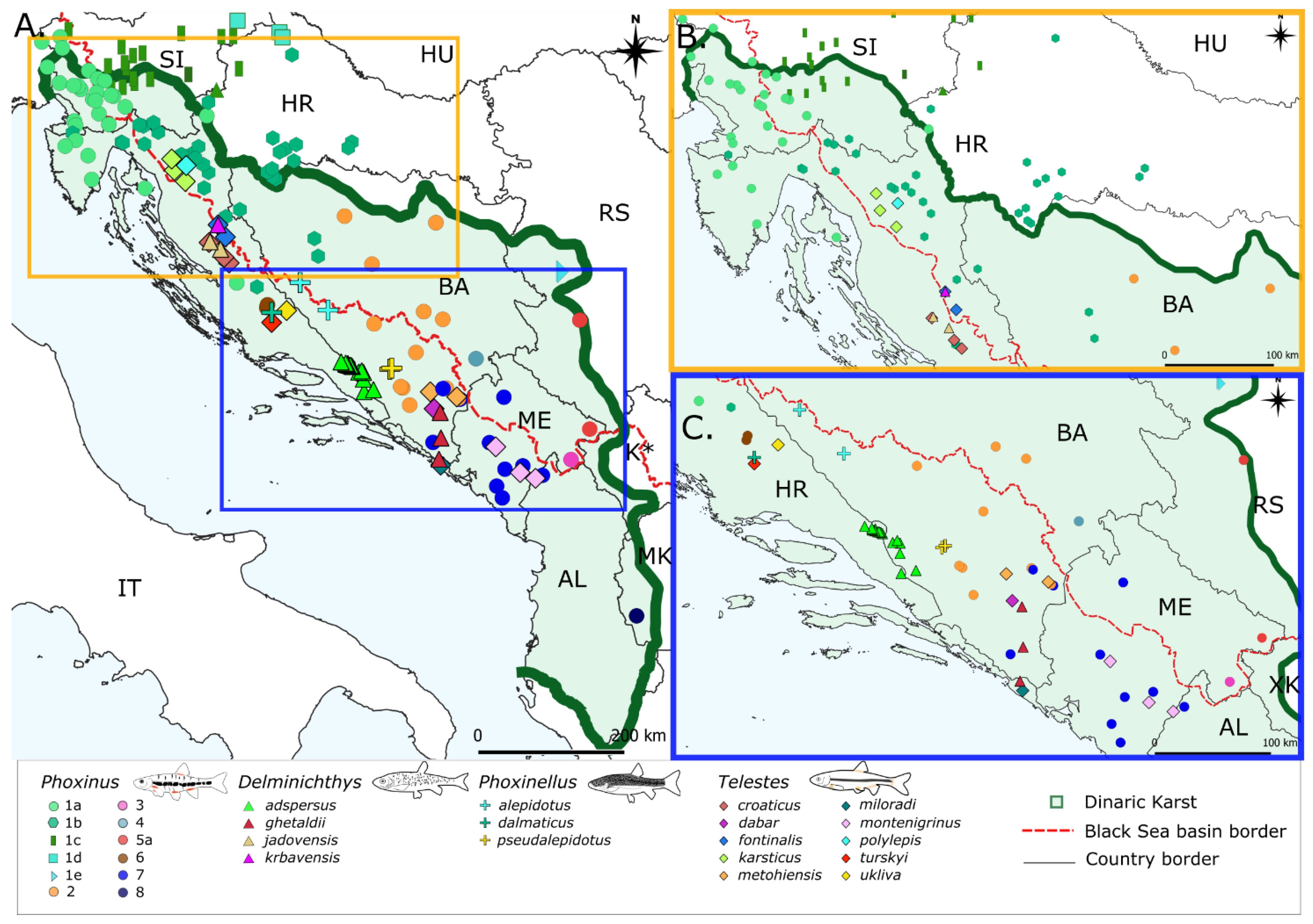
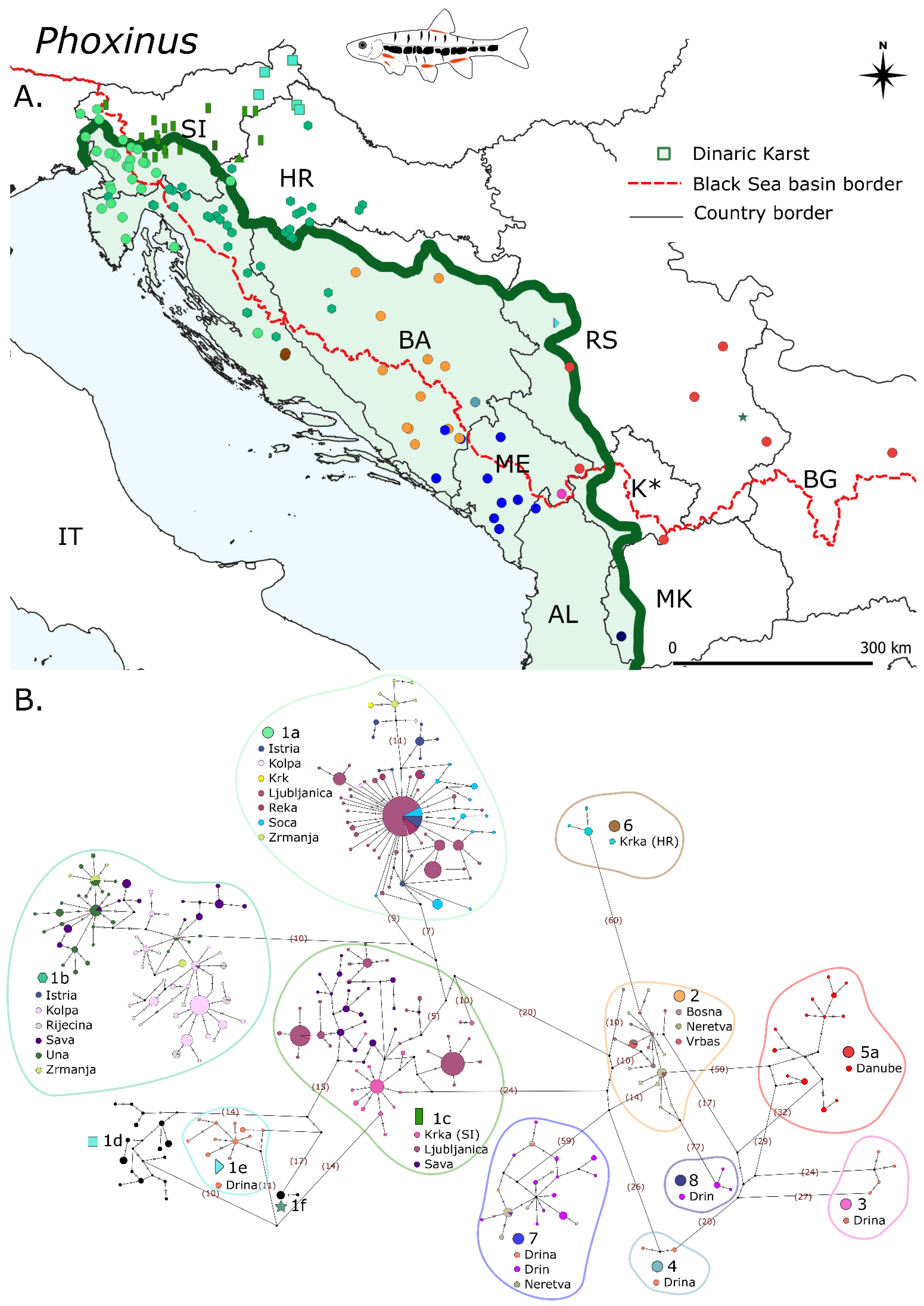
3.1. Phoxinus
3.1.1. Haplotype Networks and Distribution Patterns, Diversity Indices and p-Distances
3.1.2. Divergence Times and Ancestral Biogeography
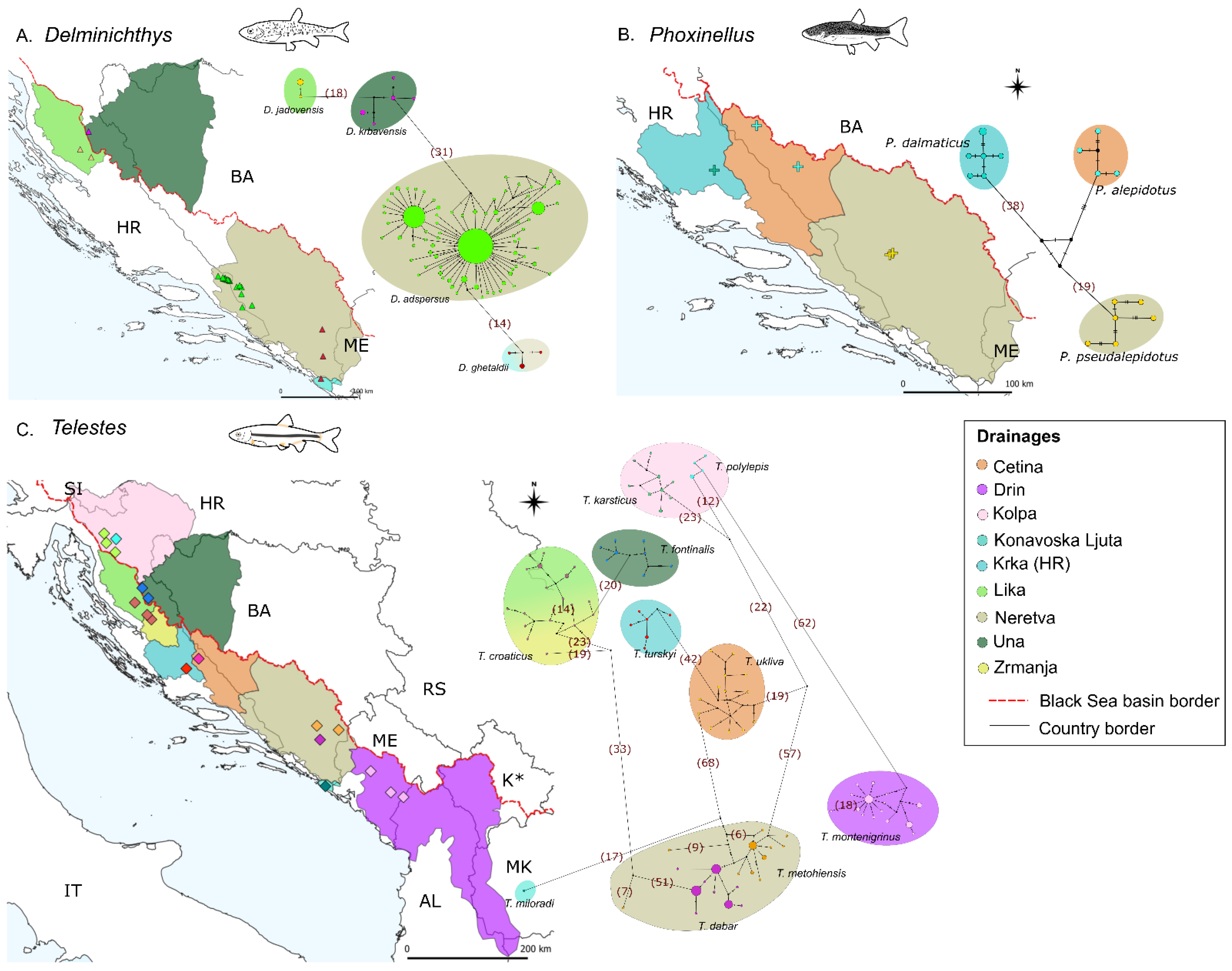
3.2. Delminichthys
3.2.1. Haplotype Networks and Distribution Patterns, Diversity Indices and p-Distances
3.2.2. Divergence Times and Ancestral Biogeography
3.3. Phoxinellus
3.3.1. Haplotype Networks and Distribution Patterns, Diversity Indices and p-Distances
3.3.2. Divergence Times and Ancestral Biogeography
3.4. Telestes
3.4.1. Haplotype Networks and Distribution Patterns, Diversity Indices and p-Distances
3.4.2. Divergence Times and Ancestral Biogeography
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bonacci, O. Karst Hydrology - With Special Reference to the Dinaric Karst; Springer Berlin: Heidelberg, Germany, 1987; 184p. [Google Scholar] [CrossRef]
- Zupan Hajna, N. Dinaric Karst—Geography and Geology. In Encyclopedia of Caves; Elsevier: Amsterdam, The Netherlands, 2019; pp. 353–362. ISBN 978-0-12-814124-3. [Google Scholar]
- Sket, B. Diversity Patterns in the Dinaric Karst. In Encyclopedia of Caves; Culver, D.C., White, W.B., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2012; pp. 228–238. [Google Scholar]
- Džukić, G. Kalezić The Biodiversity of Amphibians and Reptiles in the Balkan Peninsula. In Balkan Biodiversity: Pattern and Process in the European Hotspot; Griffiths, H.I., Kryštufek, B., Reed, J.M., Griffiths, H.I., Eds.; Kluwer Acad: Dordrecht, The Netherlands, 2004; ISBN 978-1-4020-2854-0. [Google Scholar]
- Krystufek, B.; Buzan, E.V.; Hutchinson, W.F.; Hänfling, B. Phylogeography of the Rare Balkan Endemic Martino’s Vole, Dinaromys bogdanovi, Reveals Strong Differentiation within the Western Balkan Peninsula. Mol. Ecol. 2007, 16, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Mihoci, I.; Hršak, V.; Kučinić, M.; Mičetić Stanković, V.; Delić, A.; Tvrtković, N. Butterfly Diversity and Biogeography on the Croatian Karst Mountain Biokovo: Vertical Distribution and Preference for Altitude and Aspect? Eur. J. Entomol. 2011, 108, 623–633. [Google Scholar] [CrossRef] [Green Version]
- Previšić, A.; Walton, C.; Kučinić, M.; Mitrikeski, P.T.; Kerovec, M. Pleistocene Divergence of Dinaric Drusus Endemics (Trichoptera, Limnephilidae) in Multiple Microrefugia within the Balkan Peninsula. Mol. Ecol. 2009, 18, 634–647. [Google Scholar] [CrossRef] [PubMed]
- Sket, B. The Nature of Biodiversity in Hypogean Waters and How It Is Endangered. Biodivers. Conserv. 1999, 8, 1319–1338. [Google Scholar] [CrossRef]
- Bănărescu, P.M. Distribution Pattern of the Aquatic Fauna of the Balkan Peninsula. In Balkan Biodiversity; Griffiths, H.I., Kryštufek, B., Reed, J.M., Eds.; Springer Netherlands: Dordrecht, Netherlands, 2004; pp. 203–217. ISBN 978-90-481-6732-6. [Google Scholar]
- Blondel, J.; Aronson, J.; Bodiou, J.-Y.; Boeuf, G. The Mediterranean Region: Biological Diversity in Space and Time; OUP Oxford: Oxford, UK, 2010; ISBN 978-0-19-157428-3. [Google Scholar]
- Tierno de Figueroa, J.M.; López-Rodríguez, M.J.; Fenoglio, S.; Sánchez-Castillo, P.; Fochetti, R. Freshwater Biodiversity in the Rivers of the Mediterranean Basin. Hydrobiologia 2013, 719, 137–186. [Google Scholar] [CrossRef]
- Dominique Durand, J.; Erhan, Ü.; Doadrio, I.; Pipoyan, S.; Templeton, A.R. Origin, Radiation, Dispersion and Allopatric Hybridization in the Chub Leuciscus cephalus. Proc. R. Soc. Lond. B 2000, 267, 1687–1697. [Google Scholar] [CrossRef]
- Kotlik, P.; Berrebi, P. Phylogeography of the Barbel (Barbus barbus) Assessed by Mitochondrial DNA Variation. Mol. Ecol. 2001, 10, 2177–2185. [Google Scholar] [CrossRef]
- Salzburger, W.; Brandstätter, A.; Gilles, A.; Parson, W.; Hempel, M.; Sturmbauer, C.; Meyer, A. Phylogeography of the Vairone (Leuciscus souffia, Risso 1826) in Central Europe. Mol. Ecol. 2003, 12, 2371–2386. [Google Scholar] [CrossRef] [Green Version]
- Šlechtová, V.; Bohlen, J.; Freyhof, J.; Persat, H.; Delmastro, G.B. The Alps as Barrier to Dispersal in Cold-Adapted Freshwater Fishes? Phylogeographic History and Taxonomic Status of the Bullhead in the Adriatic Freshwater Drainage. Mol. Phylogenetics Evol. 2004, 33, 225–239. [Google Scholar] [CrossRef]
- Falniowski, A.; Pešić, V.; Lewarne, B.; Grego, J.; Rysiewska, A.; Osikowski, A.; Hofman, S. Isolation and Endemism in Subterranean Aquatic Snails: Unexpected Case of Montenegrospeum bogici (Pešić et Glöer, 2012) (Gastropoda: Truncatelloidea: Hydrobiidae). Hydrobiologia 2021, 848, 4967–4990. [Google Scholar] [CrossRef]
- Hewitt, G.M. Genetic Consequences of Climatic Oscillations in the Quaternary. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2004, 359, 183–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verovnik, R.; Sket, B.; Trontelj, P. The Colonization of Europe by the Freshwater Crustacean Asellus aquaticus (Crustacea: Isopoda) Proceeded from Ancient Refugia and Was Directed by Habitat Connectivity: Colonization of europe by asellus aquaticus. Mol. Ecol. 2005, 14, 4355–4369. [Google Scholar] [CrossRef]
- Trontelj, P.; Gorički, Š.; Polak, S.; Verovnik, R.; Zakšek, V.; Sket, B. Age Estimates for Some Subterranean Taxa and Lineages in the Dinaric Karst. AC 2007, 36, 183–189. [Google Scholar] [CrossRef]
- De Leeuw, A.; Mandic, O.; Vranjković, A.; Pavelić, D.; Harzhauser, M.; Krijgsman, W.; Kuiper, K.F. Chronology and Integrated Stratigraphy of the Miocene Sinj Basin (Dinaride Lake System, Croatia). Palaeogeogr. Palaeoclimatol. Palaeoecol. 2010, 292, 155–167. [Google Scholar] [CrossRef]
- Jurišić-Polšak, Z.; Krizmanić, Z.; Hajek-Tedesse, V. Freshwater Miocene of Krbavsko Polje in Lika (Croatia))—Slatkovodni Miocen Krbavskog Polja u Lici (Hrvatska). Geol. Croat. 1993, 46, 213–228. [Google Scholar]
- Harzhauser, M.; Mandic, O.; Latal, C.; Kern, A. Stable Isotope Composition of the Miocene Dinaride Lake System Deduced from Its Endemic Mollusc Fauna. Hydrobiologia 2012, 682, 27–46. [Google Scholar] [CrossRef]
- Hewitt, G.M. Post-Glacial Re-Colonization of European Biota. Biol. J. Linn. Soc. 1999, 68, 87–112. [Google Scholar] [CrossRef]
- Palandačić, A.; Matschiner, M.; Zupančič, P.; Snoj, A. Fish Migrate Underground: The Example of Delminichthys adspersus (Cyprinidae). Mol. Ecol. 2012, 21, 1658–1671. [Google Scholar] [CrossRef]
- Palandačić, A.; Bonacci, O.; Snoj, A. Molecular Data as a Possible Tool for Tracing Groundwater Flow in Karst Environment: Example of Delminichthys adspersus in Dinaric Karst System: Fish as Groundwater Tracers in Karst. Ecohydrology 2012, 5, 791–797. [Google Scholar] [CrossRef]
- Trontelj, P. Vicariance and Dispersal in Caves. In Encyclopedia of Caves; Elsevier: Amsterdam, The Netherlands, 2019; pp. 1103–1109. ISBN 978-0-12-814124-3. [Google Scholar]
- Sanmartín, I. Historical Biogeography: Evolution in Time and Space. Evol. Educ. Outreach 2012, 5, 555–568. [Google Scholar] [CrossRef] [Green Version]
- Kappus, B.; Peissner, T.; Rawer-Jost, C. Distribution and Habitat Conditions of Crayfish Populations in the Urban Freshwater Systems of Stuttgart (Baden-Württemberg, Germany). Freshw. Crayfish 1999, 12, 778–785. [Google Scholar]
- Jelić, D.; Jelić, M. Telestes miloradi Bogutskaya, Zupancic, Bogut & Naseka, 2012 and Delminichthys ghetaldii (Steindachner, 1882) Re-Discovered in Croatia, Requiring Urgent Protection. J. Appl. Ichthyol. 2015, 31, 1133–1136. [Google Scholar] [CrossRef]
- Kottelat, M.; Freyhof, J. Handbook of European Freshwater Fishes; Kottelat, Cornol and Freyhof: Berlin, Germany, 2007; 646p. [Google Scholar]
- Ćaleta, M.; Marčić, Z.; Buj, I.; Zanella, D.; Mustafić, P.; Duplić, A.; Horvatić, S. A Review of Extant Croatian Freshwater Fish and Lampreys: Annotated List and Distribution. Croat. J. Fish. 2019, 77, 137–234. [Google Scholar] [CrossRef] [Green Version]
- Buj, I.; Marčić, Z.; Čavlović, K.; Ćaleta, M.; Tutman, P.; Zanella, D.; Duplić, A.; Raguž, L.; Ivić, L.; Horvatić, S.; et al. Multilocus Phylogenetic Analysis Helps to Untangle the Taxonomic Puzzle of Chubs (Genus Squalius: Cypriniformes: Actinopteri) in the Adriatic Basin of Croatia and Bosnia and Herzegovina. Zool. J. Linn. Soc. 2020, 189, 953–974. [Google Scholar] [CrossRef]
- Buj, I.; Knjaz, M.; Ćaleta, M.; Marčić, Z.; Ivić, L.; Onorato, L.; Šanda, R.; Vukić, J.; Horvatić, S.; Zanella, D.; et al. Phylogeographic Pattern, Genetic Diversity, and Evolutionary History of the Enigmatic Freshwater Fish Species Aulopyge huegelii (Actinopterygii: Cyprinidae). J. Zool. Syst. Evol. Res. 2021, 59, 2086–2102. [Google Scholar] [CrossRef]
- Hrbek, T.; Stölting, K.N.; Bardakci, F.; Küçük, F.; Wildekamp, R.H.; Meyer, A. Plate Tectonics and Biogeographical Patterns of the Pseudophoxinus (Pisces: Cypriniformes) Species Complex of Central Anatolia, Turkey. Mol. Phylogenetics Evol. 2004, 32, 297–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doadrio, I.; Carmona, J.A. Phylogenetic Relationships and Biogeography of the Genus Chondrostoma Inferred from Mitochondrial DNA Sequences. Mol. Phylogenetics Evol. 2004, 33, 802–815. [Google Scholar] [CrossRef]
- Perea, S.; Böhme, M.; Zupančič, P.; Freyhof, J.; Šanda, R.; Özuluğ, M.; Abdoli, A.; Doadrio, I. Phylogenetic Relationships and Biogeographical Patterns in Circum-Mediterranean Subfamily Leuciscinae (Teleostei, Cyprinidae) Inferred from Both Mitochondrial and Nuclear Data. BMC Evol. Biol. 2010, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Palandačić, A.; Kruckenhauser, L.; Ahnelt, H.; Mikschi, E. European Minnows through Time: Museum Collections Aid Genetic Assessment of Species Introductions in Freshwater Fishes (Cyprinidae: Phoxinus Species Complex). Heredity 2020, 124, 410–422. [Google Scholar] [CrossRef]
- Bogutskaya, N.G.; Jelić, D.; Vucić, M.; Jelić, M.; Diripasko, O.A.; Stefanov, T.; Klobučar, G. Description of a New Species of Phoxinus from the Upper Krka River (Adriatic Basin) in Croatia (Actinopterygii: Leuciscidae), First Discovered as a Molecular Clade. J. Fish Biol. 2020, 96, 378–393. [Google Scholar] [CrossRef]
- Palandačić, A.; Bravničar, J.; Zupančič, P.; Šanda, R.; Snoj, A. Molecular Data Suggest a Multispecies Complex of Phoxinus (Cyprinidae) in the Western Balkan Peninsula. Mol. Phylogenetics Evol. 2015, 92, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Vučić, M.; Jelić, D.; Žutinić, P.; Grandjean, F.; Jelić, M. Distribution of Eurasian Minnows (Phoxinus: Cypriniformes) in the Western Balkans. Knowl. Manag. Aquat. Ecosyst. 2018, 419, 11. [Google Scholar] [CrossRef] [Green Version]
- Zaccara, S.; Stefani, F.; Delmastro, G.B. Phylogeographical Structure of Vairone Telestes muticellus (Teleostei, Cyprinidae) within Three European Peri-Mediterranean Districts. Zool. Scr. 2007, 36, 443–453. [Google Scholar] [CrossRef]
- Marchetto, F.; Zaccara, S.; Muenzel, F.M.; Salzburger, W. Phylogeography of the Italian Vairone (Telestes muticellus, Bonaparte 1837) Inferred by Microsatellite Markers: Evolutionary History of a Freshwater Fish Species with a Restricted and Fragmented Distribution. BMC Evol. Biol. 2010, 10, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sket, B. Can We Agree on an Ecological Classification of Subterranean Animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Jelić, M.; Klobučar, G.I.V.; Grandjean, F.; Puillandre, N.; Franjević, D.; Futo, M.; Amouret, J.; Maguire, I. Insights into the Molecular Phylogeny and Historical Biogeography of the White-Clawed Crayfish (Decapoda, Astacidae). Mol. Phylogenetics Evol. 2016, 103, 26–40. [Google Scholar] [CrossRef] [PubMed]
- Ćurčić, V. Narodno ribarstvo u Bosni i Hercegovini, II. Herzegovina. Glas. Zemalj. Muz. Bosne I Hercegovine 1913, 25, 421–514. [Google Scholar]
- Trgovčević, L. Paraphoxinus Blkr. i Telestes Bonap. U vodama Like I Krbave. Nastavni Vjesn. 1905, 14, 331–335. [Google Scholar]
- Vuković, T.; Ivanović, B. Slatkovodne Ribe Jugoslavije; Zemaljski musej BiH-Prirodnjačko odjeljenje: Sarajevo, Bosnia and Herzegovina, 1971. [Google Scholar]
- Vuković, T. Ribe Bosne I Herzigovine; Svjetlost: Sarajevo, Bosnia and Herzegovina, 1977. [Google Scholar]
- Bogutskaya, N.G.; Zupancic, P. Phoxinellus pseudalepidotus (Teleostei: Cyprinidae), a New Species from the Neretva Basin with an Overview of the Morphology of Phoxinellus Species of Croatia and Bosnia-Herzegovina. Ichthyol. Explor. Freshw. 2003, 14, 369–383. [Google Scholar]
- Mrakovčić, M.; Brigić, A.; Buj, I.; Ćaleta, M.; Mustafić, P.; Zanella, D. Red Book of Freshwater Fish of Croatia; Ministry of Culture, State Institute for Nature Protection: Zagreb, Croatia, 2006. [Google Scholar]
- Zanella, D.; Mihaljevic, Z.; Mrakovcic, M.; Caleta, M. Ecology and Diet of the Endemic Telestes ukliva (Cyprinidae) in the Cetina River System, Croatia. Cybium 2009, 33, 97–105. [Google Scholar]
- Scharnweber, K. Morphological and Trophic Divergence of Lake and Stream Minnows (Phoxinus phoxinus). Ecol. Evol. 2020, 10, 8358–8367. [Google Scholar] [CrossRef] [PubMed]
- Marćić, Z.; Komljenović, I.; Ćaleta, M.; Buj, I.; Zanella, D.; Mustafić, P. Age, Growth, and Population Structure of Endemic Telestes karsticus (Actinopterygii: Cyprinidae, Leuciscinae) from Sušik Creek, Croatia. AIeP 2021, 51, 225–232. [Google Scholar] [CrossRef]
- Schönhuth, S.; Vukić, J.; Šanda, R.; Yang, L.; Mayden, R.L. Phylogenetic Relationships and Classification of the Holarctic Family Leuciscidae (Cypriniformes: Cyprinoidei). Mol. Phylogenetics Evol. 2018, 127, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Buj, I.; Marčić, Z.; Ćaleta, M.; Šanda, R.; Geiger, M.F.; Freyhof, J.; Machordom, A.; Vukić, J. Ancient Connections among the European Rivers and Watersheds Revealed from the Evolutionary History of the Genus Telestes (Actinopterygii; Cypriniformes). PLoS ONE 2017, 12, e0187366. [Google Scholar] [CrossRef] [Green Version]
- Freyhof, J.; Lieckfeldt, D.; Bogutskaya, N.G.; Pitra, C.; Ludwig, A. Phylogenetic Position of the Dalmatian Genus Phoxinellus and Description of the Newly Proposed Genus Delminichthys (Teleostei: Cyprinidae). Mol. Phylogenetics Evol. 2006, 38, 416–425. [Google Scholar] [CrossRef] [PubMed]
- Palandačić, A.; Zupančič, P.; Snoj, A. Revised Classification of Former Genus Phoxinellus Using Nuclear DNA Sequences. Biochem. Syst. Ecol. 2010, 38, 1069–1073. [Google Scholar] [CrossRef]
- Mustafić, P.; Buj, I.; Opašić, M.; Zanella, D.; Marčić, Z.; Ćaleta, M.; Šanda, R.; Horvatić, S.; Mrakovčić, M. Morphological Comparison of Delminichthys ghetaldii (Steindachner, 1882), D. adspersus (Heckel, 1843), D. jadovensis (Zupančič & Bogutskaya, 2002) and D. krbavensis (Zupančič & Bogutskaya, 2002), Endemic Species of the Dinaric Karst, Croatia. J. Appl. Ichthyol. 2017, 33, 256–262. [Google Scholar] [CrossRef]
- Marčić, Z.; Buj, I.; Duplić, A.; Ćaleta, M.; Mustafić, P.; Zanella, D.; Zupančič, P.; Mrakovčić, M. A New Endemic Cyprinid Species from the Danube Drainage. J. Fish Biol. 2011, 79, 418–430. [Google Scholar] [CrossRef]
- Matas, M. Krš Hrvatske: Geografski Pregled i Značenje; Geografsko društvo: Split, Croatia, 2009. [Google Scholar]
- Petrič, M.; Ravbar, N.; Gostinčar, P.; Krsnik, P.; Gacin, M. GIS Database of Groundwater Flow Characteristics in Carbonate Aquifers: Tracer Test Inventory from Slovenian Karst. Appl. Geogr. 2020, 118, 102191. [Google Scholar] [CrossRef]
- Yu, Y.; Harris, A.J.; He, X. S-DIVA (Statistical Dispersal-Vicariance Analysis): A Tool for Inferring Biogeographic Histories. Mol. Phylogenet. Evol. 2010, 56, 848–850. [Google Scholar] [CrossRef]
- Gams, I. The Polje: The Problem of Definition. Z. Für Geomorphol. 1978, 22, 170–181. [Google Scholar]
- Bonacci, O. 6.11 Poljes, Ponors and Their Catchments. In Treatise on Geomorphology; Shroder, J.F., Ed.; Academic Press, Elsevier: San Diego, CA, USA, 2013; pp. 112–120. ISBN 978-0-08-088522-3. [Google Scholar]
- Milanović, P. Dinaride Poljes. In Encyclopedia of Caves and Karst Science; Gunn, J., Ed.; Fitzroy Dearborn: New York, NY, USA, 2004; pp. 291–293. [Google Scholar]
- Milanović, P. Water Resources Engineering in Karst; CRC press: Boca Raton, FL, USA, 2004. [Google Scholar]
- Šerko, A. Kraški Pojavi v Jugoslaviji. Geogr. Vestn. 1948, XIX, 43–70. [Google Scholar]
- Vogt, J.; Soille, P.; De Jager, A.; Rimaviciute, E.; Mehl, W.; Foisneau, S.; Bodis, K.; Dusart, J.; Paracchini, M.; Haastrup, P.; et al. A Pan-European River and Catchment Database. Report No. EUR 22920; Office for Official Publications of the European Communities: Luxembourg, Luxembourg, 2007. [Google Scholar]
- DanubeGIS Danube River Basin GIS Maps and Layers Documentation. 2016. Available online: https://www.danubegis.org (accessed on 4 June 2021).
- QGIS Development Team; Open Source Geospatial Foundation. QGIS Geographic Information System. 2009. Available online: http://qgis.osgeo.org (accessed on 13 November 2021).
- Bergsten, J.; Engl, M.; Ericsson, P. A DNA Key to All Swedish Vertebrates. 2014. Available online: https://www.ncbi.nlm.nih.gov/nuccore/KJ128752 (accessed on 31 August 2021).
- Leigh, J.W.; Bryant, D. POPART: Full-feature Software for Haplotype Network Construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Böhme, M.; Ilg, A. Fossil Fishes, Amphibians, Reptiles—FosFARbase. 2013. Available online: www.wahre-staerke.com (accessed on 3 April 2021).
- Mödden, C.; Schäfer, P.; Reichenbacher, B.; Schwarz, J.; Kadolsky, D. Säugetiere, Fisch-Otolithen, Ostracoden, Mollusken Und Charophyten Aus Den Süßwasser-Schichten (Oligozän) von Wolfsheim Im Mainzer Becken. Paläontologische Z. 2000, 74, 343–361. [Google Scholar] [CrossRef]
- Bouckaert, R.; Vaughan, T.G.; Barido-Sottani, J.; Duchêne, S.; Fourment, M.; Gavryushkina, A.; Heled, J.; Jones, G.; Kühnert, D.; De Maio, N.; et al. BEAST 2.5: An Advanced Software Platform for Bayesian Evolutionary Analysis. PLOS Comput. Biol. 2019, 15, e1006650. [Google Scholar] [CrossRef] [Green Version]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Corral-Lou, A.; Perea, S.; Aparicio, E.; Doadrio, I. Phylogeography and Species Delineation of the Genus Phoxinus Rafinesque, 1820 (Actinopterygii: Leuciscidae) in the Iberian Peninsula. J. Zool. Syst. Evol. Res. 2019, 57, 926–941. [Google Scholar] [CrossRef]
- Dowling, T.E.; Tibbets, C.A.; Minckley, W.L.; Smith, G.R. Evolutionary Relationships of the Plagopterins (Teleostei: Cyprinidae) from cytochrome b Sequences. Copeia 2002, 2002, 665–678. [Google Scholar] [CrossRef] [Green Version]
- Drummond, A.J.; Ho, S.Y.W.; Phillips, M.J.; Rambaut, A. Relaxed Phylogenetics and Dating with Confidence. PLoS Biol. 2006, 4, e88. [Google Scholar] [CrossRef] [PubMed]
- Drummond, A.J.; Rambaut, A. BEAST: Bayesian Evolutionary Analysis by Sampling Trees. BMC Evol. Biol. 2007, 7, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, M.A.; Lloyd, G.T. Strap: Stratigraphic Tree Analysis for Palaeontology, R. package version 1.4; R Foundation for Statistical Computing: Vienna, Austria, 2014. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Yu, Y.; Harris, A.J.; Blair, C.; He, X. RASP (Reconstruct Ancestral State in Phylogenies): A Tool for Historical Biogeography. Mol. Phylogenetics Evol. 2015, 87, 46–49. [Google Scholar] [CrossRef] [PubMed]
- Denys, G.P.J.; Dettai, A.; Persat, H.; Daszkiewicz, P.; Hautecœur, M.; Keith, P. Revision of Phoxinus in France with the Description of Two New Species (Teleostei, Leuciscidae). Cybium 2020, 44, 205–237. [Google Scholar] [CrossRef]
- Krivokapić, M. Struktura Populacije Endemične Podvrste Leuciscus soffia montenegrinus (Vuković, 1963.) Iz Rijeke Morače (Crna Gora, Jugoslavija). Glas. Republičkog Zavoda Zaštite Prir. Prir. Muz. Podgor. 1992, 25, 83–91. [Google Scholar]
- Krivokapic, M. Growth of Telestes montenegrinus from River Moraca, Montenegro (Cyprinidae). Ital. J. Zool. 1998, 65, 241–242. [Google Scholar] [CrossRef]
- Frost, W.E. The Natural History of the Minnow, Phoxinus phoxinus. J. Anim. Ecol. 1943, 12, 139. [Google Scholar] [CrossRef]
- Cheverie, J.C.; Lynn, W.G. High Temperature Tolerance and Thyroid Activity in the Teleost Fish, Tanichthys albonubes. Biol. Bull. 1963, 124, 153–162. [Google Scholar] [CrossRef]
- Keith, P.; Persat, H.; Allardi, J. Les Poissons d’eau Douce de France. In Collection Inventaires & biodiversité, 2nd ed.; revue, augmentée et mise à jour; Biotope Muséum National D’histoire Naturelle: Paris, France, 2011; ISBN 978-2-914817-69-1. [Google Scholar]
- Lelek, A. The Freshwater Fishes of Europe. v. 9: Threatened Fishes of Europe; AULA-Verlag: Wiesbaden, Germany, 1987. [Google Scholar]
- Huet, M. Aperçu Des Relations Entre La Pente et Les Populations Piscicoles Des Eaux Courantes. Schweiz. Z. Für Hydrol. 1949, 11, 332–351. [Google Scholar] [CrossRef]
- Huet, M. Biologie, Profils En Long et En Travers Des Eaux Courantes. Bull. Français De Piscic. 1954, 175, 41–53. [Google Scholar] [CrossRef]
- Huet, M. Profiles and Biology of Western European Streams as Related to Fish Management. Trans. Am. Fish. Soc. 1959, 88, 155–163. [Google Scholar] [CrossRef]
- Aarts, B.G.W.; Nienhuis, P.H. Fish Zonations and Guilds as the Basis for Assessment of Ecological Integrity of Large Rivers. Hydrobiologia 2003, 500, 157–178. [Google Scholar] [CrossRef]
- Museth, J.; Hesthagen, T.; Sandlund, O.T.; Thorstad, E.B.; Ugedal, O. The History of the Minnow Phoxinus phoxinus (L.) in Norway: From Harmless Species to Pest. J. Fish Biol. 2007, 71, 184–195. [Google Scholar] [CrossRef]
- Reier, S.; Kruckenhauser, L.; Snoj, A.; Trontelj, P.; Palandačić, A. The Minnow Phoxinus lumaireul (Leuciscidae) Shifts the Adriatic/Black Sea Basin Divide in the North-Western Dinaric Karst. In prod. Ecohydrology. 2022. [Google Scholar] [CrossRef]
- Zupan Hajna, N.; Mihevc, A.; Pruner, P.; Bosak, P. Age of Dinaric Karst Cave Sediments in SW Slovenia. In Proceedings of the Book of Abstracts, Sarajevo, Bosnia and Herzegovina, 13 October 2011. [Google Scholar]
- Milanović, P. Karst of Eastern Herzegovina, the Dubrovnik Littoral and Western Montenegro. Env. Earth Sci. 2015, 74, 15–35. [Google Scholar] [CrossRef]
- Palandačić, A.; Witman, K.; Spikmans, F. Molecular Analysis Reveals Multiple Native and Alien Phoxinus Species (Leusciscidae) in the Netherlands and Belgium. Biol. Invasions 2022, 1–11. [Google Scholar] [CrossRef]
- Helfman, G.S.; Collette, B.; Facey, D.; Bowen, B.W. The Diversity of Fishes: Biology, Evolution, and Ecology, 2nd ed.; Wiley-Blackwell: Chichester, UK; Hoboken, NJ, USA, 2009; ISBN 978-1-4051-2494-2. [Google Scholar]
- Borowsky, R. Cavefishes. Curr. Biol. 2018, 28, R60–R64. [Google Scholar] [CrossRef] [Green Version]
- Howarth, F.G.; Moldovan, O.T. The Ecological Classification of Cave Animals and Their Adaptations. In Cave Ecology; Moldovan, O.T., Kováč, Ľ., Halse, S., Eds.; Springer International Publishing: Cham, The Netherlands, 2018; pp. 41–67. ISBN 978-3-319-98852-8. [Google Scholar]
- Zupančič, P. Rijetke I Ugrozene Slatkovodne Ribe Jadranskog Slijeva Hrvatske, Slovenije i Bosne i Hercegovine (Rare and Endangered Freshwater Fishes of Croatia, Slovenia and Bosnia and Hercegovina). AZV Dolsko 2008, 79. [Google Scholar]
- Bogutskaya, N.G. The morphological basis for the classification of cyprinid fishes (Leuciscinae, Cyprinidae). Communication 2. Vopr. Ikhtiologii 1990, 30, 920–933. [Google Scholar]
- Cavender, T.; Coburn, M. Phylogenetic Relationships of North American Cyprinidae. In Systematics, Historical Ecology, and North American Freshwater Fishes; Mayden, R.L., Ed.; Stanford University Press: Stanford, CA, USA, 1992; pp. 293–327. [Google Scholar]
- Ramler, D.; Palandačić, A.; Delmastro, G.B.; Wanzenböck, J.; Ahnelt, H. Morphological Divergence of Lake and Stream Phoxinus of Northern Italy and the Danube Basin Based on Geometric Morphometric Analysis. Ecol Evol 2017, 7, 572–584. [Google Scholar] [CrossRef]
- Klobučar, G.I.V.; Podnar, M.; Jelić, M.; Franjević, D.; Faller, M.; Štambuk, A.; Gottstein, S.; Simić, V.; Maguire, I. Role of the Dinaric Karst (Western Balkans) in Shaping the Phylogeographic Structure of the Threatened Crayfish Austropotamobius torrentium. Freshw. Biol. 2013, 58, 1089–1105. [Google Scholar] [CrossRef]
- Žganec, K.; Lunko, P.; Stroj, A.; Mamos, T.; Grabowski, M. Distribution, Ecology and Conservation Status of Two Endemic Amphipods, Echinogammarus acarinatus and Fontogammarus dalmatinus, from the Dinaric Karst Rivers, Balkan Peninsula. Ann. Limnol. Int. J. Lim. 2016, 52, 13–26. [Google Scholar] [CrossRef] [Green Version]
- Konec, M.; Delić, T.; Trontelj, P. DNA Barcoding Sheds Light on Hidden Subterranean Boundary between Adriatic and Danubian Drainage Basins: Subterranean Drainage Basins Elucidated by DNA Barcoding. Ecohydrology 2016, 9, 1304–1312. [Google Scholar] [CrossRef]
- Gorički, Š.; Trontelj, P. Structure and Evolution of the Mitochondrial Control Region and Flanking Sequences in the European Cave Salamander Proteus anguinus. Gene 2006, 378, 31–41. [Google Scholar] [CrossRef] [PubMed]
- Pârvulescu, L.; Pérez-Moreno, J.L.; Panaiotu, C.; Drăguț, L.; Schrimpf, A.; Popovici, I.; Zaharia, C.; Weiperth, A.; Gál, B.; Schubart, C.D.; et al. A Journey on Plate Tectonics Sheds Light on European Crayfish Phylogeography. Ecol. Evol. 2019, 9, 1957–1971. [Google Scholar] [CrossRef] [Green Version]
- Lovrenčić, L.; Bonassin, L.; Boštjančić, L.L.; Podnar, M.; Jelić, M.; Klobučar, G.; Jaklič, M.; Slavevska-Stamenković, V.; Hinić, J.; Maguire, I. New Insights into the Genetic Diversity of the Stone Crayfish: Taxonomic and Conservation Implications. BMC Evol. Biol. 2020, 20, 146. [Google Scholar] [CrossRef]
- Bromham, L.; Woolfit, M. Explosive Radiations and the Reliability of Molecular Clocks: Island Endemic Radiations as a Test Case. Syst. Biol. 2004, 53, 758–766. [Google Scholar] [CrossRef] [Green Version]
- Warnock, R.C.M.; Yang, Z.; Donoghue, P.C.J. Testing the Molecular Clock Using Mechanistic Models of Fossil Preservation and Molecular Evolution. Proc. R. Soc. B. 2017, 284, 20170227. [Google Scholar] [CrossRef] [Green Version]
- Buj, I.; Šanda, R.; Marčić, Z.; Ćaleta, M.; Mrakovčić, M. Combining Morphology and Genetics in Resolving Taxonomy—A Systematic Revision of Spined Loaches (Genus Cobitis; Cypriniformes, Actinopterygii) in the Adriatic Watershed. PLoS ONE 2014, 9, e99833. [Google Scholar] [CrossRef] [Green Version]
- Previšić, A.; Schnitzler, J.; Kučinić, M.; Graf, W.; Ibrahimi, H.; Kerovec, M.; Pauls, S.U. Microscale Vicariance and Diversification of Western Balkan Caddisflies Linked to Karstification. Freshw. Sci. 2014, 33, 250–262. [Google Scholar] [CrossRef] [Green Version]
- Bilandžija, H.; Morton, B.; Podnar, M.; Ćetković, H. Evolutionary History of Relict Congeria (Bivalvia: Dreissenidae): Unearthing the Subterranean Biodiversity of the Dinaric Karst. Front. Zool. 2013, 10, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genus | Newly Sequenced | Previous Studies | Alignment Length (bp) |
|---|---|---|---|
| Deliminichthys | 15 | 344 | 1006 |
| Phoxinus | 2 | 1006 | 1092 |
| Phoxinellus | 9 | 8 | 1121 |
| Telestes | 7 | 190 | 1043 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reier, S.; Bogutskaya, N.; Palandačić, A. Comparative Phylogeography of Phoxinus, Delminichthys, Phoxinellus and Telestes in Dinaric Karst: Which Factors Have Influenced Their Current Distributions? Diversity 2022, 14, 526. https://doi.org/10.3390/d14070526
Reier S, Bogutskaya N, Palandačić A. Comparative Phylogeography of Phoxinus, Delminichthys, Phoxinellus and Telestes in Dinaric Karst: Which Factors Have Influenced Their Current Distributions? Diversity. 2022; 14(7):526. https://doi.org/10.3390/d14070526
Chicago/Turabian StyleReier, Susanne, Nina Bogutskaya, and Anja Palandačić. 2022. "Comparative Phylogeography of Phoxinus, Delminichthys, Phoxinellus and Telestes in Dinaric Karst: Which Factors Have Influenced Their Current Distributions?" Diversity 14, no. 7: 526. https://doi.org/10.3390/d14070526
APA StyleReier, S., Bogutskaya, N., & Palandačić, A. (2022). Comparative Phylogeography of Phoxinus, Delminichthys, Phoxinellus and Telestes in Dinaric Karst: Which Factors Have Influenced Their Current Distributions? Diversity, 14(7), 526. https://doi.org/10.3390/d14070526