
Fungal Flora in Adult Females of the Rearing Population of Ambrosia Beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does It Differ from the Wild Population?


Abstract
:1. Introduction
2. Materials and Methods
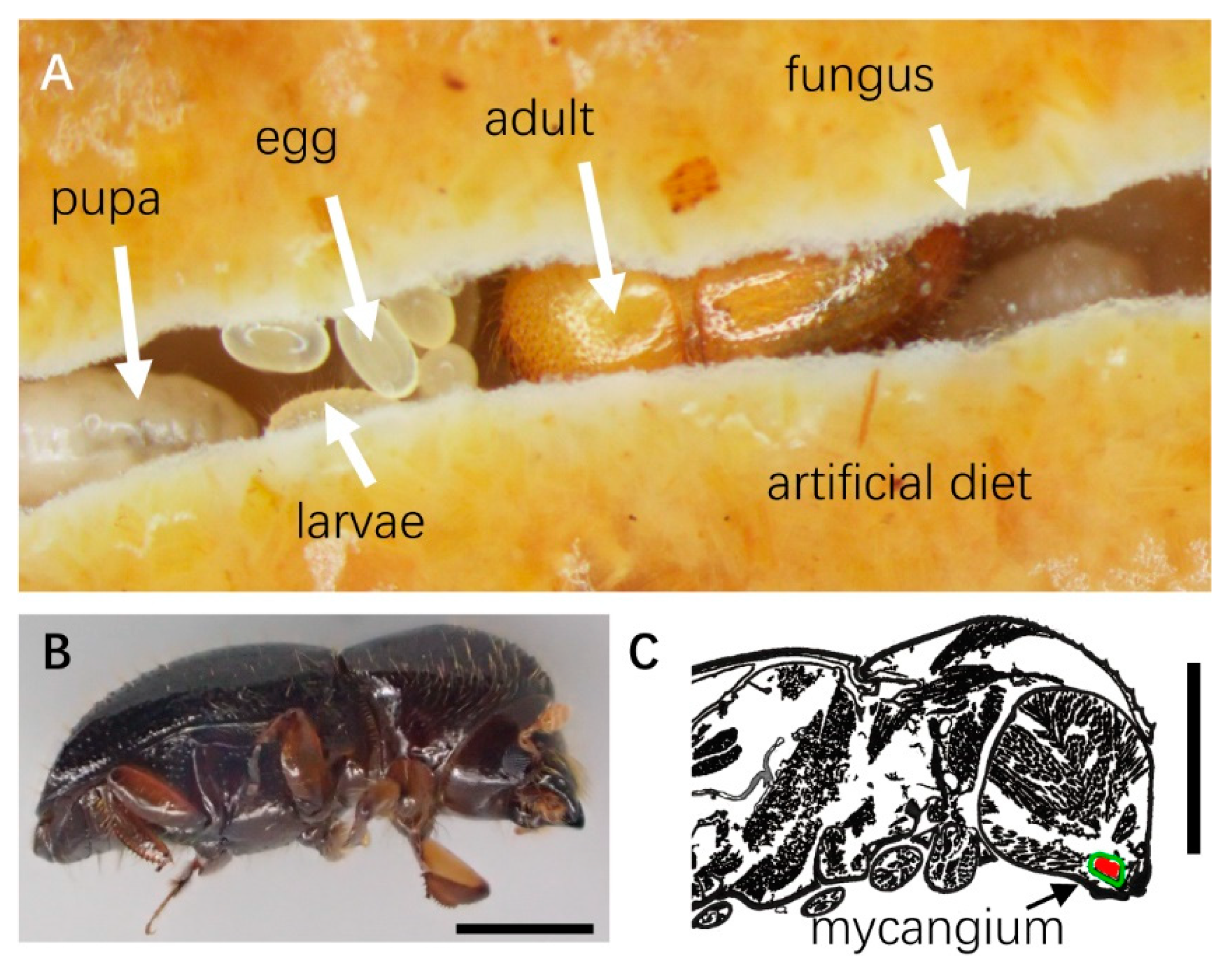
2.1. Ambrosia Beetle Collection
2.2. Fungal Isolation and Culturing
2.3. Fungal Identification
2.4. DNA Extracting and Sequencing
2.5. Phylogenetic Analyses
2.6. Quantitative Culturing of Fungi
2.7. Fungal Growth
2.8. Subsection
3. Results
3.1. Fungal Flora
3.2. Phylogenetic Analyses
3.3. Relative Dominance and Frequency of Occurrence
3.4. Quantitative Culturing
3.5. Fungal Growth
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, D.; Daugherty, S.C.; van Aken, S.E.; Pai, G.H.; Watkins, K.L.; Khouri, H.; Tallon, L.J.; Zaborsky, J.M.; Dunbar, H.E.; Tran, P.L.; et al. Metabolic Complementarity and Genomics of the Dual Bacterial Symbiosis of Sharpshooters. PLoS Biol. 2006, 4, e188. [Google Scholar] [CrossRef]
- Hulcr, J.; Stelinski, L.L. The Ambrosia Symbiosis: From Evolutionary Ecology to Practical Management. Annu. Rev. Entomol. 2017, 62, 285–303. [Google Scholar] [CrossRef] [Green Version]
- Moran, N.A.; Degnan, P.H.; Santos, S.R.; Dunbar, H.E.; Ochman, H. The Players in a Mutualistic Symbiosis: Insects, Bacteria, Viruses, and Virulence Genes. Proc. Natl Acad. Sci. USA 2005, 102, 16919–16926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, L.R. Ambrosia Beetles and Their Associated Fungi: Research Trends and Techniques. Proc. Indian Acad. Sci. 1985, 94, 137–148. [Google Scholar] [CrossRef]
- Li, Y.; Ruan, Y.Y.; Stanley, E.L.; Skelton, J.; Hulcr, J. Plasticity of Mycangia in Xylosandrus Ambrosia Beetles. Insect Sci. 2019, 26, 732–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joseph, R.; Keyhani, N.O. Fungal Mutualisms and Pathosystems: Life and Death in the Ambrosia Beetle Mycangia. Appl. Microbiol. Biotechnol. 2021, 105, 3393–3410. [Google Scholar] [CrossRef]
- Bentz, B.J.; Six, D.L. Ergosterol Content of Fungi Associated with Dendroctonus ponderosae and Dendroctonus rufipennis (Coleoptera: Curculionidae, Scolytinae). Ann. Entomol. Soc. Am. 2006, 99, 189–194. [Google Scholar] [CrossRef]
- Klepzig, K.D.; Adams, A.S.; Handelsman, J.; Raffa, K.F. Symbioses: A Key Driver of Insect Physiological Processes, Ecological Interactions, Evolutionary Diversification, and Impacts on Humans. Environ. Entomol. 2009, 38, 67–77. [Google Scholar] [CrossRef] [Green Version]
- Endoh, R.; Suzuki, M.; Okada, G.; Takeuchi, Y.; Futai, K. Fungus Symbionts Colonizing the Galleries of the Ambrosia Beetle Platypus quercivorus. Microb. Ecol. 2011, 62, 106–120. [Google Scholar] [CrossRef]
- Li, Y.; Simmons, D.R.; Bateman, C.C.; Short, D.P.; Kasson, M.T.; Rabaglia, R.J.; Hulcr, J. New Fungus-Insect Symbiosis: Culturing, Molecular, and Histological Methods Determine Saprophytic Polyporales Mutualists of Ambrosiodmus Ambrosia Beetles. PLoS ONE 2015, 10, e0137689. [Google Scholar] [CrossRef] [Green Version]
- Kirkendall, L.R.; Biedermann, P.H.W.; Jordal, B.H. Evolution and Diversity of Bark and Ambrosia Beetles. In Bark Beetles: Biology and Ecology of Native and Invasive Species; Vega, F.E., Hofstetter, R.W., Eds.; Elsevier: London, UK, 2015; pp. 85–156. [Google Scholar]
- Kajimura, H.; Hijii, N. Reproduction and Resource Utilization of the Ambrosia Beetle, Xylosandrus mutilatus, in Field and Experimental Populations. Entomol. Exp. Appl. 1994, 71, 121–132. [Google Scholar] [CrossRef]
- Peer, K.; Taborsky, M. Outbreeding Depression, but No Inbreeding Depression in Haplodiploid Ambrosia Beetles with Regular Sibling Mating. Evolution 2005, 59, 317–323. [Google Scholar] [CrossRef] [PubMed]
- Biedermann, P.H.W.; Klepzig, K.D.; Taborsky, M. Fungus Cultivation by Ambrosia Beetles: Behavior and Laboratory Breeding Success in Three Xyleborine Species. Environ. Entomol. 2009, 38, 1096–1105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roy, K.; Jaenecke, K.A.; Peck, R.W. Ambrosia beetle (Coleoptera: Curculionidae) communities and frass production in ʻŌhiʻa (Myrtales: Myrtaceae) infected with Ceratocystis (Microascales: Ceratocystidaceae) fungi responsible for rapid ʻŌhiʻa death. Environ. Entomol. 2020, 49, 1345–1354. [Google Scholar] [CrossRef]
- Jiang, Z.R.; Kajimura, H. Earwig Preying on Ambrosia Beetle: Evaluating Predatory Process and Prey Preference. J. Appl. Entomol. 2020, 144, 743–750. [Google Scholar] [CrossRef]
- Mizuno, T.; Kajimura, H. Effects of Ingredients and Structure of Semi-artificial Diet on the Reproduction of an Ambrosia Beetle, Xyleborus pfeili (Ratzeburg) (Coleoptera: Curculionidae: Scolytinae). Appl. Entomol. Zool. 2009, 44, 363–370. [Google Scholar] [CrossRef]
- Cooperband, M.F.; Stouthamer, R.; Carrillo, D.; Eskalen, A.; Thibault, T.; Cossé, A.A.; Castrillo, L.A.; Vandenberg, J.D.; Rugman-Jones, P.F. Biology of Two Members of the Euwallacea fornicatus Species Complex (Coleoptera: Curculionidae: Scolytinae), Recently Invasive in the U.S.A., Reared on an Ambrosia Beetle Artificial Diet. Agric. For. Entomol. 2016, 18, 223–237. [Google Scholar] [CrossRef]
- Lake Maner, M.; Hanula, J.L.; Kristine Braman, S. Rearing Redbay Ambrosia Beetle, Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae), on Semi-artificial Media. Fla. Entomol. 2013, 96, 1042–1051. [Google Scholar] [CrossRef] [Green Version]
- Nuotclà, J.A.; Biedermann, P.H.W.; Taborsky, M. Pathogen Defence Is a Potential Driver of Social Evolution in Ambrosia Beetles. Proc. Biol. Sci. 2019, 286, 20192332. [Google Scholar] [CrossRef]
- Freeman, S.; Sharon, M.; Dori-Bachash, M.; Maymon, M.; Belausov, E.; Maoz, Y.; Margalit, O.; Protasov, A.; Mendel, Z. Symbiotic Association of Three Fungal Species Throughout the Life Cycle of the Ambrosia Beetle Euwallacea nr. fornicatus. Symbiosis 2016, 68, 115–128. [Google Scholar] [CrossRef]
- Carrillo, J.D.; Dodge, C.; Stouthamer, R.; Eskalen, A. Fungal Symbionts of the Polyphagous and Kuroshio Shot Hole Borers (Coleoptera: Scolytinae, Euwallacea spp.) in California Can Support Both Ambrosia Beetle Systems on Artificial Media. Symbiosis 2020, 80, 155–168. [Google Scholar] [CrossRef]
- Samuelson, G.A. A Synopsis of Hawaiian Xyleborini (Coleoptera: Scolytidae). Pac. Insects. 1981, 23, 50–92. [Google Scholar]
- Landi, L.; Braccini, C.L.; Knížek, M.; Pereyra, V.A.; Marvaldi, A.E. A Newly Detected Exotic Ambrosia Beetle in Argentina: Euwallacea interjectus (Coleoptera: Curculionidae: Scolytinae). Fla. Entomol. 2019, 102, 240–242. [Google Scholar] [CrossRef]
- Aoki, T.; Smith, J.A.; Kasson, M.T.; Freeman, S.; Geiser, D.M.; Geering, A.D.W.; O’Donnell, K. Three Novel Ambrosia Fusarium Clade species Producing Clavate Macroconidia Known (F. floridanum and F. obliquiseptatum) or Predicted (F. tuaranense) to Be Farmed by Euwallacea spp. (Coleoptera: Scolytinae) on Woody Hosts. Mycologia 2019, 111, 919–935. [Google Scholar] [CrossRef]
- Wang, Z.; Li, Y.; Ernstsons, A.S.; Sun, R.; Hulcr, J.; Gao, L. The Infestation and Habitat of the Ambrosia Beetle Euwallacea interjectus (Coleoptera: Curculionidae: Scolytinae) in the Riparian Zone of Shanghai, China. Agric. For. Entomol. 2021, 23, 104–109. [Google Scholar] [CrossRef]
- Kajitani, Y. The Possibility of Transmission of Fig Ceratocystis canker Disease by an Ambrosia Beetle (Xyleborus interjectus Eichhoff). Ann. Phytopathol. Soc. Jpn. 1996, 62, 275. [Google Scholar]
- Kajitani, Y. The Dispersal Period of the Xyleborus interjectus (Coleoptera, Scolytidae), a Vector of the Fig Ceratocystis canker, and the Organ Carrying the Causal Fungus. Ann. Phytopathol. Soc. Jpn. 1999, 65, 377. [Google Scholar]
- Kajitani, Y.; Masuya, H. Ceratocystis ficicola sp. nov., a Causal Fungus of Fig Canker in Japan. Mycoscience 2011, 52, 349–353. [Google Scholar] [CrossRef]
- Nitta, H.; Morita, T.; Wakasaki, Y.; Kakogawa, K. Relationship Between Ceratocystis canker and Ambrosia Beetle in Fig Orchards. Ann. Rept. Kansai Pl. Prot. Kansai Pl. Prot. 2005, 47, 95–98. [Google Scholar] [CrossRef] [Green Version]
- Morita, T.; Hara, H.; Mise, D.; Jikumaru, S. A Case Study of Ceratocystis canker Epidemic in Relation with Euwallacea interjectus Infestation. Ann. Rept Kansai Pl. Prot. Kansai Pl. Prot. 2012, 54, 29–34. [Google Scholar] [CrossRef] [Green Version]
- Kajii, C.; Morita, T.; Jikumaru, S.; Kajimura, H.; Yamaoka, Y.; Kuroda, K. Xylem Dysfunction in Ficus carica Infected with Wilt Fungus Ceratocystis ficicola and the Role of the Vector Beetle Euwallacea interjectus. IAWA J. 2013, 34, 301–312. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Z.R.; Kinoshita, S.I.; Sasaki, O.; Cognato, A.I.; Kajimura, H. Non-destructive Observation of the Mycangia of Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae) Using X-ray Computed Tomography. Entomol. Sci. 2019, 22, 173–181. [Google Scholar] [CrossRef]
- Jiang, Z.R.; Masuya, H.; Kajimura, H. Novel Symbiotic Association Between Euwallacea Ambrosia Beetle and Fusarium Fungus on Fig Trees in Japan. Front. Microbiol. 2021, 12, 725210. [Google Scholar] [CrossRef]
- Bateman, C.; Huang, Y.T.; Simmons, D.R.; Kasson, M.T.; Stanley, E.L.; Hulcr, J. Ambrosia Beetle Premnobius cavipennis (Scolytinae: Ipini) Carries Highly Divergent Ascomycotan Ambrosia Fungus, Afroraffaelea ambrosiae gen. nov. et sp. nov. (Ophiostomatales). Fungal Ecol. 2017, 25, 41–49. [Google Scholar] [CrossRef] [Green Version]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- O’Donnell, K.; Kistler, H.C.; Cigelnik, E.; Ploetz, R.C. Multiple Evolutionary Origins of the Fungus Causing Panama Disease of Banana: Concordant Evidence from Nuclear and Mitochondrial Gene Genealogies. Proc. Natl Acad. Sci. USA 1998, 95, 2044–2049. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Madan, A. CAP3: A DNA Sequence Assembly Program. Genome Res. 1999, 9, 868–877. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A Fast and Lightweight Alignment Viewer and Editor for Large Data Sets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef]
- Nguyen, L.T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Kalyaanamoorthy, S.; Minh, B.Q.; Wong, T.K.F.; Von Haeseler, A.; Jermiin, L.S. ModelFinder: Fast Model Selection for Accurate Phylogenetic Estimates. Nat. Methods 2017, 14, 587–589. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Ruan, Y.Y.; Kasson, M.T.; Stanley, E.L.; Gillett, C.P.D.T.; Johnson, A.J.; Zhang, M.; Hulcr, J. Structure of the Ambrosia Beetle (Coleoptera: Curculionidae) Mycangia Revealed Through Micro-computed Tomography. J. Insect Sci. 2018, 18, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batra, L.R. Ambrosia Fungi: Extent of Specificity to Ambrosia Beetles. Science 1966, 153, 193–195. [Google Scholar] [CrossRef] [PubMed]
- Kühnholz, S.; Borden, J.H.; Uzunovic, A. Secondary Ambrosia Beetles in Apparently Healthy Trees: Adaptations, Potential Causes and Suggested Research. Integr. Pest. Manag. Rev. 2001, 6, 209–219. [Google Scholar] [CrossRef]
- Marini, L.; Haack, R.A.; Rabaglia, R.J.; Toffolo, E.P.; Battisti, A.; Faccoli, M. Exploring Associations Between International Trade and Environmental Factors with Establishment Patterns of Exotic Scolytinae. Biol. Invasions 2011, 13, 2275–2288. [Google Scholar] [CrossRef]
- Rassati, D.; Faccoli, M.; Battisti, A.; Marini, L. Habitat and Climatic Preferences Drive Invasions of Non-native Ambrosia Beetles in Deciduous Temperate Forests. Biol. Invasions 2016, 18, 2809–2821. [Google Scholar] [CrossRef]
- Umeda, C.; Paine, T. Temperature Can Limit the Invasion Range of the Ambrosia Beetle Euwallacea nr. fornicatus. Agr. For. Entomol. 2019, 21, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gugliuzzo, A.; Criscione, G.; Biondi, A.; Aiello, D.; Vitale, A.; Polizzi, G.; Tropea Garzia, G. Seasonal Changes in Population Structure of the Ambrosia Beetle Xylosandrus compactus and Its Associated Fungi in a Southern Mediterranean Environment. PLoS ONE 2020, 15, e0239011. [Google Scholar] [CrossRef]
- Formby, J.P.; Rodgers, J.C.; Koch, F.H.; Krishnan, N.; Duerr, D.A.; Riggins, J.J. Cold Tolerance and Invasive Potential of the Redbay Ambrosia Beetle (Xyleborus glabratus) in the Eastern United States. Biol. Invasions 2018, 20, 995–1007. [Google Scholar] [CrossRef]
- Norris, D.M.; Baker, J.M. A Minimal Nutritional Substrate Required by Fusarium solani to Fulfill Its Mutualistic Relationship with Xyleborus ferrugineus. Ann. Entomol. Soc. Am. 1968, 61, 1473–1475. [Google Scholar] [CrossRef]
- Mendel, Z.; Protasov, A.; Sharon, M.; Zveibil, A.; Ben Yehuda, S.B.; O’Donnell, K.; Rabaglia, R.; Wysoki, M.; Freeman, S. An Asian Ambrosia Beetle Euwallacea fornicatus and Its Novel Symbiotic Fungus Fusarium sp. Pose a Serious Threat to the Israeli Avocado Industry. Phytoparasitica 2012, 40, 235–238. [Google Scholar] [CrossRef]
- Kasson, M.T.; O’Donnell, K.; Rooney, A.P.; Sink, S.; Ploetz, R.C.; Ploetz, J.N.; Konkol, J.L.; Carrillo, D.; Freeman, S.; Mendel, Z.; et al. An Inordinate Fondness for Fusarium: Phylogenetic Diversity of Fusaria Cultivated by Ambrosia Beetles in the Genus Euwallacea on Avocado and Other Plant Hosts. Fungal Genet. Biol. 2013, 56, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Egonyu, J.P.; Torto, B. Responses of the Ambrosia Beetle Xylosandrus compactus (Coleoptera: Curculionidae: Scolytinae) to Volatile Constituents of Its Symbiotic Fungus Fusarium solani (Hypocreales: Nectriaceae). Arthropod Plant Interact. 2018, 12, 9–20. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Crous, P.W. Removing Chaos from Confusion: Assigning Names to Common Human and Animal Pathogens in Neocosmospora. Persoonia 2018, 41, 109–129. [Google Scholar] [CrossRef] [PubMed]
- Al-Hatmi, A.M.S.; Ahmed, S.A.; van Diepeningen, A.D.; Drogari-Apiranthitou, M.; Verweij, P.E.; Meis, J.F.; De Hoog, G.S. Fusarium metavorans sp. nov.: The Frequent Opportunist “FSSC6”. Med. Mycol. 2018, 56, 144–152. [Google Scholar] [CrossRef]
- Sandoval-Denis, M.; Lombard, L.; Crous, P.W. Back to the Roots: A Reappraisal of Neocosmospora. Persoonia 2019, 43, 90–185. [Google Scholar] [CrossRef]
- Na, F.; Carrillo, J.D.; Mayorquin, J.S.; Ndinga-Muniania, C.; Stajich, J.E.; Stouthamer, R.; Huang, Y.T.; Lin, Y.T.; Chen, C.Y.; Eskalen, A. Two Novel Fungal Symbionts Fusarium kuroshium sp. nov. and Graphium kuroshium sp. nov. of Kuroshio Shot Hole Borer (Euwallacea sp. nr. fornicatus) Cause Fusarium Dieback on Woody Host Species in California. Plant Dis. 2018, 102, 1154–1164. [Google Scholar] [CrossRef] [Green Version]
- Aoki, T.; Liyanage, P.N.H.; Konkol, J.L.; Ploetz, R.C.; Smith, J.A.; Kasson, M.T.; Freeman, S.; Geiser, D.M.; O’Donnell, K. Three Novel Ambrosia Fusarium Clade species Producing Multiseptate “Dolphin-Shaped” Conidia, and an Augmented Description of Fusarium kuroshium. Mycologia 2021, 113, 1089–1109. [Google Scholar] [CrossRef]
- Eskalen, A.; Stouthamer, R.; Lynch, S.C.; Rugman-Jones, P.F.; Twizeyimana, M.; Gonzalez, A.; Thibault, T. Hosts. Host Range of Fusarium Dieback and Its Ambrosia Beetle (Coleoptera: Scolytinae) Vector in Southern California. Plant Dis. 2013, 97, 938–951. [Google Scholar] [CrossRef] [Green Version]
- Freeman, S.; Sharon, M.; Maymon, M.; Mendel, Z.; Protasov, A.; Aoki, T.; Eskalen, A.; O’Donnell, K. Fusarium euwallaceae sp. nov.—a Symbiotic Fungus of Euwallacea sp., an Invasive Ambrosia Beetle in Israel and California. Mycologia 2013, 105, 1595–1606. [Google Scholar] [CrossRef] [Green Version]
- Morales-Ramos, J.A.; Rojas, M.G.; Sittertz-Bhatkar, H.; Saldaña, G. Symbiotic Relationship Between Hypothenemus hampei (Coleoptera: Scolytidae) and Fusarium solani (Moniliales: Tuberculariaceae). Ann. Entomol. Soc. Am. 2000, 93, 541–547. [Google Scholar] [CrossRef]
- Masuya, H.; Kajimura, H.; Tomisawa, N.; Yamaoka, Y. Fungi Associated with Scolytogenes birosimensis (Coleoptera: Curculionidae) Infesting Pittosporum tobira. Environ. Entomol. 2012, 41, 255–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kuroda, K.; Chuma, I.; Kihara, T.; Murakami, T.; Takashina, K.; Hiraoka, D.; Kameyama, N. First Report of Fusarium solani species Complex as a Causal Agent of Erythrina variegata Decline and Death After Gall Formation by Quadrastichus erythrinae on Okinawa Island, Japan. J. Gen. Plant Pathol. 2017, 83, 344–357. [Google Scholar] [CrossRef] [Green Version]
- Takashina, K.; Chuma, I.; Kajimura, H.; Kameyama, N.; Goto, C.; Kuroda, K. Pathogenicity and Distribution of Fusarium solani Isolates Associated with Erythrina Decline in Japan. Plant Dis. 2020, 104, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Yun, Y.H.; Suh, D.Y.; Yoo, H.D.; Oh, M.H.; Kim, S.H. Yeast Associated with the Ambrosia Beetle, Platypus koryoensis, the Pest of Oak Trees in Korea. Mycobiology 2015, 43, 458–466. [Google Scholar] [CrossRef] [Green Version]
- Stefanini, I. Yeast-Insect Associations: It Takes Guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Francke-Grosmann, H. Some New Aspects in Forest Entomology. Annu. Rev. Entomol. 1963, 8, 415–438. [Google Scholar] [CrossRef]
- Takagi, K. The Storage Organ of Symbiotic Fungus in the Ambrosia Beetle Xyleborus rubricollis Eichhoff: Coleoptera: Scolytidae. Appl. Entomol. Zool. 1967, 2, 168–170. [Google Scholar] [CrossRef] [Green Version]
- Cognato, A.I.; Hulcr, J.; Dole, S.A.; Jordal, B.H. Phylogeny of Haplo–Diploid, Fungus-Growing Ambrosia Beetles (Curculionidae: Scolytinae: Xyleborini) Inferred from Molecular and Morphological Data. Zool. Scr. 2011, 40, 174–186. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Suzuki, M. Phylogenetic Analysis of Ascomycete Yeasts That Form Coenzyme Q-9 and the Proposal of the New Genera Babjeviella, Meyerozyma, Millerozyma, Priceomyces, and Scheffersomyces. Mycoscience 2010, 51, 2–14. [Google Scholar] [CrossRef]
- Kurtzman, C.P.; Kurtzman, M.; Suzuki, M. The Yeasts: A Taxonomic Study, 5th ed.; Kurtzman, C.P., Fell, J.W., Boekhout, T., Eds.; Elsevier: London, UK, 2010; Volume 2011, pp. 622–623. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fungal Species | No. of Fungal Isolates and Relative Dominance (RD, %) a in Each Body Part | No. of Beetles from Which Each Fungal Species Was Isolated and Frequency of Occurrence (FO, %) b in Each Body Part | |||||
|---|---|---|---|---|---|---|---|
| Head | Thorax | Abdomen | Head | Thorax | Abdomen | ||
| Neocosmospora metavorans | 32(46.4) | 22(35.5) | 23(34.8) | 32(59.3) | 22(40.7) | 23(42.6) | |
| Fusarium sp. | 13(18.8) | 2(3.2) | 5(7.6) | 13(24.1) | 2(3.7) | 5(9.3) | |
| Meyerozyma guilliermondii | 12(17.4) | 3(4.8) | 1(1.5) | 12(22.2) | 3(5.6) | 1(1.9) | |
| Byssochlamys nivea | 5(7.2) | 18(29.0) | 17(25.8) | 5(9.3) | 18(33.3) | 17(31.5) | |
| Penicillium citrinum | 1(1.4) | 4(6.5) | 10(15.2) | 1(1.9) | 4(7.4) | 10(18.5) | |
| Paecilomyces sinensis | — | 1(1.6) | 4(6.1) | — | 1(1.9) | 4(7.4) | |
| Phaeoacremonium inflatipes | — | 1(1.6) | — | — | 1(1.9) | — | |
| Pestalotiopsis mangiferae | — | — | 1(1.5) | — | — | 1(1.9) | |
| Arthrinium sp. | — | 1(1.6) | — | — | 1(1.9) | — | |
| Fusarium tricinctum | — | 1(1.6) | — | — | 1(1.9) | — | |
| Unknown 3 | 2(2.9) | 2(3.2) | — | 2(3.7) | 2(3.7) | — | |
| Unknown 4 | — | 1(1.6) | 1(1.5) | — | 1(1.9) | 1(1.9) | |
| Unknown 5 | 4(5.8) | 6(9.7) | 3(4.5) | 4(7.4) | 6(11.1) | 3(5.6) | |
| Unknown 6 | — | — | 1(1.5) | — | — | 1(1.9) | |
| Total c | 69 | 62 | 66 | Number d | 54 | 54 | 54 |
| Beetle Individual | Visible Density (VD, CFUs/Head) a | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Neocosmospora metavorans | Meyerozyma guilliermondii | Fusarium sp. | Byssochlamys nivea | |||||||||
| Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | Mean | Min | Max | |
| A | — | — | — | 18,000 | 9500 | 23,500 | — | — | — | — | — | — |
| B | 1290 | 900 | 2000 | 270 | 50 | 400 | — | — | — | — | — | — |
| C | 360 | 200 | 600 | 115 | 50 | 200 | — | — | — | — | — | — |
| D | — | — | — | 190 | 50 | 900 | — | — | — | 1490 | 1000 | 1850 |
| E | 650 | 350 | 1000 | 200 | 100 | 250 | — | — | — | — | — | — |
| F | 1540 | 400 | 3050 | 830 | 50 | 1500 | — | — | — | — | — | — |
| G | 2020 | 1500 | 2300 | — | — | — | — | — | — | — | — | — |
| H | 2240 | 1650 | 3150 | — | — | — | — | — | — | — | — | — |
| I | 2620 | 1850 | 3300 | — | — | — | — | — | — | — | — | — |
| J | — | — | — | 27,500 | 22,500 | 43,000 | — | — | — | — | — | — |
| K | 1290 | 800 | 1500 | — | — | — | — | — | — | — | — | — |
| L | — | — | — | 140 | 50 | 250 | 920 | 550 | 1400 | — | — | — |
| M | 1700 | 1250 | 2350 | 170 | 300 | 4150 | — | — | — | — | — | — |
| N | 1790 | 1000 | 2250 | — | — | — | — | — | — | — | — | — |
| O | 1550 | 1200 | 1800 | — | — | — | — | — | — | — | — | — |
| P | 750 | 500 | 1250 | 2120 | 1800 | 2350 | — | — | — | — | — | — |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiang, Z.-R.; Masuya, H.; Kajimura, H. Fungal Flora in Adult Females of the Rearing Population of Ambrosia Beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does It Differ from the Wild Population? Diversity 2022, 14, 535. https://doi.org/10.3390/d14070535
Jiang Z-R, Masuya H, Kajimura H. Fungal Flora in Adult Females of the Rearing Population of Ambrosia Beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does It Differ from the Wild Population? Diversity. 2022; 14(7):535. https://doi.org/10.3390/d14070535
Chicago/Turabian StyleJiang, Zi-Ru, Hayato Masuya, and Hisashi Kajimura. 2022. "Fungal Flora in Adult Females of the Rearing Population of Ambrosia Beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does It Differ from the Wild Population?" Diversity 14, no. 7: 535. https://doi.org/10.3390/d14070535
APA StyleJiang, Z. -R., Masuya, H., & Kajimura, H. (2022). Fungal Flora in Adult Females of the Rearing Population of Ambrosia Beetle Euwallacea interjectus (Blandford) (Coleoptera: Curculionidae: Scolytinae): Does It Differ from the Wild Population? Diversity, 14(7), 535. https://doi.org/10.3390/d14070535