
Examination of Forest Steppe Species in the Case of Areas Where Traditional Cultivation Was Abandoned

 , and
, and
Abstract
:1. Introduction
- (1)
- How does the total species richness, relative cover and diversity of forest steppe species of abandoned land vary by former cultivation type and age group?
- (2)
- How did the relative species number and cover of habitat categories in cultivation types change after abandonment?
- (3)
- What were the typical coenogroups in the case of forest steppe species and in the studied types and age groups, and how did their proportion change after abandonment?
2. Materials and Methods
2.1. Study Site
2.2. Botanical Sampling
2.3. Data Evaluation
3. Results
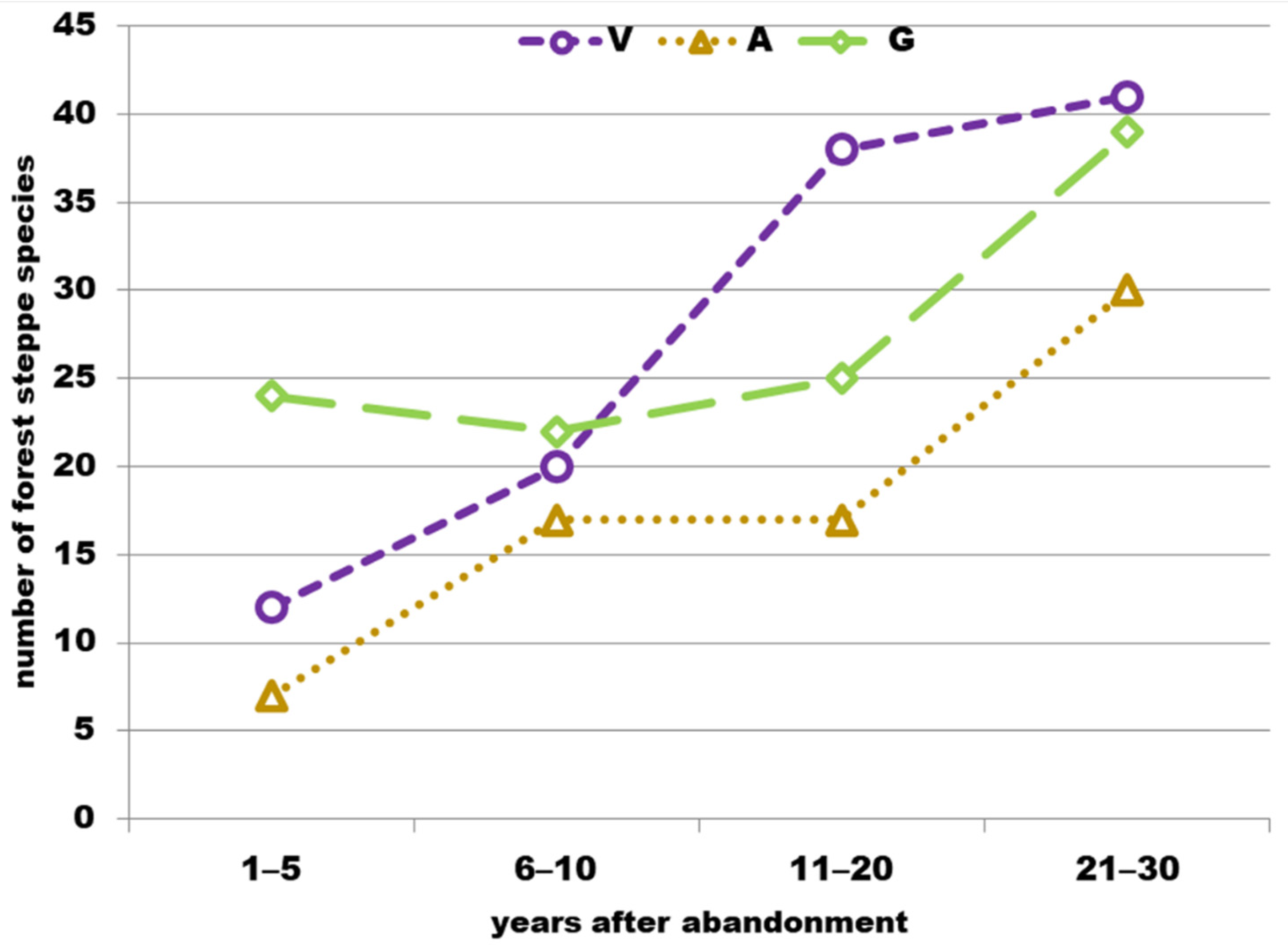
3.1. Species Richness
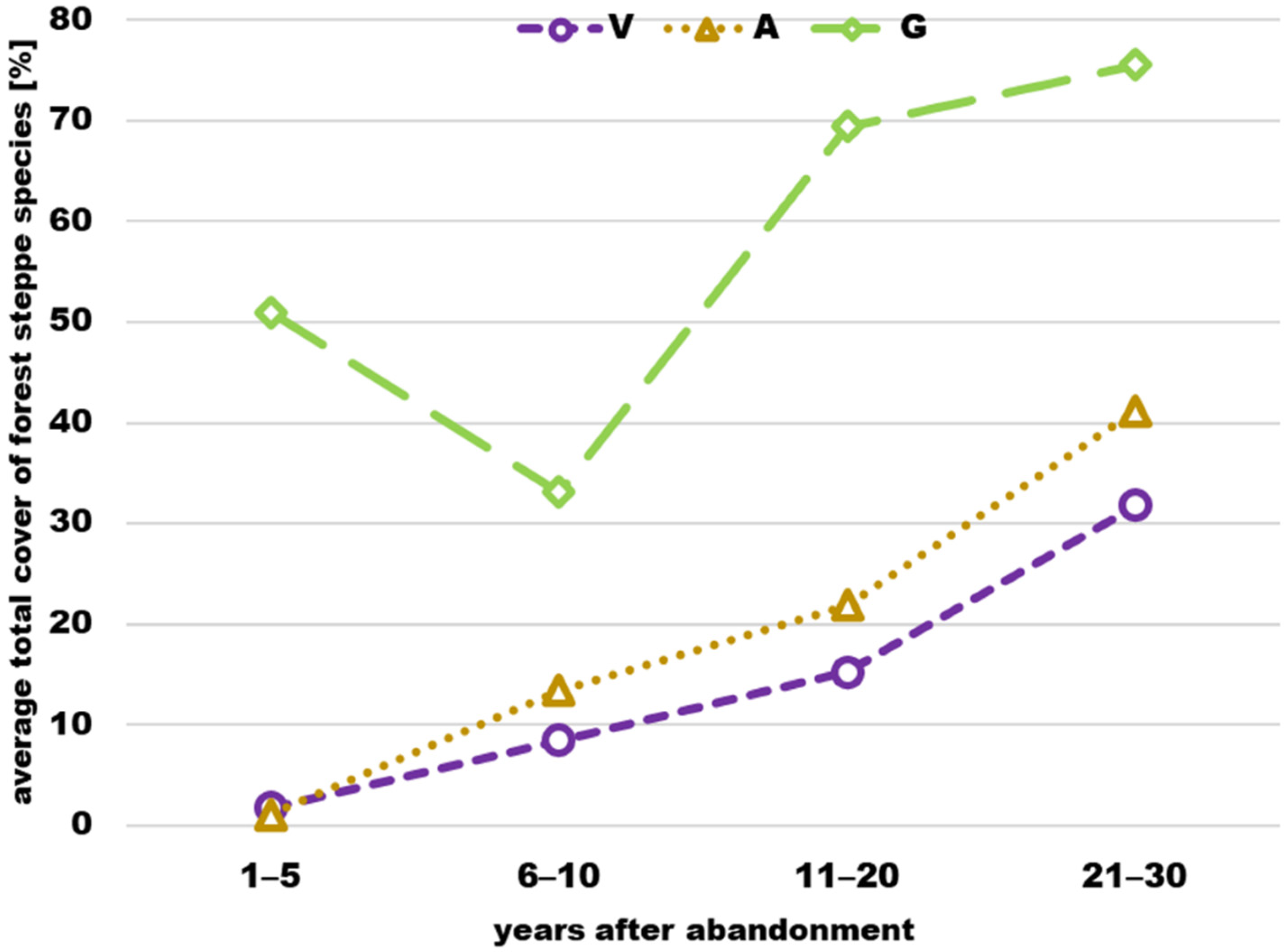
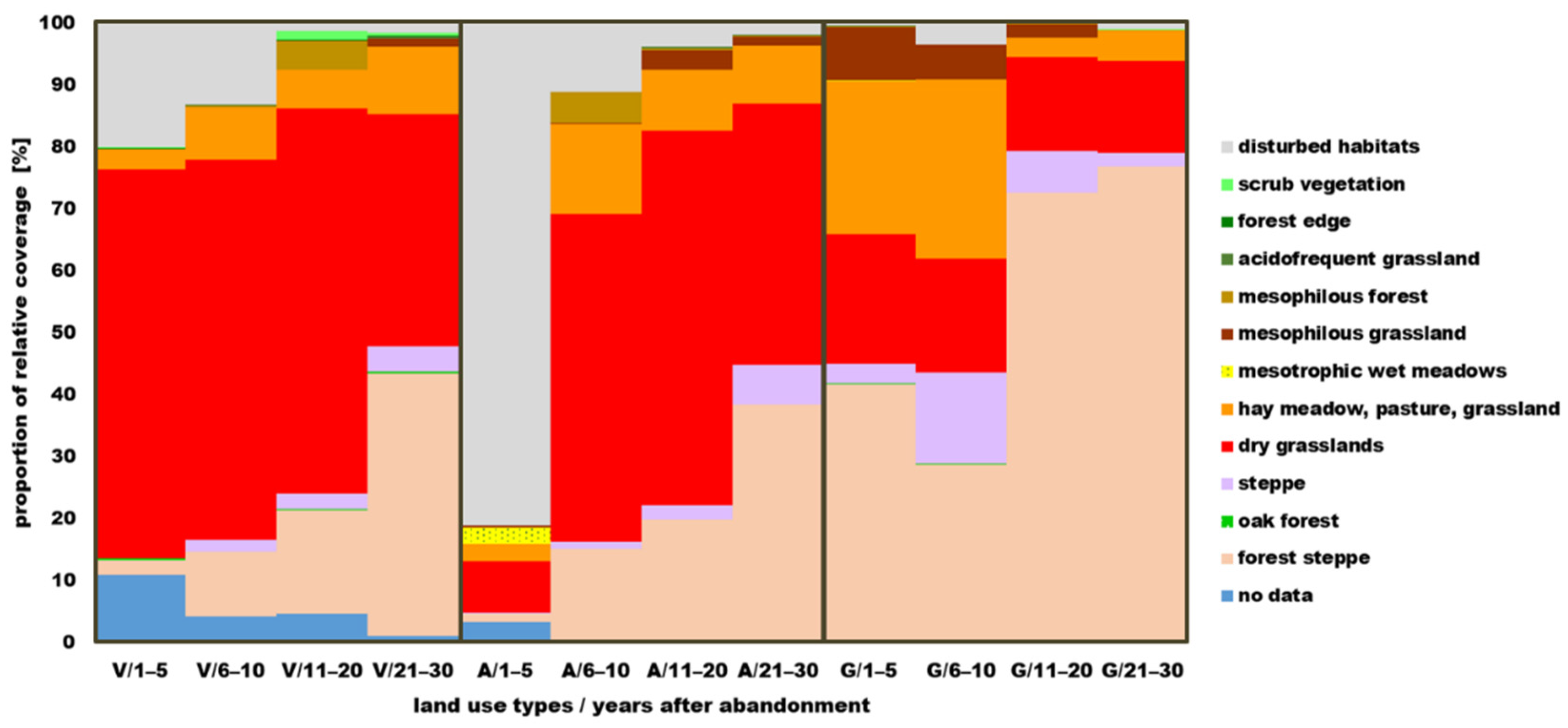
3.2. Cover
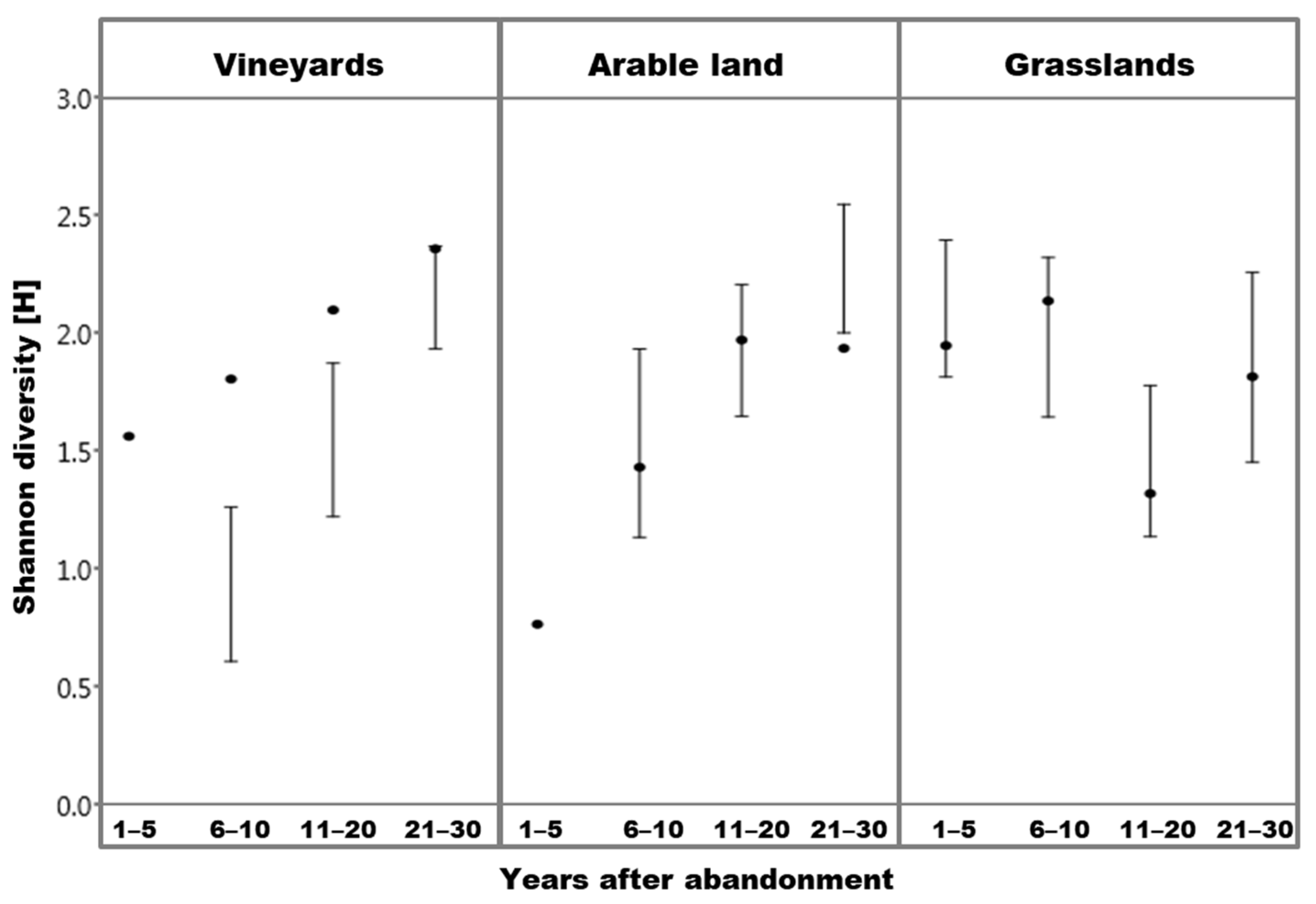
3.3. Diversity
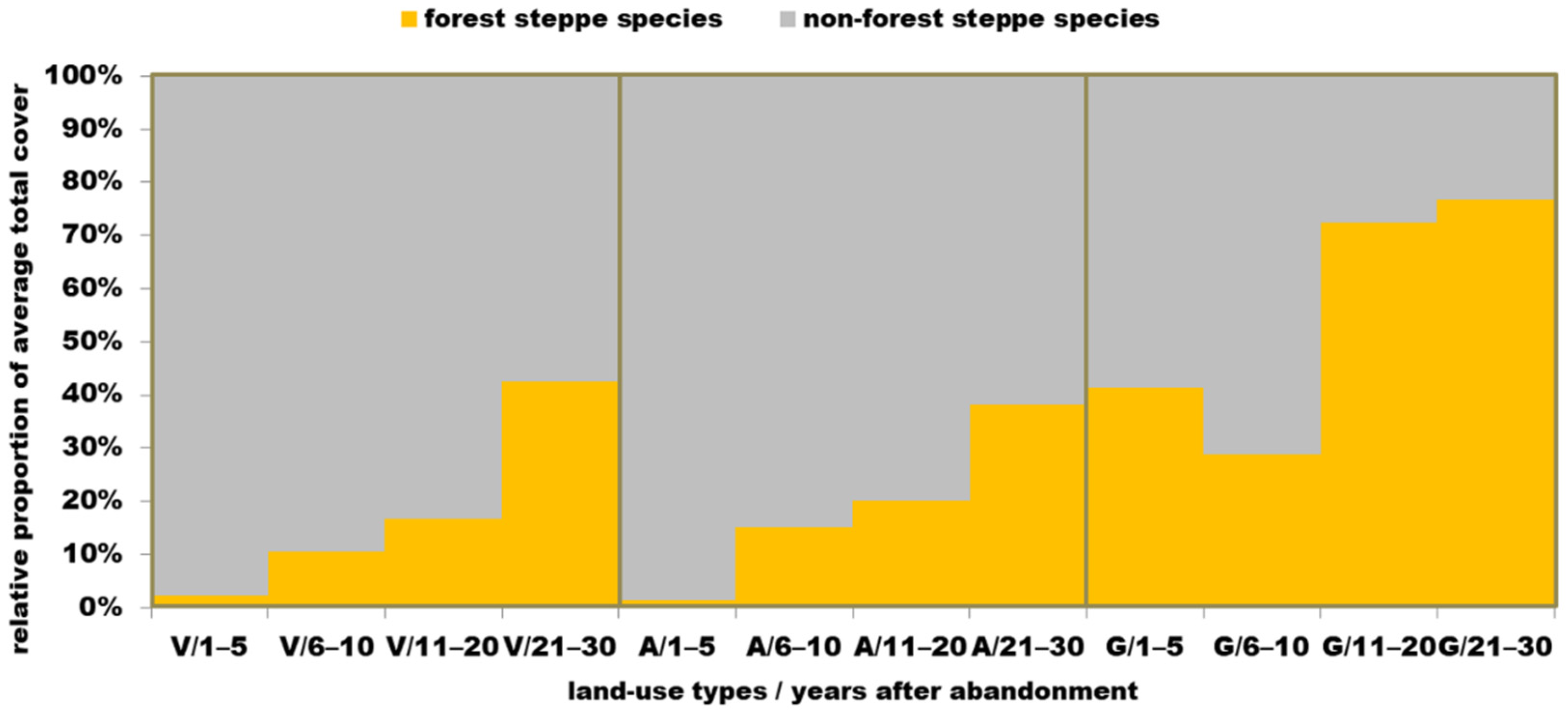
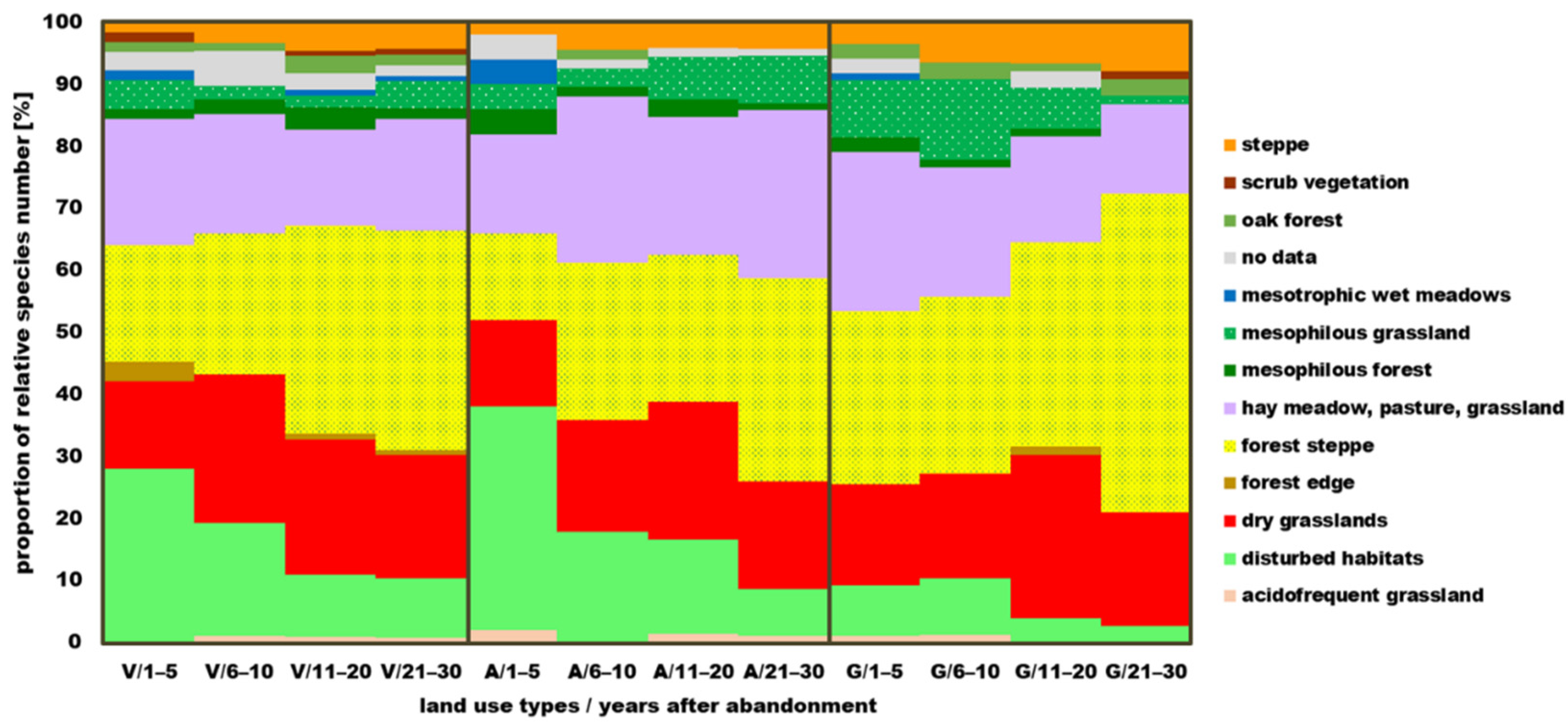
3.4. Habitat Preference
3.5. Coenosystematical Analysis
3.5.1. Coenosystematical Characteristics of Abandoned Lands
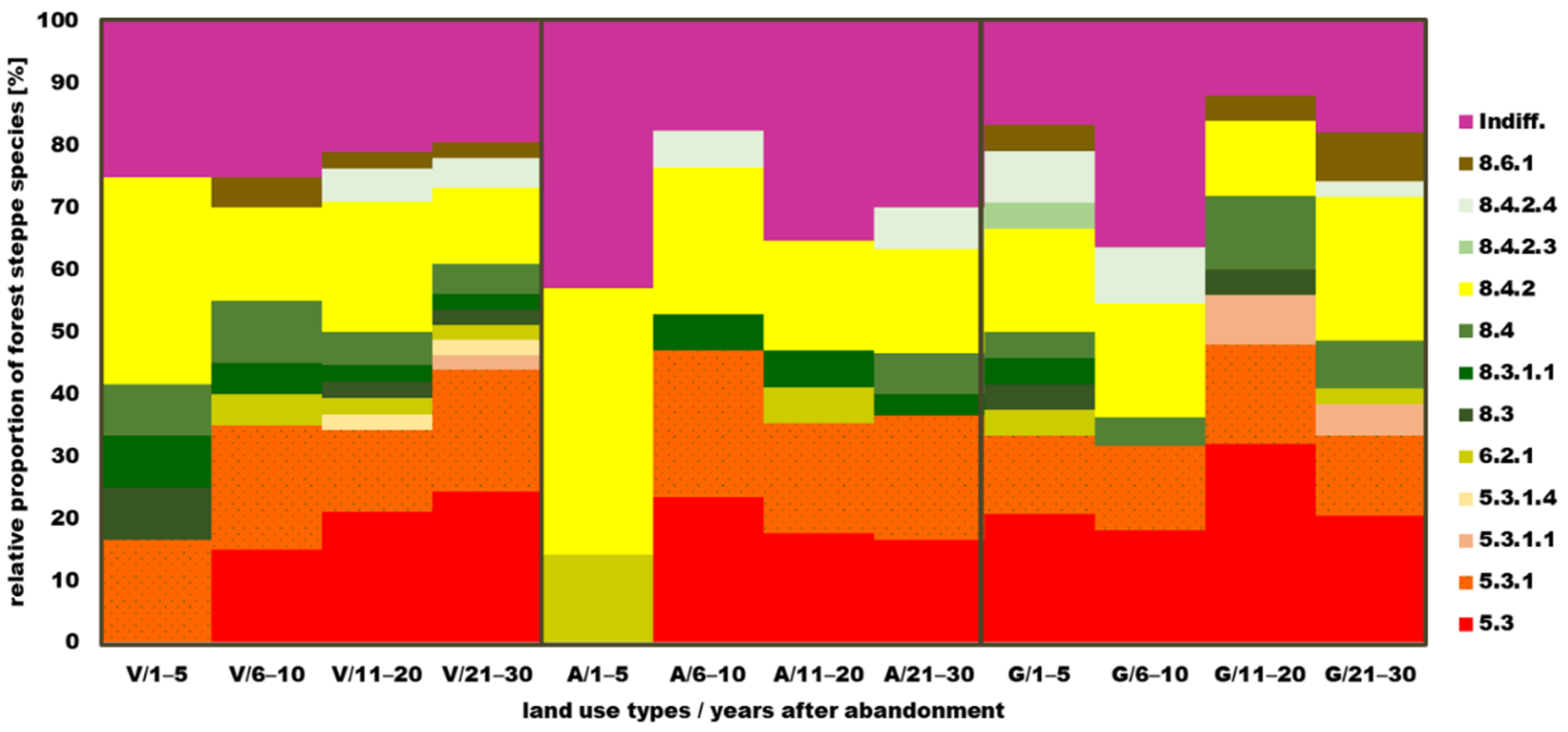
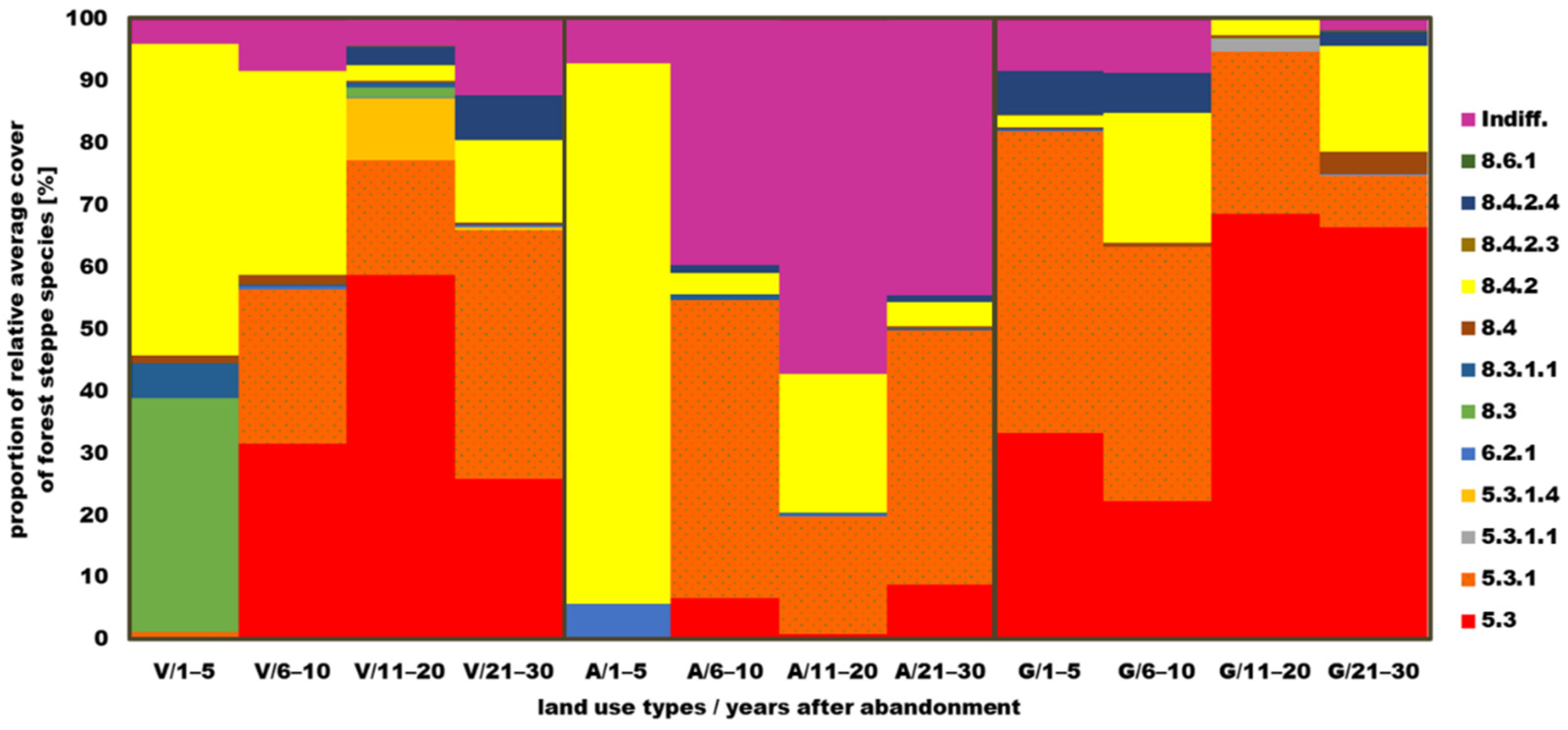
3.5.2. Coenosystematical Characteristics of Forest Steppe Species
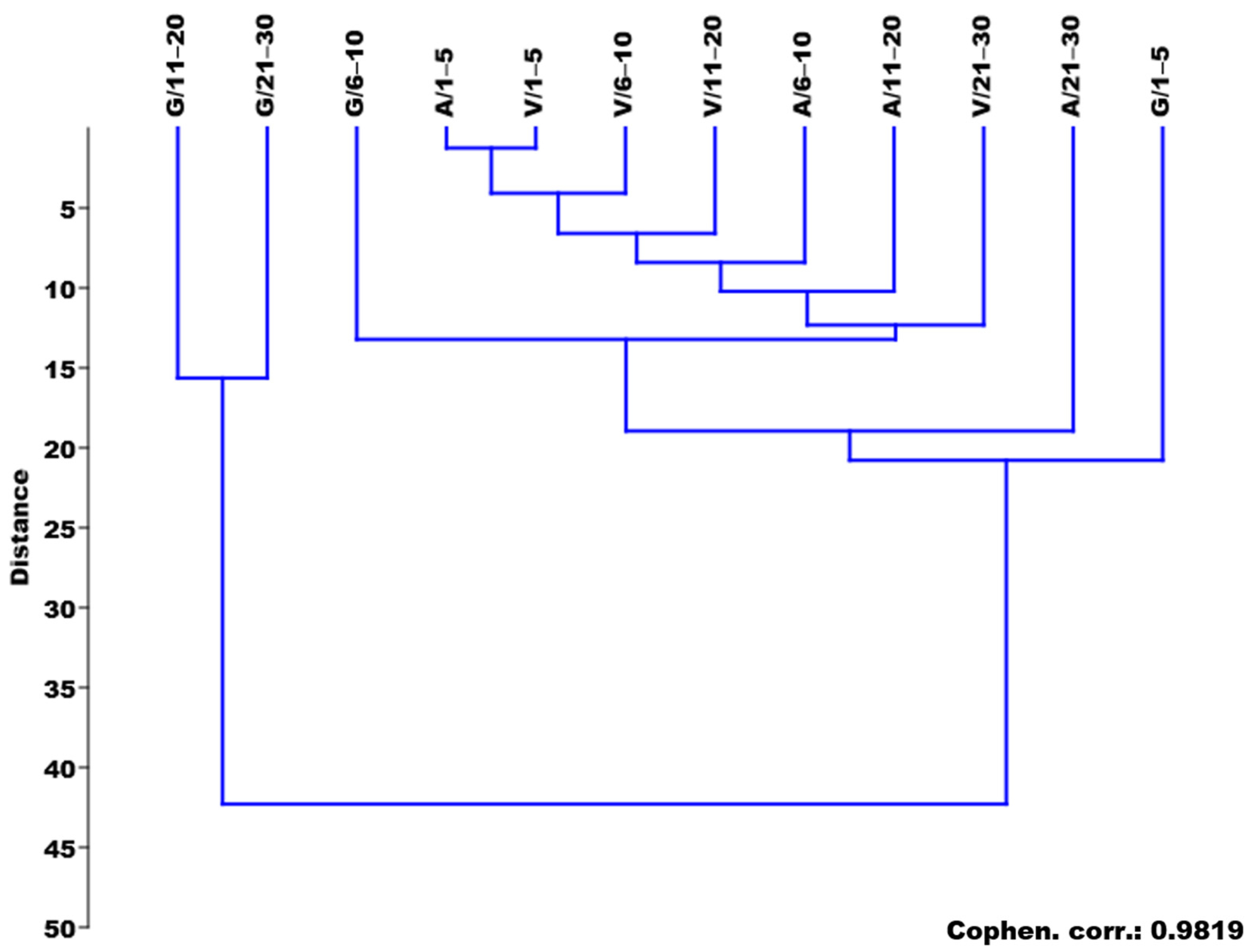
3.6. Multivariate Analysis
4. Discussion
4.1. Species Richness
4.2. Cover
4.3. Diversity
4.4. Habitat Preference
4.5. Coenosystematical Analysis
4.5.1. Coenosystematical Characteristics of Abandoned Lands
4.5.2. Coenosystematical Characteristics of Forest Steppe Species
4.6. Multivariate Analysis
4.7. Despite of Fragmentation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bátori, Z.; Erdős, L.; Kelemen, A.; Deák, B.; Valkó, O.; Gallé, R.; Bragina, T.M.; Kiss, P.J.; Kröel-Dulay, G.Y.; Tölgyesi, C.S. Diversity patterns in sandy forest-steppes: A comparative study from the western and central Palaearctic. Biodivers. Conserv. 2018, 27, 1011–1030. [Google Scholar] [CrossRef]
- Chibilyov, A. Steppe and forest-steppe. In The Physical Geography of Northern Eurasia; Shahgedanova, M., Ed.; Oxford University Press: Oxford, UK, 2002; pp. 248–266. [Google Scholar]
- Hoekstra, J.M.; Boucher, T.M.; Ricketts, T.H.; Roberts, C. Confronting a biome crisis: Global disparities of habitat loss and protection. Ecol. Lett. 2005, 8, 23–29. [Google Scholar] [CrossRef]
- Erdős, L.; Ambarli, D.; Anenkhonov, O.A.; Bátori, Z.; Cserhalmi, D.; Kiss, M.; Kröel-Dulay, G.Y.; Liu, H.; Magnes, M.; Molnár, Z.S.; et al. Where forests meet grasslands: Forest-steppes in Eurasia. Palaeartic Grassl. 2019, 40, 22–26. [Google Scholar] [CrossRef]
- Fekete, G.; Molnár, Z.S.; Magyari, E.; Somodi, I.; Varga, Z. A new framework for understanding Pannonian vegetation patterns: Regularities, deviations and uniqueness. Community Ecol. 2014, 15, 12–26. [Google Scholar] [CrossRef] [Green Version]
- Kun, A.; Rév, S.Z.; Verő, G.Y.; Nagy, I.; Demeter, L. Erdőssztyepp-erdők kezelése. In Az Erdőgazdálkodás Hatása az Erdők Biológiai Sokféleségére; Korda, M., Ed.; Duna–Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2016; pp. 501–532. [Google Scholar]
- Chytrý, M. Vegetation of the Czech Republic: Diversity, ecology, history and dynamics. Preslia 2012, 84, 427–504. [Google Scholar]
- Fekete, G.; Molnár, Z.; Kun, A.; Botta-Dukát, Z. On the structure of the Pannonian forest steppe: Grasslands on sand. Acta Zool. Hung. 2002, 48, 137–150. [Google Scholar]
- Borhidi, A. Magyarország Növénytársulásai; Akadémiai Kiadó: Budapest, Hungary, 2003; 610p, (In Hungarian). [Google Scholar] [CrossRef]
- Zólyomi, B. Budapest és környékének természetes növénytakarója. In Budapest Természeti Képe; Pécsi, M., Ed.; Akadémiai Kiadó: Budapest, Hungary, 1958; pp. 509–642. (In Hungarian) [Google Scholar]
- Borhidi, A. Klímadiagramme und klímazonale Karte Ungarns. Annales Univ. Sci. Budapest. Sec. Biol. 1961, 4, 21–50. [Google Scholar]
- Molnár, Z.; Kun, A. Alföldi erdőssztyeppmaradványok Magyarországon. WWF Füzetek 2000, 15, 1–56. (In Hungarian) [Google Scholar]
- Molnár, Z.; Varga, Z.; Molnár, A. Tiszai-Alföld és Dunai-Alföld. In Magyarország Tájainak Növényzete és Állatvilága; Fekete, G., Varga, Z., Eds.; MTA Társadalomkutató Központ: Budapest, Hungary, 2006; pp. 101–195. (In Hungarian) [Google Scholar]
- Szirmai, O.; Czóbel, S.; Nagy, J. Relationship between land use changes and forest steppe species in new forest steppe area. Cereal Res. Commun. 2005, 33, 313–316. [Google Scholar] [CrossRef]
- Kun, A.; Bölöni, J. Felnyíló erdők lágyszárú fajainak védelmi lehetőségei–különös tekintettel az erdőssztyepp-erdők megőrzésére. In Az Erdőgazdálkodás Hatása az Erdők Biológiai Sokféleségére; Korda, M., Ed.; Duna–Ipoly Nemzeti Park Igazgatóság: Budapest, Hungary, 2016; pp. 89–106. (In Hungarian) [Google Scholar]
- Illyés, E.; Bauer, N.; Botta-Dukát, Z. Classification of semi-dry grassland vegetation in Hungary. Preslia 2009, 81, 239–260. [Google Scholar]
- Zólyomi, B. Fitocenozy i lesomelirozacii obnaženij gor Budy. Acta Biol. Acad. Sci. Hung. 1950, 1, 7–67. [Google Scholar]
- Schmotzer, A.; Vojtkó, A. Investigation of Brachypodium pinnatum-dominated semi-dry grasslands in the Bükk Mountains (North-East Hungary). In Proceedings of the Research, Conservation, Management Conference, Aggtelek, Hungary, 1–5 May 1996; pp. 385–391. [Google Scholar]
- Schmotzer, A.; Vojtkó, A. Félszáraz gyepek bükki állományainak cönológiai összevetése az eredeti erdőtársulások aljnövényzetével. Kitaibelia 1997, 2, 304. (In Hungarian) [Google Scholar]
- Fekete, G.; Virágh, K.; Aszalós, R.; Orlóci, L. Landscape and coenological differentation of Brachypodium pinnatum grasslands in Hungary. Coenoses 1998, 13, 39–53. [Google Scholar]
- Fekete, G.; Virágh, K.; Aszalós, R.; Précsényi, I. Static and dynamic approaches to landscape heterogeneity in the Hungarian forest-steppe zone. J. Veg. Sci. 2000, 11, 375–382. [Google Scholar] [CrossRef]
- Varga, Z.; Varga-Sipos, J.; Orci, M.K.; Rácz, I. Felszáraz gyepek az Aggteleki karszton. In Vegetáció és Dinamizmus; Virágh, K., Kun, A., Eds.; Institute of Ecology and Botany of the Hungarian Academy of Sciences: Vácrátót, Hungary, 2000; pp. 195–238. (In Hungarian) [Google Scholar]
- Mojzes, A. A tollas szálkaperje (Brachypodium pinnatum (L.) Beauv.) és az általa dominált félszáraz gyeptársulások jellemvonásai Nyugat-Európában és hazánkban. Természetvédelmi Közlemények 2003, 10, 51–72. (In Hungarian) [Google Scholar]
- Kun, A. 6210 Meszes alapkőzetű féltermészetes száraz gyepek és cserjésedett változataik. In Natura 2000 Fajok és Élőhelyek Magyarországon; Haraszthy, L., Ed.; Pro Vértes Közalapítvány: Csákvár, Budapest, 2014; pp. 800–803. (In Hungarian) [Google Scholar]
- Willner, W.; Kuzemko, A.; Dengler, J.; Chytrý, M.; Bauer, N.; Becker, T.; Biţă-Nicolae, C.; Botta-Dukát, Z.; Čarni, A.; Csiky, J.; et al. A higher-level classification of the Pannonian and western Pontic steppe grasslands (Central and Eastern Europe). Appl. Veg. Sci. 2017, 20, 143–158. [Google Scholar] [CrossRef] [Green Version]
- Virágh, K.; Horváth, A.; Bartha, S.; Somodi, I. Kompozíciós diverzitás és términtázati rendezettség a szálkaperjés erdőssztyepprét természetközeli és zavart állományaiban. In Kutatás, Oktatás, Értékteremtés; Molnár, E., Ed.; MTA ÖBKI: Vácrátót, Hungary, 2006; pp. 89–110. (In Hungarian) [Google Scholar]
- Illyés, E. Bevezető gondolatok. In Lejtősztyepek, Löszgyepek és Erdőssztyeprétek Magyarországon; Illyés, E., Bölöni, J., Eds.; MTA ÖBKI: Budapest, Hungary, 2007; pp. 8–10. (In Hungarian) [Google Scholar]
- Illyés, E. Félszáraz irtásrétek, erdőssztyeprétek. In Lejtősztyepek, Löszgyepek és Erdőssztyeprétek Magyarországon; Illyés, E., Bölöni, J., Eds.; MTA ÖBKI: Budapest, Hungary, 2007; pp. 61–68. (In Hungarian) [Google Scholar]
- Virágh, K.; Bartha, S. Interspecific associations in different successional stages of Brachypodium pinnatum grassland after deforestation in Hungary. Tiscia 1998, 31, 3–12. [Google Scholar]
- Molnár, Z.; Horváth, F.; Botta-Dukát, Z. A parlagok kiterjedése, elterjedése és regenerációs potenciálja Magyarországon (adatok a MÉTA-adatbázisból). In Hol az a Táj Szab az Életnek Teret, Mit az Isten Csak Jókedvében Teremt; Molnár, C.S., Molnár, Z.S., Varga, A., Eds.; MTA ÖBKI: Vácrátót, Hungary, 2010; pp. 444–446. (In Hungarian) [Google Scholar]
- Szirmai, O.; Saláta, D.; Benedek, L.K.; Czóbel, S. Investigation of the Secondary Succession of Abandoned Areas from Different Cultivation in the Pannonian Biogeographic Region. Agronomy 2022, 12, 773. [Google Scholar] [CrossRef]
- Osboronova, J.; Kovarova, M.; Leps, J.; Prach, K. Succession in Abandoned Fields, Studies in Central Bohemia, Czechoslovakia; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1990; 230p. [Google Scholar]
- Pickett, S.T.A. Space-for-time substitution as an alternative to long-term studies. In Long-term Studies in Ecology: Approaches and Alternatives; Likens, G.E., Ed.; Springer: New York, NY, USA, 1989; pp. 110–135. [Google Scholar]
- Sojneková, M.; Chytrý, M. From arable land to species-rich semi-natural grasslands: Succession in abandoned fields in a dry region of central Europe. Ecol. Eng. 2015, 77, 373–381. [Google Scholar] [CrossRef]
- Jakucs, P. Die phytozönologischen Verhältnisse der Flaumeichen-Buschwälder Südost-Mitteleuropas; Akadémiai Kiadó: Budapest, Hungary, 1961; 314p. [Google Scholar]
- Zólyomi, B. Der Tatarenahorn-Eichen-Lösswald der Zonalen Waldsteppe (Acereto Tatarici-Quercetum). Acta Bot. Hung. 1957, 3, 401–429. [Google Scholar]
- Schmotzer, A. Adatok a Heves–Borsodi-sík flórájához I. Erdei, erdőssztyepp- és sztyeppfajok elterjedése. Kitaibelia 2019, 24, 16–65. [Google Scholar] [CrossRef] [Green Version]
- Szirmai, O.; Czóbel, S. Plant relations studies in agro-environmental ecosystems under different water-regimes. In Proceedings of the III. Alps-Adria Scientific Workshop, Dubrovnik, Croatia; Hidvégi, S., Gyuricza, C., Eds.; Akaprint: Budapest, Hungary, 2004; pp. 77–82. [Google Scholar]
- Szirmai, O.; Czóbel, S. Long term scale changes of land use in peasant farming of the Tardona Hills. Cereal Res. Commun. 2006, 34, 837–840. [Google Scholar] [CrossRef]
- Szirmai, O.; Czóbel, S. A Tardonai-dombság egyik vonulatának aktuális vegetációtérképe. Kitaibelia 2008, 13, 190. (In Hungarian) [Google Scholar]
- Simon, T. A Magyarországi Edényes Flóra Határozója. Harasztok-Virágos Növények; Nemzeti Tankönyvkiadó: Budapest, Hungary, 2000; 845p. (In Hungarian) [Google Scholar]
- Borhidi, A. Social behaviour types, the naturalness and relative ecological indicator values of the higher plants in the Hungarian Flora. Acta Bot. Hung. 1995, 39, 97–181. [Google Scholar]
- Hammer, Ø. PAST–PAleontological STatictics Version 4.08 Reference Manual; Natural History Museum–University of Oslo: Oslo, Norway, 1999–2022; 296p. [Google Scholar]
- Baráth, Z. Növénytakaró vizsgálatok felhagyott szőlőkben. Földrajzi Értesítő 1963, 12, 341–356. (In Hungarian) [Google Scholar]
- Házi, J.; Bartha, S. A siskanád (Calamagrostis epigeios L. Roth.) visszaszorításának lehetőségei kaszálással. Kitaibelia 2006, 11, 54. (In Hungarian) [Google Scholar]
- Házi, J.; Barth, S.; Szentes, S.; Wichmann, B.; Penksza, K. Seminatural grassland management by mowing of Calamagrostis epigejos in Hungary. Plant Biosyst. 2011, 145, 699–707. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Plant species diversity in old field successional ecosystems in Southern Illinois. Ecology 1975, 56, 485–488. [Google Scholar] [CrossRef]
- Ruprecht, E. Secondary succession in old-fields in the Transylvanian Lowland (Romania). Preslia 2005, 77, 145–157. [Google Scholar]
- Ruprecht, E. Successfully Recovered Grassland: A Promising Example from Romanian Old-Fields. Restor. Ecol. 2006, 14, 473–480. [Google Scholar] [CrossRef]
- Tyler, G. Studies in the ecology of Baltic seashore meadows II. Flora and vegetation. Opera Bot. 1969, 25, 1–101. [Google Scholar]
- Jutila, H.M. Vascular plant species richness in grazed and ungrazed coastal meadows, SW Finland. Ann. Bot. Fennici 1997, 34, 245–263. [Google Scholar]
- Chen, S.P.; Bai, Y.F.; Lin, G.H.; Liang, Y.; Han, Y.G. Effects of grazing on photosynthetic characteristics of major steppe species in the Xilin River Basin, Inner Mongolia, China. Photosynthetica 2005, 43, 559–565. [Google Scholar] [CrossRef]
- Tamm, C.O. Composition of vegetation in grazed and mown sections of a former haymeadow. Oikos 1956, 7, 144–157. [Google Scholar] [CrossRef]
- Luoto, M.; Pykala, J.; Kuussaari, M. Decline of landscape-scale habitat and species diversity after the end of cattle grazing. J. Nat. Conserv. 2003, 11, 171–178. [Google Scholar] [CrossRef]
- Pykala, J.; Luoto, M.; Heikkinen, R.K.; Kontula, T. Plant species richness and persistence of rare plants in abandoned semi-natural grasslands in northern Europe. Basic Appl. Ecol. 2005, 6, 25–33. [Google Scholar] [CrossRef]
- Prévosto, B.; Kuiters, L.; Bernhardt-Römermann, M.; Dölle, M.; Schmidt, W.; Hoffmann, M.; Van Uytvanck, J.; Bohner, A.; Kreiner, D.; Stadler, J.; et al. Impacts of Land Abandonment on Vegetation: Successional Pathways in European Habitats. Folia Geobot. 2011, 46, 303–325. [Google Scholar] [CrossRef] [Green Version]
- Virágh, K.; Bartha, S. The effect of current dynamical state of a loess steppe community on its responses to disturbances. Tiscia 1996, 3, 3–13. [Google Scholar]
- Bakker, J.P. The impact of grazing on plant communities. In Grazing and Conservation Management; Wallis De Vries, M.F., Bakker, J., Van Wieren, P.S.E., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1998; pp. 137–184. [Google Scholar]
- Olff, H.; Ritchie, M.E. Effects of herbivores on grassland plant diversity. Trends Ecol. Evol. 1998, 13, 261–265. [Google Scholar] [CrossRef] [Green Version]
- Proulx, M.; Mazumder, A. Reversal of grazing impact on plant species richness in nutrient-poor vs. nutrient-rich ecosystems. Ecology 1998, 79, 2581–2592. [Google Scholar] [CrossRef]
- Alard, D.; Bance, J.F.; Frileux, P.N. Grassland Vegetation as an Indicator of the Main Agro-Ecological Factors in a Rural Landscape: Consequences for Biodiversity and Wildlife Conservation in Central Normandy (France). J. Environm. Manag. 1994, 42, 91–109. [Google Scholar] [CrossRef]
- Bartha, S. Kompozíció, differenciálódás és dinamika az erdőssztyep biom gyepjeiben. In Lejtősztyepek, löszgyepek és erdőssztyeprétek Magyarországon; Illyés, E., Bölöni, J., Eds.; MTA ÖBKI: Budapest, Hungary, 2007; pp. 72–103. (In Hungarian) [Google Scholar]
- Tatoni, T.; Roche, P. Comparison of old-field and forest revegetation dynamics in Provence. J. Veg. Sci. 1994, 5, 295–302. [Google Scholar] [CrossRef]
- Debussche, M.; Escarré, J.; Lepart, J.; Houssard, C.; Lavorel, S. Changes in Mediterranean plant succession: Old-fields revisited. J. Veg. Sci. 1996, 7, 519–526. [Google Scholar] [CrossRef]
- Bazzaz, F.A. Plants in Changing Environments: Linking Physiological, Population, and Community Ecology; Cambridge University Press: Cambridge, UK, 1996; p. 320. [Google Scholar]
- Molnár, Z.; Botta-Dukát, Z. Improved space-for-time substitution for hypothesis generation: Secondary grasslands with documented site history in SE-Hungary. Phytocoenologia 1998, 28, 1–29. [Google Scholar] [CrossRef]
- Monk, C.D. Relationship of life forms and diversity in old fields succession. Bull. Torrey Bot. Club 1983, 112, 383–392. [Google Scholar] [CrossRef]
- Belsky, A.J. Effects of Grazing, Competition, Disturbance and Fire on Species Composition and Diversity in Grassland Communities. J. Veg. Sci. 1992, 3, 187–200. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 Anemone sylvestris * | 25 Galium mollugo | 49 Pulmonaria mollis * |
| 2 Aster amellus | 26 Geranium sanguineum * | 50 Pulsatilla grandis |
| 3 Aster linosyris | 27 Hieracium bauhinii | 51 Pyrus pyraster |
| 4 Astragalus glycyphyllos | 28 Hieracium cymosum | 52 Quercus robur |
| 5 Betonica officinalis * | 29 Hypochoeris maculata * | 53 Ranunculus polyanthemos * |
| 6 Brachypodium pinnatum | 30 Inula ensifolia | 54 Rosa canina |
| 7 Buglossoides purpureo-coer. | 31 Inula hirta * | 55 Rosa gallica * |
| 8 Carex humilis * | 32 Inula salicina | 56 Salvia pratensis |
| 9 Carex montana * | 33 Iris variegata * | 57 Sedum maximum * |
| 10 Chamaecytisus albus * | 34 Lathyrus lathifolius | 58 Serratula tictoria |
| 11 Cirsium pannonicum * | 35 Lathyrus niger | 59 Solidago virga-aurea |
| 12 Clematis recta * | 36 Lembotopis nigricans | 60 Stachys recta |
| 13 Colutea arborescens | 37 Libanotis pyrenaica | 61 Stipa pulcherrima |
| 14 Crataegus monogyna | 38 Linum flavum | 62 Tanacetum corymbosum * |
| 15 Crepis praemorsa * | 39 Lychnis viscaria | 63 Teucrium chamaedrys * |
| 16 Doronicum hungaricum * | 40 Origanum vulgare | 64 Thesium linophyllon * |
| 17 Echium maculatum | 41 Peucedanum alsaticum * | 65 Trifolium alpestre |
| 18 Elymus hispidus | 42 Peucedanum cervaria * | 66 Trifolium montanum * |
| 19 Euphorbia cyparissias | 43 Poa angustifolia | 67 Trifolium rubens |
| 20 Euphorbia polychroma | 44 Polygala major | 68 Ulmus minor |
| 21 Euphorbia virgata | 45 Potentilla alba | 69 Valeriana officinalis |
| 22 Falcaria vulgaris | 46 Potentilla argentea | 70 Verbascum phoeniceum |
| 23 Festuca rupicola | 47 Prunella grandiflora | 71 Vincetoxicum hirundinaria * |
| 24 Filipendula vulgaris * | 48 Prunus spinosa | 72 Viola hirta |
| Coeno. Groups | Land-Use Types/Years after Abandonment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V/1–5 | V/6–10 | V/11–20 | V/21–30 | A/1–5 | A/6–10 | A/11–20 | A/21–30 | G/1–5 | G/6–10 | G/11–20 | G/21–30 | |
| 1.5.1 | 0.00 | 0.00 | 0.00 | 0.00 | 1.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 3.3 | 4.62 | 2.25 | 1.77 | 0.85 | 3.77 | 2.82 | 2.67 | 1.05 | 1.14 | 2.53 | 2.67 | 1.23 |
| 3.4 | 3.08 | 3.37 | 0.88 | 1.69 | 5.66 | 4.23 | 1.33 | 1.05 | 3.41 | 0.00 | 0.00 | 1.23 |
| 3.5 | 4.62 | 3.37 | 3.54 | 1.69 | 3.77 | 1.41 | 4.00 | 1.05 | 3.41 | 1.27 | 2.67 | 1.23 |
| 3.6.1.1 | 1.54 | 1.12 | 0.88 | 0.85 | 1.89 | 1.41 | 1.33 | 1.05 | 1.14 | 1.27 | 1.33 | 1.23 |
| 3.7 | 1.54 | 0.00 | 0.00 | 0.00 | 1.89 | 0.00 | 0.00 | 0.00 | 1.14 | 0.00 | 0.00 | 0.00 |
| 3.8.1 | 0.00 | 1.12 | 0.00 | 0.85 | 0.00 | 0.00 | 0.00 | 1.05 | 0.00 | 1.27 | 0.00 | 1.23 |
| 3.9.1 | 0.00 | 0.00 | 0.00 | 0.00 | 1.89 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.1.1 | 0.00 | 0.00 | 0.00 | 0.85 | 0.00 | 0.00 | 0.00 | 0.00 | 1.14 | 0.00 | 0.00 | 0.00 |
| 5.2 | 3.08 | 1.12 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.3. | 1.54 | 13.48 | 17.70 | 19.49 | 3.77 | 14.08 | 9.33 | 9.47 | 10.23 | 12.66 | 26.67 | 22.22 |
| 5.3.1 | 3.08 | 5.62 | 7.08 | 10.17 | 0.00 | 8.45 | 6.67 | 8.42 | 7.95 | 5.06 | 9.33 | 8.64 |
| 5.3.1.1 | 0.00 | 0.00 | 0.00 | 0.85 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 2.67 | 3.70 |
| 5.3.1.4 | 0.00 | 0.00 | 0.88 | 0.85 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.3.3.1 | 0.00 | 1.12 | 0.88 | 0.85 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.33 | 1.23 |
| 5.3.3.2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.05 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.4 | 7.69 | 4.49 | 4.42 | 5.93 | 1.89 | 7.04 | 6.67 | 12.63 | 11.36 | 12.66 | 8.00 | 2.47 |
| 5.5 | 1.54 | 1.12 | 1.77 | 1.69 | 3.77 | 4.23 | 2.67 | 3.16 | 1.14 | 3.80 | 2.67 | 2.47 |
| 6.2 | 1.54 | 1.12 | 0.88 | 0.85 | 1.89 | 0.00 | 1.33 | 0.00 | 1.14 | 0.00 | 0.00 | 1.23 |
| 8.1.1 | 1.54 | 1.12 | 0.88 | 0.85 | 0.00 | 1.41 | 0.00 | 0.00 | 1.14 | 0.00 | 1.33 | 0.00 |
| 8.3. | 3.08 | 1.12 | 1.77 | 1.69 | 0.00 | 1.41 | 1.33 | 1.05 | 2.27 | 0.00 | 1.33 | 0.00 |
| 8.4 | 6.15 | 3.37 | 7.08 | 5.08 | 5.66 | 5.63 | 4.00 | 5.26 | 5.68 | 5.06 | 4.00 | 11.11 |
| 8.4.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.27 | 0.00 | 0.00 |
| 8.4.2 | 6.15 | 3.37 | 7.08 | 5.08 | 5.66 | 5.63 | 4.00 | 5.26 | 5.68 | 5.06 | 4.00 | 11.11 |
| 8.4.2.2 | 0.00 | 0.00 | 0.00 | 0.85 | 0.00 | 1.41 | 1.33 | 1.05 | 1.14 | 0.00 | 1.33 | 0.00 |
| 8.4.2.3 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.14 | 0.00 | 0.00 | 0.00 |
| 8.4.2.4 | 0.00 | 0.00 | 1.77 | 1.69 | 0.00 | 1.41 | 0.00 | 2.11 | 2.27 | 2.53 | 0.00 | 1.23 |
| 8.4.3 | 1.54 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 8.4.3.2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.33 | 0.00 |
| 8.6.1 | 0.00 | 1.12 | 0.88 | 0.85 | 0.00 | 0.00 | 0.00 | 1.05 | 1.14 | 0.00 | 1.33 | 4.94 |
| Indiff. | 44.62 | 43.82 | 36.28 | 35.59 | 50.94 | 39.44 | 52.00 | 43.16 | 34.09 | 45.57 | 26.67 | 23.46 |
| n.d. | 3.08 | 6.74 | 3.54 | 0.85 | 5.66 | 0.00 | 1.33 | 1.05 | 2.27 | 0.00 | 1.33 | 0.00 |
| Coeno. Groups | Land-Use Types/Years after Abandonment | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| V/1–5 | V/6–10 | V/11–20 | V/21–30 | A/1–5 | A/6–10 | A/11–20 | A/21–30 | G/1–5 | G/6–10 | G/11–20 | G/21–30 | |
| 1.5.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.50 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 3.3 | 0.36 | 0.55 | 0.02 | 0.03 | 0.19 | 2.81 | 0.11 | 0.03 | 0.15 | 0.09 | 0.10 | 0.01 |
| 3.4 | 0.14 | 0.33 | 0.04 | 0.15 | 1.60 | 2.77 | 1.64 | 0.16 | 0.67 | 0.00 | 0.00 | 0.01 |
| 3.5 | 0.27 | 1.87 | 0.67 | 0.93 | 0.09 | 0.07 | 0.35 | 0.02 | 1.29 | 0.32 | 0.01 | 0.25 |
| 3.6.1.1 | 0.11 | 0.19 | 0.05 | 0.13 | 0.87 | 0.62 | 0.14 | 0.01 | 0.16 | 0.02 | 0.01 | 0.95 |
| 3.7 | 0.01 | 0.00 | 0.00 | 0.00 | 0.10 | 0.00 | 0.00 | 0.00 | 0.05 | 0.00 | 0.00 | 0.00 |
| 3.8.1 | 0.00 | 0.02 | 0.00 | 0.45 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 8.41 | 0.00 | 0.76 |
| 3.9.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.32 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.1.1 | 0.00 | 0.00 | 0.00 | 0.18 | 0.00 | 0.00 | 0.00 | 0.00 | 0.11 | 0.00 | 0.00 | 0.00 |
| 5.2 | 0.09 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.3. | 0.00 | 8.45 | 23.61 | 17.71 | 0.41 | 20.57 | 16.44 | 17.69 | 18.61 | 8.49 | 61.49 | 58.79 |
| 5.3.1 | 0.03 | 4.29 | 5.03 | 20.51 | 0.00 | 11.14 | 5.88 | 23.78 | 23.19 | 17.02 | 22.77 | 6.80 |
| 5.3.1.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 1.51 | 0.20 |
| 5.3.1.4 | 0.00 | 0.00 | 1.69 | 0.18 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.3.3.1 | 0.00 | 0.08 | 0.77 | 0.84 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 3.69 | 2.21 |
| 5.3.3.2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 | 0.00 | 0.00 |
| 5.4 | 0.70 | 0.65 | 0.69 | 2.20 | 0.08 | 1.00 | 3.89 | 3.75 | 17.05 | 10.75 | 2.28 | 0.46 |
| 5.5 | 0.09 | 0.09 | 0.34 | 0.32 | 0.10 | 2.35 | 9.88 | 4.67 | 0.65 | 0.49 | 0.92 | 0.65 |
| 6.2 | 0.00 | 0.04 | 0.02 | 0.11 | 0.08 | 0.00 | 0.07 | 0.00 | 0.10 | 0.00 | 0.00 | 0.19 |
| 8.1.1 | 10.94 | 4.16 | 4.52 | 1.05 | 0.00 | 0.01 | 0.00 | 0.00 | 1.74 | 0.00 | 0.01 | 0.00 |
| 8.3. | 0.98 | 0.04 | 0.40 | 0.09 | 0.00 | 0.12 | 0.08 | 0.10 | 0.09 | 0.00 | 0.07 | 0.00 |
| 8.4 | 0.50 | 0.17 | 0.37 | 0.37 | 0.00 | 0.00 | 0.00 | 0.13 | 0.27 | 0.23 | 0.33 | 2.67 |
| 8.4.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.00 |
| 8.4.2 | 1.14 | 3.42 | 0.40 | 5.76 | 1.23 | 0.54 | 4.48 | 1.46 | 0.80 | 6.04 | 1.82 | 13.07 |
| 8.4.2.2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.12 | 0.09 | 0.01 | 0.11 | 0.00 | 0.01 | 0.00 |
| 8.4.2.3 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | 0.00 | 0.00 |
| 8.4.2.4 | 0.00 | 0.00 | 0.52 | 3.05 | 0.00 | 0.19 | 0.00 | 0.43 | 2.98 | 1.86 | 0.00 | 1.70 |
| 8.4.3 | 0.04 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| 8.4.3.2 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.07 | 0.00 |
| 8.6.1 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | 0.01 | 0.00 | 0.38 |
| Indiff. | 84.66 | 75.33 | 60.33 | 46.28 | 90.76 | 57.55 | 57.31 | 47.62 | 32.32 | 46.54 | 4.48 | 10.42 |
| n.d. | 0.01 | 0.10 | 0.54 | 0.09 | 3.27 | 0.00 | 0.01 | 0.02 | 0.04 | 0.00 | 0.01 | 0.00 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szirmai, O.; Saláta, D.; Penksza, K.; Schellenberger, J.; Czóbel, S. Examination of Forest Steppe Species in the Case of Areas Where Traditional Cultivation Was Abandoned. Diversity 2022, 14, 561. https://doi.org/10.3390/d14070561
Szirmai O, Saláta D, Penksza K, Schellenberger J, Czóbel S. Examination of Forest Steppe Species in the Case of Areas Where Traditional Cultivation Was Abandoned. Diversity. 2022; 14(7):561. https://doi.org/10.3390/d14070561
Chicago/Turabian StyleSzirmai, Orsolya, Dénes Saláta, Károly Penksza, Judit Schellenberger, and Szilárd Czóbel. 2022. "Examination of Forest Steppe Species in the Case of Areas Where Traditional Cultivation Was Abandoned" Diversity 14, no. 7: 561. https://doi.org/10.3390/d14070561
APA StyleSzirmai, O., Saláta, D., Penksza, K., Schellenberger, J., & Czóbel, S. (2022). Examination of Forest Steppe Species in the Case of Areas Where Traditional Cultivation Was Abandoned. Diversity, 14(7), 561. https://doi.org/10.3390/d14070561