1. Introduction
The lesser kestrel (
Falco naumanni) is a colonial small falcon that breeds in the holes and fissures of urban buildings or farmhouses in the countryside [
1]. This species prefers steppe-like grasslands and cultivated landscapes with short vegetation and extensive crops [
2]. It is a spring and summer visitor in western Europe, migrating to Africa in autumn [
3]. It is present in Annex I of the EU Wild Birds Directive (2009/147/EEC) and its important breeding habitats have been designated as SPAs (Special Protection Areas) of the Natura 2000 network in several nations, including Greece, Italy, Portugal, and Spain [
4]. This species is also considered an important biological indicator for monitoring environmental changes because it is associated with high value habitats of the western Palaearctic [
5], where it dwells in open ecosystems that are altered by agriculture.
Although it was once considered one of the most abundant raptors of the Palaearctic region, lesser kestrel populations have undergone substantial decline across Europe, its Asian range, and its African wintering grounds [
3]. This population decline was primarily due to a reduction in the availability of nest sites, increases in the use of pesticides, and changes in land use [
2,
6]. In addition, many Palaearctic lesser kestrels are trans-Saharan migrants [
7], and there is increasing evidence of the effects of climate change on their habitat associations, timing of arrival, and travel risks. Thus, the effects of climate change may add to those associated with local scale factors [
8].
In Italy, the lesser kestrel has been widely studied in the Alta Murgia (Apulia region, Southern Italy) (
Figure 1), an area that hosts the highest number of individuals in Italy in several urban colonies, and probably presents the highest densities of lesser kestrels in urban areas worldwide [
9,
10]. Recent studies have revealed the space use and flight attributes of this lesser kestrel population [
11], its unexpected attitude towards frequent and long-distance nocturnal flights [
12,
13], and its elevated within-colony home range overlap and between-colony home range segregation [
14,
15,
16].
In this work, we first investigated the biotic community of the lesser kestrel in the Alta Murgia. We aimed to understand how changes (caused by external agents or by endogenous modifications) to the species interacting with the lesser kestrel can induce changes (both positive and negative) to its population stock. Hence, we used a qualitative modelling approach, which provided a method that is particularly useful whenever species and their interactions are known but not quantified [
17,
18]. Through simulations, we sought to detect (a) which interventions with the interacting species could help to preserve the lesser kestrel in the study area, and (b) how natural changes in the population parameters of the interacting species could be used to pro-actively predict the consequent impacts on the lesser kestrel population stock.
2. Materials and Methods
Firstly, based on our 10 years of field experience in the study area [
9,
10], we detected all of the species and/or taxonomic groups (players henceforth) that directly interact with the lesser kestrel. We detected six predators (the beech marten, the brown rat, the domestic cat, the magpie, the lanner falcon, and the peregrine falcon), three competitors (the European roller, the jackdaw, and the starling) and two kinds of prey (primary and secondary). Invertebrates (in particular, grasshoppers) are the primary prey of the lesser kestrel. In the absence or shortage of invertebrates, lesser kestrels feed on swifts, chiroptera, small reptiles, and small mammals (secondary prey).
Secondly, we split these players into two groups based on their frequent or infrequent interactions with lesser kestrels (
Figure 2). The brown rat, the domestic cat, the magpie, the peregrine falcon, the jackdaw, and the invertebrates belonged to the former group (frequent players), whereas the beech marten, the lanner falcon, the European roller, the starling, and the secondary prey belonged to the latter group (infrequent players).
Thirdly, we built the current ecological network of the lesser kestrel in the study area, which represents all of the ecological interactions that are currently active among the frequent players and the species of interest.
Next, we built two what-if scenarios where we simulated the introduction of the two infrequent players (the beech marten and the secondary prey) into the current ecological network. We used these scenarios to determine whether a possible increase in the interactions between these infrequent players and the lesser kestrel could change the dynamics of the current ecological network. The latter scenario (secondary prey) simulated the future absence or shortage of invertebrates in the study area due to unfit agricultural practices (i.e., more intensive farming and an increased use of insecticides), which would force the lesser kestrels to shift their diet toward swifts, chiroptera, small reptiles, and small mammals (secondary prey). We avoided simulating alternative scenarios with the lanner falcon, the European roller, and the starling as, based on our 10 years of field experience, these species are rare in the study area, in fact the lanner falcon population is in decline. Therefore, the probability that these species become frequent players in the future is very low.
Next, we simulated the dynamics of the current scenario using qualitative modeling. The central feature of our methodological approach was that all ecological communities are composed of species connected to each other both directly and indirectly through a shared network of species interactions [
19]. The presence of indirect effects implies that species need not interact directly in order to affect each other’s populations, and that direct effects may be counteracted by indirect effects [
20,
21]. We used the loop analysis [
22] as a means of encapsulating the topology of all the pairwise interactions present in the ecological network of the lesser kestrel in the Alta Murgia. The loop analysis is a qualitative modeling method that allows the evolution of the equilibrium values of system variables (i.e., species in our case study) to be predicted, following perturbations (inputs) that occur by permanently changing one or more parameters (e.g., mortality or fecundity) in the growth rate of the variables. Changes in the parameter values define new equilibrium points with new values for the levels of the variables. Inputs can be positive or negative and be caused by external agents or endogenous modifications. Because of the connections between components of the whole system, inputs propagate beyond their direct targets to all of the variables under study, and the reticulate interaction chains can magnify or dampen the effects of each direct interaction [
22]. For example, indirect effects can coalesce and result in positive species responses even in systems where each species’ direct effect on another is negative [
23]. Any system variable can be the target of perturbations, and when an input (positive or negative) acts on a variable, there will be consequences both on the target variable and on the variables that directly or indirectly interact with it. In mathematical terms, each species’ dynamics can be thought of as:
where
Xj is the variable for which the equilibrium value is calculated, the function
fj represents any imaginable relationship between the species
Xj and the other species
Xh≠j with population parameters
ch≠j. In a loop analysis, the community matrix is a table where the rows represent the variables starting the effect and the columns represent the variables that receive the effect. Each
hj element of the community matrix represents the partial derivative of
Xj, with respect to the growth rate of species
h. The matrix values can be −1, 0, or 1. The loop analysis allows us to understand whether the equilibrium value of
Xj (after simulating perturbations in the population parameters
ch of species
Xh≠j) is expected to increase (i.e.,
> 0), decrease (i.e.,
< 0), or remain the same (i.e.,
= 0). In mathematical terms,
where
Xj is the variable (species) for which the equilibrium value is calculated;
ch is the parameter (growth rate) of the
hth species that is changing;
∂ƒh/∂ch describes whether the growth rate of the
hth variable increased or decreased after changing the parameter
ch;
Pjh(k) is the product of the signs of the links comprising a causal pathway between variable
Xj and
Xh, including
k variables; and
Fn–k(comp) represents feedback in the complementary subsystem of
Pjh(k).
Fn describes the overall feedback of the system, which is defined as:
where
L(
m,
n) is the product of the signs of
m disjunct loops in
n variables for
m = 1 up to
n, and (−1)
m+1 ensures that the sign of the feedback is corrected for
n being even.
Equations (2) and (3) produce the table of predictions, i.e., a matrix whose entries denote the net effect (the sum of the direct and indirect effects) on variable
j, resulting from a perturbation on variable
h, i.e., the variations expected in the lesser kestrel population stock in response to positive/negative parameter inputs affecting any other species. Variations can be positive (population growth), negative (population decline), or null (when positive and negative effects tend to counteract each other, resulting in a net change of zero or a small change that is considered negligible). As the system variables are often connected to each other by multiple pathways, if such pathways have opposite effects, then the qualitative model can yield ambiguous predictions. Therefore, to safeguard against ambiguous predictions, we randomly assigned numerical values from a uniform distribution to the coefficients of the community matrix: −1 values were replaced with random values in the [−1, 0) interval and +1 values were replaced with random values in the (0, 1] interval. This was performed 100 ×
n2 times, where
n was the number of species. The frequency of positive or negative effects after these randomizations defined the sign and the probability of the net effect on variable
j (i.e., the lesser kestrel), following a perturbation on variable
h (e.g., the peregrine falcon). In order to meet the stability criterion, we only retained the simulations where the overall feedback (
Fn) was negative [
24].
We then repeated these simulations for the what-if scenarios.
4. Discussion
In this work, we have investigated the biotic community of the lesser kestrel in Southern Italy for the first time. We have used its ecological network to detect fit management options aimed at preserving this important bird species in a region that presents the highest density of lesser kestrels in urban areas worldwide. Our approach sought to distill the complexity of this biotic community down to manageable and measurable levels that allow reliable predictions of its behaviors [
25,
26]. The structure of the community interaction network was fundamental to our understanding of the factors that affect the lesser kestrel’s dynamics through the propagation of different types of perturbations [
27,
28].
In the study area, the biotic community of the lesser kestrel is composed of nine species and two functional groups, i.e., primary and secondary prey. The peregrine falcon and the domestic cat are the top predators. The former preys on adult lesser kestrels and the latter preys on chicks. The beech marten, the brown rat, the domestic cat, the magpie, and the lanner falcon are intermediate predators. The beech marten and the brown rat feed on eggs, chicks, and adult lesser kestrels. The magpie feeds on eggs and chicks, whereas the lanner falcon preys on adult lesser kestrels. Two out of three biotic competitors (the jackdaw and the starling) compete with the lesser kestrels for the artificial nests that are placed on top of many buildings in the study area. The third competitor (the European roller) contends for food. In fact, it forages in the same areas (in particular, the pseudo-steppes, where invertebrates are more accessible and more abundant [
29]) and sends the lesser kestrels away during their foraging attempts. The two types of prey can be considered alternative prey as the lesser kestrels largely prefer to feed on invertebrates. Thus, the secondary prey (swifts, chiroptera, small reptiles, and small mammals) enter the lesser kestrel’s ecological network only when there is a substantial decline in invertebrate populations (in particular, grasshoppers) due to unfit agricultural practices (i.e., intensive agriculture with a large use of insecticides).
Currently, these interacting players present very different population stocks and trends. Based on our 10 years of field experience [
9,
10], the peregrine falcon and the brown rat are frequent and increasing in the study area, whereas the magpie, the jackdaw, the beech marten, and the domestic cat are frequent and stable. Although the beech marten is common in the study area, our field observations indicate that its interactions with the lesser kestrels are still infrequent. The lanner falcon is rare and its population is in decline, whereas the European roller and the starling are rare but their populations are slightly increasing. This qualitative assessment of the population stocks and trends required us to distinguish between the current biotic community (composed of the frequent players) and two alternative scenarios that could possibly occur in the future.
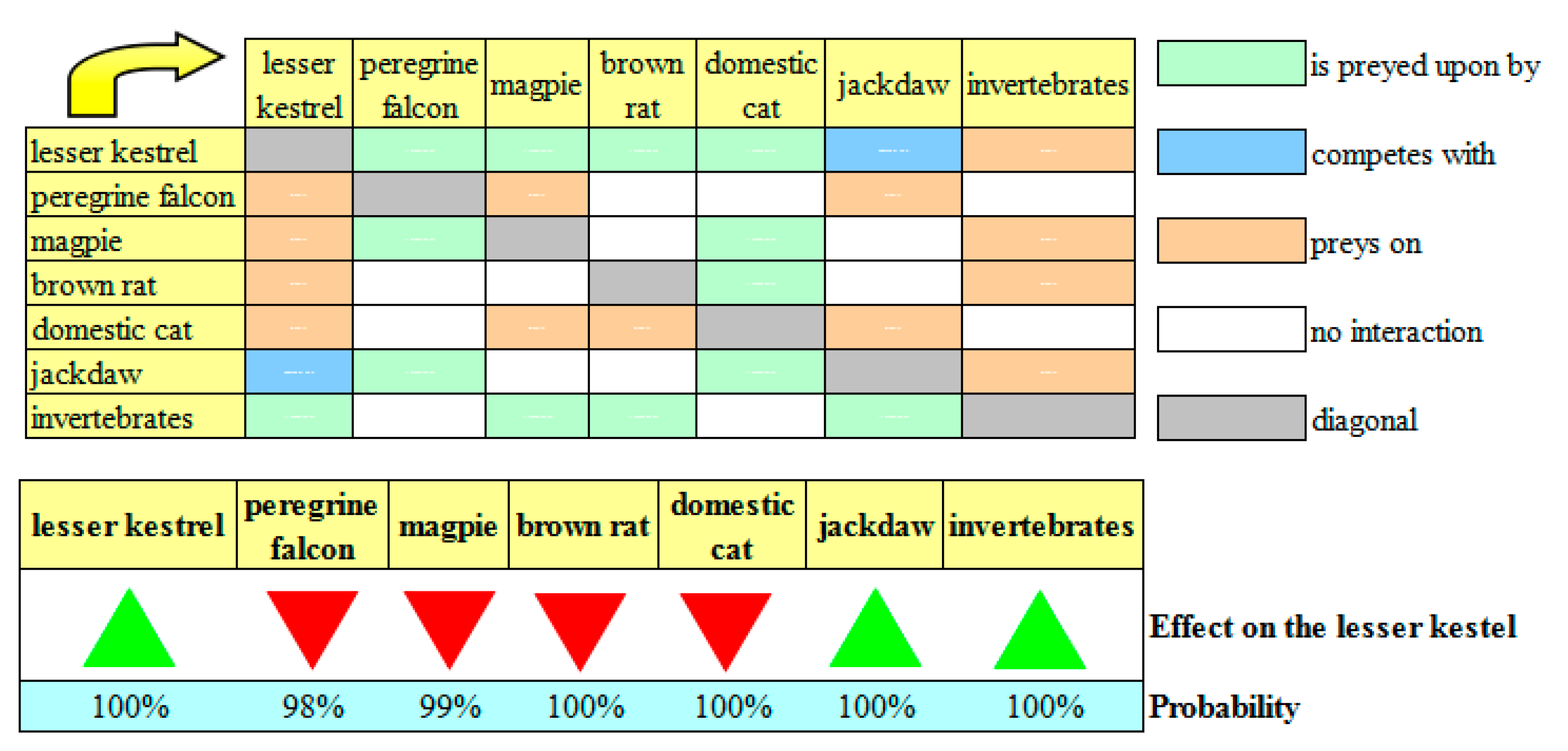
Interestingly, in all the simulated scenarios, two dynamics were shown to favor the lesser kestrel population stock with 100% probability. Any action aimed at increasing the lesser kestrel’s growth rate will result in an increase in its population stock. Although this may seem obvious, it actually was not. In fact, a positive input on the lesser kestrel also favors its predators (the peregrine falcon, the magpie, the brown rat, the domestic cat, and the beech marten), which could increase their predation rates upon the lesser kestrel. However, due to the structural properties of the biotic community of the lesser kestrel, the net effect of an increase in the lesser kestrel’s growth rate is expected to always be positive. Thus, any species-level conservation activity (e.g., the placement of new artificial nests atop urban buildings and farmhouses in order to increase the lesser kestrel’s reproductive rates) will help to preserve the lesser kestrel population stock in the study area. The second type of action that favors the lesser kestrel population stock with 100% probability in all the simulated scenarios is purposed to increase the growth rate (and thus the abundance) of the primary prey. This may also seem obvious, however an increase in the growth rate of invertebrates will also favor the two predators (the magpie and the brown rat) and a competitor (the jackdaw) of the lesser kestrel. These species also feed on invertebrates (
Figure 3). Due to the structural properties of the biotic community of the lesser kestrel, the net effect of these contrasting dynamics on the species of interest was positive in all of the simulated scenarios. Thus, a habitat-level approach based on (a) the preservation of the pseudo-steppes, (b) the optimal time for harvesting, and (c) the reduced use of pesticides will increase the abundance of invertebrates, which in turn will increase the lesser kestrel population stock. Of course, the opposite is also true. For example, if the artificial nests present in the study area are damaged or destroyed by meteorological events, or if agricultural practices become more intense, then the net effect on the lesser kestrel population stock will be negative with 100% probability.
The results of the simulations for the current scenario showed a counterintuitive result: an increase in the growth rate of the jackdaw would cause an increase in the lesser kestrel population stock (
Figure 3). As the jackdaw is a competitor of the lesser kestrel, the opposite result was expected. However, an increase in the growth rate of the jackdaw will favor the domestic cat (which preys on the lesser kestrel, the magpie, the brown rat, and the jackdaw) and the peregrine falcon (which preys on the lesser kestrel, the magpie, and the jackdaw), and the net effect of these contrasting dynamics will be positive for the lesser kestrel with 100% probability. The results of the current scenario also showed several intuitive results: any action purposed to decrease (a) the growth rates of the brown rat and the magpie, or (b) the predation rate of the domestic cat will favor the lesser kestrel population stock. For example, frequent rodent control in the study area is desirable (i.e., using ad hoc traps and avoiding the use of rat poisons that could otherwise enter the food web of the lesser kestrel). In addition, the artificial nests used by the lesser kestrels should be placed out of reach of domestic cats, so as to lower their predation rate on young lesser kestrels. In contrast, as the peregrine falcon is a protected species in Italy, no action can be taken to decrease its growth rate. Thus, this species can only be used as an indirect indicator of the lesser kestrel population stock as it is negatively correlated to it.
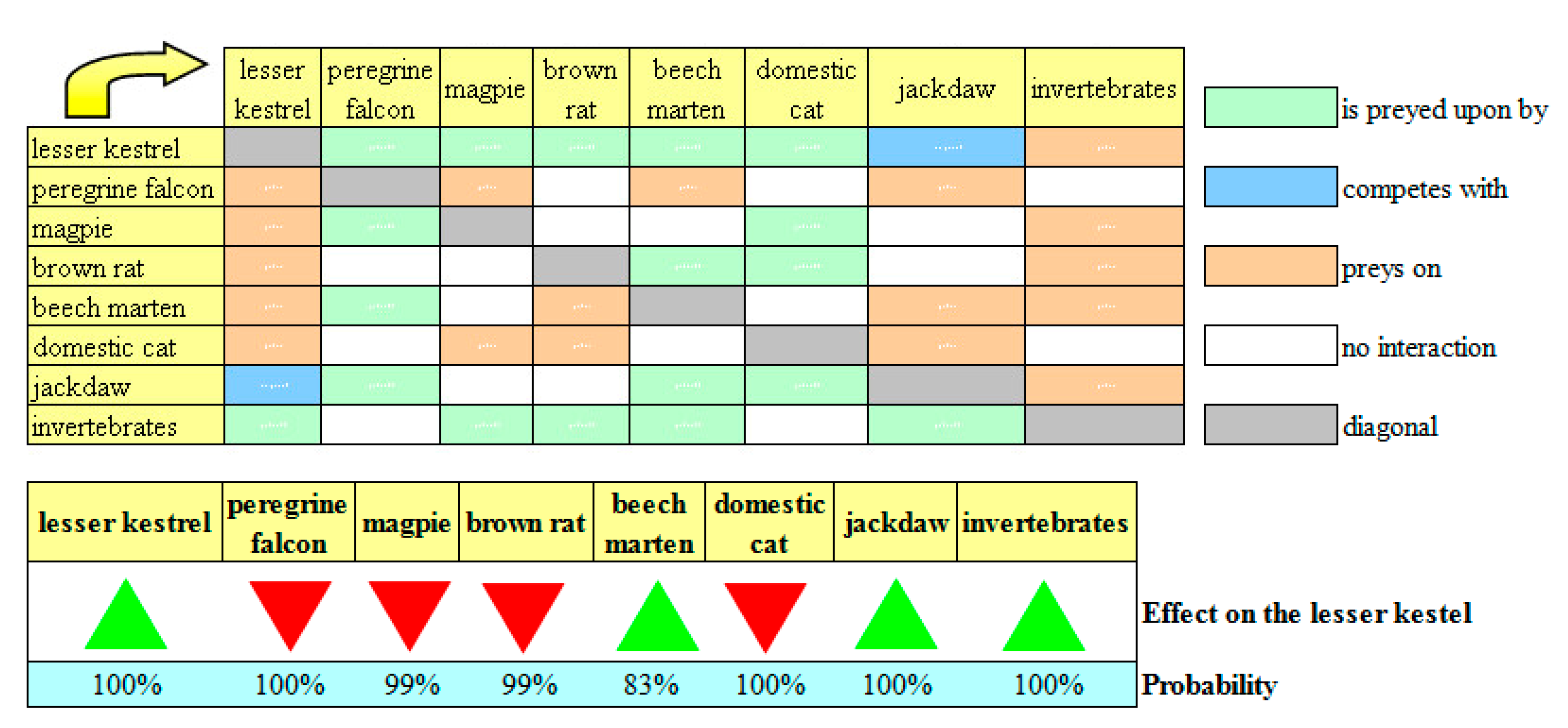
Although the beech marten feeds on the lesser kestrel, the “beech marten” scenario showed that an increase in the population stock and growth rate of this species should not be impeded (
Figure 4). In fact, the beech marten also feeds on the brown rat and the jackdaw, and the net effect of these contrasting dynamics will be positive for the lesser kestrel with 83% probability. In this scenario, the effects of the other species on the lesser kestrel would remain unchanged with respect to the current ecological network, including the positive effects of the jackdaw.
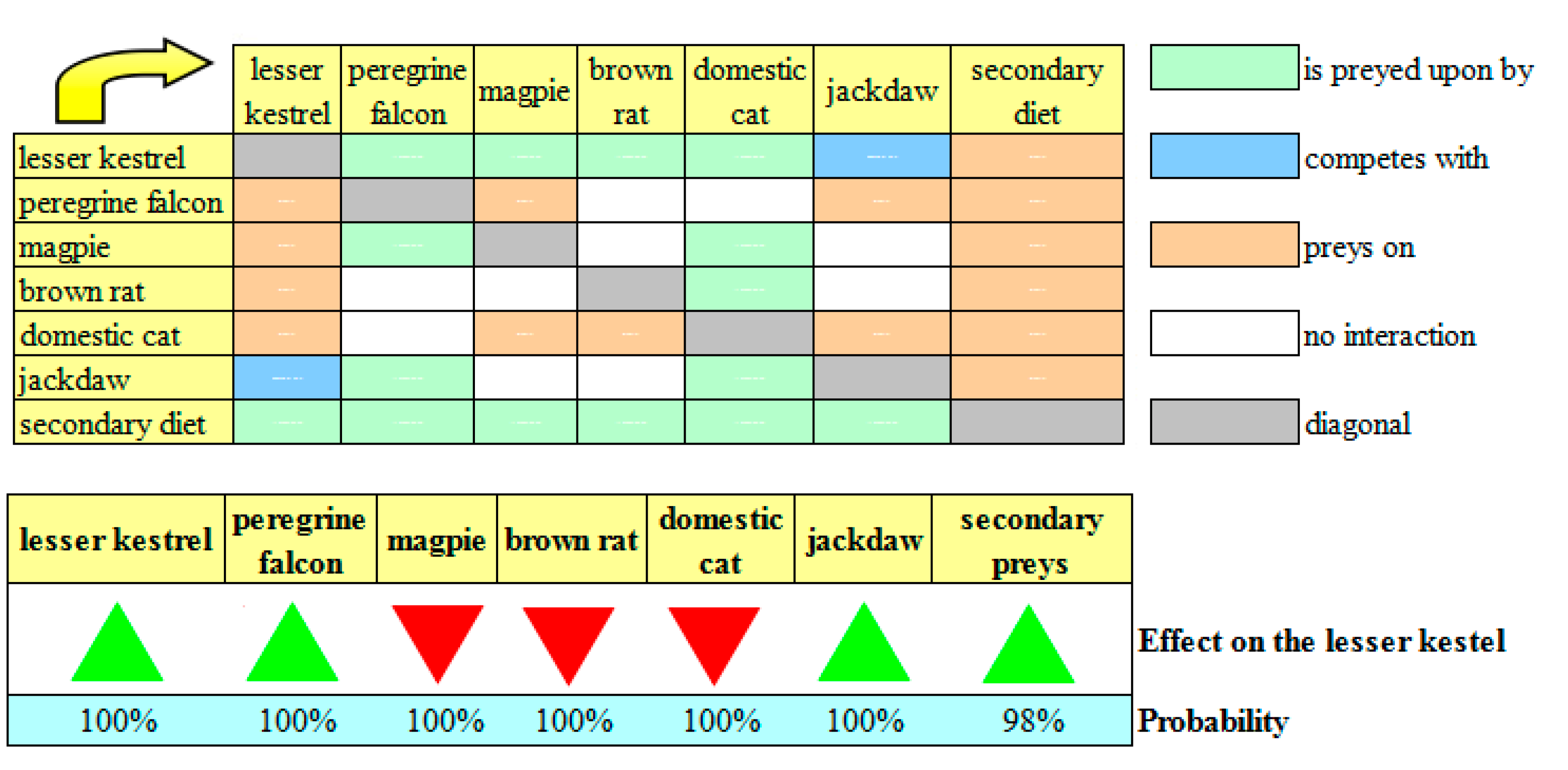
The “second prey” scenario showed another counterintuitive result (
Figure 5): If there was a substantial decline in invertebrate populations (in particular, grasshoppers), an increase in the growth rate of the peregrine falcon would favor the lesser kestrel population stock. This would occur because the magpie, the brown rat, and the jackdaw would replace the invertebrates as the lesser kestrel’s prey. However, in contrast to the current network, the secondary prey would also be preyed upon by the peregrine falcon and the domestic cat. Therefore, an increase in the growth rate of the peregrine falcon would subtract the food resources of two predators and one competitor of the lesser kestrel, and the net effect of these conflicting dynamics would be positive for the lesser kestrel with 100% probability. The peregrine falcon may also be useful to the lesser kestrel in the “starling” scenario as it is used to feed on the starling, which would become a frequent competitor of the lesser kestrel for the artificial nests that are placed on top of many buildings in the study area. This scenario was not simulated in this study because it is considered very improbable.
The methodological framework used in this study should be considered as a local-scale approach which cannot be generalized to regional or national areas because the ecological network of a certain species can change considerably from one area to another. For example, the lesser kestrel is also present in Northern Italy (in the Emilia-Romagna region) but, based on our field experience, its biotic community is very different in this region: the lanner falcon and the peregrine falcon are absent, however other species (the red-footed falcon, the common kestrel, the hooded crow, and the common buzzard) commonly interact with the lesser kestrel. In southern Portugal, the biotic community of the lesser kestrel was found to also include the barn owl and the little owl [
30].
Qualitative modelling relies on the interactions between system components without quantitative data requirements [
31]. Rather than focusing on known direct effects, qualitative modelling follows the pathways through which influences cascade in a biotic community [
32]. While developing and using our qualitative modelling approach, a fundamental step was to detect the variables considered relevant and the interactions that represented how such variables influenced each other. Gaining insight into the ecological network of the study species is among the most fruitful results of qualitative modeling [
33,
34]. Our approach required neither detailed exact equations nor expensive field measurements, but instead focused on the qualitative character of the interactions (positive, negative, or zero) and the structure of the network. Using this approach, which sacrificed precision for generality and realism [
35,
36], we circumvented many of the uncertainties (e.g., observation error and variable functional responses) that impede quantitative models [
37]. Recent applications of qualitative modelling to ecological and environmental systems are found in [
38,
39,
40,
41,
42].
Although qualitative modelling makes simplified assumptions due to the absence of quantitative data, it allowed us to create a relatively complex picture of the dynamics of the lesser kestrel biotic community. By focusing on the direction of responses, our approach provided useful guidance about different management options without the need to collect quantitative data [
43]. Firstly, a species-level approach purposed to directly increase the growth rate of this species will produce the desired effect. Secondly, a habitat-level approach aimed at preserving and extending the foraging habitats (where invertebrates are more abundant and easier to capture) will also increase the lesser kestrel population stock. As before, this outcome was not so obvious due to the complex structure of the ecological network of the lesser kestrel in the Alta Murgia. Thirdly, the presence of the jackdaw in the study area does not threaten the lesser kestrel because the net effect of its direct (negative) and indirect (positive) interactions with the lesser kestrel is positive. As the jackdaw is protected in Italy, favoring the presence of this species in the study area would produce two beneficial effects at the same time. Fourthly, the lesser kestrel would benefit from frequent rodent control. Fifthly, an increase in the abundance of beech marten in the study area should not be impeded. In fact, the net effect of its direct (negative) and indirect (positive) interactions with the lesser kestrel is positive. Sixthly, the interaction strength between the domestic cat and the lesser kestrel should be reduced, for example, by placing artificial nests where the domestic cats cannot reach. Lastly, the peregrine falcon is a threat to the lesser kestrel in the current biotic community, however, counteracting any decline in the lesser kestrel population stock would be essential in case invertebrate populations decrease in the study area due to unfit agricultural practices. In this instance, targeted actions that favor the growth rate of the peregrine falcon would help to increase the lesser kestrel population stock, despite its shift towards a secondary prey type.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}




