
Nematode Assemblages Development Twenty-One Years after the Introduction of Meadow Soil into Bare Post Mining Spoil Heap



Abstract
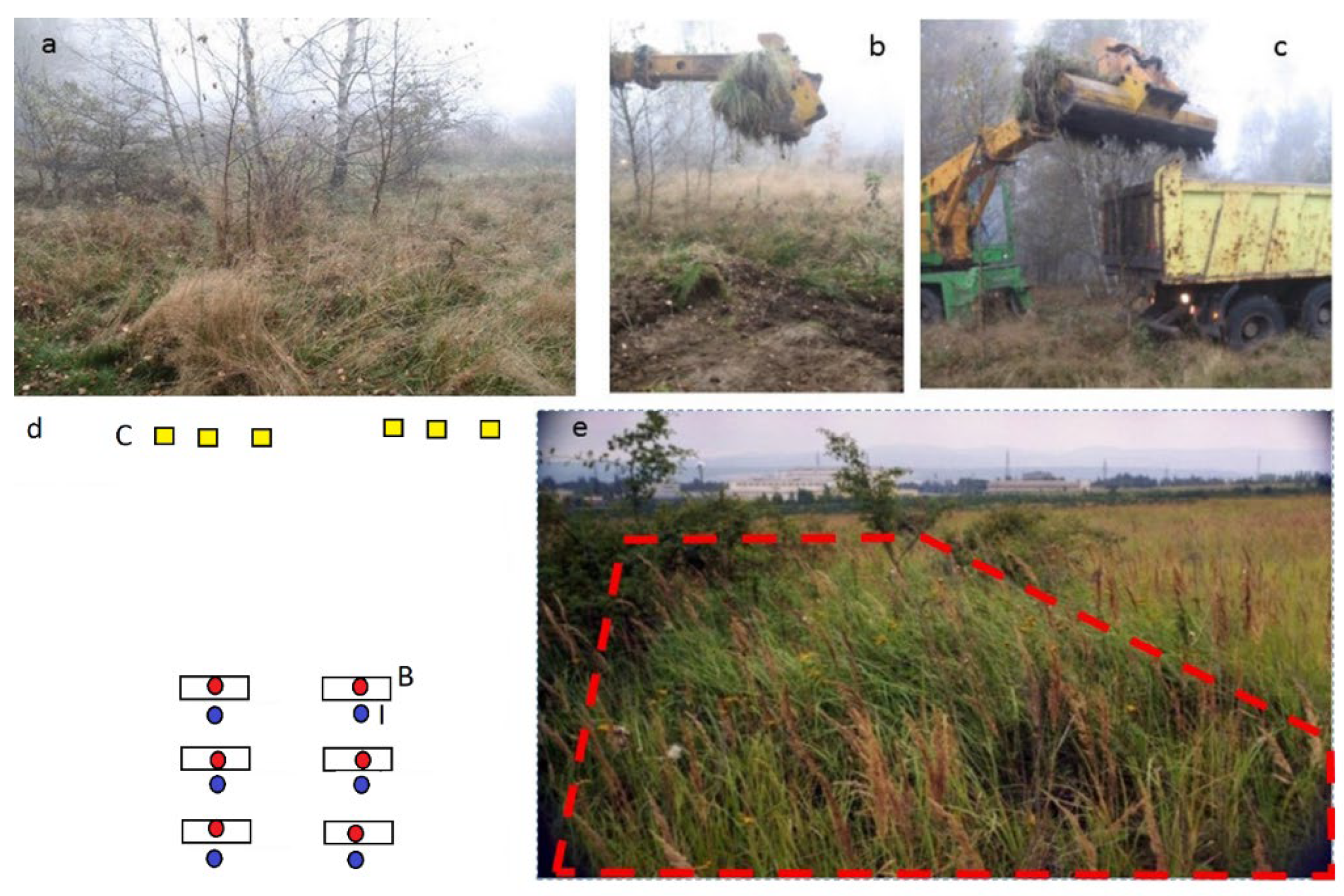
:1. Introduction
2. Materials and Methods
3. Results
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bradshaw, A. Restoration of mined lands—Using natural processes. Ecol. Eng. 1997, 8, 255–269. [Google Scholar] [CrossRef]
- Bradshaw, A.D. The Reconstruction of Ecosystems. J. Appl. Ecol. 1983, 20, 1–17. [Google Scholar] [CrossRef]
- Curry, J.P.; Good, J.A. Soil faunal degradation and restoration. In Soil Restoration; Lal, R., Steward, B.A., Eds.; Springer: New York, NY, USA, 1992; pp. 171–215. [Google Scholar]
- Bröring, U.; Wiegleb, G. Soil zoology II: Colonization, distribution, and abundance of terrestrial Heteroptera in open landscapes of former brown coal mining areas. Ecol. Eng. 2005, 24, 135–147. [Google Scholar] [CrossRef]
- Ingimarsdóttir, M.; Caruso, T.; Ripa, J.; Magnúsdóttir, B.; Migliorini, M.; Hedlund, K. Primary assembly of soil communities: Disentangling the effect of dispersal and local environment. Oecologia 2012, 170, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Brady, V.J.; Cardinale, B.J.; Gathman, J.P.; Burton, T.M. Does Facilitation of Faunal Recruitment Benefit Ecosystem Restoration? An Experimental Study of Invertebrate Assemblages in Wetland Mesocosms. Restor. Ecol. 2002, 10, 617–626. [Google Scholar] [CrossRef] [Green Version]
- Grimbacher, P.S.; Catterall, C.P. How much do site age, habitat structure and spatial isolation influence the restoration of rainforest beetle species assemblages? Biol. Conserv. 2007, 135, 107–118. [Google Scholar] [CrossRef]
- Yeates, G.W. Soil nematodes in terrestrial ecosystems. J. Nematol. 1979, 11, 213–229. [Google Scholar] [PubMed]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef] [PubMed]
- De Goede, R.G.M.; Bongers, T.; Ettema, C.H. Graphical presentation and interpretation of nematode community structure: c-p triangles. Med. Fac. Landbouww. Univ. Gent. 1993, 58, 743–750. [Google Scholar]
- Fiscus, D.A.; Neher, D.A. Distinguishing sensitivity of free-living soil nematode genera to physical and chemical disturbances. Ecol. Appl. 2002, 12, 565–575. [Google Scholar] [CrossRef]
- Yeates, G.W. Modification and qualification of the nematode Maturity Index. Pedobiologia (Jena) 1994, 38, 97–101. [Google Scholar]
- Borůvka, L.; Kozák, J.; Mühlhanselová, M.; Donátová, H.; Nikodem, A.; Němeček, K.; Drábek, O. Effect of covering with natural topsoil as a reclamation measure on brown-coal mining dumpsites. J. Geochem. Explor. 2012, 113, 118–123. [Google Scholar] [CrossRef]
- Benetková, P.; Tichý, L.; Háněl, L.; Kukla, J.; Vicentini, F.; Frouz, J. The effect of soil and plant material transplants on vegetation and soil biota during forest restoration in a limestone quarry: A case study. Ecol. Eng. 2020, 158, 106039. [Google Scholar] [CrossRef]
- Benetková, P.; van Diggelen, R.; Háněl, L.; Vicentini, F.; Moradi, R.; Weijters, M.; Bobbink, R.; Harris, J.A.; Frouz, J. Soil fauna development during heathland restoration from arable land: Role of soil modification and material transplant. Ecol. Eng. 2022, 176, 106531. [Google Scholar] [CrossRef]
- Van der Bij, A.; Weijters, M.; Bobbink, R.; Harris, J.; Pawlett, M.; Ritz, K.; Benetková, P.; Moradi, J.; Frouz, J.; van Diggelen, R. Facilitating ecosystem assembly: Plant-soil interactions as a restoration tool. Biol. Conserv. 2018, 220, 272–279. [Google Scholar] [CrossRef]
- Frouz, J.; Prach, K.; Pižl, V.; Háněl, L.; Starý, J.; Tajovský, K.; Materna, J.; Balík, V.; Kalčík, J.; Řehounková, K. Interactions between soil development, vegetation and soil fauna during spontaneous succession in post mining sites. Eur. J. Soil Biol. 2008, 44, 109–121. [Google Scholar] [CrossRef]
- Háněl, L. Nematodes in clay colliery spoil dumps and experimentally introduced strips of meadow soil. In Nematodes in Clay Colliery Spoil Dumps and Experimentally Introduced Strips of Meadow Soil; Tajovský, K., Schlaghamerský, J., Pižl, V., Eds.; Institute of Soil Biology and Biogeochemistry, Biology Centre CAS: České Budějovice, Czech Republic, 2005; pp. 19–23. [Google Scholar]
- Moradi, J.; Vicentini, F.; Šimáčková, H.; Pižl, V.; Tajovský, K.; Stary, J.; Frouz, J. An investigation into the long-term effect of soil transplant in bare spoil heaps on survival and migration of soil meso and macrofauna. Ecol. Eng. 2018, 110, 158–164. [Google Scholar] [CrossRef]
- Bongers, T. De Nematoden van Nederland, 2nd ed.; Pirola: Schoorl, The Netherlands, 1994. [Google Scholar]
- Andrássy, I. Free-Living Nematodes of Hungary; Hungarian Natural History Museum: Budapest, Hungary, 2005; Volume I. [Google Scholar]
- Andrássy, I. Free-Living Nematodes of Hungary; Hungarian Natural History Museum: Budapest, Hungary, 2007; Volume II. [Google Scholar]
- Andrássy, I. Free-Living Nematodes of Hungary; Hungarian Natural History Museum: Budapest, Hungary, 2009; Volume III. [Google Scholar]
- Sieriebriennikov, B.; Ferris, H.; de Goede, R.G. NINJA: An automated calculation system for nematode-based biological monitoring. Eur. J. Soil Biol. 2014, 61, 90–93. [Google Scholar] [CrossRef]
- Boyer, S.; Wratten, S.; Pizey, M.; Weber, P. Impact of soil stockpiling and mining rehabilitation on earthworm communities. Pedobiologia (Jena) 2011, 54, S99–S102. [Google Scholar] [CrossRef]
- Radujković, D.; van Diggelen, R.; Bobbink, R.; Weijters, M.; Harris, J.; Pawlett, M.; Vicca, S.; Verbruggen, E. Initial soil community drives heathland fungal community trajectory over multiple years through altered plant–soil interactions. New Phytol. 2019, 225, 2140–2151. [Google Scholar] [CrossRef]
- Wubs, E.R.J.; van der Putten, W.H.; Bosch, M.; Bezemer, T.M. Soil inoculation steers restoration of terrestrial ecosystems. Nat. Plants 2016, 2, 16107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Háněl, L. Development of soil nematode communities on coal-mining dumps in two different landscapes and recla-mation practices. Eur. J. Soil Biol. 2002, 38, 167–171. [Google Scholar] [CrossRef]
- Ettema, C.H.; Bongers, T. Characterization of nematode colonization and succession in disturbed soil using the Ma-turity Index. Biol. Fertil. Soils 1993, 16, 79–85. [Google Scholar] [CrossRef]
- Bongers, T.; Haar, J. On the potential of basing an ecological typology of aquatic sediments on the nematode fauna: An example from the River Rhine. Hydrobiol. Bull. 1990, 24, 37–45. [Google Scholar] [CrossRef]
- Frouz, J.; Nováková, A. Development of soil microbial properties in topsoil layer during spontaneous succession in heaps after brown coal mining in relation to humus microstructure development. Geoderma 2005, 129, 54–64. [Google Scholar] [CrossRef]
- Rantalainen, M.-L.; Fritze, H.; Haimi, J.; Pennanen, T.; Setälä, H. Colonisation of newly established habitats by soil decomposer organisms: The effect of habitat corridors in relation to colonisation distance and habitat size. Appl. Soil Ecol. 2005, 28, 67–77. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Bacterial Feeders | Fungal Feeders | Plant Feeders | Omnivores | Predators | Algal Feeders | |
|---|---|---|---|---|---|---|
| B1997 | 21.5 ± 4.5 | 30.5 ± 8.3 | 33.0 ± 5.9 b | 10.6 ± 2.7 a | 4.2 ± 1.7 | 0.1 ± 0.2 |
| I1997 | 16.8 ± 7.5 | 28.1 ± 11.3 | 21.9 ± 10.6 ab | 30.6 ± 8.9 b | 2.6 ± 2.6 | 0.0 |
| C1997 | 15.8 ± 0.4 | 24.5 ± 2.3 | 9.8 ± 0.3 a | 49.7 ± 2.8 b | 0.1 ± 0.2 | 0.0 |
| B2016 | 24.8 ± 7.7 b | 26.2 ± 10.1 b | 40.5 ± 12.7 | 6.7 ± 3.4 | 1.1 ± 0.9 a | 0.8 ± 1.6 ab |
| I2016 | 10.8 ± 5.6 a | 17.9 ± 5.3 ab | 52.1 ± 8.3 | 12.5 ± 5.9 | 4.4 ± 6.4 ab | 2.3 ± 3.6 b |
| C2016 | 19.0 ± 11.0 ab | 10.5 ± 4.6 a | 40.2 ± 23.3 | 13.8 ± 6.8 | 16.5 ± 16.0 b | 0.0 a |
| Position (B,I,C) | 0.04 | 0.04 | n.s. | <0.0005 | n.s. | n.s. |
| Year | n.s. | 0.007 | 0.0006 | <0.0005 | n.s. | n.s. |
| Position + Year | n.s. | n.s. | n.s. | <0.0005 | n.s. | n.s. |
| Treatment | Phase | MI | ƩMI | PPI | CI | BI | EI | SI | N |
|---|---|---|---|---|---|---|---|---|---|
| B | Initial | 2.64 ± 0.18 | 2.66 ± 0.11 a | 2.68 ± 0.16 | 45.45 ± 8.93 a | 18.93 ± 1.38 | 60.09 ± 4.55 | 72.93 ± 4.93 | 4 |
| I | Initial | 3.00 ± 0.32 | 2.80 ± 0.23 ab | 2.22 ± 0.23 | 72.06 ± 15.38 b | 17.02 ± 7.04 | 49.68 ± 7.71 | 79.19 ± 9.68 | 4 |
| C | Initial | 3.33 ± 0.09 | 3.24 ± 0.04 b | 2.42 ± 0.50 | 65.97 ± 0.30 ab | 11.26 ± 1.00 | 50.38 ± 0.98 | 87.28 ± 1.34 | 2 |
| B | Late | 2.38 ± 0.22 a | 2.41 ± 0.18 a | 2.50 ± 0.17 | 80.14 ± 15.23 | 39.09 ± 11.31 b | 37.08 ± 5.50 | 48.10 ± 18.59 a | 6 |
| I | Late | 2.96 ± 0.46 ab | 2.85 ± 0.28 ab | 2.74 ± 0.21 | 69.21 ± 18.4 | 18.75 ± 12.54 a | 41.14 ± 8.83 | 77.17 ± 16.91 b | 6 |
| C | Late | 3.32 ± 0.48 b | 3.05 ± 0.44 b | 2.72 ± 0.61 | 67.24 ± 20.05 | 13.63 ± 12.06 a | 22.81 ± 19.21 | 85.21 ± 13.74 b | 6 |
| Treatment | 0.001 | 0.005 | n.s. | n.s. | 0.05 | 0.05 | 0.005 | ||
| Phase | n.s. | n.s. | n.s. | n.s. | n.s. | 0.0005 | n.s. | ||
| Treatment * Phase | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | n.s. | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benetková, P.; Háněl, L.; Frouz, J. Nematode Assemblages Development Twenty-One Years after the Introduction of Meadow Soil into Bare Post Mining Spoil Heap. Diversity 2022, 14, 567. https://doi.org/10.3390/d14070567
Benetková P, Háněl L, Frouz J. Nematode Assemblages Development Twenty-One Years after the Introduction of Meadow Soil into Bare Post Mining Spoil Heap. Diversity. 2022; 14(7):567. https://doi.org/10.3390/d14070567
Chicago/Turabian StyleBenetková, Petra, Ladislav Háněl, and Jan Frouz. 2022. "Nematode Assemblages Development Twenty-One Years after the Introduction of Meadow Soil into Bare Post Mining Spoil Heap" Diversity 14, no. 7: 567. https://doi.org/10.3390/d14070567
APA StyleBenetková, P., Háněl, L., & Frouz, J. (2022). Nematode Assemblages Development Twenty-One Years after the Introduction of Meadow Soil into Bare Post Mining Spoil Heap. Diversity, 14(7), 567. https://doi.org/10.3390/d14070567