
Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study


{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
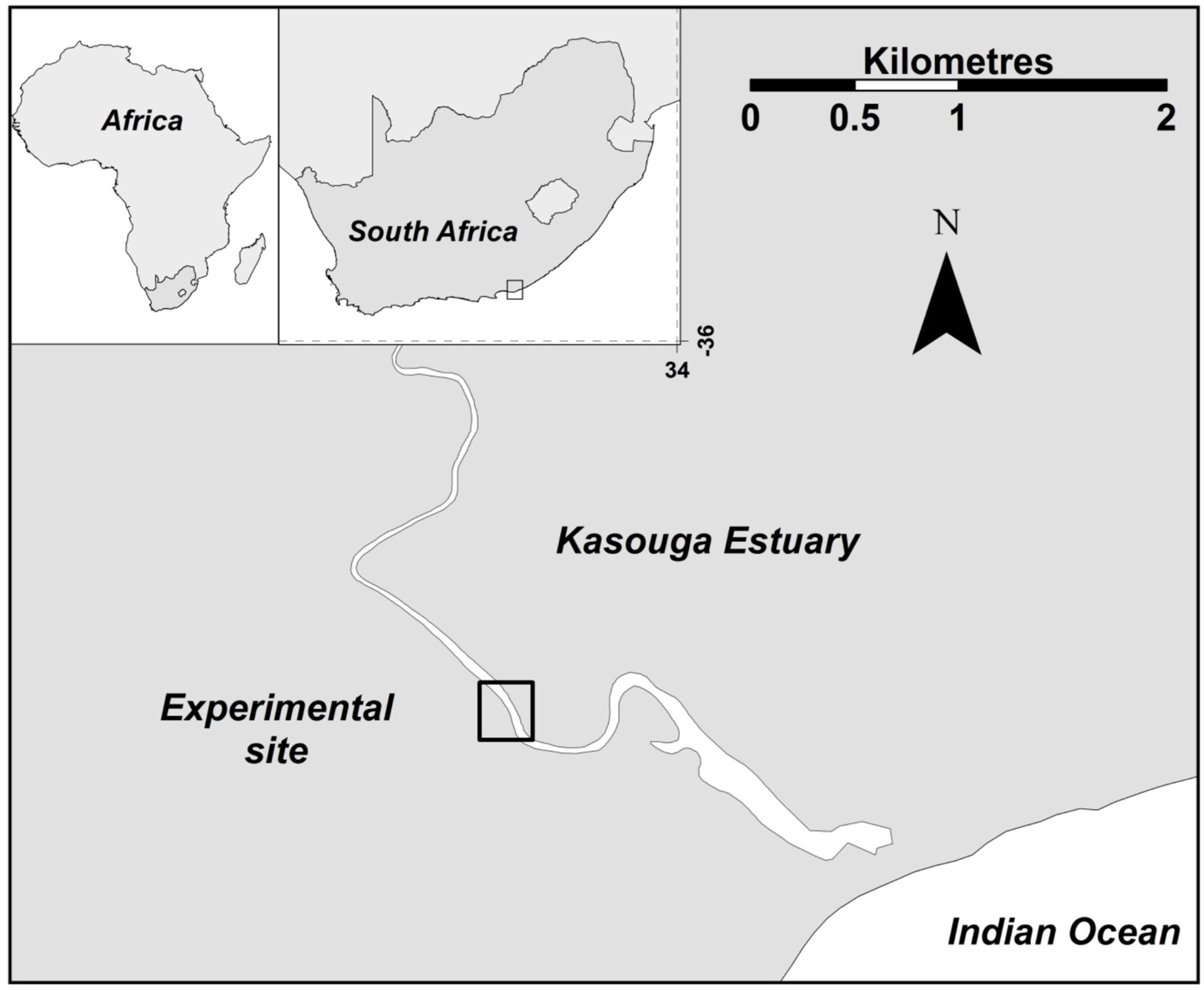
2.1. Study Site
2.2. Experimental Set-Up
2.3. Statistical Analyses
3. Results
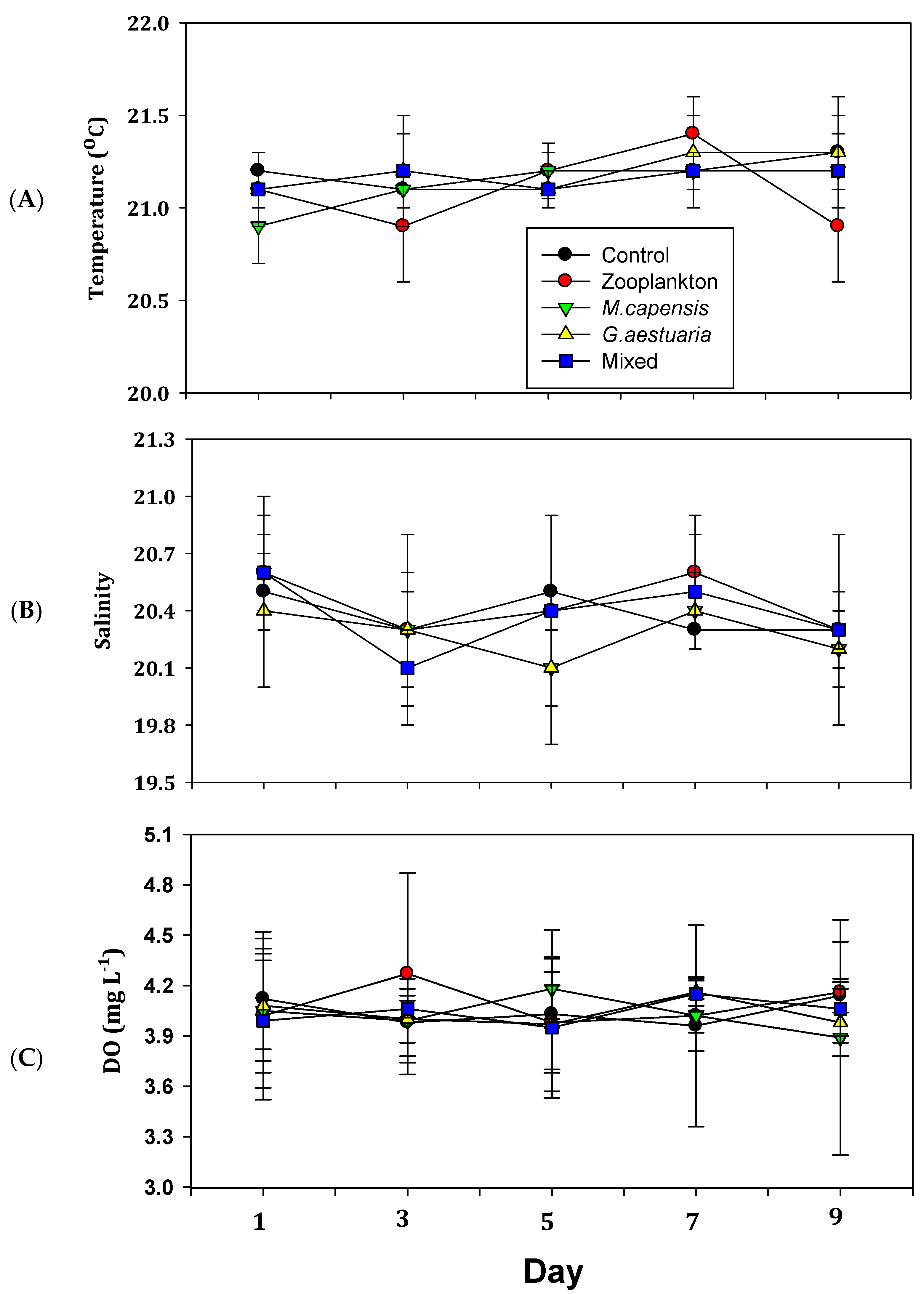
3.1. Physico-Chemical Variables
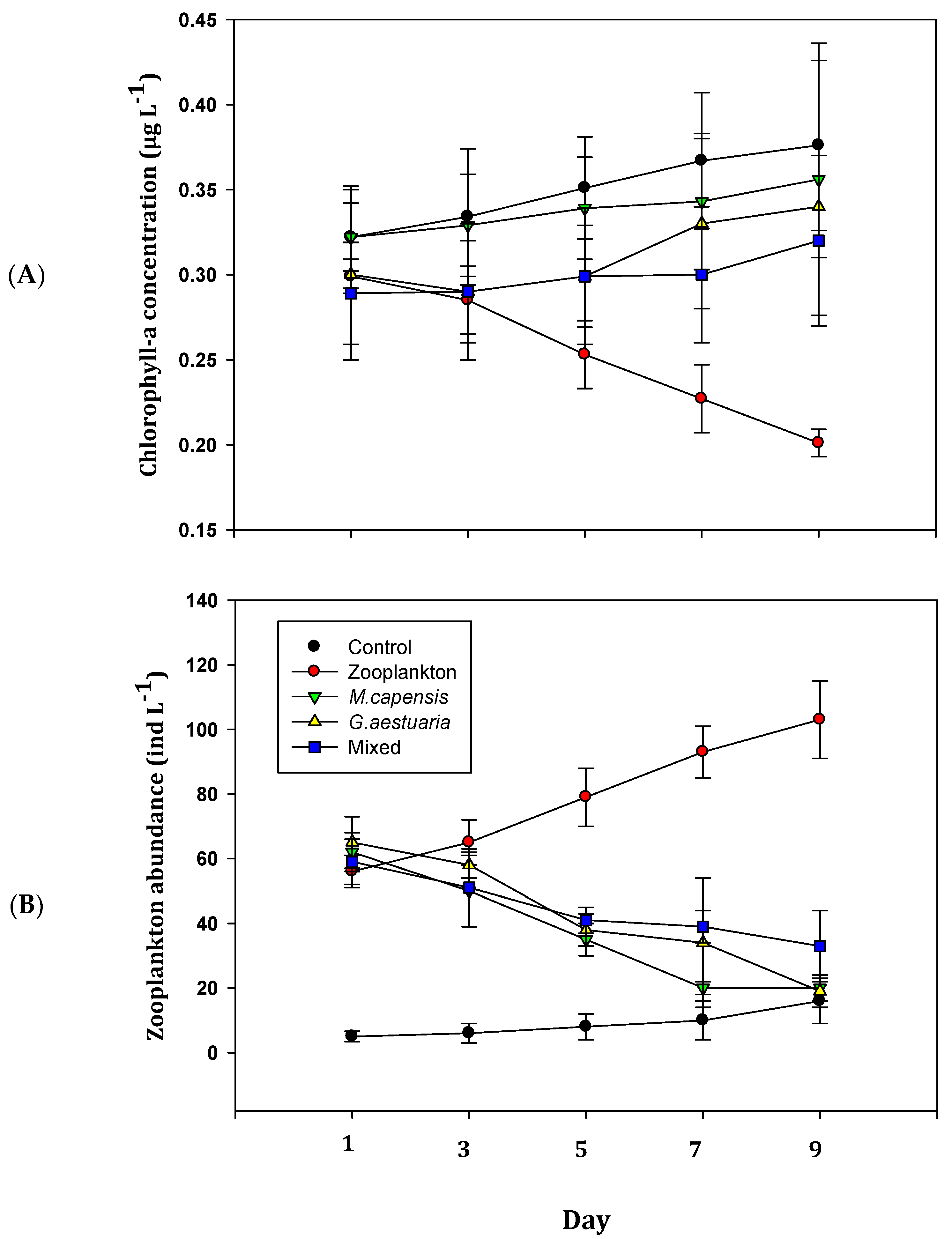
3.2. Biological Variables
4. Discussion
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bruno, J.F.; O’Connor, M.I. Cascading effects of predator diversity and omnivory in a marine food web. Ecol. Lett. 2005, 8, 1048–1056. [Google Scholar] [CrossRef]
- Wasserman, R.J.; Vink, T.J.F.; Kramer, R.; Froneman, P.W. Hyperbenthic and pelagic predators regulate alternate key planktonic predators in shallow temperate estuaries. Mar. Freshw. Res. 2014, 65, 791–801. [Google Scholar] [CrossRef]
- Wasserman, R.J.; Matcher, G.F.; Vink, T.J.F.; Froneman, P.W. Preliminary evidence for the organization of a bacterial community by zooplanktivores at the top of an estuarine planktonic food web. Microbial. Ecol. 2015, 69, 245–253. [Google Scholar] [CrossRef] [PubMed]
- Pace, M.L.; Cole, J.; Carpenter, S.R.; Kitchell, R.F. Trophic cascades revealed in diverse ecosystems. TREE 1999, 14, 483–488. [Google Scholar] [CrossRef]
- Sommer, U. Trophic cascades in marine and freshwater plankton. Int. Rev. Hydrobiol. 2008, 93, 506–516. [Google Scholar] [CrossRef]
- Finke, D.L.; Denno, R.F. Predator diversity dampens trophic cascades. Nature 2004, 429, 407–410. [Google Scholar] [CrossRef]
- Schmitz, O.J. Predator diversity and trophic interactions. Ecology 2007, 88, 2415–2426. [Google Scholar] [CrossRef]
- Polis, G.A.; Holt, R.D. Intra-guild predation: The dynamics of complex trophic interactions. TREE 1999, 7, 151–155. [Google Scholar]
- Sih, A.; Englund, G.; Wooster, D. Emergent impacts of multiple predators on prey. TREE 1998, 13, 35–355. [Google Scholar] [CrossRef]
- Ives, A.R.; Cardinale, B.J.; Snyder, W.E. A synthesis of subdisciplines: Predator-prey interactions, and biodiversity and ecosystem functioning. Ecol. Lett. 2005, 8, 102–116. [Google Scholar] [CrossRef]
- Vance-Chalcraft, H.D.; Soluk, D.A. Multiple predator effects result in risk reduction for prey across multiple prey densities. Oecologia 2005, 144, 472–480. [Google Scholar] [CrossRef]
- Griffen, B.D. Detecting emergent effects of multiple predator species. Oecologia 2006, 148, 702–709. [Google Scholar] [CrossRef] [PubMed]
- Soluk, D.A.; Collins, N.C. Synergistic Interactions between Fish and Stoneflies: Facilitation and Interference among Stream Predators. Oikos 1988, 52, 94–100. [Google Scholar] [CrossRef]
- Losey, J.E.; Denno, R.F. Positive predator- predator interactions: Enhanced predation rates and synergistic suppression of aphid populations. Ecology 1998, 79, 2143–2152. [Google Scholar]
- Whitfield, A.K. Biology and Ecology of Fishes in Southern African Estuaries; Ichthyological Monographs of the J.L.B Smith Institute of Ichthyology: Grahamstown, South Africa, 1998; Volume 2, 223p. [Google Scholar]
- Franco, M.; Elliott, P.; Franzoi, P.; Torricelli, P. Life strategies of fishes in European estuaries: The functional guild approach. Mar. Ecol. Prog. Ser. 2008, 354, 219–228. [Google Scholar] [CrossRef] [Green Version]
- Kemp, J.O.G.; Froneman, P.W. Recruitment of ichthyoplankton and macrozooplankton during overtopping events into a temporarily open/closed southern African estuary. Est. Coast. Shelf Sci. 2004, 61, 529–537. [Google Scholar] [CrossRef]
- Strydom, N.A. Patterns in Larval Fish Diversity, Abundance, and Distribution in Temperate South African Estuaries. Estuaries Coasts 2015, 38, 268–284. [Google Scholar] [CrossRef]
- James, N.C.; Leslie, T.D.; Potts, W.N.; Whitfield, A.K.; Rajakaran, A. The importance of different juvenile habitats as nursery areas for a ubiquitous estuarine-dependent marine fish species. Estuar. Coast. Shelf Sci. 2019, 226, 106270. [Google Scholar] [CrossRef]
- Whitfield, A.K. Ichthyofaunal assemblages in estuaries: A South African case study. Rev. Fish Biol. Fish. 1999, 9, 151–186. [Google Scholar] [CrossRef]
- Vorwerk, P.D.; Whitfield, A.K.; Cowley, P.D.; Paterson, A.W. A survey of selected Eastern Cape estuaries with particular reference to the ichthyofauna. Ichthyol. Bull. 2001, 72, 1–52. [Google Scholar]
- Wasserman, R.J.; Noyon, M.; Avery, T.S.; Froneman, P.W. Trophic level stability-inducing effects of predaceous early juvenile fish in an estuarine mesocosm study. PLoS ONE 2013, 8, e61019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitfield, A.K.; Cowley, P.D. The status of fish conservation in South African estuaries. J. Fish Biol. 2011, 76, 2067–2089. [Google Scholar] [CrossRef] [PubMed]
- Froneman, P.W. Food web structure in three contrasting estuaries determined using stable isotope (δ13C) analysis. Afr. J. Aquat. Sci. 2002, 27, 107–115. [Google Scholar] [CrossRef]
- Froneman, P.W. Seasonal variations in selected physico-chemical and biological variables in the temporarily open/closed Kasouga estuary (South Africa). Afr. J. Aquat. Sci. 2002, 27, 117–123. [Google Scholar] [CrossRef]
- Froneman, P.W. The importance of phytoplankton size in mediating trophic interactions in a temperate, southern African estuary. Estuar. Coast. Shelf Sci. 2006, 70, 693–700. [Google Scholar] [CrossRef]
- Holm-Hansen, O.; Riemann, B. Chlorophyll a Determination: Improvements in Methodology. Oikos 1978, 30, 438–447. [Google Scholar] [CrossRef]
- Venables, W.N.; David, M.; Smith, D.M.; The R development core team. An Introduction to R, Version 1.0; The R development core team: Vienna, Austria, 2003.
- Auffan, M.; Tella, M.; Santaella, C.; Brousset, L.; Paillès, C.; Barakat, M.; Espinasse, B.; Artells, E.; Issartel, J.; Masion, A.; et al. An adaptable mesocosm platform for performing integrated assessments of nanomaterial risk in complex environmental systems. Sci. Rep. 2014, 4, 5608. [Google Scholar] [CrossRef]
- Wilson, C.; Caughlin, T.T.; Civitello, D.J.; Flory, S.L. Combining mesocosm and field experiments to predict invasive plant performance: A hierarchical Bayesian approach. Ecology 2015, 96, 1084–1092. [Google Scholar] [CrossRef] [Green Version]
- Davis, M.; Hodgkins, G.A.; Stoner, A.W. A mesocosm system for ecological research with marine invertebrate larvae. Mar. Ecol. Prog. Ser. 1996, 130, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, R.G.; Kadin, M. The importance of benthic-pelagic coupling for marine ecosystem functioning. Glob. Change Biol. 2017, 23, 2179–2196. [Google Scholar] [CrossRef] [Green Version]
- Froneman, P.W.; Cuthbert, R.N. Habitat complexity alters predator-prey interactions in a shallow water ecosystem. Diversity 2022, 14, 431. [Google Scholar] [CrossRef]
- Allan, E.L.; Froneman, P.W. Trophic interactions amongst the plankton in a temporarily open/closed estuary, South Africa. Afr. J. Aquat. Sci. 2008, 33, 167–174. [Google Scholar] [CrossRef]
- Power, M.E. Top-down and bottom-up forces in food web: Do plants have primacy. Ecology 1992, 73, 733–746. [Google Scholar] [CrossRef]
- Lynam, C.P.; Llope, M.; Mollmann, C.; Steneth, N.C. Interaction between top-down and bottom-up control in marine food webs. PNAS 2017, 114, 1952–1957. [Google Scholar] [CrossRef] [Green Version]
- Edgar, N.B.; Green, J.D. Calanoid copepod grazing on phytoplankton: Seasonal experiments on natural communities. Hydrobiologia 1994, 273, 147–161. [Google Scholar] [CrossRef]
- Lonsdale, D.J.; Cosper, E.M.; Doall, M. Effects of zooplankton grazing on phytoplankton size-structure and biomass in the lower Hudson River estuary. Estuaries 1996, 19, 874–889. [Google Scholar] [CrossRef]
- McIntyre, P.B.; Flecker, A.S.; Vanni, M.J.; Hood, J.M.; Taylor, B.W.; Thomas, S.A. Fish distributions and nutrient cycling in streams: Can fish create biogeochemical hotspots? Ecology 2008, 89, 2335–2346. [Google Scholar] [CrossRef] [Green Version]
- Blaber, S.J.M.; Whitfield, A.K. The feeding ecology of juvenile mullet (mugilidae) in south-east African estuaries. Biol. J. Linnaean Soc. 1997, 9, 277–284. [Google Scholar] [CrossRef]
- Dagg, M.J. Copepod grazing and the fate of phytoplankton in the northern Gulf of Mexico. Cont. Shelf Res. 1995, 15, 1303–1317. [Google Scholar] [CrossRef]
- Froneman, P.W.; Vorwerk, P. Feeding ecology of juvenile Gilchristella aestuaria and Atherina breviceps (Pisces) in a temperate estuary, South Africa. Afr. J. Aquat. Sci. 2003, 28, 35–41. [Google Scholar] [CrossRef]
- Kane, J. The feeding habits of co-occurring cod and haddock larvae from Georges Bank. Mar. Ecol. Prog. Ser. 1984, 16, 9–20. [Google Scholar] [CrossRef]
- Holt, R.D.; Polis, A. A theoretical framework for intraguild predation. Am. Nat. 1997, 149, 745–776. [Google Scholar] [CrossRef]
- Nunn, A.D.; Vickers, L.H.; Mazik, K.; Bolland, J.D.; Peirson, G.; Axford, S.N.; Henshaw, A.; Cowx, I.G. Dynamic competition and resource partitioning during the early life of two widespread, abundant and ecologically similar fishes. Hydrobiologia 2020, 847, 2211–2224. [Google Scholar] [CrossRef]
- Marshall, S. The structure and functioning of the fish assemblage of the Humber estuary, UK. Ph.D. Thesis, University of Hull, Hull, UK, 1995. [Google Scholar]
- Melo-Silva, M.; da Silva, J.C.; Bialetzki, A. Community structure of fish larvae in different biotopes of a neotropical river. Community Ecol. 2022, 23, 1–12. [Google Scholar] [CrossRef]
- Otto, S.B.; Blow, E.L.; Rank, N.E.; Smiley, J.; Bose, U. Predator diversity and identity drive interaction strength and trophic cascades in a foodweb. Ecology 2008, 89, 134–144. [Google Scholar] [CrossRef] [Green Version]
- Orlando, S. Daphnia as keystone predators: Effects on phytoplankton diversity and grazing resistance. J. Plankton Res. 2005, 27, 1229–1238. [Google Scholar]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Froneman, P.W. Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study. Diversity 2022, 14, 584. https://doi.org/10.3390/d14080584
Froneman PW. Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study. Diversity. 2022; 14(8):584. https://doi.org/10.3390/d14080584
Chicago/Turabian StyleFroneman, Pierre William. 2022. "Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study" Diversity 14, no. 8: 584. https://doi.org/10.3390/d14080584
APA StyleFroneman, P. W. (2022). Predator Diversity Does Not Contribute to Increased Prey Risk: Evidence from a Mesocosm Study. Diversity, 14(8), 584. https://doi.org/10.3390/d14080584