
Anopheles maculipennis Complex in The Netherlands: First Record of Anopheles daciae (Diptera: Culicidae)

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. DNA-Based Species Identification
2.3. Habitat Characterization
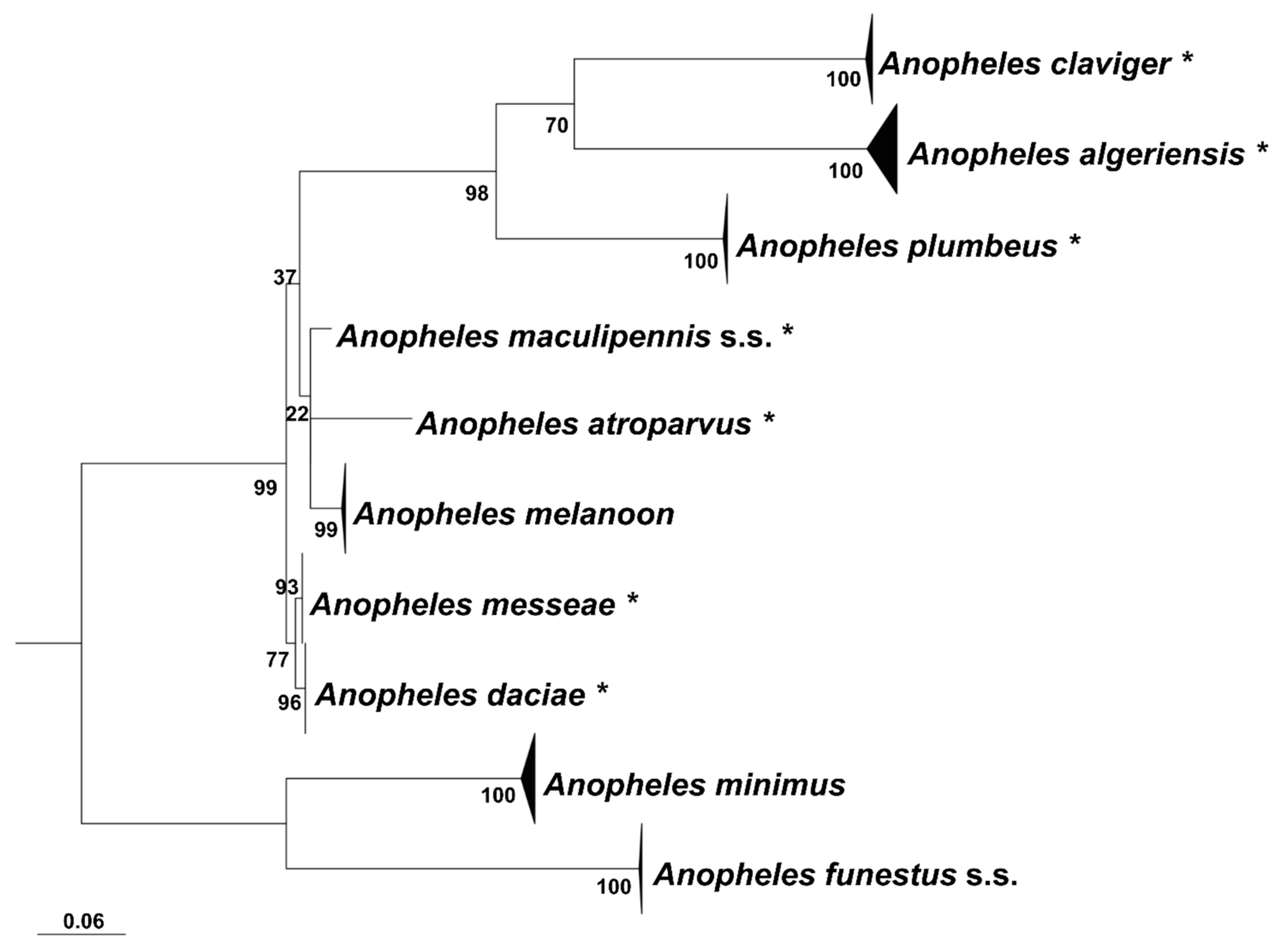
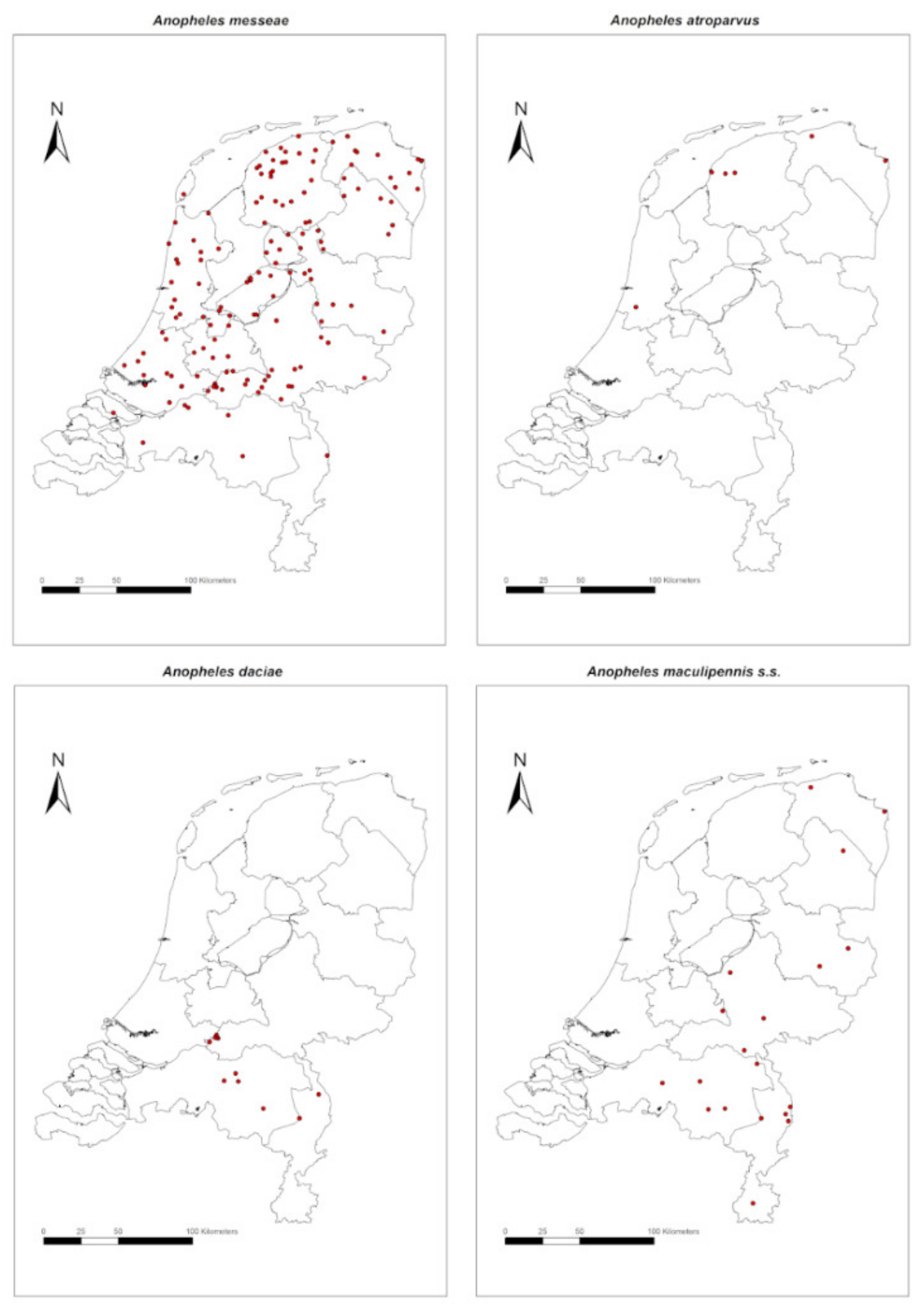
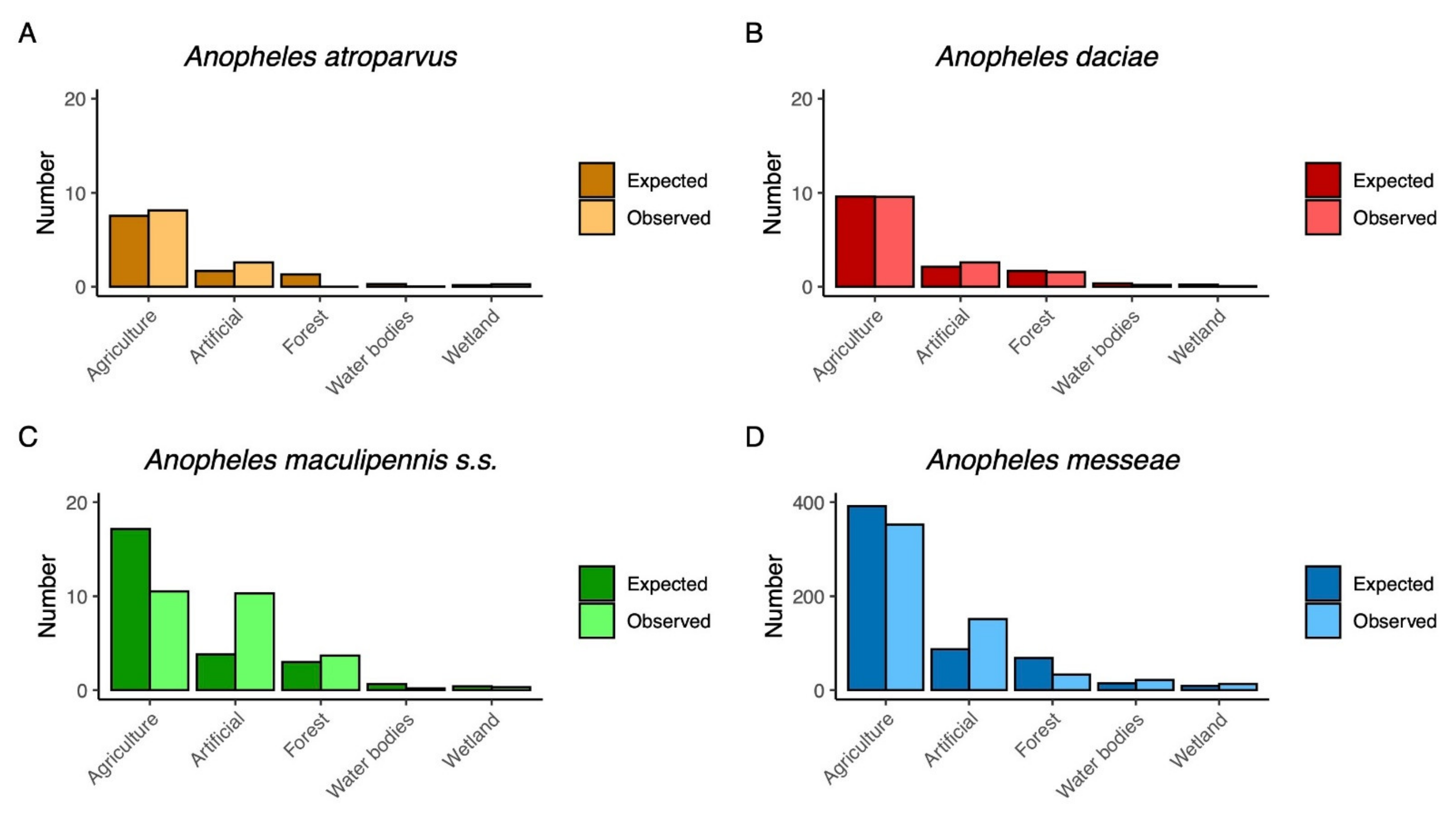
3. Results
4. Discussion and Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Verdonschot, P.F.M.; Beuk, P.L.T. Family Culicidae. In Checklist of the Diptera of the Netherlands; Beuk, P.L.T., Ed.; KNNV Uitgeverij: Utrecht, The Netherlands, 2019; Available online: https://diptera-info.nl/news.php?fam=Culicidae (accessed on 13 June 2022).
- Deblauwe, I.; Ibanez-Justicia, A.; De Wolf, K.; Smitz, N.; Schneider, A.; Stroo, A.; Jacobs, F.; Vanslembrouck, A.; Gombeer, S.; Dekoninck, W.; et al. First detections of Culiseta longiareolata (Diptera: Culicidae) in Belgium and the Netherlands. J. Med. Entomol. 2021, 58, 2524–2532. [Google Scholar] [CrossRef] [PubMed]
- Takken, W.; Kager, P.A.; Verhave, J.P. Will malaria return to North-West Europe? In Emerging Pests and Vector-Borne Diseases in Europe; Takken, W., Knols, B.G.J., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2007; pp. 23–24. [Google Scholar]
- Scholte, E.J.; Mars, M.H.; Braks, M.; Den Hartog, W.; Ibañez-Justicia, A.; Koopmans, M.; Koenraadt, C.J.; De Vries, A.; Reusken, C. No evidence for the persistence of Schmallenberg virus in overwintering mosquitoes. Med. Vet. Entomol. 2013, 28, 110–115. [Google Scholar] [CrossRef] [PubMed]
- Collins, W.E.; Jeffery, G.M. Plasmodium ovale: Parasite and disease. Clin. Microbiol. Rev. 2005, 18, 570–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takken, W.; Geene, R.; Adam, W.; Jetten, T.H.; van der Velden, J.A. Distribution and dynamics of larval populations of Anopheles messeae and A. atroparvus in the delta of the rivers Rhine and Meuse, The Netherlands. Ambio 2002, 31, 212–218. [Google Scholar] [CrossRef]
- Rodhain, J.; Van Hoof, M.T. Recherches sur l’anophélisme en Belgique. Ann. De La Société Belg. De Médecine Trop. 1942, 21, 19–43. [Google Scholar]
- Van Seventer, H.A. The disappearance of malaria in the Netherlands. Ned. Tijdschr. Voor Geneeskd. 1969, 113, 2055–2056. [Google Scholar]
- ECDC. Multiple Reports of Locally-Acquired Malaria Infections in the EU. Available online: https://www.ecdc.europa.eu/sites/portal/files/documents/RRA-Malaria-EU-revised-September-2017_0.pdf (accessed on 20 May 2022).
- Ibañez-Justicia, A.; Stroo, A.; Dik, M.; Beeuwkes, J.; Scholte, E.J. National mosquito (Diptera: Culicidae) survey in The Netherlands 2010-2013. J. Med. Entomol. 2015, 52, 185–198. [Google Scholar] [CrossRef]
- Danabalan, R.; Monaghan, M.T.; Ponsonby, D.J.; Linton, Y.M. Occurrence and host preferences of Anopheles maculipennis group mosquitoes in England and Wales. Med. Vet. Entomol. 2014, 28, 169–178. [Google Scholar] [CrossRef]
- Nicolescu, G.; Linton, Y.M.; Vladimirescu, A.; Howard, T.M.; Harbach, R.E. Mosquitoes of the Anopheles maculipennis group (Diptera: Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull. Entomol. Res. 2004, 94, 525–535. [Google Scholar] [CrossRef]
- Proft, J.; Maier, W.A.; Kampen, H. Identification of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol. Res. 1999, 85, 837–843. [Google Scholar] [CrossRef]
- Bertola, M.; Mazzucato, M.; Pombi, M.; Montarsi, F. Updated occurrence and bionomics of potential malaria vectors in Europe: A systematic review (2000-2021). Parasit Vectors 2022, 15, 88. [Google Scholar] [CrossRef] [PubMed]
- Luhken, R.; Czajka, C.; Steinke, S.; Jost, H.; Schmidt-Chanasit, J.; Pfitzner, W.; Becker, N.; Kiel, E.; Kruger, A.; Tannich, E. Distribution of individual members of the mosquito Anopheles maculipennis complex in Germany identified by newly developed real-time PCR assays. Med. Vet. Entomol. 2016, 30, 144–154. [Google Scholar] [CrossRef]
- Weitzel, T.; Gauch, C.; Becker, N. Identification of Anopheles daciae in Germany through ITS2 sequencing. Parasitol. Res. 2012, 111, 2431–2438. [Google Scholar] [CrossRef] [PubMed]
- Smitz, N.; De Wolf, K.; Gheysen, A.; Deblauwe, I.; Vanslembrouck, A.; Meganck, K.; De Witte, J.; Schneider, A.; Verle, I.; Dekoninck, W.; et al. DNA identification of species of the Anopheles maculipennis complex and first record of An. daciae in Belgium. Med. Vet. Entomol. 2021, 35, 442–450. [Google Scholar] [CrossRef]
- Ibañez-Justicia, A.; Koenraadt, C.J.M.; Stroo, A.; van Lammeren, R.; Takken, W. Risk-based and adaptive invasive mosquito surveillance at Lucky bamboo and used tire importers in the Netherlands. J. Am. Mosq. Control. Assoc. 2020, 36, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Koenraadt, C.J.M.; Mohlmann, T.W.R.; Verhulst, N.O.; Spitzen, J.; Vogels, C.B.F. Effect of overwintering on survival and vector competence of the West Nile virus vector Culex pipiens. Parasit Vectors 2019, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Becker, N.; Petric, D.; Zgomba, M.; Boase, C.; Madon, M.; Dahl, C.; Kaiser, A. Mosquitoes and Their Control; Kluwer Academic/Plenum Publishers: Heidelberg, Germany; Dordrecht, The Nertherlands; London, UK; New York, NY, USA, 2010. [Google Scholar]
- Vogels, C.B.; van de Peppel, L.J.; van Vliet, A.J.; Westenberg, M.; Ibanez-Justicia, A.; Stroo, A.; Buijs, J.A.; Visser, T.M.; Koenraadt, C.J. Winter Activity and Aboveground Hybridization Between the Two Biotypes of the West Nile Virus Vector Culex pipiens. Vector Borne Zoonotic Dis. 2015, 15, 619–626. [Google Scholar] [CrossRef]
- Robert, V.; Günay, F.; Le Goff, G.; Boussès, P.; Sulesco, T.; Khalin, A.; Medlock, J.M.; Kampen, H.; Petrić, D.; Schaffner, F. Distribution chart for Euro-Mediterranean mosquitoes (western Palaearctic region). J. Eur. Mosq. Control Assoc. 2019, 37, 1–28. [Google Scholar]
- Posada, D.; Crandall, K.A. MODELTEST: Testing the model of DNA substitution. Bioinformatics 1998, 14, 817–818. [Google Scholar] [CrossRef] [Green Version]
- Tao, N.; Richardson, R.; Bruno, W.; Kuiken, C. FindModel. 2016. Available online: http://hiv.lanl.gov/content/sequence/findmodel/findmodel.html (accessed on 18 February 2021).
- RStudioTeam. RStudio: Integrated Development for R. RStudio. 2020 Boston, MA: PBC. Available online: https://rstudio.com/ (accessed on 26 February 2021).
- Brown, S.D.; Collins, R.A.; Boyer, S.; Lefort, M.C.; Malumbres-Olarte, J.; Vink, C.J.; Cruickshank, R.H. Spider: An R package for the analysis of species identity and evolution, with particular reference to DNA barcoding. Mol. Ecol. Resour. 2012, 12, 562–565. [Google Scholar] [CrossRef]
- ESRI. ArcGIS Desktop: Release 10; Environmental Systems Research Institute: Redlands, CA, USA, 2011. [Google Scholar]
- Copernicus. CORINE Land Cover. Available online: https://land.copernicus.eu/pan-european/corine-land-cover (accessed on 17 April 2022).
- Bezzhonova, O.V.; Goryacheva, I.I. Intragenomic heterogeneity of rDNA internal transcribed spacer 2 in Anopheles messeae (Diptera: Culicidae). J. Med. Entomol. 2008, 45, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Culverwell, C.L.; Vapalahti, O.P.; Harbach, R.E. Anopheles daciae, a new country record for Finland. Med. Vet. Entomol. 2020, 34, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Lilja, T.; Eklof, D.; Jaenson, T.G.T.; Lindstrom, A.; Terenius, O. Single nucleotide polymorphism analysis of the ITS2 region of two sympatric malaria mosquito species in Sweden: Anopheles daciae and Anopheles messeae. Med. Vet. Entomol. 2020, 34, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ICZN. International Code of Zoological Nomenclature, 4th ed.; International Trust for Zoological Nomenclature: London, UK, 1999; 306p. [Google Scholar]
- Calzolari, M.; Desiato, R.; Albieri, A.; Bellavia, V.; Bertola, M.; Bonilauri, P.; Callegari, E.; Canziani, S.; Lelli, D.; Mosca, A.; et al. Mosquitoes of the Maculipennis complex in Northern Italy. Sci. Rep. 2021, 11, 6421. [Google Scholar] [CrossRef] [PubMed]
- Mukete, B.; Vermaat, J.E.; van Straalen, N.M. Variability in microhabitats prevalence in low lying peat polder ditches of the Netherlands. Appl. Ecol. Environ. Res. 2013, 11, 629–643. [Google Scholar] [CrossRef]
- Becker, N.; Petrić, D.; Zgomba, M.; Boase, C.; Madon, M.B.; Dahl, C.; Kaiser, A. Mosquitoes: Identification, Ecology and Control; Springer Nature Switzerland AG: Cham, Switzerland, 2020; 570p. [Google Scholar]
- Dakić, Z.; Zoran, K.; Novica, S.; Mijomir, P.; Miloje, C.; Stanimirovic, Z.; Nikola, I.; Poluga, J.; Pavlovic, M. Ecology of Anopheles mosquitoes in Belgrade area. Estimating vector potential for malaria retransmission. Acta Veterinaria 2008, 58, 603–614. [Google Scholar]
- Weyer, F. Die geographische Verbreitung der Rassen von Anopheles maculipennis in Deutschland. Z. Für Parasitenkd. 1938, 10, 437–463. [Google Scholar] [CrossRef]
- Ramsdale, C.D.; Wilkes, T.J. Some aspects of overwintering in southern England of the mosquitoes Anopheles atroparvus and Culiseta annulata (Diptera: Culicidae). Ecol. Entomol. 1985, 10, 449–454. [Google Scholar] [CrossRef]
- Weyer, F. Versuche zur Übertragung der affen-malaria durch stechmucken. Arch. Schiff. f. Tropenhyg. 1937, 41, 167–172. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| n | NH | NP | Average Interspecific K2P (%) ± Range (%) | |
|---|---|---|---|---|
| An. atroparvus | 11 | 1 | 0 | 8.258 ± 0.000 |
| An. daciae | 9 + 7 | 1 | 0 * | 0.881 ± 0.327 |
| An. maculipennis s.s. | 25 | 1 | 0 | 3.114 ± 0.000 |
| An. messeae | 496 + 82 | 1 | 0 | 0.687 ± 0.003 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ibáñez-Justicia, A.; Smitz, N.; Blom, R.; Vanderheyden, A.; Jacobs, F.; Meganck, K.; Gombeer, S.; Backeljau, T.; Koenraadt, C.J.M.; Griep, J.S.; et al. Anopheles maculipennis Complex in The Netherlands: First Record of Anopheles daciae (Diptera: Culicidae). Diversity 2022, 14, 636. https://doi.org/10.3390/d14080636
Ibáñez-Justicia A, Smitz N, Blom R, Vanderheyden A, Jacobs F, Meganck K, Gombeer S, Backeljau T, Koenraadt CJM, Griep JS, et al. Anopheles maculipennis Complex in The Netherlands: First Record of Anopheles daciae (Diptera: Culicidae). Diversity. 2022; 14(8):636. https://doi.org/10.3390/d14080636
Chicago/Turabian StyleIbáñez-Justicia, A., Nathalie Smitz, Rody Blom, Ann Vanderheyden, Frans Jacobs, Kenny Meganck, Sophie Gombeer, Thierry Backeljau, Constantianus J. M. Koenraadt, J. S. Griep, and et al. 2022. "Anopheles maculipennis Complex in The Netherlands: First Record of Anopheles daciae (Diptera: Culicidae)" Diversity 14, no. 8: 636. https://doi.org/10.3390/d14080636
APA StyleIbáñez-Justicia, A., Smitz, N., Blom, R., Vanderheyden, A., Jacobs, F., Meganck, K., Gombeer, S., Backeljau, T., Koenraadt, C. J. M., Griep, J. S., De Meyer, M., & Stroo, A. (2022). Anopheles maculipennis Complex in The Netherlands: First Record of Anopheles daciae (Diptera: Culicidae). Diversity, 14(8), 636. https://doi.org/10.3390/d14080636