
The Spread of the Japanese Beetle in a European Human-Dominated Landscape: High Anthropization Favors Colonization of Popillia japonica


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Study Species and Data
2.3. Predictor Variables
2.4. Data Analysis
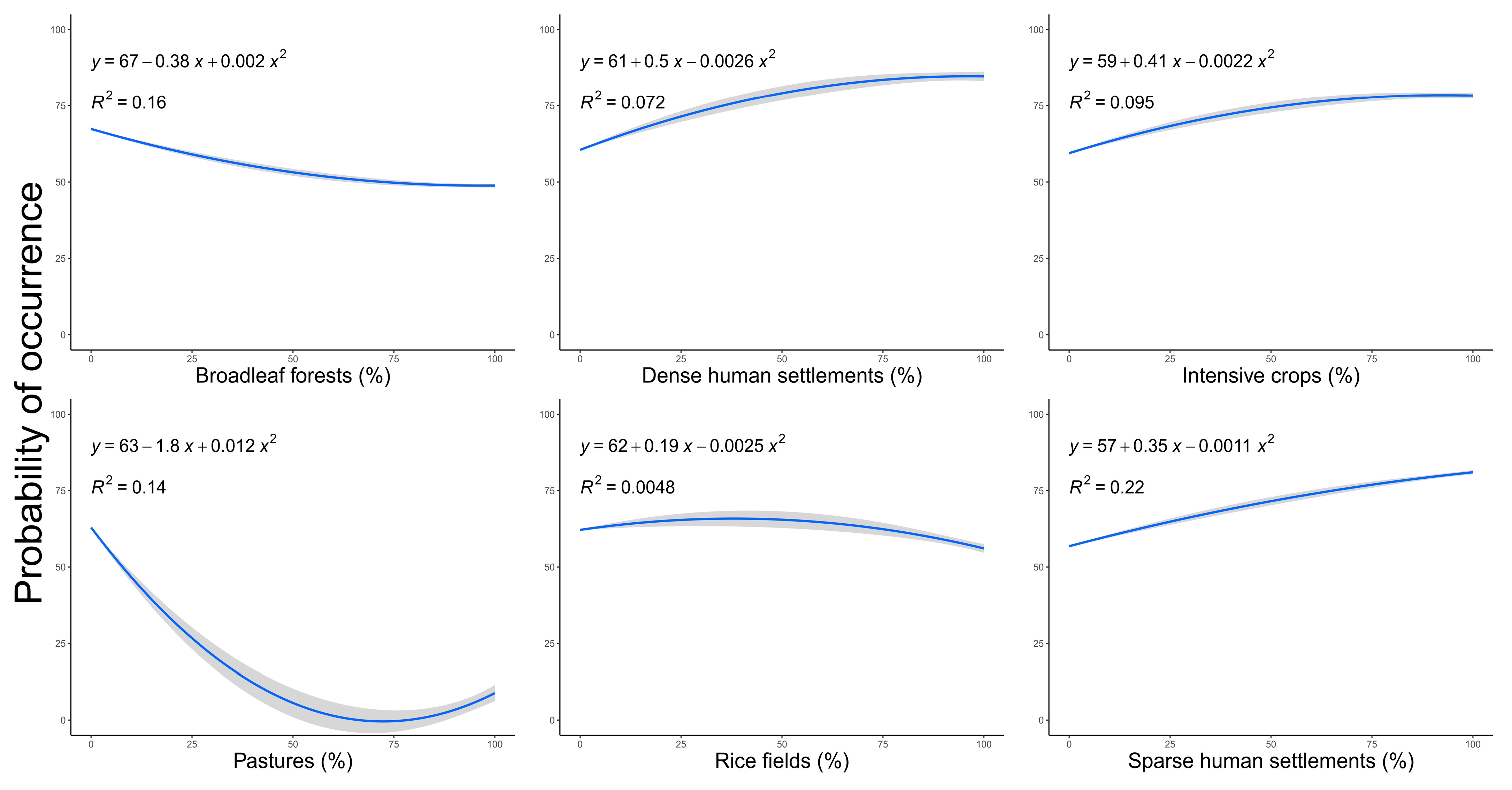
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Keller, R.P.; Geist, J.; Jeschke, J.; Kühn, I. Invasive species in Europe: Ecology, status, and policy. Environ. Sci. Eur. 2011, 23, 23. [Google Scholar] [CrossRef]
- Bellard, C.; Genovesi, P.; Jeschke, J.M. Global patterns in threats to vertebrates by biological invasions. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152454. [Google Scholar] [CrossRef]
- Bradshaw, C.J.A.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.-M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Díaz, S.; Settele, J.; Brondízio, E.S.; Ngo, H.T.; Agard, J.; Arneth, A.; Balvanera, P.; Brauman, K.A.; Butchart, S.H.M.; Chan, K.M.A.; et al. Pervasive human-driven decline of life on Earth points to the need for transformative change. Science 2019, 366, eaax3100. [Google Scholar] [CrossRef] [PubMed]
- Montagnani, C.; Gentili, R.; Brundu, G.; Caronni, S.; Citterio, S. Accidental Introduction and Spread of Top Invasive Alien Plants in the European Union through Human-Mediated Agricultural Pathways: What Should We Expect? Agronomy 2022, 12, 423. [Google Scholar] [CrossRef]
- Lodge, D.M.; Williams, S.; MacIsaac, H.J.; Hayes, K.R.; Leung, B.; Reichard, S.; Mack, R.N.; Moyle, P.B.; Smith, M.; Andow, D.A.; et al. Biological invasions: Recommendations for US policy and management. Ecol. Appl. 2006, 16, 2035–2054. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, transport and trouble: Managing invasive species pathways in an era of globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Barbet-Massin, M.; Rome, Q.; Villemant, C.; Courchamp, F. Can species distribution models really predict the expansion of invasive species? PLoS ONE 2018, 13, e0193085. [Google Scholar] [CrossRef] [PubMed]
- EPPO. EPPO Global Database. 2022. Available online: https://gd.eppo.int (accessed on 10 February 2022).
- Pavesi, M.A. Popillia japonica specie aliena invasiva segnalata in Lombardia. L’Informatore Agrar. 2014, 32, 53–55. [Google Scholar]
- Hungate, B.A.; Kearns, D.N.; Ogle, K.; Caron, M.; Marks, J.C.; Rogg, H.W. Hydrogen isotopes as a sentinel of biological invasion by the Japanese beetle, Popillia japonica (Newman). PLoS ONE 2016, 11, e0149599. [Google Scholar] [CrossRef]
- Fleming, W.E. Biology of the Japanese Beetle; Technical Bulletin No. 1449; United States Department of Agriculture: Washington, DC, USA, 1972; pp. 1–129. [Google Scholar]
- Caton, B.P.; Fang, H.; Manoukis, N.C.; Pallipparambil, G.R. Quantifying insect dispersal distances from trapping detections data to predict delimiting survey radii. J. Appl. Entomol. 2021, 146, 203–216. [Google Scholar] [CrossRef]
- EFSA Plant Health Panel; Bragard, C.; Dehnen-Schmutz, K.; Di Serio, F.; Gonthier, P.; Jacques, M.A.; Jaques Miret, J.A.; Justesen, A.F.; Magnusson, C.S.; Milonas, P.; et al. Scientific Opinion on the pest categorisation of Popillia japonica. EFSA J. 2018, 16, e05438. [Google Scholar]
- PM 1/2(28); EPPO Standards: EPPO A1 and A2 Lists of Pests Recommended for Regulation as Quarantine Pests. EPPO: Paris, France, 2019.
- Della Rocca, F.; Milanesi, P. The New Dominator of the World: Modeling the Global Distribution of the Japanese Beetle under Land Use and Climate Change Scenarios. Land 2022, 11, 567. [Google Scholar] [CrossRef]
- Kistner-Thomas, E.J. The Potential Global Distribution and Voltinism of the Japanese Beetle (Coleoptera: Scarabaeidae) Under Current and Future Climates. J. Insect Sci. 2019, 19, 16. [Google Scholar] [CrossRef]
- Ludwig, D. The Effects of Temperature on the Development of an Insect (Popillia japonica Newman). Physiol. Zool. 1928, 1, 358–389. [Google Scholar] [CrossRef]
- Regniere, J.; Rabb, R.L.; Stinner, R.E. Popillia japonica: Simulation of temperature-dependent development of the immatures, and prediction of adult emergence. Environ. Entomol. 1981, 10, 290–296. [Google Scholar] [CrossRef]
- Bourke, P.A. Climatic Aspects of the Possible Establishment of the Japanese Beetle in Europe. In Technical Note; World Meteorological Organization: Geneva, Switzerland, 1961; Chapter 41; pp. 1–9. [Google Scholar]
- Romano, B.; Zullo, F. Half a century of urbanization in southern European lowlands: A study on the Po Valley (Northern Italy). Urban Res. Pract. 2016, 9, 109–130. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Bacher, S.; Blackburn, T.M.; Dick, J.T.A.; Essl, F.; Evans, T.; Gaertner, M.; Hulme, P.E.; Kühn, I.; Mrugała, A.; et al. Defining the Impact of Non-Native Species. Conserv. Biol. 2014, 28, 1188–1194. [Google Scholar] [CrossRef]
- Hulme, P.E. Invasion pathways at a crossroad: Policy and research challenges for managing alien species introductions. J. Appl. Ecol. 2015, 52, 1418–1424. [Google Scholar] [CrossRef]
- Dyer, E.E.; Cassey, P.; Redding, D.W.; Collen, B.; Franks, V.; Gaston, K.J.; Jones, K.; Kark, S.; Orme, C.D.L.; Blackburn, T.M. The Global Distribution and Drivers of Alien Bird Species Richness. PLoS Biol. 2017, 15, e2000942. [Google Scholar] [CrossRef]
- Mori, E.; Menchetti, M.; Zozzoli, R.; Milanesi, P. The importance of taxonomy in species distribution models at a global scale: The case of an overlooked alien squirrel facing taxonomic revision. J. Zool. 2019, 307, 43–52. [Google Scholar] [CrossRef]
- Milanesi, P.; Mori, E.; Menchetti, M. Observer-oriented approach improves species distribution models from citizen science data. Ecol. Evol. 2020, 10, 12104–12114. [Google Scholar] [CrossRef] [PubMed]
- Rue, H.; Martino, S.; Chopin, N. Approximate Bayesian inference for latent Gaussian model by using integrated nested Laplace approximations (with discussion). J. R. Stat. Soc. Ser. B 2009, 71, 319–392. [Google Scholar] [CrossRef]
- Villa, L.; Maksimov, P.; Luttermann, C.; Tuschy, M.; Gazzonis, A.L.; Zanzani, S.A.; Mortarino, M.; Conraths, F.J.; Manfredi, M.T.; Schares, G. Spatial distance between sites of sampling associated with genetic variation among Neospora caninum in aborted bovine foetuses from northern Italy. Parasites Vectors 2021, 14, 47. [Google Scholar] [PubMed]
- Zampieri, M.; Scoccimarro, E.; Gualdi, S.; Navarra, A. Observed shift towards earlier spring discharge in the main Alpine rivers. Sci. Total Environ. 2015, 503–504, 222–232. [Google Scholar] [CrossRef] [PubMed]
- Zampieri, M.; Ceglar, A.; Manfron, G.; Toreti, A.; Duveiller, G.; Romani, M.; Rocca, C.; Scoccimarro, E.; Podrascanin, Z.; Djurdjevic, V. Adaptation and sustainability of water management for rice agriculture in temperate regions: The Italian case-study. Land Degrad. Dev. 2019, 30, 2033–2047. [Google Scholar] [CrossRef]
- Ajassa, R.; Beretta, E.; Biagini, E.; Biancotti, A.; Bonansea, E.; Boni, P.; Brancucci, G.; Carton, A.; Cerutti, A.V.; Ferrari, R.; et al. Mountains, hills and plains in north-western Italy. Suppl. Di Geogr. Fis. E Din. Quat. 1997, 2, 49–78. [Google Scholar]
- Monteleone, B.; Martina, M. Improving Climate Resilience of Agricultural Systems through the Development of Drought Vulnerability Curves. In Book of Abstracts Modelling for Action with a Flood of Data and a Cloud of Uncertainty; Modelling and Simulation Society of Australia and New Zealand Inc.: Canberra, Australia, 2021. [Google Scholar]
- Zullo, F.; Fazio, G.; Romano, B.; Marucci, A.; Fiorini, L. Effects of urban growth spatial pattern (UGSP) on the land surface temperature (LST): A study in the Po Valley (Italy). Sci. Total Environ. 2018, 650, 1740–1751. [Google Scholar] [CrossRef]
- Pileri, P.; Sartori, F. Monitoring biodiversity at a wide land scale to support sustainable planning and policy: The proposal of a key indicator based on vegetation cover data deriving from maps. In Monitoring and Indicators of Forest Biodiversity in Europe—From Ideas to Operationality; Marchetti, M., Ed.; European Forest Institute Proceedings: Saarijarvi, Finland, 2004; Chapter 51; pp. 455–463. [Google Scholar]
- Lassini, P.; Monzani, F.; Pileri, P. A green vision for the renewal of the Lombardy landscape. In Europe’s LIVING landscapes. Essays on Exploring Our Identity in the Countryside. Landscape Europe; Pedroli, B., van Doorn, A., de Blust, G., Eds.; KNNV Publishing: Amsterdam, The Netherland, 2007; pp. 83–100. [Google Scholar]
- Della Rocca, F.; Stefanelli, S.; Bogliani, G. Robinia pseudoacacia as a surrogate for native tree species for saproxylic beetles inhabiting the riparian mixed forests of northern Italy. Agric. For. Entomol. 2016, 18, 250–259. [Google Scholar] [CrossRef]
- iNaturalist. Available online: www.inaturalist.org (accessed on 22 April 2022).
- Barve, V.; Hart, E.; Guillou, S. Rinat: Access iNaturalist Data through APIs, R Package Version 0.1.8; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://CRAN.R-project.org/package=rinat (accessed on 22 April 2022).
- Calenge, C.; Fortmann-Roe, S. adehabitatHR: Home Range Estimation, R Package Version 0.4, 19; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://CRAN.R-project.org/package=adehabitatHR (accessed on 22 April 2022).
- Barbet-Massin, M.; Jiguet, F.; Albert, C.H.; Thuiller, W. Selecting pseudo-absences for species distribution models: How, where and how many? Methods Ecol. Evol. 2012, 3, 327–338. [Google Scholar] [CrossRef]
- Italian Digital Elevation Model. Available online: http://www.sinanet.isprambiente.it/it/sia-ispra/download-mais/dem20/view (accessed on 22 April 2022).
- CORINE Land Cover 2018 (CLC2018), 4th Level Detailed for Italy. Available online: https://groupware.sinanet.isprambiente.it/uso-copertura-e-consumo-di-suolo/library/copertura-del-suolo/corine-land-cover/corine-land-cover-2018-iv-livello (accessed on 22 April 2022).
- Zuur, A.F.; Ieno, E.N.; Elphick, C.S. A protocol for data exploration to avoid common statistical problems. Methods Ecol. Evol. 2010, 1, 3–14. [Google Scholar] [CrossRef]
- Naimi, B. usdm: Uncertainty Analysis for Species Distribution Models, R Software Package. 2014. Available online: https://CRAN.R-project.org/package=usdm (accessed on 22 April 2022).
- Beguin, J.; Martino, S.; Rue, H.; Cumming, S.G. Hierarchical analysis of spatially autocorrelated ecological data using integrated nested Laplace approximation. Methods Ecol. Evol. 2012, 3, 921–929. [Google Scholar] [CrossRef]
- Lindgren, F.; Rue, H.; Lindström, J. An explicit link between Gaussian fields and Gaussian Markov random fields: The stochastic partial differential equation approach. J. R. Stat. Soc. Ser. B Stat. Methodol. 2011, 73, 423–498. [Google Scholar] [CrossRef]
- Blangiardo, M.; Cameletti, M.; Baio, G.; Rue, H. Spatial and spatio-temporal models with R-INLA. Spat. Spatiotemporal Epidemiol. 2013, 4, 33–49. [Google Scholar] [CrossRef] [PubMed]
- Thuiller, W.; Lafourcade, B.; Engler, R.; Araújo, M.B. BIOMOD—A platform for ensemble forecasting of species distributions. Ecography 2009, 32, 369–373. [Google Scholar] [CrossRef]
- Allouche, O.; Tsoar, A.; Kadmon, R. Assessing the accuracy of species distribution models: Prevalence, kappa and the true skill statistic (TSS). J. Appl. Ecol. 2006, 43, 1223–1232. [Google Scholar] [CrossRef]
- Thuiller, W.; Georges, D.; Engler, R.; Breiner, F.; Georges, M.D.; Thuiller, C.W. Package ‘biomod2′. Species Distribution Modeling within an Ensemble Forecasting Framework. 2016. Available online: https://CRAN.R-project.org/package=biomod2 (accessed on 22 April 2022).
- Dietz, H.; Edwards, P.J. Recognition that Causal Processes Change during Plant Invasion Helps Explain Conflicts in Evidence. Ecology 2006, 87, 1359–1367. [Google Scholar] [CrossRef]
- Sádlo, J.; Chytrý, M.; Pyšek, P. Regional species pools of vascular plants in habitats of the Czech Republic. Preslia 2007, 79, 303–321. [Google Scholar]
- Chytrý, M.; Maskell, L.C.; Pino, J.; Pyšek, P.; Vilà, M.; Font, X.; Smart, S.M. Habitat invasions by alien plants: A quantitative comparison among Mediterranean, subcontinental and oceanic regions of Europe. J. Appl. Ecol. 2008, 45, 448–458. [Google Scholar] [CrossRef]
- Roura-Pascual, N.; Hui, C.; Ikeda, T.; Leday, G.; Richardson, D.M.; Carpintero, S.; Espadaler, X.; Gómez, C.; Guénard, B.; Hartley, S.; et al. Relative roles of climatic suitability and anthropogenic influence in determining the pattern of spread in a global invader. Proc. Natl. Acad. Sci. USA 2010, 108, 220–225. [Google Scholar] [CrossRef]
- Beans, C.M.; Kilkenny, F.F.; Galloway, L.F. Climate suitability and human influences combined explain the range expansion of an invasive horticultural plant. Biol. Invasions 2012, 14, 2067–2078. [Google Scholar] [CrossRef]
- Vezzoli, R.; Mercogliano, P.; Coppola, V. Climate-hydrological modelling of Calore Irpino River basin. In CMCC Research Paper; Centro Euro-Mediterraneo per i Cambiamenti Climatici: Lecce, Italy, 2015; p. 46. [Google Scholar]
- Bocchiola, D. Impact of potential climate change on crop yield and water footprint of rice in the Po valley of Italy. Agric. Syst. 2015, 139, 223–237. [Google Scholar] [CrossRef]
- Regniere, J.; Rabb, R.L.; Stinner, R.E. Popillia japonica (Coleoptera: Scarabaeidae): A mathematical model of oviposition in heterogeneous agroecosystems. Can. Entomol. 1979, 111, 1271–1280. [Google Scholar] [CrossRef]
- Allsopp, P.G. Japanese beetle, Popillia japonica Newman (Coleoptera: Scarabaeidae): Rate of movement and potential distribution of an immigrant species. Coleopt. Bull. 1996, 50, 81–95. [Google Scholar]
- Potter, D.A.; Powell, A.J.; Spicer, P.G.; Williams, D.W. Cultural Practices Affect Root-Feeding White Grubs (Coleoptera: Scarabaeidae) in Turfgrass. J. Econ. Entomol. 1996, 89, 156–164. [Google Scholar] [CrossRef]
- Tilman, D.; Fargione, J.; Wolff, B.; D’Antonio, C.; Dobson, A.; Howarth, R.; Schindler, D.; Schlesinger, W.H.; Simberloff, D.; Swackhamer, D. Forecasting Agriculturally Driven Global Environmental Change. Science 2001, 292, 281–284. [Google Scholar] [CrossRef] [PubMed]
- Newton, I. The recent declines of farmland bird populations in Britain: An appraisal of causal factors and conservation actions. Ibis 2004, 146, 579–600. [Google Scholar] [CrossRef]
- Mineau, P.; Whiteside, M. Pesticide Acute Toxicity Is a Better Correlate of U.S. Grassland Bird Declines than Agricultural Intensification. PLoS ONE 2013, 8, e57457. [Google Scholar] [CrossRef]
- Sánchez-Bayo, F.; Wyckhuys, K.A.G. Worldwide decline of the entomofauna: A review of its drivers. Biol. Conserv. 2019, 232, 8–27. [Google Scholar] [CrossRef]
- Glazer, I.; Santoiemma, G.; Battisti, A.; De Luca, F.; Fanelli, E.; Troccoli, A.; Tarasco, E.; Sacchi, S.; Bianchi, A.; Gilioli, G.; et al. Invasion of Popillia japonica in Lombardy, Italy: Interactions with soil entomopathogenic nematodes and native grubs. Agric. For. Entomol. 2022. [Google Scholar] [CrossRef]
- Ebbenga, D.N.; Burkness, E.C.; Hutchison, W.D. Optimizing the Use of Semiochemical-Based Traps for Efficient Monitoring of Popillia japonica (Coleoptera: Scarabaeidae): Validation of a Volumetric Approach. J. Econ. Entomol. 2022, 115, 869–876. [Google Scholar] [CrossRef] [PubMed]
- Mori, N.; Santoiemma, G.; Glazer, I.; Gilioli, G.; Ciampitti, M.; Cavagna, B.; Battisti, A. Management of Popillia japonica in container-grown nursery stock in Italy. Phytoparasitica 2022, 50, 83–89. [Google Scholar] [CrossRef]
- Potter, D.A.; Held, D.W. Biology and Management of the Japanese Beetle. Annu. Rev. Entomol. 2002, 47, 175–205. [Google Scholar] [CrossRef]
- Altieri, M.A.; Letourneau, D.K. Vegetation management and biological control in agroecosystems. Crop Prot. 1982, 1, 405–430. [Google Scholar] [CrossRef]
- Marianelli, L.; Paoli, F.; Torrini, G.; Mazza, G.; Benvenuti, C.; Binazzi, F.; Sabbatini Peverieri, G.; Bosio, G.; Venanzio, D.; Giacometto, E.; et al. Entomopathogenic nematodes as potential biological control agents of Popillia japonica (Coleoptera, Scarabaeidae) in Piedmont Region (Italy). J. Appl. Entomol. 2018, 142, 311–318. [Google Scholar] [CrossRef]
- Marianelli, L.; Paoli, F.; Peverieri, G.S.; Benvenuti, C.; Barzanti, G.P.; Bosio, G.; Venanzio, D.; Giacometto, E.; Roversi, P.F. Long-lasting insecticide-treated nets: A new integrated pest management approach for Popillia japonica (Coleoptera: Scarabaeidae). Integr. Environ. Assess. Manag. 2019, 15, 259–265. [Google Scholar] [CrossRef]
- Guerrero, R.; Margulis, L.; Berlanga, M. Symbiogenesis: The holobiont as a unit of evolution. Int. Microbiol. 2013, 16, 133–143. [Google Scholar]
- Kajtoch, Ł. Evolutionary and ecological signals in Wolbachia-beetle relationships: A review. Eur. J. Entomol. 2022, 119, 215–226. [Google Scholar] [CrossRef]
- Mazza, G.; Paoli, F.; Strangi, A.; Torrini, G.; Marianelli, L.; Peverieri, G.S.; Binazzi, F.; Bosio, G.; Sacchi, S.; Benvenuti, C.; et al. Hexamermis popilliae n. sp. (Nematoda: Mermithidae) parasitizing the Japanese beetle Popillia japonica Newman (Coleoptera: Scarabaeidae) in Italy. Syst. Parasitol. 2017, 94, 915–926. [Google Scholar] [CrossRef]
- Paoli, F.; Marianelli, L.; Torrini, G.; Mazza, G.; Benvenuti, C.; Bosio, G.; Venanzio, D.; Tarasco, E.; Klein, M.; Roversi, P.F. Differential susceptibility of Popillia japonica 3rd instars to Heterorhabditis bacteriophora (Italian strain) at three different seasons Biocont. Sci. Tech. 2017, 27, 439–444. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Variable | Unit | VIF |
|---|---|---|
| Altitude | m a.s.l. | >3 |
| Slope | ° | >3 |
| Bare areas | % | >3 |
| Deciduous forests | % | 1.283 |
| Coniferous forests | % | >3 |
| Grasslands | % | 1.126 |
| Shrublands | % | >3 |
| Intensive crops | % | 1.302 |
| Rice fields | % | 1.101 |
| Dense human settlements | % | 1.039 |
| Sparse human settlements | % | 1.103 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Della Rocca, F.; Milanesi, P. The Spread of the Japanese Beetle in a European Human-Dominated Landscape: High Anthropization Favors Colonization of Popillia japonica. Diversity 2022, 14, 658. https://doi.org/10.3390/d14080658
Della Rocca F, Milanesi P. The Spread of the Japanese Beetle in a European Human-Dominated Landscape: High Anthropization Favors Colonization of Popillia japonica. Diversity. 2022; 14(8):658. https://doi.org/10.3390/d14080658
Chicago/Turabian StyleDella Rocca, Francesca, and Pietro Milanesi. 2022. "The Spread of the Japanese Beetle in a European Human-Dominated Landscape: High Anthropization Favors Colonization of Popillia japonica" Diversity 14, no. 8: 658. https://doi.org/10.3390/d14080658
APA StyleDella Rocca, F., & Milanesi, P. (2022). The Spread of the Japanese Beetle in a European Human-Dominated Landscape: High Anthropization Favors Colonization of Popillia japonica. Diversity, 14(8), 658. https://doi.org/10.3390/d14080658