
Spatial-Temporal Variations in Dietary Consumption of Two Dominant Rodent Species (Rhabdomys dilectus and Lophuromys acquilus) on Mount Kilimanjaro, Tanzania


Abstract
:1. Introduction
2. Materials and Methods
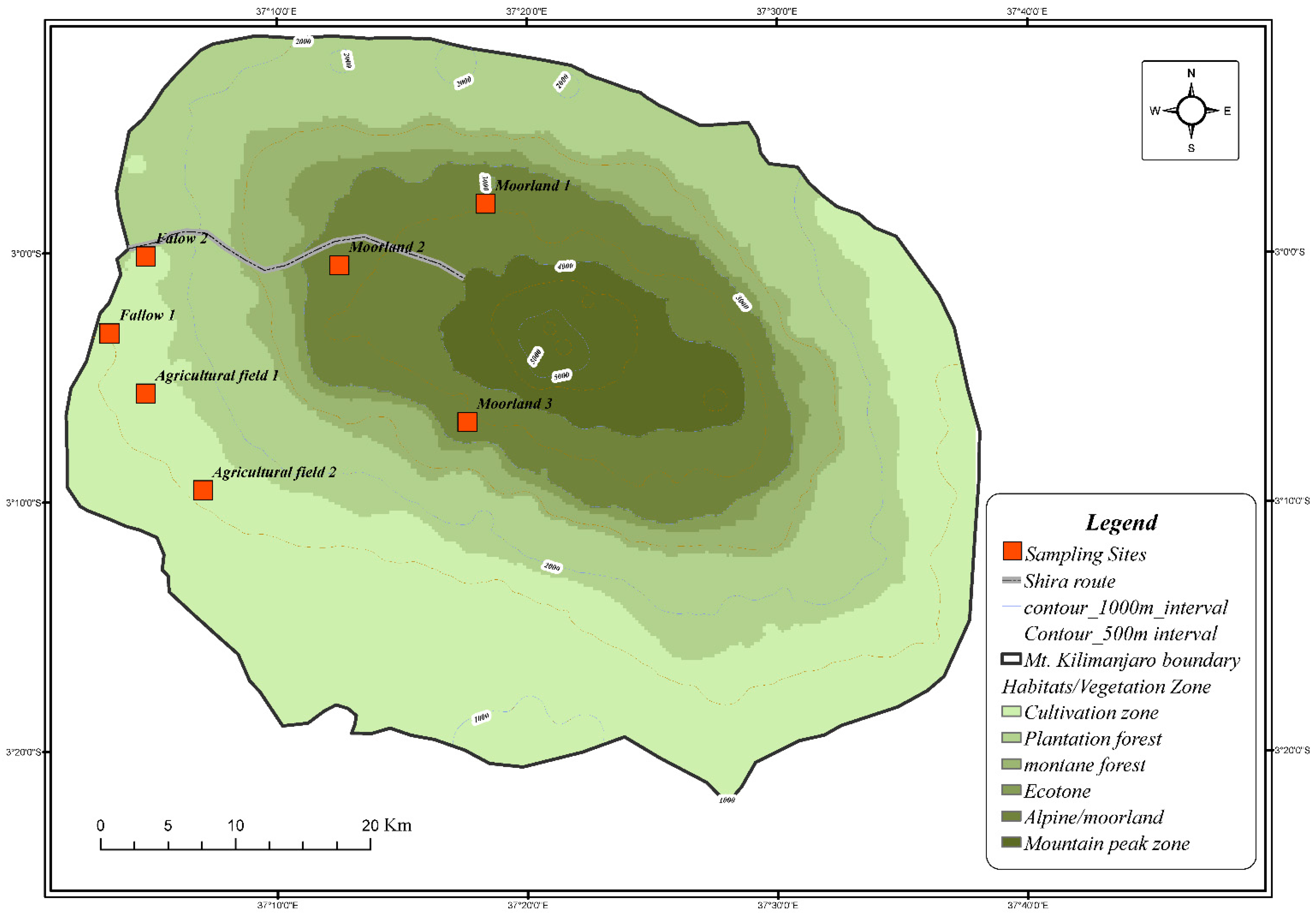
2.1. Study Area
2.2. Rodent Trapping
2.3. Assessment of Food Availability
2.4. Stomach Dissection and Data Processing
2.5. Data Analysis
3. Results
3.1. Species Composition
3.2. Percentage Occurrence
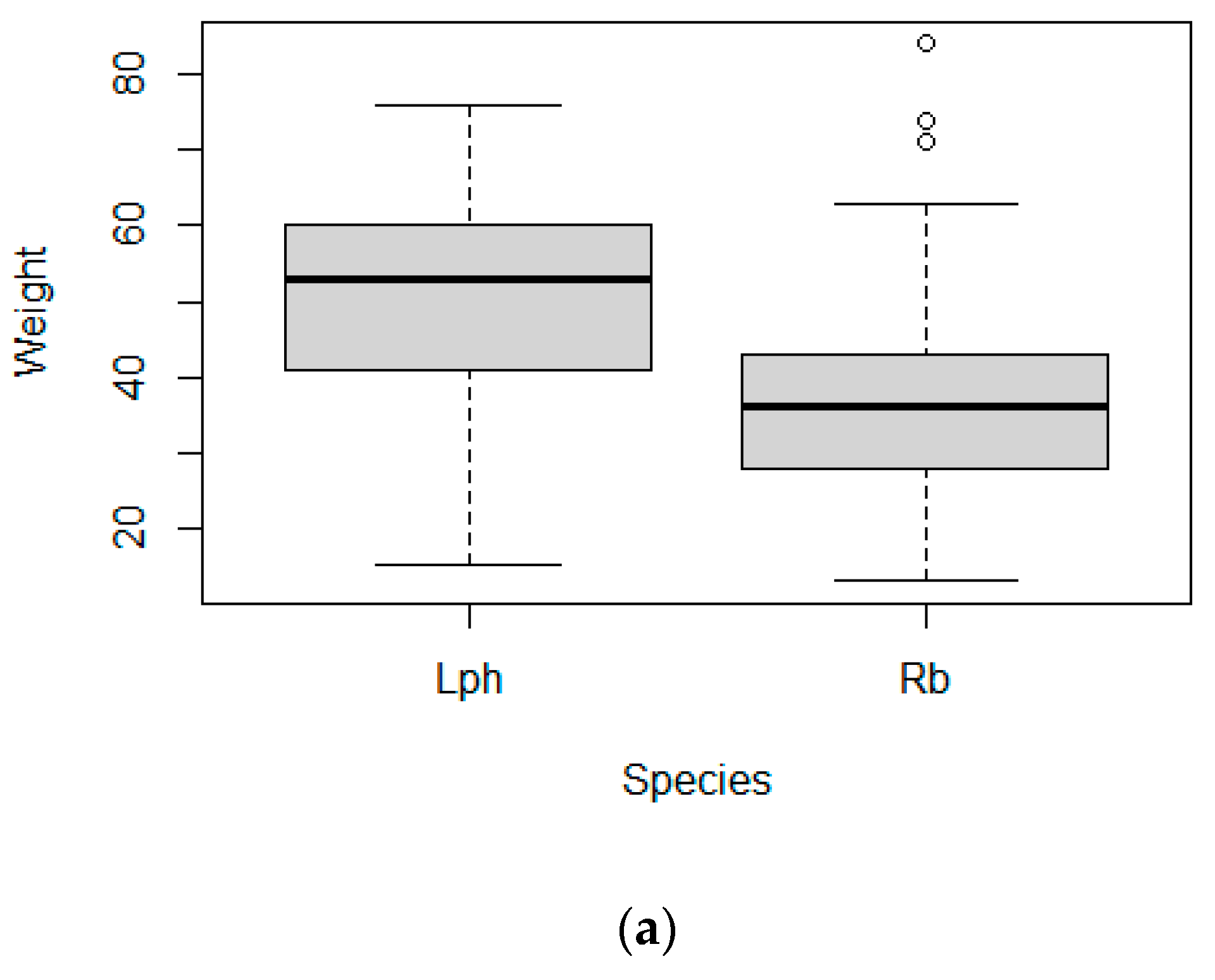
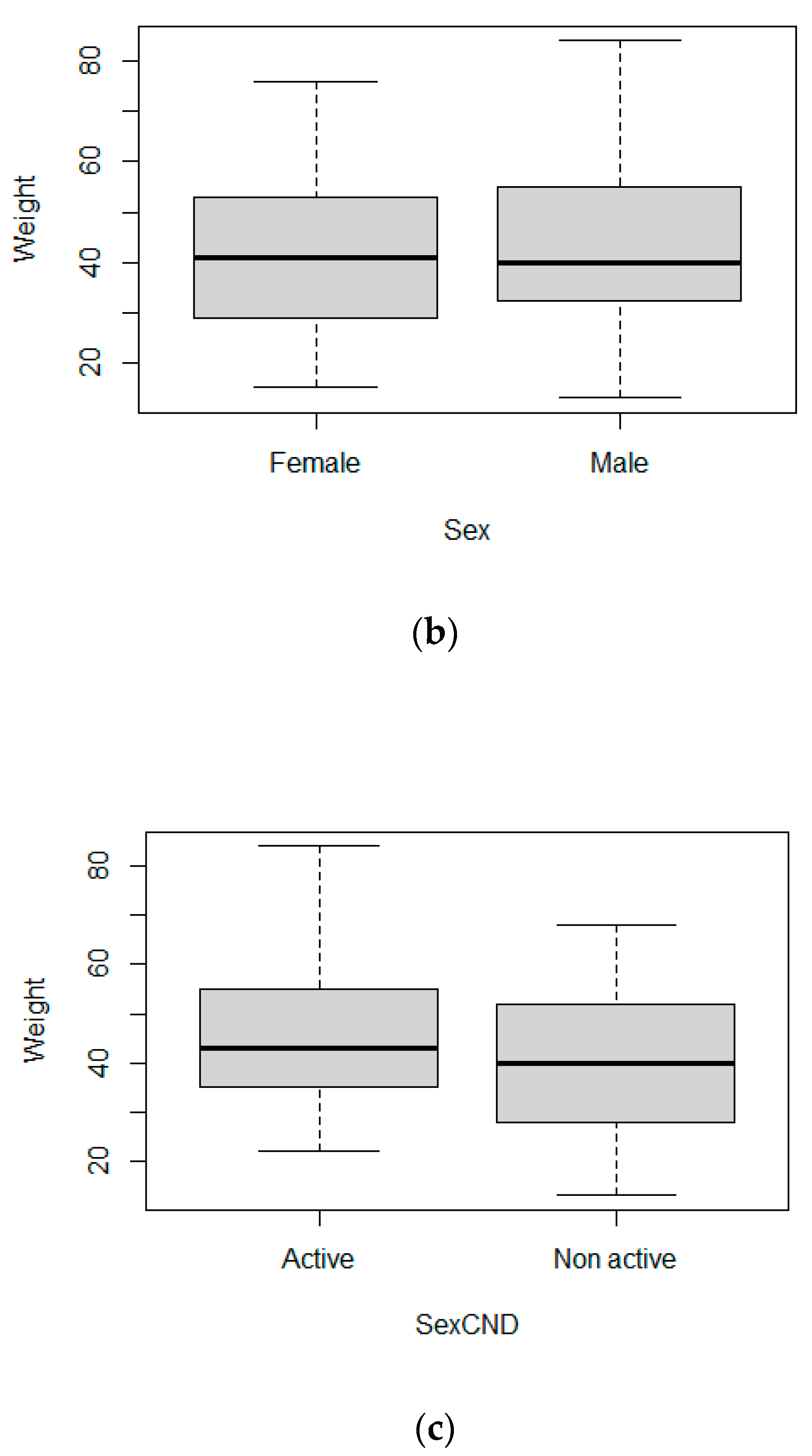
3.3. Body Size
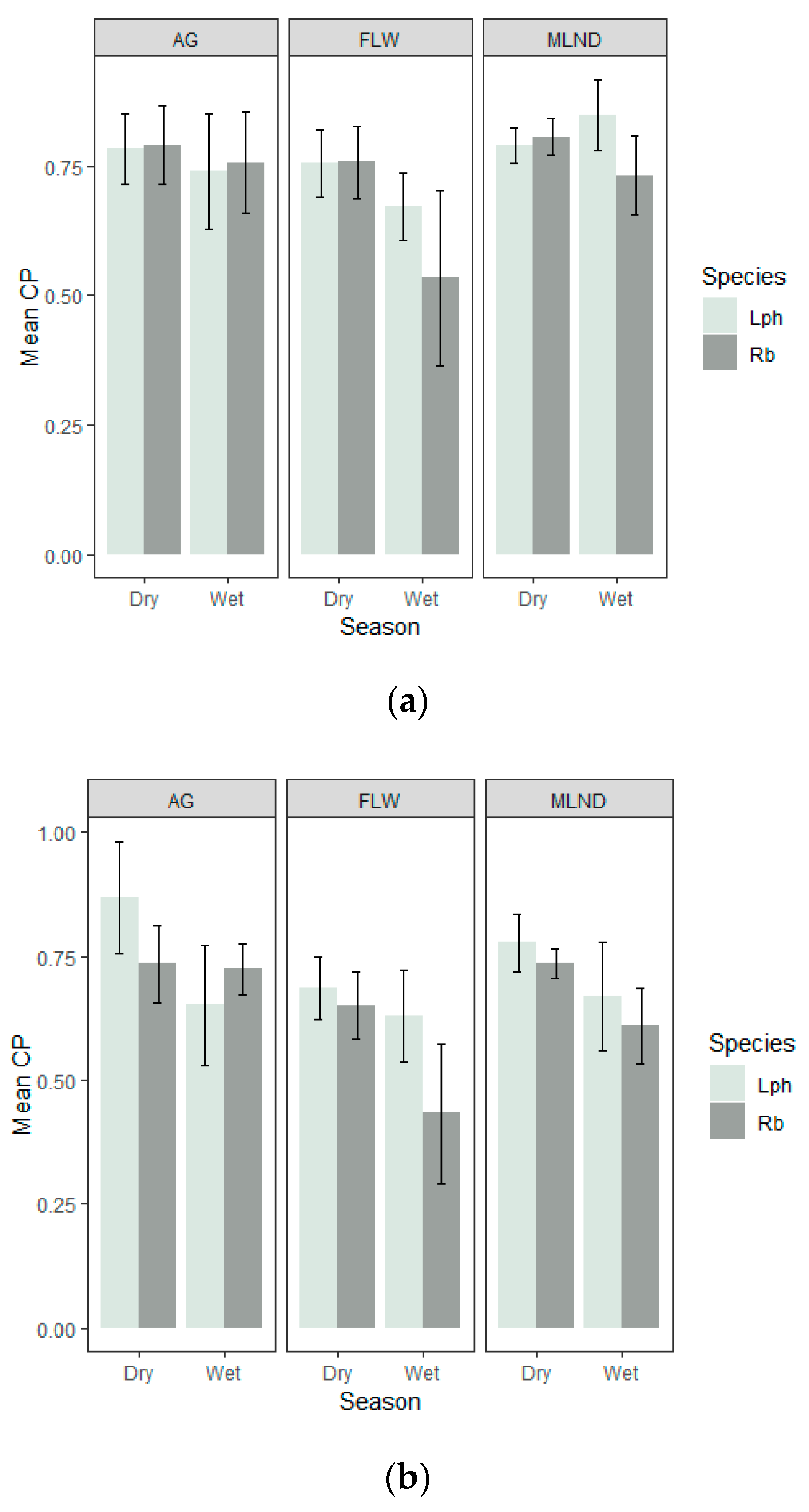
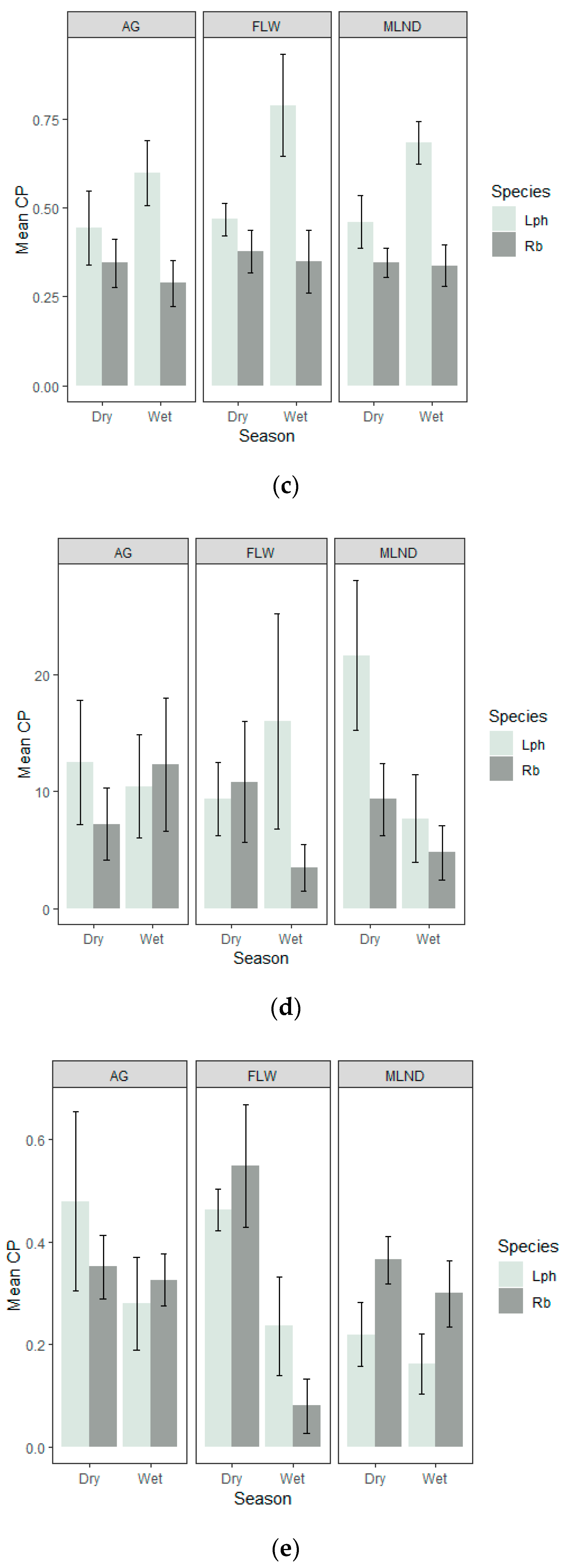
3.4. Percentage Contribution
3.5. Relative Importance
3.6. Diet Diversity, Niche Breadth, and Overlap
4. Discussion
5. Conclusions and Recommendations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Datiko, D.; Bekele, A.; Belay, G. Feeding ecology of pest rodents from Arbaminch forest and farmlands, Ethiopia. SINET Ethiop. J. Sci. 2008, 30, 127–134. [Google Scholar] [CrossRef]
- Pianka, E.R. Niche Overlap and Diffuse Competition. Proc. Natl. Acad. Sci. USA 1974, 71, 2141–2145. [Google Scholar] [CrossRef] [PubMed]
- Sassi, P.L.; Borghi, C.E.; Dacar, M.A.; Bozinovic, F. Geographic and seasonal variability in feeding behaviour of a small herbivorous rodent. Acta Thériol. 2010, 56, 35–43. [Google Scholar] [CrossRef]
- Finke, D.L.; Snyder, W.E. Niche Partitioning Increases Resource Exploitation by Diverse Communities. Science 2008, 321, 1488–1490. [Google Scholar] [CrossRef]
- Curtis, B.; Perrin, M. Food preferences of the vlei rat (Otomys Irroratus) and the four-striped mouse (Rhabdomys pumilio). S. Afr. J. Zool. 1979, 14, 224–229. [Google Scholar] [CrossRef]
- Luo, J.; Fox, B.J. Seasonal and successional dietary shifts of two sympatric rodents in coastal heathland: A possible mechanism for coexistence. Austral Ecol. 1996, 21, 121–132. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Massawe, A.W.; Kennis, J.; Crauwels, D.; Eiseb, S.; Mahlaba, T.A.; Monadjem, A.; Makundi, R.H.; Katakweba, A.A.; Leirs, H.; et al. Differences in diet between two rodent species, Mastomys natalensisandGerbilliscus vicinus, in fallow land habitats in central Tanzania. Afr. Zool. 2011, 46, 387–392. [Google Scholar] [CrossRef]
- Soininen, E.M.; Ravolainen, V.T.; Bråthen, K.A.; Yoccoz, N.G.; Gielly, L.; Ims, R.A. Arctic Small Rodents Have Diverse Diets and Flexible Food Selection. PLoS ONE 2013, 8, e68128. [Google Scholar] [CrossRef]
- Clausnitzer, V.; Churchfield, S.; Hutterer, R. Habitat occurrence and feeding ecology of Crocidura montis and Lophuromys flavopunctatus on Mt. Elgon, Uganda. Afr. J. Ecol. 2003, 41, 1–8. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Mahlaba, T.A.; Massawe, A.W.; Kennis, J.; Crauwels, D.; Eiseb, S.; Monadjem, A.; Makundi, R.H.; Katakweba, A.A.S.; Leirs, H.; et al. Dietary differences of the multimammate mouse, Mastomys natalensis (Smith, 1834), across different habitats and seasons in Tanzania and Swaziland. Wildl. Res. 2011, 38, 640–646. [Google Scholar] [CrossRef]
- Odhiambo, R.O.; Makundi, R.H.; Leirs, H.; Verhagen, R. Dietary selection in Mastomys natalensis (Rodentia: Muridae) in the maize agro-ecosystems of central and southwestern Tanzania. Mammalia 2008, 72. [Google Scholar] [CrossRef]
- Mulungu, L.; Mlyashimbi, E.; Ngowo, V.; Mdangi, M.; Katakweba, A.; Tesha, P.; Mrosso, F.; Mchomvu, M.; Kilonzo, B.; Belmain, S. Food preferences of the multi-mammate mouse, Mastomys natalensis, in irrigated rice habitats in Tanzania. Int. J. Pest Manag. 2014, 60, 1–8. [Google Scholar] [CrossRef]
- Bomford, M. Food and reproduction of wild house mice. 2. A field experiment to examine the effect of food availability and food quality on breeding in spring. Wildl. Res. 1987, 14, 197–206. [Google Scholar] [CrossRef]
- Bomford, M. Food and reproduction of wild house mice. 1. Diet and breeding seasons in various habitats on irrigated cereal farms in New South Wales. Wildl. Res. 1987, 14, 183–196. [Google Scholar] [CrossRef]
- Tann, C.; Singleton, G.; Coman, B. Diet of the House Mouse, Mus domesticus, in the Mallee Wheatlands of North-Western Victoria. Wildl. Res. 1991, 18, 1–12. [Google Scholar] [CrossRef]
- Htwe, N.M.; Singleton, G.R. Is quantity or quality of food influencing the reproduction of rice-field rats in the Philippines? Wildl. Res. 2014, 41, 56–63. [Google Scholar] [CrossRef]
- Brettschneider, H.; Julius, R.S.; Bastos, A.D.; Chimimba, C.T. Dynamics of Rodent-Borne Zoonotic Diseases and Their Reservoir Hosts: Invasive Rattus in South Africa. Proc. Vertebr. Pest Conf. 2012, 25. [Google Scholar] [CrossRef]
- Meerburg, B.G.; Singleton, G.; Kijlstra, A. Rodent-borne diseases and their risks for public health. Crit. Rev. Microbiol. 2009, 35, 221–270. [Google Scholar] [CrossRef]
- Davies, M.; Béné, C.; Arnall, A.; Tanner, T.; Newsham, A.; Coirolo, C. Promoting resilient livelihoods through adaptive social protection: Lessons from 124 programmes in South Asia. Dev. Policy Rev. 2013, 31, 27–58. [Google Scholar] [CrossRef]
- Makundi, R.H.; Massawe, A.W. Ecologically based rodent management in Africa: Potential and challenges. Wildl. Res. 2011, 38, 588–595. [Google Scholar] [CrossRef]
- Ralaizafisoloarivony, N.A.; Kimaro, D.N.; Kihupi, N.I.; Mulungu, L.S.; Leirs, H.; Msanya, B.M.; Deckers, J.A.; Gulinck, H. Vegetation habitats and small mammals in a plague endemic area in Western Usambara Mountains, Tanzania. Tanzan. J. Health Res. 2014, 16, 194–205. [Google Scholar] [CrossRef] [PubMed]
- Oguge, N.O. Diet, seasonal abundance and microhabitats of Praomys (Mastomys) natalensis (Rodentia: Muridae) and other small rodents in a Kenyan sub-humid grassland community. Afr. J. Ecol. 1995, 33, 211–223. [Google Scholar] [CrossRef]
- Leirs, H.; Verhagen, R.; Verheyen, W.; Mwanjabe, P.; Mbise, T. Forecasting Rodent Outbreaks in Africa: An Ecological Basis for Mastomys Control in Tanzania. J. Appl. Ecol. 1996, 33, 937. [Google Scholar] [CrossRef]
- Mwanjabe, P.S.; Sirima, F.B.; Lusingu, J. Crop losses due to outbreaks of Mastomys natalensis (Smith, 1834) Muridae, Rodentia, in the Lindi Region of Tanzania. Int. Biodeterior. Biodegrad. 2002, 49, 133–137. [Google Scholar] [CrossRef]
- Shiels, A.B.; Flores, C.A.; Khamsing, A.; Krushelnycky, P.D.; Mosher, S.M.; Drake, D.R. Dietary niche differentiation among three species of invasive rodents (Rattus rattus, R. exulans, Mus musculus). Biol. Invasions 2012, 15, 1037–1048. [Google Scholar] [CrossRef]
- Corti, M.; Castiglia, R.; Verheyen, W. A note on three new cytotypes of Lophuromys flavopunctatus sensu lato (Rodentia, Muridae) from Tanzania. Mammalia 2004, 68, 69–74. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Makundi, R.H.; Massawe, A.W.; Machang’U, R.S.; Mbije, N.E. Diversity and distribution of rodent and shrew species associated with variations in altitude on Mount Kilimanjaro, Tanzania. Mammalia 2008, 72. [Google Scholar] [CrossRef]
- Stanley, W.T.; Rogers, M.A.; Kihaule, P.M.; Munissi, M.J. Elevational Distribution and Ecology of Small Mammals on Africa’s Highest Mountain. PLoS ONE 2014, 9, e109904. [Google Scholar] [CrossRef]
- Gebrezgiher, G.B.; Makundi, R.H.; Meheretu, Y.; Mulungu, L.S.; Katakweba, A.A.S. A Decade-Long Change in the Elevational Distribution of Non-Volant Small Mammals on Mount Meru, Tanzania. Diversity 2022, 14, 454. [Google Scholar] [CrossRef]
- Hanney, P. The Harsh-Furred Rat in Nyasaland. J. Mammal. 1964, 45, 345–358. [Google Scholar] [CrossRef]
- Bekele, A.; Corti, M. Multivariate morphometries of the Ethiopian populations of harsh-furred rat (Lophuromys: Mammalia, Rodentia). J. Zool. 1994, 232, 675–689. [Google Scholar] [CrossRef]
- Ademola, O.J.; Massawe, A.W.; Mulungu, L.S.; Makundi, R.H. Dietary composition of rodents across different habitats in the Ukaguru Mountains, Tanzania. Afr. J. Ecol. 2022. [Google Scholar] [CrossRef]
- Krug, C.B. Reproduction of Rhabdomys pumilio in the Namib Desert: Pattern and possible control. Basic Appl. Dryland Res. 2007, 1, 67–85. [Google Scholar] [CrossRef]
- Perrin, M.; Kotler, B. A test of five mechanisms of species coexistence between rodents in a southern African savanna. Afr. Zool. 2005, 40, 55–61. [Google Scholar] [CrossRef]
- Henke, Z.; Sahd, L.; Matthee, S.; Kotzé, S.H. Morphometric analysis of the gastrointestinal tract of four African muroid rodent species (Rhabdomys dilectus, Rhabdomys pumilio, Aethomys chrysophilus, and Lemniscomys rosalia). J. Morphol. 2018, 279, 1282–1289. [Google Scholar] [CrossRef]
- Mulungu, L.S.; Ngowo, V.; Mdangi, M.; Katakweba, A.S.; Tesha, P.; Mrosso, F.P.; Mchomvu, M.; Sheyo, P.M.; Kilonzo, B.S. Population dynamics and breeding patterns of multimammate mouse, Mastomys natalensis (Smith 1834), in irrigated rice fields in Eastern Tanzania. Pest Manag. Sci. 2012, 69, 371–377. [Google Scholar] [CrossRef]
- Field, A.C. Seasonal changes in reproduction, diet and body composition of two equatorial rodents. Afr. J. Ecol. 1975, 13, 221–235. [Google Scholar] [CrossRef]
- Leirs, H.; Verhagen, R.; Verheyen, W. The basis of reproductive seasonally in Mastomys rats (Rodentia: Muridae) in Tanzania. J. Trop. Ecol. 1994, 10, 55–66. [Google Scholar] [CrossRef]
- Thomas, S.; Mulungu, L.; Soka, G. Influence of Vegetation Structure, Seasonality, and Soil Physical Properties on Rodent Species Diversity and Community Assemblages on West Mount Kilimanjaro, Tanzania. Authorea Prepr. 2021. [Google Scholar] [CrossRef]
- Monadjem, A. Stomach contents of 19 species of small mammals from Swaziland. S. Afr. J. Zool. 1997, 32, 23–26. [Google Scholar] [CrossRef]
- Mulangu, F.; Kraybill, D. Climate Change and the Future of Mountain Farming on Mt. Kilimanjaro. In The Future of Mountain Agriculture; Springer: Berlin/Heidelberg, Germany, 2012; pp. 73–88. [Google Scholar] [CrossRef]
- Hemp, A. Continuum or zonation? Altitudinal gradients in the forest vegetation of Mt. Kilimanjaro. Plant Ecol. 2005, 184, 27–42. [Google Scholar] [CrossRef]
- Monica, S.; Flora, J.M.; Joseph, O.O.; Eivin, R. Small mammal community demography and reproductive seasonality under protection, pastoralism and agriculture in the Serengeti Ecosystem, Tanzania. Int. J. Biodivers. Conserv. 2020, 12, 253–269. [Google Scholar] [CrossRef]
- Welegerima, K.; Meheretu, Y.; Haileselassie, T.H.; Gebre, B.; Kidane, D.; Massawe, A.W.; Mbije, N.E.; Makundi, R.H. Abundance and microhabitat use of rodent species in crop fields and bushland in Ethiopia. J. Vertebr. Biol. 2020, 69, 1–12. [Google Scholar] [CrossRef]
- Shorrocks, B. Mammals of Africa Volume 1. Introductory Chapters and Afrotheria Jonathan Kingdon, David Happold, Michael Hoffman Thomas Butynski, Meredith Happold and Jan Kalina. Bloomsbury Publishing, London (2013) 351 pp. ISBN 978-1-4081-2251-8. Afr. J. Ecol. 2013, 52, 254–255. [Google Scholar] [CrossRef]
- Sikes, R.S. The Animal Care and Use Committee of the American Society of Mammalogists 2016 Guidelines of the American Society of Mammalogists for the use of wild mammals in research and education. J. Mammal. 2016, 97, 663–688. [Google Scholar] [CrossRef]
- Mlyashimbi, E.C.M.; Mariën, J.; Kimaro, D.N.; Tarimo, A.J.P.; Isabirye, M.; Makundi, R.H.; Massawe, A.W.; Mdangi, M.E.; Kifumba, D.; Nakiyemba, A.; et al. Relationships between seasonal changes in diet of Multimammate rat (Mastomys natalensis) and its breeding patterns in semi-arid areas in Tanzania. Cogent Food Agric. 2018, 4, 1507509. [Google Scholar] [CrossRef]
- Smith, V.; Avenant, N.; Chown, S. The diet and impact of house mice on a sub-Antarctic island. Polar Biol. 2002, 25, 703–715. [Google Scholar] [CrossRef]
- Cooper, R.; Skinner, J. Importance of Termites in the diet of the Aardwolf Proteles Cristatus in South Africa. S. Afr. J. Zool. 1979, 14, 5–8. [Google Scholar] [CrossRef]
- Levins, R. Evolution in Changing Environment; Princeton University Press: Princeton, NJ, USA, 1968. [Google Scholar]
- MacArthur, R.H.; Pianka, E.R. On Optimal Use of a Patchy Environment. Am. Nat. 1966, 100, 603–609. [Google Scholar] [CrossRef]
- Winemiller, K.; Pianka, E.R. Organization in Natural Assemblages of Desert Lizards and Tropical Fishes. Ecol. Monogr. 1990, 60, 27–55. [Google Scholar] [CrossRef]
- R Core Team. R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2013; Available online: https://cran.r-project.org/bin/windows/base/old/3.6.2 (accessed on 19 March 2022).
- Schradin, C.; Pillay, N. Female striped mice (Rhabdomys pumilio) change their home ranges in response to seasonal variation in food availability. Behav. Ecol. 2006, 17, 452–458. [Google Scholar] [CrossRef]
- Clausnitzer, V.; Denys, C.; Granjon, L.; Poulet, A. Rodents of the afro-alpine zone of Mt. Elgon. Afr. Small Mamm. 2001, 427–443. [Google Scholar]
- Clausnitzer, V.; Kityo, R. Altitudinal distribution of rodents (Muridae and Gliridae) on Mt Elgon, Uganda. Trop. Zool. 2001, 14, 95–118. [Google Scholar] [CrossRef]
- Krug, C.B. Survival in the Namib: Adaptations of the striped mouse to an arid environment. Trans. R. Soc. S. Afr. 2004, 59, 93–98. [Google Scholar] [CrossRef]
- Taylor, K.D.; Green, M.G. The influence of rainfall on diet and reproduction in four African rodent species. J. Zool. 1976, 180, 367–389. [Google Scholar] [CrossRef] [PubMed]
- Makundi, R.H.; Massawe, A.W.; Mulungu, L.S. Breeding seasonality and population dynamics of three rodent species in the Magamba Forest Reserve, Western Usambara Mountains, north-east Tanzania. Afr. J. Ecol. 2007, 45, 17–21. [Google Scholar] [CrossRef]
- Wells, K.; Corlett, R.T.; Lakim, M.B.; Kalko, E.K.V.; Pfeiffer, M. Seed consumption by small mammals from Borneo. J. Trop. Ecol. 2009, 25, 555–558. [Google Scholar] [CrossRef]
- Massawe, A.W.; Mrosso, F.P.; Makundi, R.H.; Mulungu, L. Breeding patterns of Arvicanthis neumanni in central Tanzania. Afr. J. Ecol. 2008, 46, 320–324. [Google Scholar] [CrossRef]
- Massawe, A.W.; Makundi, R.H.; Mulungu, L.S.; Katakweba, A.; Shayo, T.N. Breeding Dynamics of Rodent Species Inhabiting Farm—Fallow Mosaic Fields in Central Tanzania†. Afr. Zool. 2012, 47, 128–137. [Google Scholar] [CrossRef]
- Kitessa, H.; Tsegaye, G.; Hundera, K.; Gadisa, T. The reproductive and feeding ecology of rodents in Sekoru district, Southwest Ethiopia. J. Ecol. Nat. Environ. 2015, 7, 188–195. [Google Scholar] [CrossRef]
- Cole, L.R. Foods and foraging places of rats (Rodentia: Muridae) in the lowland evergreen forest of Ghana. J. Zool. 1975, 175, 453–471. [Google Scholar] [CrossRef]
- Brown, J.S. Desert Rodent Community Structure: A Test of Four Mechanisms of Coexistence. Ecol. Monogr. 1989, 59, 1–20. [Google Scholar] [CrossRef]
- Lema, R.; Magige, F.J. The influence of agricultural activities on the diversity of rodents in Kindoroko forest reserve and surrounding areas, North Pare Mountains, Tanzania. Tanzan. J. Sci. 2018, 44, 97–106. [Google Scholar]
- Petraitis, P.S. Likelihood Measures of Niche Breadth and Overlap. Ecology 1979, 60, 703–710. [Google Scholar] [CrossRef]
- May, R.M.; Mac Arthur, R.H. Niche Overlap as a Function of Environmental Variability. Proc. Natl. Acad. Sci. USA 1972, 69, 1109–1113. [Google Scholar] [CrossRef]
- Christian, D.P. Diurnal Activity of the Four-Striped Mouse, Rhabdomys Pumilio. Zool. Afr. 1977, 12, 238–239. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Agricultural Field | Fallow Land | Moorland | Grand Total |
|---|---|---|---|---|
| Arvicanths niloticus | 0(0) | 1(0.93) | 0(0) | 1(0.35) |
| Crocidura spp. | 1(1.1) | 2(1.85) | 2(2.3) | 5(1.75) |
| Dendromus spp. | 0(0) | 1(0.93) | 3(3.45) | 4(1.4) |
| Aethomys kaiseri | 0(0) | 2(1.85) | 0(0) | 2(0.7) |
| Gramomys dolichurus | 0(0) | 4(3.7) | 0(0) | 4(1.4) |
| Lemniscomys striatus | 0(0) | 16(14.81) | 0(0) | 16(5.59) |
| Lophuromysacquilus | 19(20.88) | 25(23.15) | 28(32.18) | 72(25.17) |
| Mastomys natalensis | 41(45.05) | 29(26.85) | 0(0) | 70(24.48) |
| Otomys spp. | 0(0) | 1(0.93) | 1(1.15) | 2(0.7) |
| Pelomys falax | 0(0) | 1(0.93) | 0(0) | 1(0.35) |
| Praomys delectroum | 1(1.1) | 4(3.7) | 1(1.15) | 6(2.1) |
| Rhabdomys dilectus | 29(31.87) | 22(20.37) | 52(59.77) | 103(36.01) |
| Grand Total | 91(100) | 108(100) | 87(100) | 286(100) |
| Species and Season | ||||
|---|---|---|---|---|
| L. acquilus | R. dilectus | |||
| Dry (n = 8) | Wet (n = 11) | Dry (n = 18) | Wet (11) | |
| Seeds/Starch | 29.83 | 23.80 | 28.86 | 29.70 |
| Vegetative materials | 28.98 | 32.32 | 32.38 | 33.03 |
| Roots | 5.33 | 4.22 | 2.30 | 4.84 |
| Hairs | 8.31 | 4.41 | 6.40 | 5.81 |
| Invertebrates | 13.05 | 16.23 | 10.99 | 6.89 |
| Others/Unidentified | 14.49 | 19.03 | 19.07 | 19.73 |
| Species and Season | ||||
|---|---|---|---|---|
| L. acquilus | R. dilectus | |||
| Dry (n = 15) | Wet (n = 10) | Dry (n = 12) | Wet (n = 10) | |
| Seeds/Starch | 23.46 | 19.52 | 25.15 | 16.44 |
| Vegetative materials | 29.04 | 27.71 | 29.66 | 22.31 |
| Roots | 4.18 | 5.29 | 4.73 | 1.37 |
| Hairs | 9.76 | 2.65 | 12.21 | 0.52 |
| Invertebrates | 27.64 | 31.57 | 14.72 | 14.40 |
| Others/Unidentified | 18.92 | 20.26 | 16.53 | 31.96 |
| Species and Season | ||||
|---|---|---|---|---|
| L. acquilus | R. dilectus | |||
| Dry (n = 15) | Wet (n = 13) | Dry (n = 30) | Wet (n = 22) | |
| Seeds/Starch | 27.66 | 23.27 | 29.58 | 23.03 |
| Vegetative materials | 29.75 | 37.85 | 33.10 | 32.93 |
| Roots | 11.08 | 1.92 | 2.80 | 1.41 |
| Hairs | 2.23 | 1.32 | 6.42 | 5.35 |
| Invertebrates | 13.02 | 14.54 | 8.49 | 10.11 |
| Others/Unidentified | 16.27 | 21.09 | 19.62 | 27.17 |
| Species | AG | FLW | MLND | |||
|---|---|---|---|---|---|---|
| Dry | Wet | Dry | Wet | Dry | Wet | |
| L. acquilus | 0.83 (1.7) | 0.87 (1.72) | 0.83 (1.7) | 0.95 (1.77) | 0.9 (1.75) | 0.86 (1.70) |
| R.dilectus | 0.78 (1.66) | 0.78 (1.67) | 0.80 (1.69) | 0.81 (1.67) | 0.79 (1.67) | 0.78 (1.66) |
| Niche overlap (O jk) | 0.94 | 0.84 | 0.98 | 0.86 | 0.88 | 0.84 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Thomas, S.M.; Soka, G.E.; Mulungu, L.S.; Makonda, F.B.S. Spatial-Temporal Variations in Dietary Consumption of Two Dominant Rodent Species (Rhabdomys dilectus and Lophuromys acquilus) on Mount Kilimanjaro, Tanzania. Diversity 2022, 14, 659. https://doi.org/10.3390/d14080659
Thomas SM, Soka GE, Mulungu LS, Makonda FBS. Spatial-Temporal Variations in Dietary Consumption of Two Dominant Rodent Species (Rhabdomys dilectus and Lophuromys acquilus) on Mount Kilimanjaro, Tanzania. Diversity. 2022; 14(8):659. https://doi.org/10.3390/d14080659
Chicago/Turabian StyleThomas, Suzana M., Geofrey E. Soka, Loth S. Mulungu, and Fortunatus B. S. Makonda. 2022. "Spatial-Temporal Variations in Dietary Consumption of Two Dominant Rodent Species (Rhabdomys dilectus and Lophuromys acquilus) on Mount Kilimanjaro, Tanzania" Diversity 14, no. 8: 659. https://doi.org/10.3390/d14080659
APA StyleThomas, S. M., Soka, G. E., Mulungu, L. S., & Makonda, F. B. S. (2022). Spatial-Temporal Variations in Dietary Consumption of Two Dominant Rodent Species (Rhabdomys dilectus and Lophuromys acquilus) on Mount Kilimanjaro, Tanzania. Diversity, 14(8), 659. https://doi.org/10.3390/d14080659