
Alien Travel Companies: The Case of Two Sea Slugs and One Bryozoan in the Mediterranean Sea



Abstract
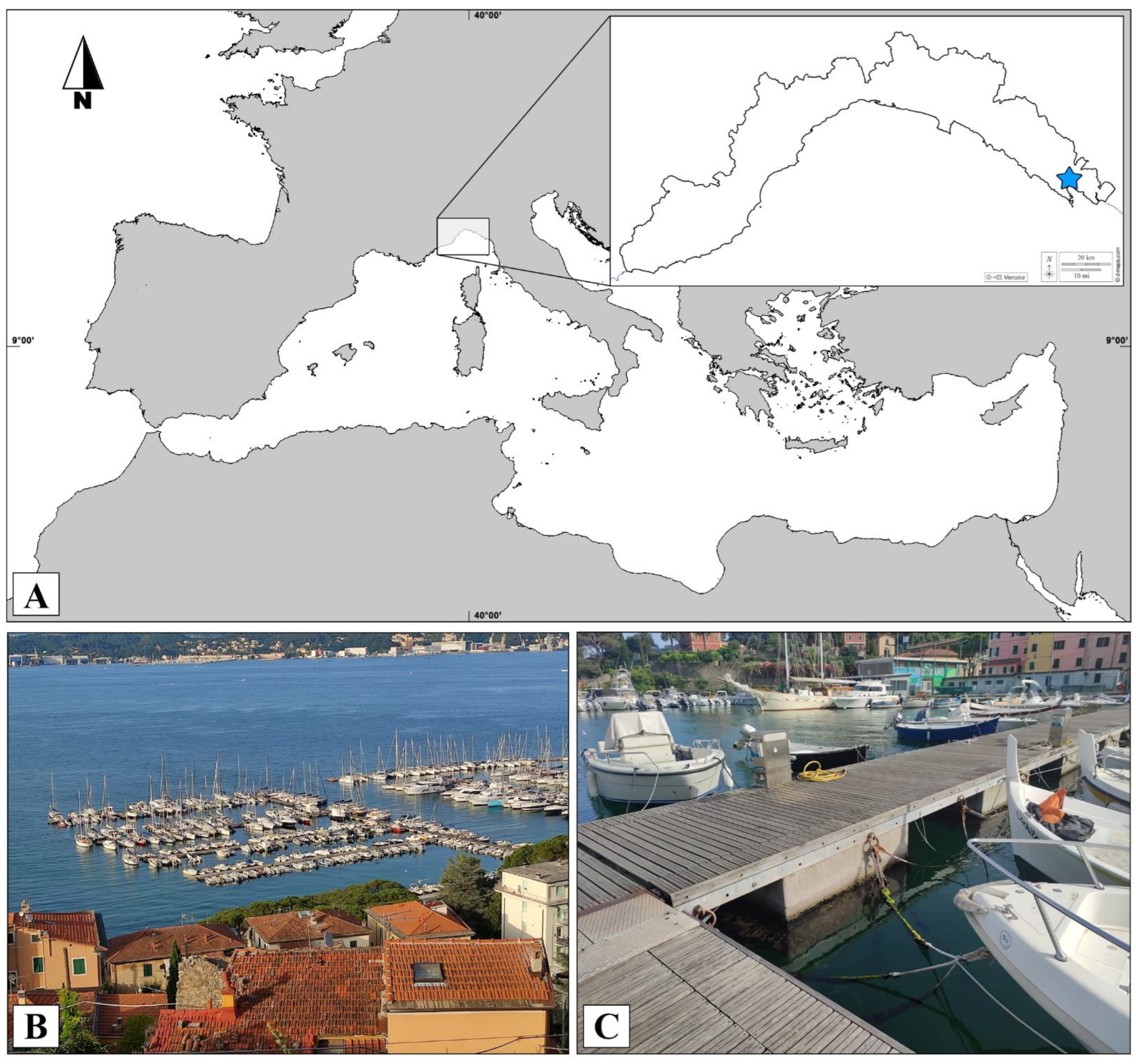
:1. Introduction
2. Materials and Methods
2.1. DNA Barcoding
2.2. Phylogenetic Analyses
3. Results
3.1. Morphological Identification
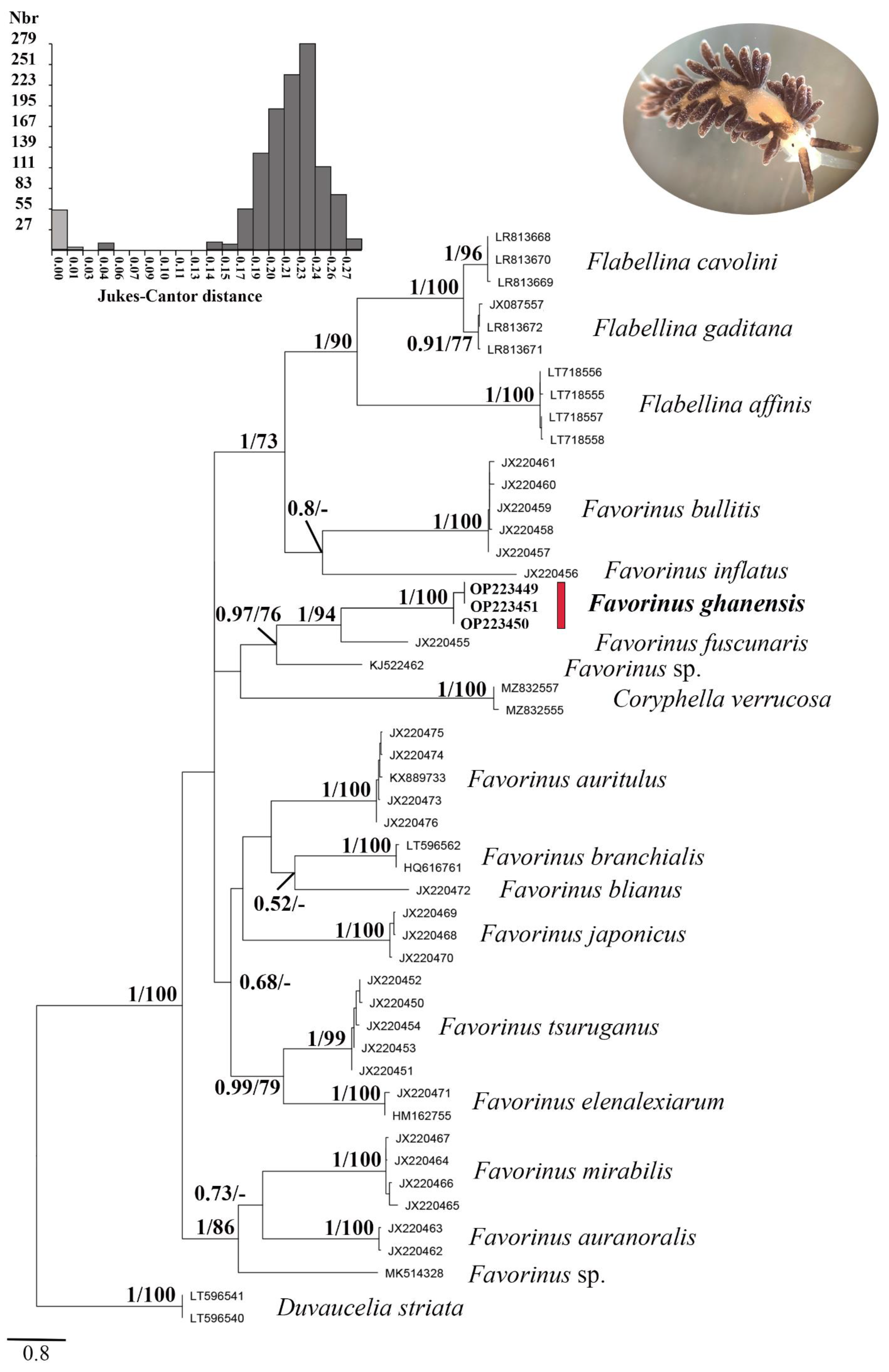
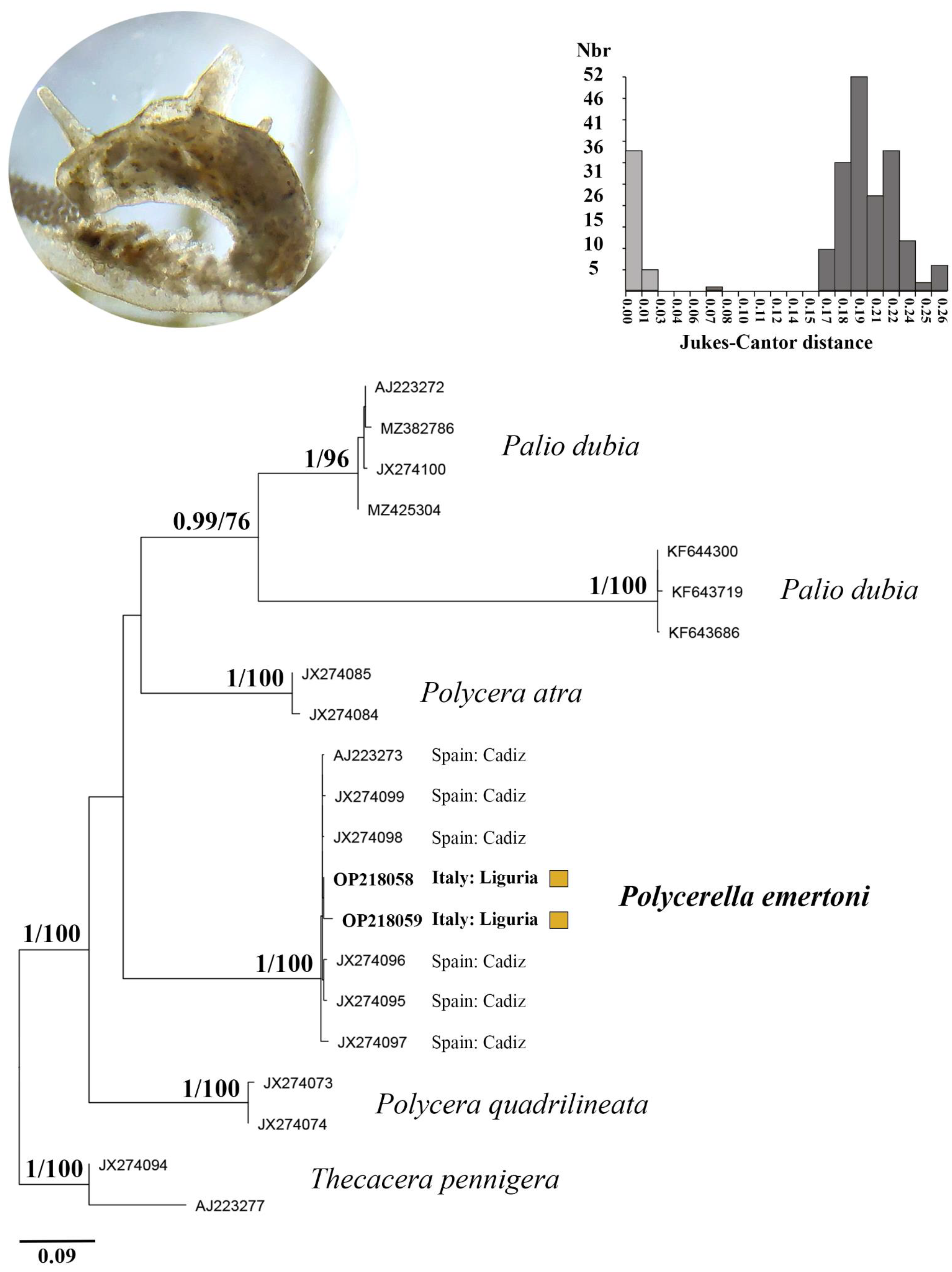
3.2. Molecular Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The Biodiversity of the Mediterranean Sea: Estimates, Patterns, and Threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danovaro, R.; Company, J.B.; Corinaldesi, C.; D’Onghia, G.; Galil, B.; Gambi, C.; Gooday, A.J.; Lampadariou, N.; Luna, G.M.; Morigi, C.; et al. Deep-Sea Biodiversity in the Mediterranean Sea: The Known, the Unknown, and the Unknowable. PLoS ONE 2010, 5, e11832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lejeusne, C.; Chevaldonné, P.; Pergent-Martini, C.; Boudouresque, C.F.; Pérez, T. Climate change effects on a miniature ocean: The highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 2010, 25, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, C.N.; Morri, C. Marine Biodiversity of the Mediterranean Sea: Situation, Problems and Prospects for Future Research. Mar. Pollut. Bull. 2000, 40, 367–376. [Google Scholar] [CrossRef]
- Mouillot, D.; Albouy, C.; Guilhaumon, F.; Lasram, F.B.R.; Coll, M.; Devictor, V.; Meynard, C.N.; Pauly, D.; Tomasini, J.A.; Troussellier, M.; et al. Protected and Threatened Components of Fish Biodiversity in the Mediterranean Sea. Curr. Biol. 2011, 21, 1044–1050. [Google Scholar] [CrossRef] [Green Version]
- Costello, M.J.; Tsai, P.; Wong, P.S.; Cheung, A.K.L.; Basher, Z.; Chaudhary, C. Marine biogeographic realms and species endemicity. Nat. Commun. 2017, 8, 1057. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Cinar, M.E.; Pancucci-Papadopoulou, M.A.; Harmelin, J.G.; Furnari, G.; Andaloro, F.; Bellou, N.; Streftaris, N.; Zibrowius, H. Annotated list of marine alien species in the Mediterranean with records of the worst invasive species. Mediterr. Mar. Sci. 2005, 6, 63–118. [Google Scholar] [CrossRef]
- Zenetos, A.; Meriç, E.; Verlaque, M.; Galli, P.; Boudouresque, C.F.; Giangrande, A.; Çinar, M.E.; Bilecenoglu, M. Additions to the annotated list of marine alien biota in the Mediterranean with special emphasis on Foraminifera and Parasites. Mediterr. Mar. Sci. 2008, 9, 119–166. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Gofas, S.; Verlaque, M.; Cinar, M.E.; Raso, J.E.G.; Bianchi, C.N.; Morri, C.; Azzurro, E.; Bilecenoglu, M.; Froglia, C.; et al. Alien species in the Mediterranean Sea by 2010. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part I. Spatial distribution. Mediterr. Mar. Sci. 2010, 11, 381. [Google Scholar] [CrossRef] [Green Version]
- Zenetos, A.; Gofas, S.; Morri, C.; Rosso, A.; Violanti, D.; Raso, J.G.; Çinar, M.E.; Almogi-Labin, A.; Ates, A.S.; Azzurro, E.; et al. Alien species in the Mediterranean Sea by 2012. A contribution to the application of European Union’s Marine Strategy Framework Directive (MSFD). Part 2. Introduction trends and pathways. Mediterr. Mar. Sci. 2012, 13, 328–352. [Google Scholar] [CrossRef] [Green Version]
- Galil, B.S. Taking stock: Inventory of alien species in the Mediterranean sea. Biol. Invasions 2009, 11, 359–372. [Google Scholar] [CrossRef]
- Galil, B.S. A sea under siege–alien species in the Mediterranean. Biol. Invasions 2000, 2, 177–186. [Google Scholar] [CrossRef]
- Bianchi, C.N.; Morri, C.; Chiantore, M.; Montefalcone, M.; Parravicini, V.; Rovere, A. Mediterranean Sea biodiversity between the legacy from the past and a future of change. In Life in the Mediterranean Sea: A Look at Habitat Changes; Nova Science Publishers: Hauppauge, NY, USA, 2012; Volume 1, p. 55. [Google Scholar]
- Ameer, A.; Olof, L. Maritime Traffic Effects on Biodiversity in the Mediterranean Sea: Review of Impacts, Priority Areas and Mitigation Measures, Malaga, Spain. Ph.D. Thesis, IUCN Centre for Mediterranean Cooperation, Gland, Switzerland, 2008; p. 184. [Google Scholar]
- Ferrario, J.; Caronni, S.; Occhipinti-Ambrogi, A.; Marchini, A. Role of commercial harbours and recreational marinas in the spread of non-indigenous fouling species. Biofouling 2017, 33, 651–660. [Google Scholar] [CrossRef]
- Renda, W.; Amati, B.; Bogi, C.; Bonomolo, G.; Capua, D.; Dell’angelo, B.; Furfaro, G.; Giannuzzi-Savelli, R.; La Perna, R.; Nofroni, I.; et al. The new Checklist of the Italian Fauna: Marine Mollusca. Biogeogr. J. Integr. Biogeogr. 2022, 37, ucl005. [Google Scholar] [CrossRef]
- Sabelli, B.; Taviani, M. The making of the Mediterranean molluscan biodiversity. In The Mediterranean Sea; Springer: Dordrecht, The Netherlands, 2014; pp. 285–306. [Google Scholar]
- Zenetos, A.; Galanidi, M. Mediterranean non indigenous species at the start of the 2020s: Recent changes. Mar. Biodivers. Rec. 2020, 13, 10. [Google Scholar] [CrossRef]
- Crocetta, F.; Macali, A.; Furfaro, G.; Cooke, S.; Villani, G.; Valdes, A. Alien molluscan species established along the Italian shores: An update, with discussions on some Mediterranean “alien species” categories. ZooKeys 2013, 277, 91. [Google Scholar] [CrossRef] [Green Version]
- Vitale, D.; Giacobbe, S.; Spinelli, A.; De Matteo, S.; Cervera, J.L. “Opisthobranch” (mollusks) inventory of the Faro Lake: A Sicilian biodiversity hot spot. Ital. J. Zool. 2016, 83, 524–530. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; De Matteo, S.; Mariottini, P.; Giacobbe, S. Ecological notes of the alien species Godiva quadricolor (Gastropoda, Nudibranchia) occurring in Faro Lake (Italy). J. Nat. Hist. 2018, 52, 645–657. [Google Scholar] [CrossRef]
- Azzola, A.; Furfaro, G.; Trainito, E.; Doneddu, M.; Montefalcone, M. Sea water warming favours the northward range extension of Lessepsian species in the Mediterranean Sea: A further example with the cephalaspidean Lamprohaminoea ovalis. JMBA 2022, 102, 1–7. [Google Scholar]
- Mioni, E.; Furfaro, G.; Merlino, S.; Mannino, A.M. Project “Percorsi nel Blu”: Ocean Literacy & Citizen Science as a valid tool to monitor Marine Heterobranchia. The case study of the first records of the alien species Favorinus ghanensis and the Mediterranean Okenia cf. longiductis, as updating records for the Italian coast. In Proceedings of the EMSEA Virtual Conference 2021, Gdynia, Poland, 7–8 October 2021. [Google Scholar]
- Mioni, E. “Percorsi nel Blu” (“Blue Paths”): A long-lasting project to integrate Ocean Literacy and Marine Citizen Science into school curricula. Mediterr. Mar. Sci. 2022, 23, 405–416. [Google Scholar] [CrossRef]
- Trainito, E.; Doneddu, M.; Furfaro, G. Aliens in changing seascapes: A newly reported non-native sacoglossan (Mollusca, Heterobranchia) in the western Mediterranean Sea. Check List 2022, 18, 545–551. [Google Scholar] [CrossRef]
- Canessa, M.; Bavestrello, G.; Cattaneo-Vietti, R.; Furfaro, G.; Doneddu, M.; Navone, A.; Trainito, E. Rocky substrate affects benthic heterobranch assemblages and prey/predator relationships. Estuar. Coast. Shelf Sci. 2021, 261, 107568. [Google Scholar] [CrossRef]
- Cervera, J.L.; Tamsouri, N.; Moukrim, A.; Villani, G. New records of two alien opisthobranch molluscs from the north-eastern Atlantic: Polycera hedgpethi and Godiva quadricolor. Mar. Biodivers. Rec. 2010, 3, e51. [Google Scholar] [CrossRef]
- Tamsouri, N.; Carmona, L.; Moukrim, A.; Cervera, J.L. Polycerella emertoni and Favorinus ghanensis: Two new alien sea slug molluscs from the Moroccan Atlantic coasts. Mar. Biodivers. Rec. 2014, 7, e13. [Google Scholar] [CrossRef]
- Verlaque, M. Checklist of the macroalgae of Thau Lagoon (Hérault, France), a hot spot of marine species introduction in Europe. Oceanol. Acta 2001, 24, 29–49. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Soto, C.; van der Meeren, G.I.; Busch, J.A.; Delany, J.; Domegan, C.; Dubsky, K.; Gorsky, G.; von Juterzenka, K.; Malfatti, F.; Mannaerts, G.; et al. Advancing Citizen Science for Coastal and Ocean Research; Position Paper 23; French, V., Kellett, P., Delany, J., McDonough, N., Eds.; European Marine Board, European Science Foundation: Ostend, Belgium, 2017; p. 112. [Google Scholar]
- Mioni, E.; Merlino, S.; Giovacchini, A. Engaging way to help students develop skills, interest and methodological research approaches in marine and environmental science. In Advances in Higher Education; de la Poza, E., Domènech, J., Lloret, J., Vincent-Vela, M.C., Zuriaga, E., Eds.; Editorial Universitat Politècnica de València: València, Spain, 2016; pp. 400–408. [Google Scholar]
- Mioni, E.; Mannino, A.M.; Merlino, S. First record of Aplysia dactylomela (Rang, 1828) (Heterobranchia, Aplysiidae) from the island of Pianosa (Alto Tirreno). In Proceedings of the Conference of the Italian Society of Marine Biology SIBM, Cesenatico, Italy, 4–8 June 2018; p. 224. [Google Scholar]
- Mioni, E.; Merlino, S.; Balistreri, P.; Mannino, A.M. First record of the invasive crab Percnon gibbesi (H. Milne Edwards, 1853) at Pianosa Island: The second goal reached by the innovative Marine Citizen Science Literacy Project “PERCORSI NEL BLU” (“BLUE PATHS”). In Proceedings of the 11th International Conference on Biological Invasions, Vodice, Croatia, 15–18 September 2020; p. 150. [Google Scholar]
- Mioni, E.; Stroobant, M.; Locritani, M.; Merlino, S.; Traverso, R. Blue Paths: 5 years motivating the participation of citizens in research activities for coastal monitoring of the Benthos. In Proceedings of the First Citizen Science Conference, Rome, Italy, 23–25 November 2017; p. 64. [Google Scholar]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Furfaro, G.; Schreier, C.; Trainito, E.; Pontes, M.; Madrenas, E.; Girard, P.; Mariottini, P. The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity. Diversity 2022, 14, 297. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.; Romano, S.; Mc Millan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Department of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 2002. [Google Scholar]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 SnRNA DNA Sequences and Arthropod Molecular Evolution. Aust. J. Zool. 1999, 46, 419–437. [Google Scholar] [CrossRef]
- Furfaro, G.; Oliverio, M.; Mariottini, P. The southernmost record of Felimida elegantula (Philippi, 1844) (Gastropoda: Nudibranchia). Mar. Biodivers. 2017, 47, 579–584. [Google Scholar] [CrossRef]
- Staden, R.; Beal, K.F.; Bonfield, J.K. The Staden package, 1998. Methods Mol. Biol. 2000, 132, 115–130. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Furfaro, G.; Mariottini, P. A New Dondice Marcus Er. 1958 (Gastropoda: Nudibranchia) from the Mediterranean Sea Reveals Interesting Insights into the Phylogenetic History of a Group of Facelinidae Taxa. Zootaxa 2020, 4731, 1–22. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Trainito, E.; Vitale, F.; Mariottini, P. When Morphology Does Not Match Phylogeny: The Puzzling Case of Two Sibling Nudibranchs (Gastropoda). Zool. Scr. 2021, 50, 439–454. [Google Scholar] [CrossRef]
- Furfaro, G.; Salvi, D.; Mancini, E.; Mariottini, P. A multilocus view on Mediterranean aeolid nudibranchs (Mollusca): Systematics and cryptic diversity of Flabellinidae and Piseinotecidae. Mol. Phylogenetic Evol. 2018, 118, 13–22. [Google Scholar] [CrossRef]
- Galià-Camps, C.A.R.L.E.S.; Cervera, J.L.; Valdés, Á.; Ballesteros, M. Attack on crypsis: Molecular and morphological study of Dendrodoris Ehrenberg, 1831 (Mollusca: Gastropoda: Nudibranchia) from the Mediterranean Sea and Northern Atlantic Ocean reinstates Dendrodoris temarana Pruvot-Fol, 1953. Zootaxa 2022, 5133, 383–406. [Google Scholar] [CrossRef]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; Boisselier, M.-C.; Cruaud, C.; Holford, M.; Samadi, S. Large-Scale Species Delimitation Method for Hyperdiverse Groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silvestro, D.; Michalak, I. RaxmlGUI: A Graphical Front-End for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Edmunds, M. Opisthobranchiate Mollusca from Ghana. J. Molluscan Stud. 1968, 38, 83–100. [Google Scholar] [CrossRef]
- Ben Soussi, J.; Zaouali, J.; Rezig, M.; Bradai, M.N.; Quignard, J.P.; Rudman, B. Contribution à l’étude de quelques récentes migrations d’espèces exotiques dans les eaux Tunisiennes. Rapp. Comm. Int. Pour L’exploration Sci. Mer Mediterr. 2004, 37, 312. [Google Scholar]
- Verrill, A.E. Notice of recent addition to the marine Invertebrata of the northeastern coast of America, with descriptions of new genera and species and critical remarks on others. Part II—Mollusca, with notes on Annelida, Echinodermata, etc., collected by the United States Fish Commission. Proc. U. S. Natl. Mus. 1880, 3, 356–409. [Google Scholar]
- Valdés, A.; Hamann, J.; Behrens, D.W.; Dupont, A. Caribbean Sea Slugs: A Field Guide to the Opisthobranch Mollusks from the Tropical Northwestern Atlantic; Sea Challengers Natural History Books: Gig Harbor, WA, USA, 2006. [Google Scholar]
- Edmunds, M. Larval development, oceanic currents, and origins of the opisthobranch fauna of Ghana. J. Molluscan Stud. 1977, 43, 301–308. [Google Scholar]
- García, J.C.; Bobo, A. Un nuevo Doridaceo para el litoral Ibérico: Polycerella emertoni Verrill (1880) 1881 (Gastropoda: Nudibranchia). Boll. Malacol. 1986, 22, 49–56. [Google Scholar]
- Cervera, J.L.; García-Gómez, J.C.; Toscano, F.; García, F.J. Polycera hedgpethi Marcus, 1964 (Gastropoda: Nudibranchia) an Indo-Pacific species discovered in the Mediterranean Sea. Iberus 1988, 8, 225–231. [Google Scholar]
- Cervera, J.L.; García-Gómez, J.C.; Sánchez-Tocino, L. Familia Polyceridae. In Moluscos Marinos de Andalucía; Gofas, S., Moreno, D., Salas, C., Eds.; Servicio de Publicaciones e Intercambio Científico, Universidad de Málaga: Málaga, Spain, 2011; pp. 467–471. [Google Scholar]
- García-Gómez, J.C.; Cervera, J.L.; García, F.J.; Ortea, J.A.; Garcia-Martin, S.F.; Medina, A.; Burnay, L.P. Resultados de la Campana Internacional de biología marina ‘Algarve-88’: Moluscos Opistobranquios. Boll. Malacol. 1991, 27, 125–138. [Google Scholar]
- Megina, C.; Cervera, J.L. Diet, prey selection and cannibalism in the hunter opistobranch Roboastra europaea. J. Mar. Biol. Assoc. UK 2003, 83, 489–495. [Google Scholar] [CrossRef]
- Camps-Castellà, J.; Ballesteros, M.; Trobajo, R.; Pontes, M.; Prado, P. Not all nudibranchs are carnivorous: Trophic ecology of Polycerella emertoni in the Ebro Delta. Mar. Ecol. Prog. Ser. 2020, 645, 67–82. [Google Scholar] [CrossRef]
- Schmekel, L. Die gattung Polycerella Verrill im Mittelmeer (Gastrp. Opisthobranchia). Pubbl. Della Stn. Zool. Napoli 1965, 34, 226–234. [Google Scholar]
- Cattaneo-Vietti, R.; Chemello, R.; Giannuzzi-Savelli, R. Atlas of Mediterranean Nudibranchs; La Conchiglia: Rome, Italy, 1990. [Google Scholar]
- Sammut, C.R.; Perroneù, A.S. A preliminary check-list of Opisthobranchia (Mollusca, Gastropoda) from the Maltese Islands. Basteria 1998, 62, 221–240. [Google Scholar]
- Sciberras, M.; Schembri, P.J. A critical review of records of alien marine species from the Maltese Islands and surrounding waters (Central Mediterranean). Mediterr. Mar. Sci. 2007, 8, 41–66. [Google Scholar] [CrossRef] [Green Version]
- Koutsoubas, D.; Arvanitidis, C.; Dounas, C.; Drummond, L. Community structure and dynamics of the molluscan fauna in a Mediterranean lagoon (Gialova Lagoon, SW Greece). Belg. J. Zool. 2000, 130, 135–142. [Google Scholar]
- Antit, M.; Gofas, S.; Salas, C.; Azzouna, A. One hundred years after Pinctada: An update on alien Mollusca in Tunisia. Mediterr. Mar. Sci. 2011, 12, 53–74. [Google Scholar] [CrossRef] [Green Version]
- West, N.P. Systematics and phylogeny of Favorinus, a clade of specialized predatory nudibranchs. In Collection: 2012 Biology: Concentration in Integrative Biology Masters Theses 2012; San Francisco State University: San Francisco, CA, USA, 2012; Available online: http://hdl.handle.net/10211.3/125190 (accessed on 1 August 2022).
- Korshunova, T.; Picton, B.; Furfaro, G.; Mariottini, P.; Pontes, M.; Prkić, J.; Fletcher, K.; Malmberg, K.; Lundin, K.; Martynov, A. Multilevel fine-scale diversity challenges the ‘cryptic species’ concept. Sci. Rep. 2019, 9, 6732. [Google Scholar] [CrossRef]
- Pola, M.; Paz-Sedano, S.; Macali, A.; Minchin, D.; Marchini, A.; Vitale, F.; Licchelli, C.; Crocetta, F. What is really out there? Review of the genus Okenia Menke, 1830 (Nudibranchia: Goniodorididae) in the Mediterranean Sea with description of two new species. PLoS ONE 2019, 14, e0215037. [Google Scholar] [CrossRef] [Green Version]
- Marchini, A.; Ferrario, J.; Minchin, J.D. Marinas may act as hubs for the spread of the pseudo-indigenous bryozoan Amathia verticillata (Delle Chiaje, 1822) and its associates. Sci. Mar. 2015, 79, 355–365. [Google Scholar] [CrossRef] [Green Version]
- Dailianis, T.; Akyol, O.K.A.N.; Babali, N.; Bariche, M.; Crocetta, F.; Gerovasileiou, V.; Ghanem, R.; Gökoğlu, M.; Hasiotis, T.; Izquierdo-Munoz, A.; et al. New Mediterranean biodiversity records (July 2016). Mediterr. Mar. Sci. 2016, 17, 608–626. [Google Scholar] [CrossRef] [Green Version]
- Olenin, S.; Leppäkoski, E. Non-native animals in the Baltic Sea: Alteration of benthic habitats in coastal inlets and lagoons. Hydrobiologia 1999, 393, 233–243. [Google Scholar] [CrossRef]
- Galil, B.S.; Gevili, R. Zoobotryon verticillatum (Bryozoa: Ctenostomatida: Vesiculariidae), a new occurrence on the Mediterranean coast of Israel. Mar. Biodivers. Rec. 2014, 7, e17. [Google Scholar] [CrossRef]
- Carlton, J.T. Biological invasions and cryptogenic species. Ecology 1996, 77, 1653–1655. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species Name | Voucher | COI | 16S | H3 |
|---|---|---|---|---|
| Favorinus ghanensis | RM3_2290 | OP223449 | OP221319 | OP251348 |
| RM3_2291 | OP223450 | OP221320 | OP251349 | |
| RM3_2292 | OP223451 | OP221321 | OP251350 | |
| RM3_2293 | -------- | -------- | -------- | |
| Polycerella emertoni | RM3_2283 | -------- | -------- | -------- |
| RM3_2284 | -------- | -------- | -------- | |
| RM3_2285 | OP218058 | OP218055 | OP251346 | |
| RM3_2286 | OP218059 | OP218056 | OP251347 | |
| RM3_2287 | -------- | -------- | -------- | |
| RM3_2288 | -------- | -------- | -------- | |
| RM3_2289 | -------- | -------- | -------- |
| Species Name | Locality | Voucher | COI |
|---|---|---|---|
| Favorinus auranoralis | Pacific Ocean: Marshall Islands, Kwajalein, Atoll, Loi Island, South Loi sandspit | CASIZ 181353 | JX220463 |
| Philippines: Luzon, Batangas Province, Maricaban Island, Bethlehem | CASIZ 182795 | JX220462 | |
| Favorinus auritulus | Atlantic Ocean: North Atlantic Ocean, Bermuda, Hamilton Parish, Shelly Bay | CASIZ 181172 | JX220476 |
| Atlantic Ocean: North Atlantic Ocean, Bermuda, Hamilton Parish, Walsingham Bay | CASIZ 181196 | JX220475 | |
| Atlantic Ocean: North Atlantic Ocean, Bermuda, Hamilton Parish, Walsingham Bay | CASIZ 181202 | JX220474 | |
| Atlantic Ocean: North Atlantic Ocean, Bermuda, Hamilton Parish, Walsingham Bay | CASIZ 181203 | JX220473 | |
| --------------------------------- | SRR1950950 | KX889733 | |
| Favorinus blianus | United Kingdom: Scotland, Outer Hebrides, St. Kilda, Hirta, Village Bay | CASIZ 118892 | JX220472 |
| Favorinus branchialis | Italy: Latium, Sabaudia, Lago di Paola | BAU2676 | LT596562 |
| Spain: Cadiz | MNCN15.05/53695 | HQ616761 | |
| Favorinus bullitis | Pacific Ocean: Marshall Islands, Kwajalein, Atoll, Gugeegue Island, Gugeegue sandspit | CASIZ 181361 | JX220461 |
| Pacific Ocean: Marshall Islands, Kwajalein, Atoll, Little Shell sandspit | CASIZ 185769A | JX220460 | |
| Pacific Ocean: Marshall Islands, Kwajalein, Atoll, Little Shell sandspit | CASIZ 185769B | JX220459 | |
| Pacific Ocean: Marshall Islands, Kwajalein, Atoll, Little Shell sandspit | CASIZ 185769C | JX220458 | |
| Philippines: Luzon, Batangas Province, Calumpan, Peninsula, Balayan Bay | CASIZ 186045 | JX220457 | |
| Favorinus elenalexiarum | Costa Rica: Guanacaste, Isla Plata | CASIZ178875 | HM162755 |
| Mexico: Jalisco, Bahia de Banderas, Puerto Vallarta, Marietas and Los Arcos | CASIZ 174060A | JX220471 | |
| Favorinus fuscunaris | Philippines: Luzon, Batangas Province, Balayan, Bay, Anilao, Anilao Harbor | CASIZ 186046 | JX220455 |
| Favorinus ghanensis | Italy: Liguria, La Spezia, Fezzano’s marina | RM3_2290 | OP223449 |
| Italy: Liguria, La Spezia, Fezzano’s marina | RM3_2291 | OP223450 | |
| Italy: Liguria, La Spezia, Fezzano’s marina | RM3_2292 | OP223451 | |
| Favorinus inflatus | Philippines: Luzon, Batangas Province, Balayan, Bay, Anilao, Anilao Harbor | CASIZ 186438 | JX220456 |
| Favorinus japonicus | Philippines: Luzon, Batangas Province, Maricaban Island, Devil’s Point | CASIZ 177286 | JX220470 |
| USA: Hawaii, Maui, Airport Beach | CASIZ 180309 | JX220469 | |
| Philippines: Sibuyan Sea, Romblon Province, Tablas Island, north tip of Tablas Island | CASIZ 182291 | JX220468 | |
| Favorinus mirabilis | Philippines: Luzon, Batangas Province, Calumpan Peninsula, Maricaban Strait, Twin Rocks | CASIZ 177474 | JX220467 |
| Philippines: Luzon, Batangas Province, Maricaban Island, Devil’s Point | CASIZ 177672A | JX220466 | |
| Pacific Ocean: Marshall Islands, Kwajalein Atoll, Kwajalein Harbor | CASIZ 181347A | JX220465 | |
| Pacific Ocean: Marshall Islands, Kwajalein Atoll, North Loi Island | CASIZ 182194A | JX220464 | |
| Favorinussp. | Indonesia | Fasp_17Bu-1 | MK514329 |
| Favorinussp. | Indonesia | Fasp_15Bu-1 | MK514328 |
| Favorinussp. | French Polynesia: Moorea | PW-2014 | KJ522462 |
| Favorinus tsuruganus | Philippines: Luzon, Batangas Province, Maricaban Island, Bethlehem | CASIZ 177373 | JX220454 |
| Philippines: Luzon, Batangas Province, Maricaban Island, Bethlehem | CASIZ 177374 | JX220453 | |
| Philippines: Luzon, Batangas Province, Balayan Bay, east of Dive’n Trek | CASIZ 177652 | JX220452 | |
| Philippines: Sibuyan Sea, Romblon Province, Romblon Island, Bon Bon Beach | CASIZ 182284 | JX220451 | |
| Philippines: Luzon, Batangas Province, Maricaban Island, Caban Island, Kirby’s Rock | CASIZ 186044 | JX220450 | |
| Coryphella verrucosa | WS14499 | MZ832555 | |
| ZMBN106122 | MZ832557 | ||
| Flabellina affinis | Italy: Latium, Ponza Is. | BAU2806 | LT718555 |
| Italy: Latium, Ponza Is. | BAU2807 | LT718556 | |
| Greece: Lefkada | BAU2808 | LT718557 | |
| Spain: Cadiz | BAU2809 | LT718558 | |
| Flabellina cavolini | Italy: Sardinia, Tavolara | RM3_1390 | LR813668 |
| Italy: Liguria, La Spezia, Scoglio Ferale | RM3_1560 | LR813669 | |
| Italy: Liguria, Genova, Portofino, Il faro | RM3_1751 | LR813670 | |
| Flabellina gaditana | Italy: Apulia, Tremiti Islands | RM3_461 | LR813671 |
| Italy: Sardinia, Tavolara Island | RM3_783 | LR813672 | |
| France: Arcachon | MCNCN/ADN51998 | JX087557 | |
| Duvaucelia striata * | Italy: Tuscany, Giannutri Is. | BAU2695 | LT596540 |
| Italy: Grosseto, Le Formiche Is. | BAU2696 | LT596541 | |
| Palio dubia | USA: Maine, Portland | CASIZ 182030 | MZ382786 |
| Russia:White Sea | ZMMU:Op-751 | MZ425304 | |
| Canada: Quebec, Baie Ste-Marguerite, Trait 7 | CCDB-15498-E04 | KF644300 | |
| Canada: Quebec, Baie Ste-Marguerite, Trait 7 | CCDB-15498-E07 | KF643719 | |
| Canada: Quebec, Baie Ste-Marguerite, Trait 7 | CCDB-15498-E06 | KF643686 | |
| Sweden:Kristineberg, Bohuslän | MNCN:15.05/55467 | JX274100 | |
| Sweden:Kristineberg, Bohuslän | -------------- | AJ223272 | |
| Polycera atra | USA: California, San Francisco Bay, USA, San Francisco Marina | CASIZ 170506b | JX274085 |
| USA: California, San Francisco Bay, USA, San Francisco Marina | CASIZ 170506a | JX274084 | |
| Polycera quadrilineata | Sweden: Gothenborg, Tjarno | MNCN:15.05/55466 | JX274073 |
| Sweden: Gothenborg, Tjarno | MNCN:15.05/55463 | JX274074 | |
| Polycerella emertoni | Italy: Liguria, La Spezia, Fezzano’s marina | RM3_2285 | OP218058 |
| Italy: Liguria, La Spezia, Fezzano’s marina | RM3_2286 | OP218059 | |
| Spain: Cadiz, Santi Petri, Pantalan | MNCN:15.05/55482 | JX274099 | |
| Spain: Cadiz, Santi Petri, Pantalan | MNCN:15.05/55482 | JX274098 | |
| Spain: Cadiz, Santi Petri, Pantalan | MNCN:15.05/55479.2 | JX274097 | |
| Spain: Cadiz, Santi Petri, Pantalan | MNCN:15.05/55479.1 | JX274096 | |
| Spain: Cadiz, Santi Petri | MNCN:15.05/55480 | JX274095 | |
| Spain: Cadiz, Andalusia | -------------- | AJ223273 | |
| Thecacera pennigera * | Spain: Cadiz, Andalusia | -------------- | AJ223277 |
| South Africa: Cape Province, Atlantic Coast, Oudekraal | CASIZ 176285 | JX274094 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mioni, E.; Furfaro, G. Alien Travel Companies: The Case of Two Sea Slugs and One Bryozoan in the Mediterranean Sea. Diversity 2022, 14, 687. https://doi.org/10.3390/d14080687
Mioni E, Furfaro G. Alien Travel Companies: The Case of Two Sea Slugs and One Bryozoan in the Mediterranean Sea. Diversity. 2022; 14(8):687. https://doi.org/10.3390/d14080687
Chicago/Turabian StyleMioni, Erika, and Giulia Furfaro. 2022. "Alien Travel Companies: The Case of Two Sea Slugs and One Bryozoan in the Mediterranean Sea" Diversity 14, no. 8: 687. https://doi.org/10.3390/d14080687
APA StyleMioni, E., & Furfaro, G. (2022). Alien Travel Companies: The Case of Two Sea Slugs and One Bryozoan in the Mediterranean Sea. Diversity, 14(8), 687. https://doi.org/10.3390/d14080687