

Mollusc Crystallins: Physical and Chemical Properties and Phylogenetic Analysis


Abstract
:
1. Introduction
2. Materials and Methods
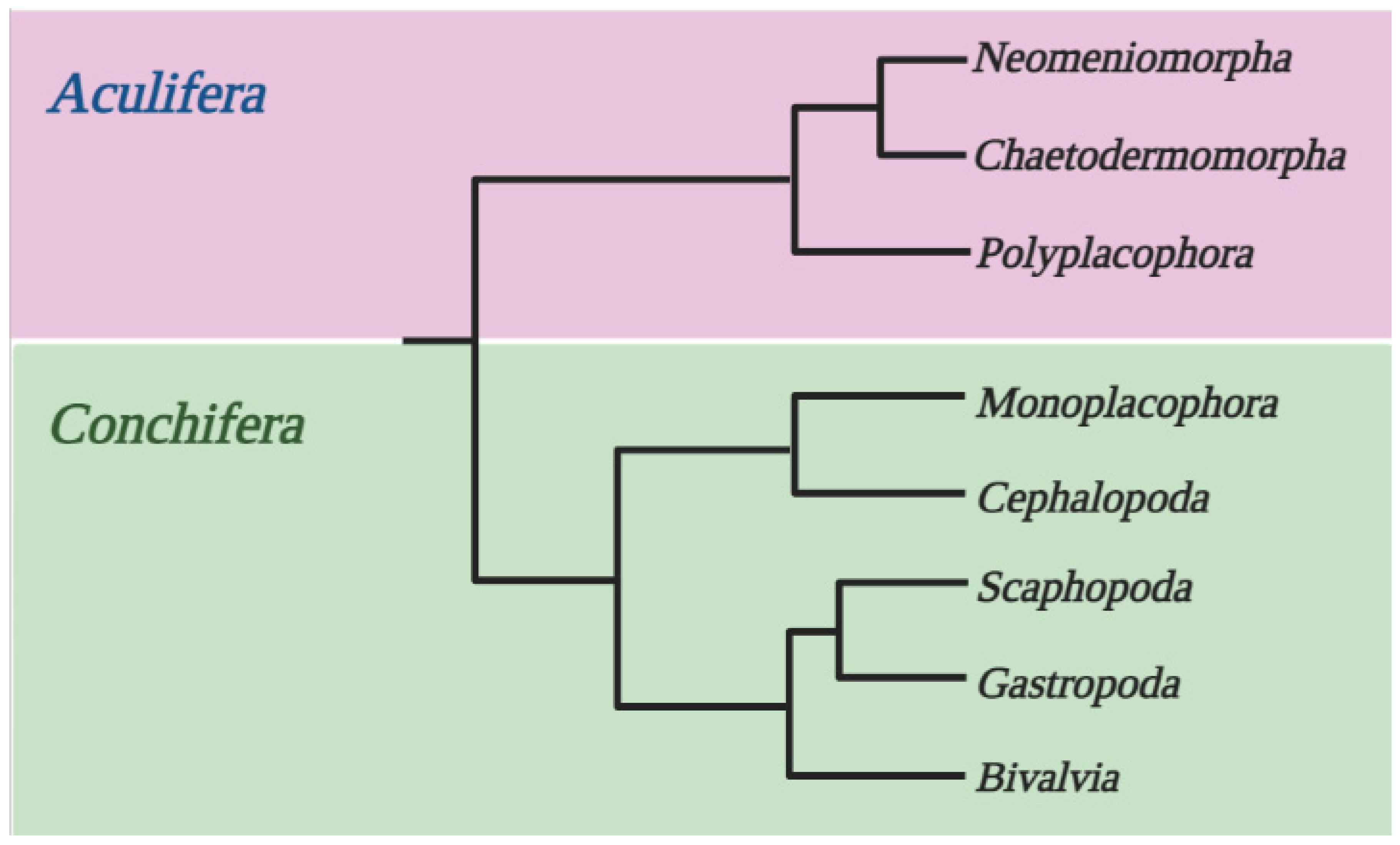
2.1. Studied Genomes and Crystallin Sequences
2.2. Analysis of Several Physical and Chemical Properties and Calculation of the Refractive Index Increment of Crystallins
2.3. Multiple Sequence Alignment and Phylogenetic Analysis
3. Results
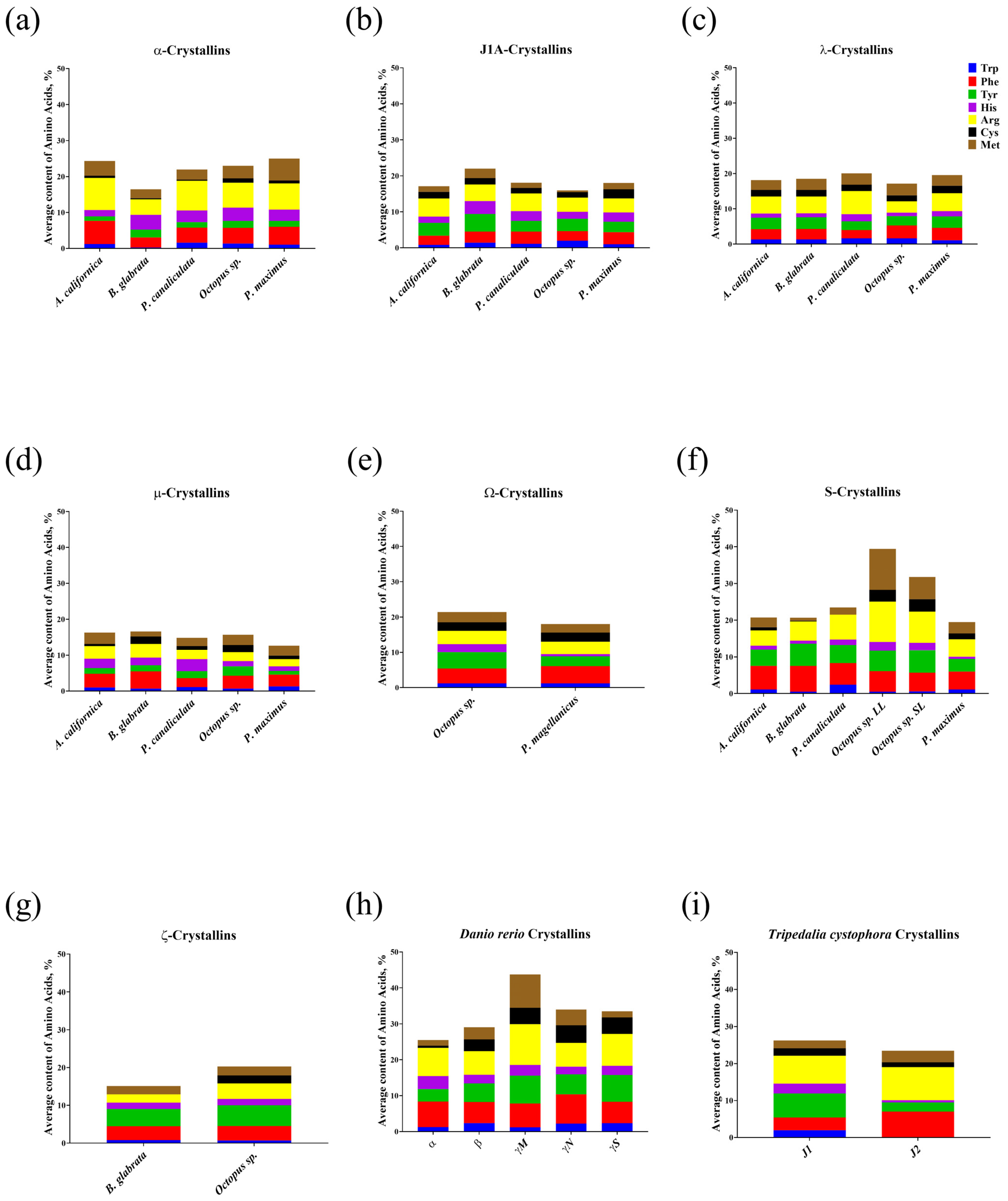
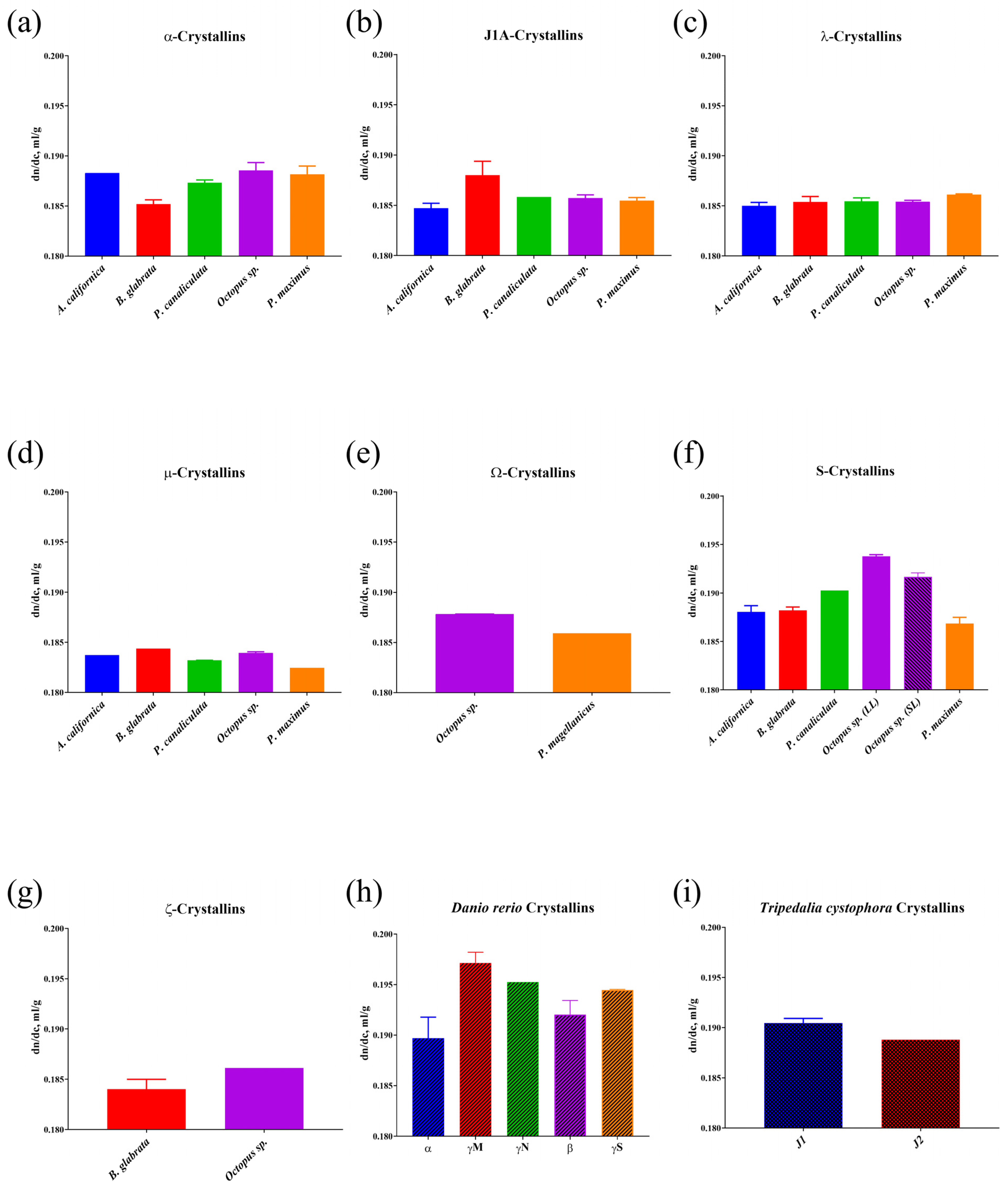
3.1. Analysis of the Amino Acid Composition and Calculation of the Refractive Index Increment of Crystallins
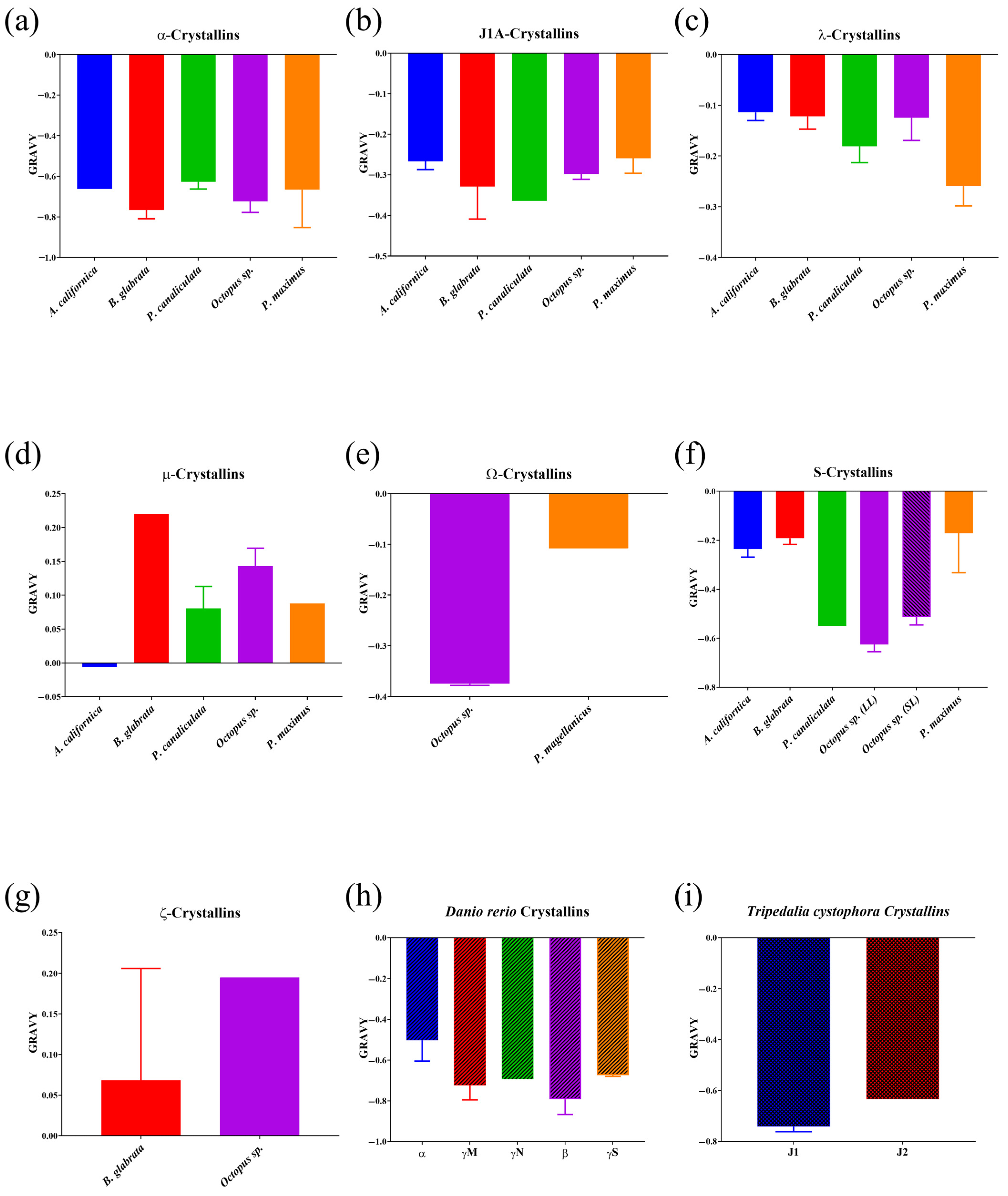
3.2. Crystalline Hydrophobicity Analysis
3.3. The Secondary Structure Analysis of Crystallins
3.4. Multiple Crystallin Sequence Alignment Analysis
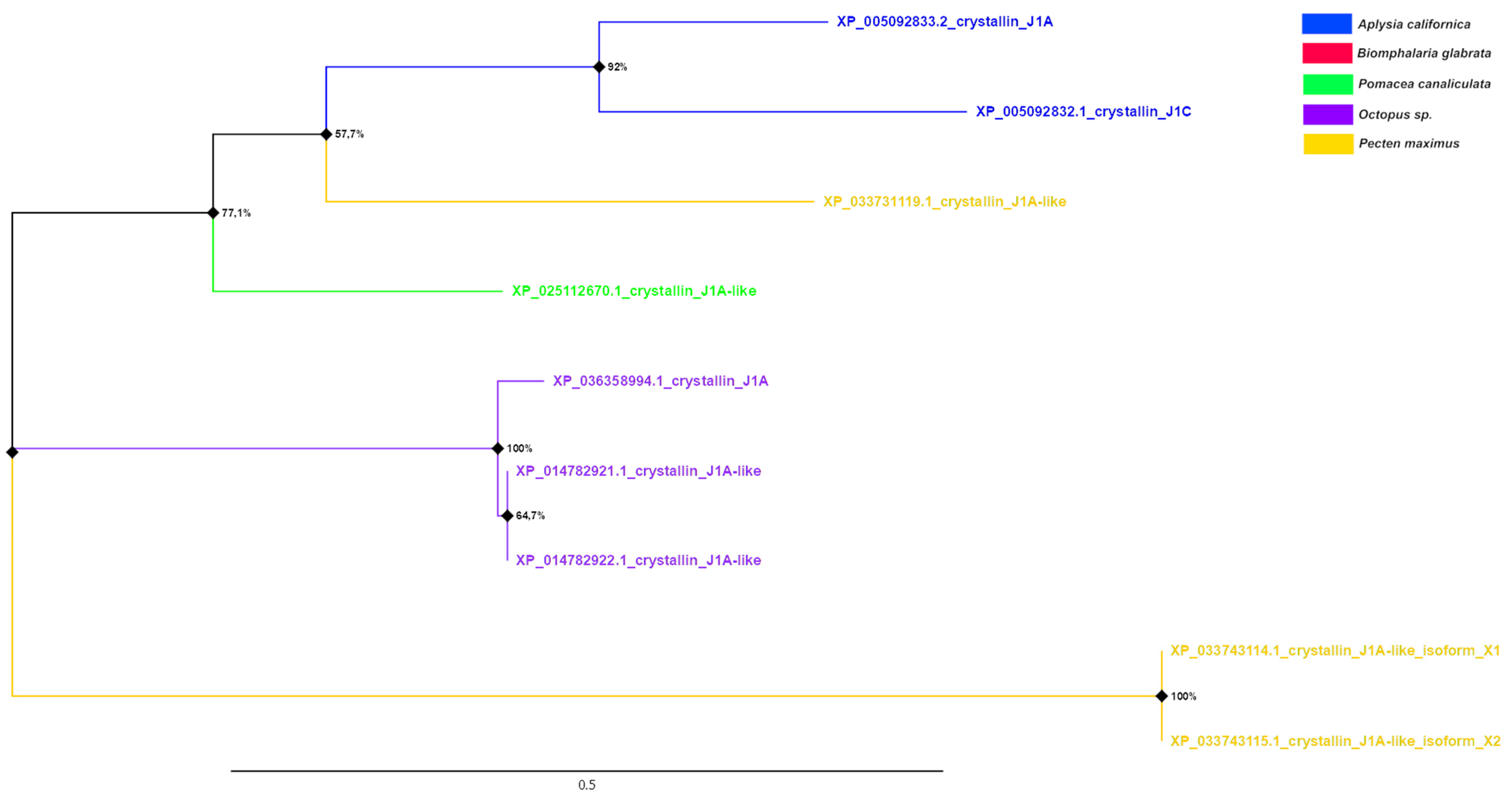
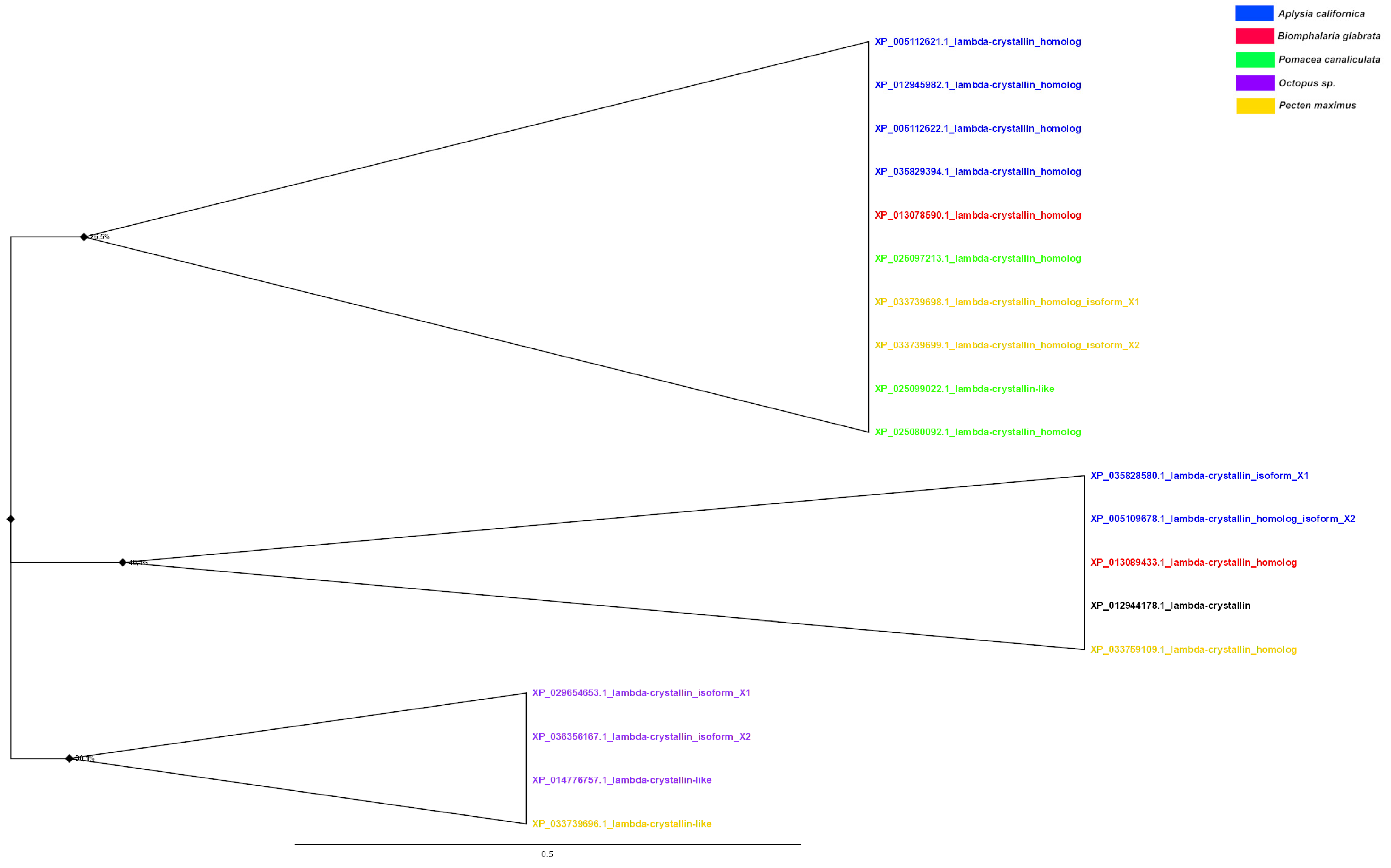
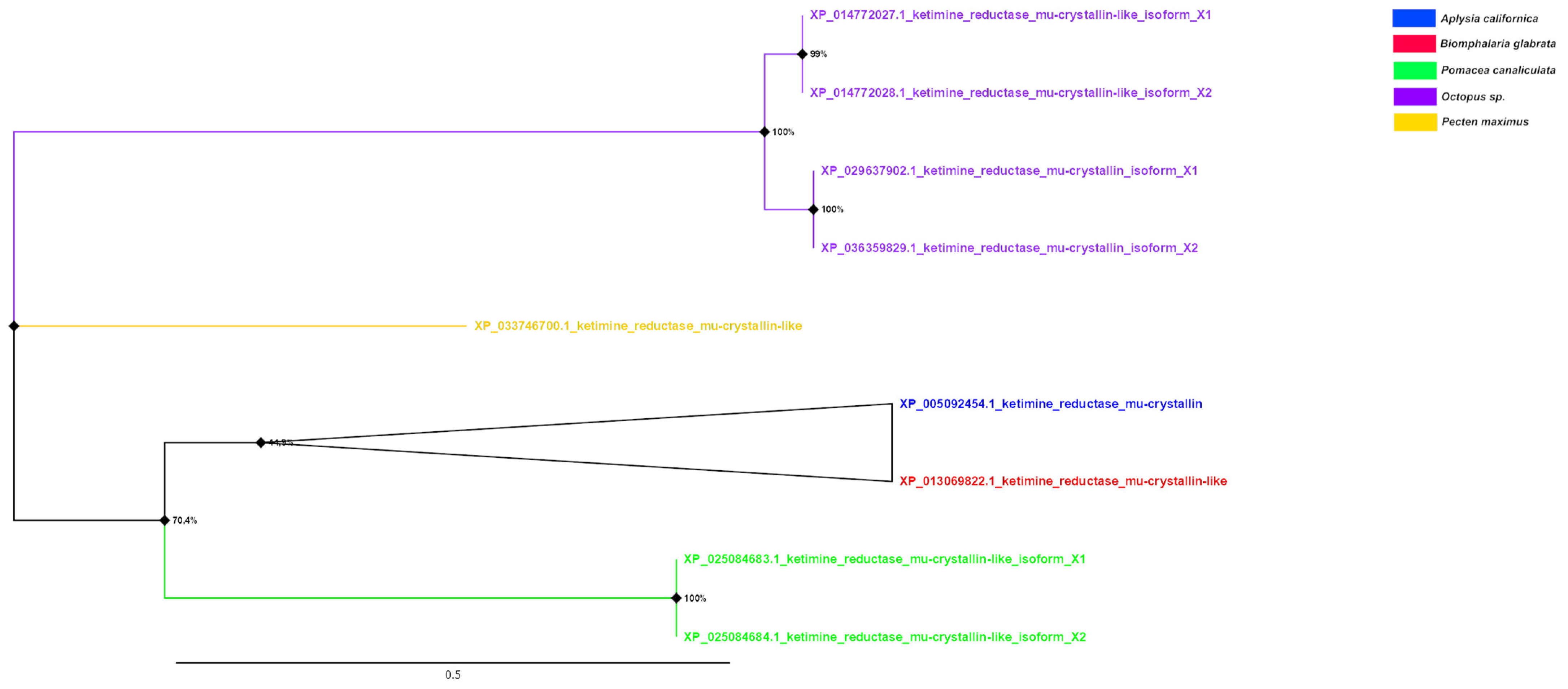
3.5. Phylogenetic Analysis of Crystallins
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wistow, G. The Human Crystallin Gene Families. Hum. Genom. 2012, 6, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Jong, W.W.; Hendriks, W.; Mulders, J.W.M.; Bloemendal, H. Evolution of Eye Lens Crystallins: The Stress Connection. Trends Biochem. Sci. 1989, 14, 365–368. [Google Scholar] [CrossRef]
- Piatigorsky, J. Lens crystallins. Innovation Associated with Changes in Gene Regulation. J. Biol. Chem. 1992, 267, 4277–4280. [Google Scholar] [CrossRef]
- Piatigorsky, J. Evolution of Mollusc Lens Crystalline: Glutathione S-Transferase/S-Crystallins and Aldehyde Dehydrogenase/Ω-Crystallins. Am. Malacol. Bull. 2008, 26, 73–81. [Google Scholar] [CrossRef]
- Chuang, C.C.; Wu, S.H.; Chiou, S.H.; Chang, G.G. Homology Modeling of Cephalopod Lens S-Crystallin: A Natural Mutant of Sigma-Class Glutathione Transferase with Diminished Endogenous Activity. Biophys. J. 1999, 76, 679–690. [Google Scholar] [CrossRef] [Green Version]
- Tomarev, S.I.; Chung, S.; Piatigorsky, J. Glutathione S-Transferase and S-Crystallins of Cephalopods: Evolution from Active Enzyme to Lens-Refractive Proteins. J. Mol. Evol. 1995, 41, 1048–1056. [Google Scholar] [CrossRef]
- Yoshida, M.A.; Ogura, A.; Ikeo, K.; Shigeno, S.; Moritaki, T.; Winters, G.C.; Kohn, A.B.; Moroz, L.L. Molecular Evidence for Convergence and Parallelism in Evolution of Complex Brains of Cephalopod Molluscs: Insights from Visual Systems. Integr. Comp. Biol. 2015, 55, 1070–1083. [Google Scholar] [CrossRef] [Green Version]
- Cvekl, A.; Zhao, Y.; McGreal, R.; Xie, Q.; Gu, X.; Zheng, D. Evolutionary Origins of Pax6 Control of Crystallin Genes. Genome Biol. Evol. 2017, 9, 2075. [Google Scholar] [CrossRef] [Green Version]
- Horwitz, J.; Ding, L.; Vasiliou, V.; Cantore, M.; Piatigorsky, J. Scallop Lens Ω-Crystallin (ALDH1A9): A Novel Tetrameric Aldehyde Dehydrogenase. Biochem. Biophys. Res. Commun. 2006, 348, 1302–1309. [Google Scholar] [CrossRef]
- Zinovieva, R.D.; Tomarev, S.I.; Piatigorsky, J. Aldehyde Dehydrogenase-Derived Omega-Crystallins of Squid and Octopus. Specialization for Lens Expression. J. Biol. Chem. 1993, 268, 11449–11455. [Google Scholar] [CrossRef]
- Montgomery, M.K.; McFall-Ngai, M.J. The Muscle-Derived Lens of a Squid Bioluminescent Organ Is Biochemically Convergent with the Ocular Lens. Evidence for Recruitment of Aldehyde Dehydrogenase as a Predominant Structural Protein. J. Biol. Chem. 1992, 267, 20999–21003. [Google Scholar] [CrossRef]
- Cox, R.L.; Glick, D.L.; Strumwasser, F. Isolation and Protein Sequence Identification of Aplysia Californica Lens Crystallins. Biol. Bull. 2016, 181, 333–335. [Google Scholar] [CrossRef]
- Tomarev, S.I.; Piatigorsky, J. Lens Crystallins of Invertebrates--Diversity and Recruitment from Detoxification Enzymes and Novel Proteins. Eur. J. Biochem. 1996, 235, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Wages, P.; Horwitz, J.; Ding, L.; Corbin, R.W.; Posner, M. Changes in Zebrafish (Danio Rerio) Lens Crystallin Content during Development. Mol. Vis. 2013, 19, 408. [Google Scholar] [PubMed]
- Piatigorsky, J.; Horwitz, J.; Norman, B.L. J1-Crystallins of the Cubomedusan Jellyfish Lens Constitute a Novel Family Encoded in at Least Three Intronless Genes. J. Biol. Chem. 1993, 268, 11894–11901. [Google Scholar] [CrossRef]
- Zhao, H.; Brown, P.H.; Schuck, P. On the Distribution of Protein Refractive Index Increments. Biophys. J. 2011, 100, 2309–2317. [Google Scholar] [CrossRef] [Green Version]
- Gasteiger, E.; Hoogland, C.; Gattiker, A.; Duvaud, S.; Wilkins, M.R.; Appel, R.D.; Bairoch, A. Protein Identification and Analysis Tools on the ExPASy Server. In The Proteomics Protocols Handbook; Springer: Totowa, NJ, USA, 2005; pp. 571–607. [Google Scholar] [CrossRef]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Magdeldin, S.; Yoshida, Y.; Li, H.; Maeda, Y.; Yokoyama, M.; Enany, S.; Zhang, Y.; Xu, B.; Fujinaka, H.; Yaoita, E.; et al. Murine Colon Proteome and Characterization of the Protein Pathways. BioData Min. 2012, 5, 11. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Li, W.; Liu, S.; Xu, J. RaptorX-Property: A Web Server for Protein Structure Property Prediction. Nucleic Acids Res. 2016, 44, W430–W435. [Google Scholar] [CrossRef]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; Varlamov, A.; Vaskin, Y.; Efremov, I.; German Grehov, O.G.; Kandrov, D.; Rasputin, K.; Syabro, M.; et al. Unipro UGENE: A Unified Bioinformatics Toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Sievers, F.; Higgins, D.G. Clustal Omega for Making Accurate Alignments of Many Protein Sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview Version 2—A Multiple Sequence Alignment Editor and Analysis Workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirny, L.A.; Shakhnovich, E.I. Universally Conserved Positions in Protein Folds: Reading Evolutionary Signals about Stability, Folding Kinetics and Function. J. Mol. Biol. 1999, 291, 177–196. [Google Scholar] [CrossRef] [Green Version]
- Valdar, W.S.J. Scoring Residue Conservation. Proteins 2002, 48, 227–241. [Google Scholar] [CrossRef]
- Taylor, W.R. The Classification of Amino Acid Conservation. J. Theor. Biol. 1986, 119, 205–218. [Google Scholar] [CrossRef]
- Procter, J.B.; Carstairs, G.M.; Soares, B.; Mourão, K.; Ofoegbu, T.C.; Barton, D.; Lui, L.; Menard, A.; Sherstnev, N.; Roldan-Martinez, D.; et al. Alignment of Biological Sequences with Jalview. Methods Mol. Biol. 2021, 2231, 203–224. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An Improved General Amino Acid Replacement Matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Felsenstein, J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution 1985, 39, 783. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 July 2022).
- Khago, D.; Bierma, J.C.; Roskamp, K.W.; Kozlyuk, N.; Martin, R.W. Protein Refractive Index Increment Is Determined by Conformation as Well as Composition. J. Phys. Condens. Matter 2018, 30, 435101. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahendiran, K.; Elie, C.; Nebel, J.C.; Ryan, A.; Pierscionek, B.K. Primary Sequence Contribution to the Optical Function of the Eye Lens. Sci. Rep. 2014, 4, 5195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kozmik, Z.; Swamynathan, S.K.; Ruzickova, J.; Jonasova, K.; Paces, V.; Vlcek, C.; Piatigorsky, J. Cubozoan Crystallins: Evidence for Convergent Evolution of Pax Regulatory Sequences. Evol. Dev. 2008, 10, 52–61. [Google Scholar] [CrossRef] [PubMed]
- Greiling, T.M.S.; Houck, S.A.; Clark, J.I. The Zebrafish Lens Proteome during Development and Aging. Mol. Vis. 2009, 15, 2313. [Google Scholar] [PubMed]
- Pierscionek, B.K.; Regini, J.W. The Gradient Index Lens of the Eye: An Opto-Biological Synchrony. Prog. Retin. Eye Res. 2012, 31, 332–349. [Google Scholar] [CrossRef]
- Cronin, T.W.; Johnsen, S.; Marshall, N.J.; Warrant, E.J. Visual Ecology; Princeton University Press: Princeton, NJ, USA, 2014; p. 428. [Google Scholar]
- Land, M.F.; Nilsson, D.-E. Animal Eyes; Oxford University Press: Oxford, UK, 2013. [Google Scholar] [CrossRef]
- Piatigorsky, J.; Kozmik, Z.; Horwitzi, J.; Ding, L.; Carosa, E.; Robison, W.G.; Steinbach, P.J.; Tamm, E.R. Omega-Crystallin of the Scallop Lens. A Dimeric Aldehyde Dehydrogenase Class 1/2 Enzyme-Crystallin. J. Biol. Chem. 2000, 275, 41064–41073. [Google Scholar] [CrossRef] [Green Version]
- Carosa, E.; Kozmik, Z.; Edward Rall, J.; Piatigorsky, J. Structure and Expression of the Scallop Omega-Crystallin Gene. Evidence for Convergent Evolution of Promoter Sequences. J. Biol. Chem. 2002, 277, 656–664. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, A.M.; des Marais, D.L.; Ban, Y.E.A.; Johnsen, S. Evolution of Graded Refractive Index in Squid Lenses. J. R. Soc. Interface 2007, 4, 685–698. [Google Scholar] [CrossRef]
- Cai, J.; Townsend, J.P.; Dodson, T.C.; Heiney, P.A.; Sweeney, A.M. Eye Patches: Protein Assembly of Index-Gradient Squid Lenses. Science 2017, 357, 564–569. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.H.; Cheng, S.C.; Liu, Y.T.; Wu, C.G.; Lin, M.H.; Chen, C.C.; Lin, C.H.; Chou, C.Y. Structure of a Highly Active Cephalopod S-Crystallin Mutant: New Molecular Evidence for Evolution from an Active Enzyme into Lens-Refractive Protein. Sci. Rep. 2016, 6, 31176. [Google Scholar] [CrossRef] [PubMed]
- Palmer, B.A.; Taylor, G.J.; Brumfeld, V.; Gur, D.; Shemesh, M.; Elad, N.; Osherov, A.; Oron, D.; Weiner, S.; Addadi, L. The Image-Forming Mirror in the Eye of the Scallop. Science 2017, 358, 1172–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, K.; Vorontsova, I.; Hoshino, M.; Uesugi, K.; Yagi, N.; Hall, J.E.; Schilling, T.F.; Pierscionek, B.K. Optical Development in the Zebrafish Eye Lens. FASEB J. 2020, 34, 5552–5562. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, D.E.; Gislén, L.; Coates, M.M.; Skogh, C.; Garm, A. Advanced Optics in a Jellyfish Eye. Nature 2005, 435, 201–205. [Google Scholar] [CrossRef] [PubMed]
- Gál, J.; Bobkova, M.V.; Zhukov, V.V.; Shepeleva, I.P.; Meyer-Rochow, V.B. Fixed Focal-Length Optics in Pulmonate Snails (Mollusca, Gastropoda): Squaring Phylogenetic Background and Ecophysiological Needs (II). Invertebr. Biol. 2004, 123, 116–127. [Google Scholar] [CrossRef]
- Zhukov, V.V.; Borissenko, S.L.; Zieger, M.V.; Vakoliuk, I.A.; Meyer-Rochow, V.B. The Eye of the Freshwater Prosobranch Gastropod Viviparus Viviparus: Ultrastructure, Electrophysiology and Behaviour. Acta Zool. 2006, 87, 13–24. [Google Scholar] [CrossRef]
- Newell, G.E. The Eye of Littorina Littorea. J. Zool. 1965, 144, 75–87. [Google Scholar] [CrossRef]
- SEYER, J.-O. Resolution and Sensitivity in the Eye of the Winkle Littorina Littorea. J. Exp. Biol. 1992, 170, 57–69. [Google Scholar] [CrossRef]
- Seyer, J.-O. Structure and Optics of the Eye of the Hawk-Wing Conch, Strombus raninus (L.). J. Exp. Zool. 1994, 268, 200–207. [Google Scholar] [CrossRef]
- Hamilton, P.V.; Ardizzoni, S.C.; Penn, J.S. Eye Structure and Optics in the Intertidal Snail, Littorina Irrorata. J. Comp. Phys. 1983, 152, 435–445. [Google Scholar] [CrossRef]
- Salvini-Plawen, L.V.; Steiner, G. The Testaria Concept (Polyplacophora + Conchifera) Updated. J. Nat. Hist. 2014, 48, 2751–2772. [Google Scholar] [CrossRef]
- Wanninger, A.; Wollesen, T. The Evolution of Molluscs. Biol. Rev. 2019, 94, 102–115. [Google Scholar] [CrossRef] [PubMed]
- Schrödl, M.; Stöger, I. A review on deep molluscan phylogeny: Old markers, integrative approaches, persistent problems. J. Nat. Hist. 2014, 48, 2773–2804. [Google Scholar] [CrossRef]
- Swardfager, W.; Mitchell, J. Purification of Visual Arrestin from Squid Photoreceptors and Characterization of Arrestin Interaction with Rhodopsin and Rhodopsin Kinase. J. Neurochem. 2007, 101, 223–231. [Google Scholar] [CrossRef]
- Matsuo, R.; Takatori, Y.; Hamada, S.; Koyanagi, M.; Matsuo, Y. Expression and Light-Dependent Translocation of β-Arrestin in the Visual System of the Terrestrial Slug Limax Valentianus. J. Exp. Biol. 2017, 220, 3301–3314. [Google Scholar] [CrossRef] [Green Version]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Crystallins | Number of Analyzed Sequences | The Length of the Analyzed Sequences, in Amino Acid Residues | Normalized Conservation Index by Mirny, Mean ± SD | Number of Identical Amino Acids | Number of Conservative Substitutions | Number of Semi-Conservative Substitutions |
|---|---|---|---|---|---|---|
| α-crystallins | 23 | Minimum: 87 Maximum: 233 Average: 156 | 0.83 ± 0.26 | 2 | 19 | 7 |
| J1A-crystallins | 9 | Minimum: 254 Maximum: 334 Average: 286 | 0.68 ± 0.26 | 51 | 32 | 20 |
| λ-crystallins | 19 | Minimum: 298 Maximum: 333 Average: 320 | 0.65 ± 0.29 | 30 | 52 | 20 |
| μ-crystallins | 9 | Minimum: 250 Maximum: 312 Average: 287 | 0.72 ± 0.27 | 72 | 46 | 27 |
| Ω-crystallins | 3 | Minimum: 492 Maximum: 495 Average: 494 | 0.85 ± 0.25 | 274 | 117 | 33 |
| S-crystallins | 51 | Minimum: 182 Maximum: 318 Average: 220 | 0.75 ± 0.25 | 1 | 4 | 6 |
| ζ-crystallins | 3 | Minimum: 105 Maximum: 323 Average: 239 | 0.64 ± 0.20 | 34 | 25 | 9 |
| Gastropoda molluscs | Crystallin | Mollusc Protein, GenBank Accession Number | Percent Identity, % | E-Value |
|---|---|---|---|---|
| versus Cephalopoda | ||||
| Aplysia californica | α (n = 1) | XP_029637045.1 O. sinensis XP_014782498.1 O. bimaculoides XP_029642860.1 O. sinensis XP_014783828.1 O. bimaculoides XP_014783827.1 O. bimaculoides | 56.02 55.15 43.37 43.37 43.37 | 4 × 10−56 3 × 10−56 2 × 10−21 2 × 10−21 3 × 10−21 |
| J1A (n = 2) | XP_014782921.1 O. bimaculoides XP_036358994.1 O. sinensis | 44.81 42.98 | 4 × 10−72 7 × 10−70 | |
| λ (n = 7) | XP_029654653.1 O. sinensis XP_014776757.1 O. bimaculoides XP_036356167.1 O. sinensis XP_036356168.1 O. sinensis XP_014787243.1 O. bimaculoides | 45.64 45.30 45.14 44.91 28.81 | 3 × 10−102 7 × 10−102 6 × 10−97 1 × 10−69 5 × 10−17 | |
| µ (n = 1) | XP_014772028.1 O. bimaculoides XP_036359829.1 O. sinensis XP_014772027.1 O. bimaculoides XP_029637902.1 O. sinensis | 47.93 47.52 45.61 45.27 | 3 × 10−76 1 × 10−74 2 × 10−90 3 × 10−88 | |
| S short loop (n = 6) | XP_014771120.1 O. bimaculoides XP_014771119.1 O. bimaculoides XP_036355061.1 O. sinensis XP_014772688.1 O. bimaculoides XP_036359314.1 O. sinensis | 38.89 38.38 35.54 33.80 33.17 | 7 × 10−18 2 × 10−15 6 × 10−18 5 × 10−16 2 × 10−22 | |
| Biomphalaria glabrata | α (n = 6) | XP_014783064.1 O. bimaculoides XP_014783063.1 O. bimaculoides XP_029654771.1 O. sinensis XP_036354796.1 O. sinensis XP_029637045.1 O. sinensis | 52.63 52.63 51.58 42.75 41.94 | 1 × 10−28 1 × 10−28 4 × 10−28 4 × 10−28 3 × 10−15 |
| J1A (n = 2) | XP_014782921.1 O. bimaculoides XP_036358994.1 O. sinensis | 43.06 41.67 | 9 × 10−40 7 × 10−38 | |
| λ (n = 2) | XP_036356167.1 O. sinensis XP_029654653.1 O. sinensis XP_014776757.1 O. bimaculoides XP_036356168.1 O. sinensis XP_014769218.1 O. bimaculoides | 48.34 47.92 47.60 44.91 29.17 | 3−110 1 × 10−114 2 × 10−115 3 × 10−70 4 × 10−12 | |
| µ (n = 1) | XP_036359829.1 O. sinensis XP_014772028.1 O. bimaculoides XP_029637902.1 O. sinensis XP_014772027.1 O. bimaculoides | 47.11 45.12 44.75 43.67 | 1 × 10−63 7 × 10−63 1 × 10−7 53 × 10−76 | |
| S short loop (n = 3) | XP_036359020.1 O. sinensis XP_029636899.1 O. sinensis XP_036358738.1 O. sinensis XP_014783449.1 O. bimaculoides XP_036355061.1 O. sinensis | 49.46 45.16 42.86 42.86 37.40 | 4 × 10−25 1 × 10−22 1 × 10−20 2 × 10−20 4 × 10−23 | |
| ζ (n = 2) | XP_036356064.1 O. sinensis XP_014777379.1 O. bimaculoides XP_029644700.1 O. sinensis XP_036359047.1 O. sinensis XP_014784767.1 O. bimaculoides | 37.93 33.33 32.52 32.52 31.28 | 3 × 10−27 3 × 10−09 1 × 10−09 1 × 10−09 5 × 10−14 | |
| Pomacea canaliculata | α (n = 8) | XP_029654771.1 O. bimaculoides XP_014783063.1 O. sinensis XP_014783064.1 O. bimaculoides XP_036354796.1 O. sinensis XP_029642860.1 O. sinensis | 45.00 45.00 44.14 43.33 41.67 | 5 × 10−23 1 × 10−22 5 × 10−24 2 × 10−24 7 × 10−12 |
| J1A (n = 1) | XP_014782921.1 O. bimaculoides XP_036358994.1 O. sinensis | 47.35 47.35 | 1 × 10−81 2 × 10−81 | |
| λ (n = 3) | XP_036356168.1 O. sinensis XP_029654653.1 O. sinensis XP_014776757.1 O. bimaculoides XP_036356167.1 O. sinensis | 47.12 44.41 44.41 43.56 | 2 × 10−51 3 × 10−80 2 × 10−79 5 × 10−75 | |
| µ (n = 2) | XP_014772028.1 O. bimaculoides XP_036359829.1 O. sinensis XP_014772027.1 O. bimaculoides XP_029637902.1 O. sinensis | 46.56 46.56 44.44 44.12 | 3 × 10−62 1 × 10−60 1 × 10−78 4 × 10−76 | |
| S long loop (n = 1) | XP_036355061.1 O. sinensis XP_036359020.1 O. sinensis XP_029637027.1 O. sinensis XP_029636899.1 O. sinensis XP_029636493.1 O. sinensis | 39.78 38.00 37.75 36.73 34.95 | 6 × 10−17 4 × 10−17 2 × 10−36 3 × 10−16 5 × 10−33 | |
| versus Bivalvia | ||||
| Aplysia californica | α (n = 1) | XP_045176285.1 M. mercenaria XP_045216391.1 M. mercenaria XP_033752285.1 P. maximus XP_045216364.1 M. mercenaria XP_021346583.1 M. yessoensis | 55.68 49.14 48.80 48.57 47.59 | 1 × 10−54 9 × 10−46 5 × 10−43 1 × 10−44 6 × 10−41 |
| J1A (n = 2) | XP_021365545.1 M. yessoensis XP_033731119.1 P. maximus XP_045200778.1 M. mercenaria XP_048774863.1 O. edulis XP_033743114.1 P. maximus | 51.95 51.29 49.21 44.87 42.86 | 4 × 10−80 3 × 10−81 1 × 10−77 6 × 10−67 1 × 10−44 | |
| λ (n = 7) | XP_033739698.1 P. maximus XP_011439697.2 C. gigas XP_022295241.1 C. virginica XP_033739699.1 P. maximus XP_048771868.1 O. edulis | 62.34 62.21 61.87 61.84 61.20 | 7 × 10−138 1 × 10−135 9 × 10−134 2 × 10−126 3 × 10−139 | |
| µ (n = 1) | XP_048762137.1 O. edulis XP_011454698.2 C. gigas XP_021361605.1 M. yessoensis XP_048762136.1 O. edulis XP_033746700.1 P. maximus | 57.49 54.64 53.02 52.82 52.68 | 1 × 10−97 4 × 10−116 3 × 10−108 8 × 10−111 2 × 10−107 | |
| S short loop (n = 6) | XP_045194312.1 M. mercenaria XP_021339566.1 M. yessoensis XP_021378340.1 M. yessoensis XP_048759013.1 O. edulis XP_045167239.1 M. mercenaria XP_033754085.1 P. maximus | 50.00 45.00 41.76 40.30 39.71 36.82 | 5 × 10−17 1 × 10−13 2 × 10−20 3 × 10−41 4 × 10−09 4 × 10−33 | |
| Biomphalaria glabrata | α (n = 6) | XP_045166562.1 M. mercenaria XP_045202791.1 M. mercenaria XP_045176285.1 M. mercenaria XP_045203286.1 M. mercenaria XP_045192081.1 M. mercenaria XP_033730743.1 P. maximus | 42.27 39.33 38.71 38.64 38.64 34.41 | 3 × 10−22 6 × 10−17 1 × 10−14 2 × 10−15 4 × 10−16 2 × 10−15 |
| J1A (n = 2) | XP_021365545.1 M. yessoensis XP_033731119.1 P. maximus XP_045200778.1 M. mercenaria XP_022323163.1 C. virginica XP_011426564.1 C. gigas | 48.00 46.97 43.18 37.86 36.43 | 4 × 10−36 7 × 10−38 4 × 10−35 5 × 10−23 3 × 10−20 | |
| λ (n = 2) | XP_048770804.1 O. edulis XP_034331263.1 C. gigas XP_034331265.1 C. gigas XP_045195917.1 M. mercenaria XP_034331611.1 C. gigas XP_033739696.1 P. maximus | 58.65 57.51 57.51 54.95 54.95 51.94 | 1 × 10−134 2 × 10−132 3 × 10−132 9 × 10−131 1 × 10−119 4 × 10−119 | |
| µ (n = 1) | XP_011454698.2 C. gigas XP_021361605.1 M. yessoensis XP_048762137.1 O. edulis XP_033746700.1 P. maximus XP_022319189.1 C. virginica | 49.49 49.32 49.19 49.15 48.12 | 8 × 10−89 1 × 10−86 4 × 10−71 3 × 10−87 4 × 10−85 | |
| S short loop (n = 3) | XP_045194312.1 M. mercenaria XP_034305106.1 C. gigas XP_021378340.1 M. yessoensis XP_033755358.1 P. maximus XP_034308304.1 C. gigas | 54.17 44.95 43.75 41.61 40.70 | 5 × 10−21 6 × 10−25 6 × 10−23 6 × 10−36 2 × 10−42 | |
| ζ (n = 2) | XP_045204266.1 M. mercenaria XP_021380229.1 M. yessoensis XP_045204273.1 M. mercenaria XP_045204248.1 M. mercenaria XP_045204257.1 M. mercenaria XP_033732342.1 P. maximus | 52.16 51.88 51.84 51.84 50.92 49.38 | 1 × 10−88 2 × 10−86 7 × 10−88 1 × 10−87 2 × 10−82 1 × 10−82 | |
| Pomacea canaliculata | α (n = 8) | XP_045166562.1 M. mercenaria XP_045202791.1 M. mercenaria XP_045206475.1 M. mercenaria XP_045203286.1 M. mercenaria XP_022298136.1 C. virginica XP_033752285.1 P. maximus | 43.43 42.11 41.76 41.05 40.70 38.20 | 1 × 10−20 6 × 10−19 1 × 10−13 3 × 10−18 1 × 10−13 2 × 10−11 |
| J1A (n = 1) | XP_045200778.1 M. mercenaria XP_021365545.1 M. yessoensis XP_033731119.1 P. maximus XP_011426564.1 C. gigas XP_048774863.1 O. edulis | 56.15 56.13 55.84 50.96 50.57 | 8 × 10−84 8 × 10−92 7 × 10−92 1 × 10−68 5 × 10−79 | |
| λ (n = 3) | XP_048770804.1 O. edulis XP_034331265.1 C. gigas XP_034331271.1 C. gigas XP_045195917.1 M. mercenaria XP_034331263.1 C. gigas XP_033739696.1 P. maximus | 46.73 46.23 45.63 45.62 45.22 43.91 | 2 × 10−82 2 × 10−79 4 × 10−21 3 × 10−85 6 × 10−78 6 × 10−72 | |
| µ (n = 2) | XP_048762137.1 O. edulis XP_033746700.1 P. maximus XP_011454698.2 C. gigas XP_048762136.1 O. edulis XP_021361605.1 M. yessoensis | 54.88 53.02 53.00 52.32 52.16 | 3 × 10−84 2 × 10−101 1 × 10−104 2 × 10−101 2 × 10−102 | |
| S long loop (n = 1) | XP_045194312.1 M. mercenaria XP_021378342.1 M. yessoensis XP_045164188.1 M. mercenaria XP_021378354.1 M. yessoensis XP_034305103.1 C. gigas XP_033755358.1 P. maximus | 44.74 42.36 42.05 41.87 41.67 38.68 | 1 × 10−16 7 × 10−43 3 × 10−14 1 × 10−47 4 × 10−40 6 × 10−36 | |
| Organism | Crystallin | GRAVY | Prevailing Secondary Structure, % | Proportion of 7 Amino Acids with the Highest Value of the Increment of Refractive Index, % | Calculated Average Increment of Refractive Index Values (dn/dc, mL/g) |
|---|---|---|---|---|---|
| Octopus sp. | α (n = 5) | −0.7226 ± 0.1231 | 63.20% ± 6.54 Coil | 23.00% ± 3.23 | 0.1885 ± 0.0018 |
| J1A (n = 3) | −0.2980 ± 0.0225 | 57.00% Alpha Helix | 15.23% ± 0.46 | 0.1857 ± 0.0006 | |
| λ (n = 4) | −0.1243 ±0.0899 | 49.75% ± 6.75 Alpha Helix | 17.15% ± 0.56 | 0.1854 ± 0.0003 | |
| µ (n = 4) | 0.1430 ± 0.0530 | 41.00% ± 2.71 Coil | 15.68% ± 0.78 | 0.1840 ± 0.0002 | |
| Ω (n = 2) | −0.3710; −0.3780 | 43.00% Coil | 21.20%; 21.60% | 0.1878 | |
| S long loop 1 (n = 21) | −0.6250 ± 0.1360 | 53.62% ± 4.40 Alpha Helix | 39.45% ± 2.91 | 0.1938 ± 0.0009 | |
| S short loop (n = 18) | −0.5134 ± 0.1373 | 54.78% ± 4.71 Alpha Helix | 31.79% ± 5.36 | 0.1917 ± 0.0018 | |
| ζ (n = 1) | 0.1950 | 44.00% Coil | 20.30% | 0.1861 | |
| Pecten maximus/Placopecten magellanicus | α (n = 3) | −0.6657 ± 0.3234 | 61.67% ± 1.53 Coil | 24.97% ± 3.09 | 0.1882 ± 0.0014 |
| J1A (n = 3) | −0.2590 ±0.0641 | 56.67% ± 1.16 Alpha Helix | 18.00% ± 0.50 | 0.1855 ± 0.0005 | |
| λ (n = 4) | −0.2590 ± 0.0786 | 51.25% ± 2.22 Alpha Helix | 19.55% ± 0.31 | 0.1861 ± 0.0002 | |
| µ (n = 1) | 0.0880 | 43.00% Coil | 12.60% | 0.1825 | |
| Ω 1 (n = 1) | −0.1080 | 42.00% Coil | 18.00% | 0.1859 | |
| S short loop (n = 6) | −0.1713 ± 0.3945 | 53.33% ± 3.39 Alpha Helix | 19.45% ± 3.12 | 0.1869 ± 0.0016 | |
| Aplysia californica | α (n = 1) | −0.6620 | 60.00% Coil | 24.30% | 0.1883 |
| J1A (n = 2) | −0.2460; −0.2870 | 51.00% & 60.00% Alpha Helix | 16.80%; 17.40 | 0.1847 | |
| λ (n = 7) | −0.1139 ± 0.0428 | 51.86% ± 0.90 Alpha Helix | 18.17% ± 0.92 | 0.1850 ± 0.0009 | |
| µ (n = 1) | −0.0060 | 44.00% Coil | 16.30% | 0.1837 | |
| S short loop (n = 6) | −0.2362 ± 0.0807 | 53.00% ± 1.79 Alpha Helix | 20.72% ± 2.38 | 0.1881 ± 0.0016 | |
| Biomphalaria glabrata | α (n = 6) | −0.7657 ± 0.1068 | 53.17% ± 3.82 Coil | 16.48% ± 2.79 | 0.1852 ±0.0010 |
| J1A (n = 2) | −0.2480; −0.4090 | 56.00% Alpha Helix; 55.00% Coil | 21.00%; 23.00% | 0.1866; 0.1894 | |
| λ (n = 2) | −0.1470; −0.0970 | 51.00% Alpha Helix | 18.10%; 19.00% | 0.1848; 0.1859 | |
| µ (n = 1) | 0.2200 | 42.00% Coil | 16.60% | 0.1844 | |
| S short loop (n = 3) | −0.1920 ± 0.0439 | 59.67% ± 11.59 Alpha Helix | 20.67% ± 1.55 | 0.1882 ± 0.0006 | |
| ζ (n = 2) | −0.0690; 0.2060 | 44.00%; 48.00% Coil | 13.00%; 17.20% | 0.1830; 0.1850 | |
| Pomacea canaliculata | α (n = 8) | −0.6266 ± 0.1022 | 60.6250% ± 6.3230 Coil | 21.96% ± 1.60 | 0.1873 ± 0.0008 |
| J1A (n = 1) | −0.3640 | 56.00% Alpha Helix | 18.10% | 0.1858 | |
| λ (n = 3) | −0.1807 ± 0.0560 | 51.33% ± 1.53 Alpha Helix | 20.10% ± 2.17 | 0.1855 ± 0.0006 | |
| µ (n = 2) | 0.1130; 0.0480 | 43.00%; 45.00% Coil | 14.90%; 14.80% | 0.1832 | |
| S long loop (n = 1) | −0.5500 | 57.00% Alpha Helix | 23.50% | 0.1903 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dominova, I.N.; Zhukov, V.V. Mollusc Crystallins: Physical and Chemical Properties and Phylogenetic Analysis. Diversity 2022, 14, 827. https://doi.org/10.3390/d14100827
Dominova IN, Zhukov VV. Mollusc Crystallins: Physical and Chemical Properties and Phylogenetic Analysis. Diversity. 2022; 14(10):827. https://doi.org/10.3390/d14100827
Chicago/Turabian StyleDominova, Irina N., and Valery V. Zhukov. 2022. "Mollusc Crystallins: Physical and Chemical Properties and Phylogenetic Analysis" Diversity 14, no. 10: 827. https://doi.org/10.3390/d14100827
APA StyleDominova, I. N., & Zhukov, V. V. (2022). Mollusc Crystallins: Physical and Chemical Properties and Phylogenetic Analysis. Diversity, 14(10), 827. https://doi.org/10.3390/d14100827