
How Many Abalone Species Live in the Mediterranean Sea?

 , ,
, ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
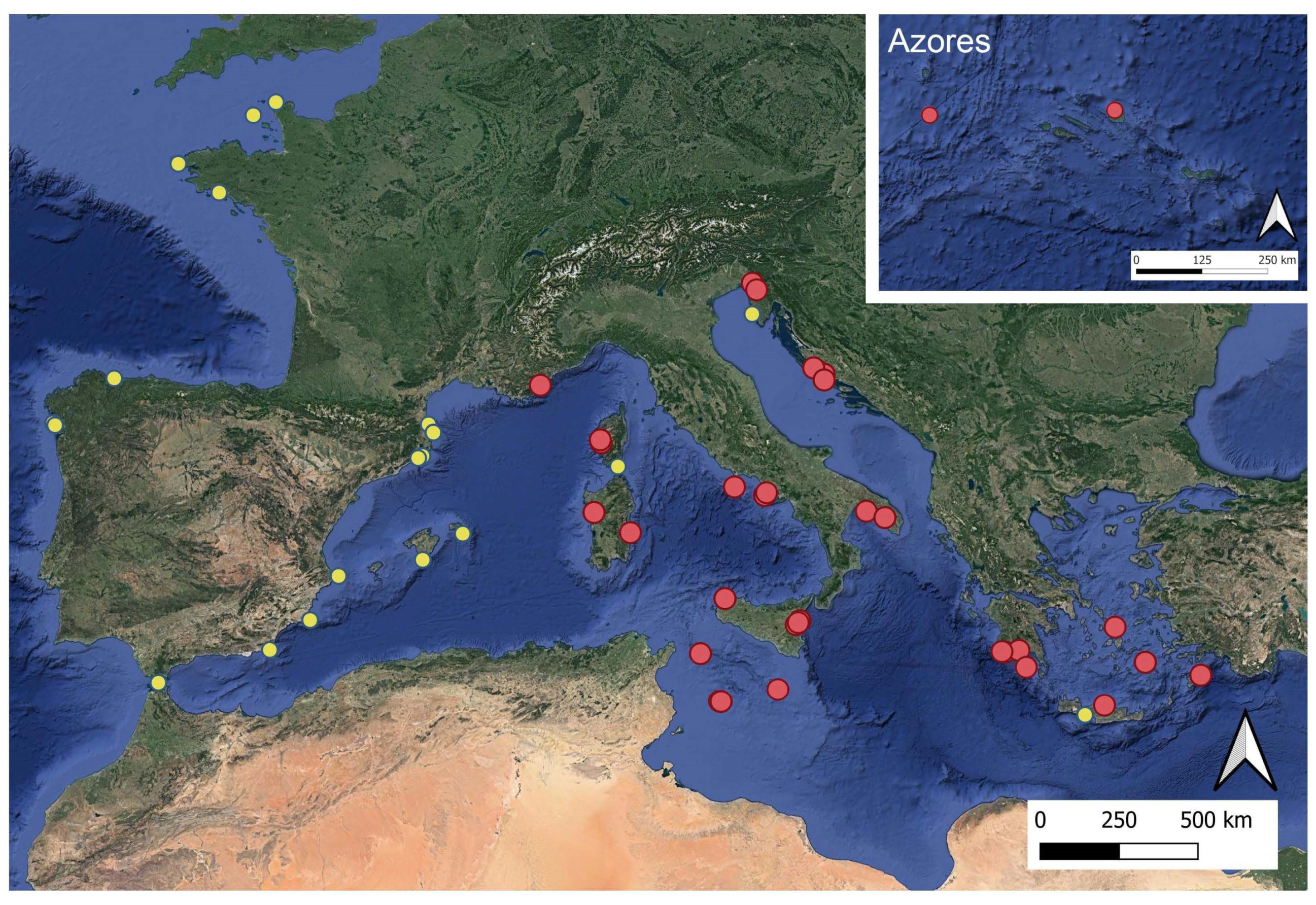
2.1. Dataset
2.2. Laboratory Analyses
2.3. Bioinformatic Analyses
3. Results
3.1. Dataset
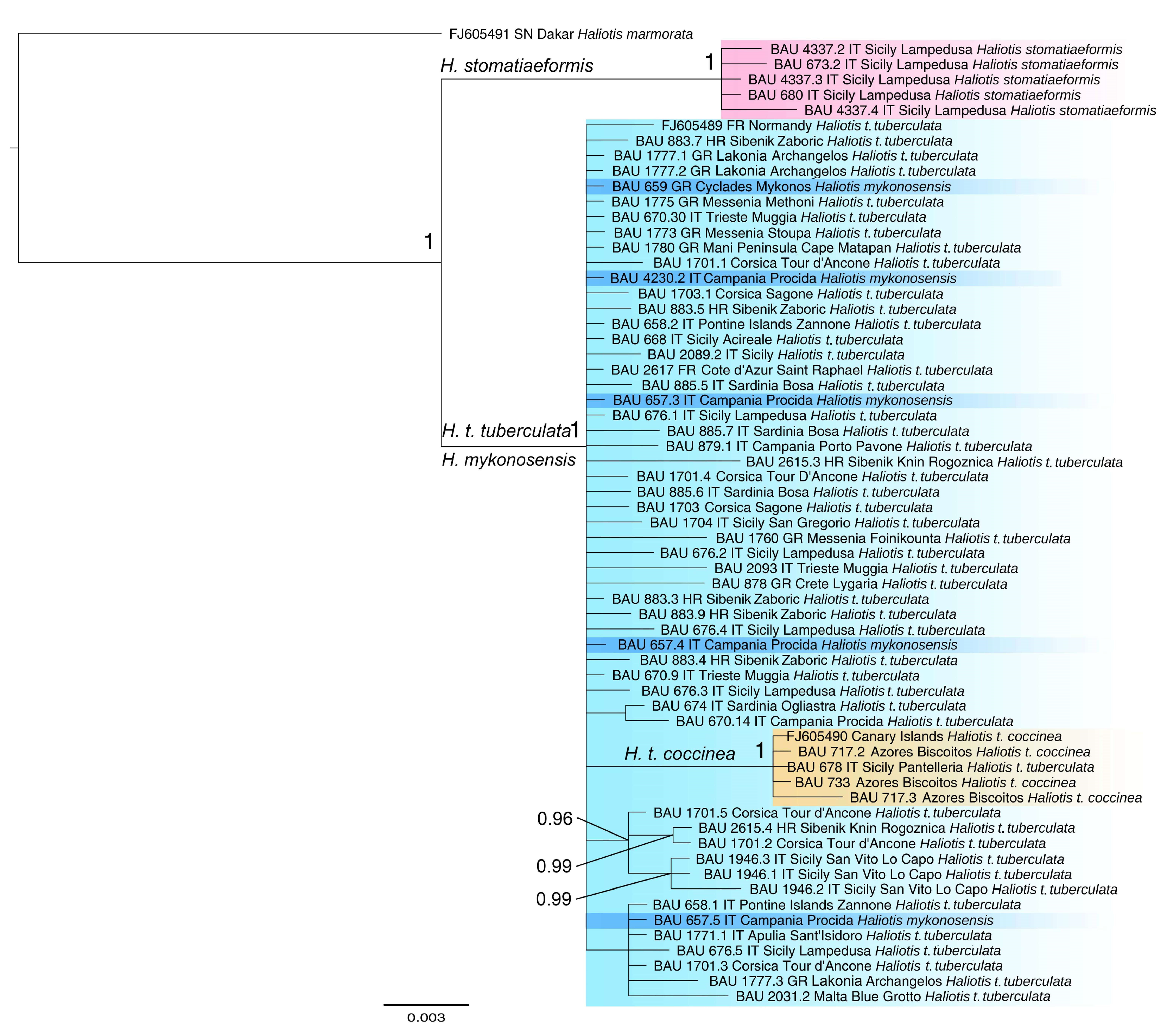
3.2. Bayesian Inference
3.3. Species Delimitation Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Puillandre, N.; Baylac, M.; Boisselier, M.-C.; Cruaud, C.; Samadi, S. An Integrative Approach to Species Delimitation in Benthomangelia (Mollusca: Conoidea). Biol. J. Linn. Soc. 2009, 96, 696–708. [Google Scholar] [CrossRef]
- Aissaoui, C.; Puillandre, N.; Bouchet, P.; Fassio, G.; Modica, M.V.; Oliverio, M. Cryptic Diversity in Mediterranean Gastropods of the Genus Aplus (Neogastropoda: Buccinidae). Sci. Mar. 2016, 80, 521–533. [Google Scholar] [CrossRef] [Green Version]
- Fassio, G.; Bouchet, P.; Lozouet, P.; Modica, M.V.; Russini, V.; Schiaparelli, S.; Oliverio, M. Becoming a Limpet: An ‘Intermittent Limpetization’ Process Driven by Host Features in the Kleptoparasitic Gastropod Family Capulidae. Mol. Phylogenet. Evol. 2021, 155, 107014. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological Identifications through DNA Barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hsu, T.H.; Gwo, J.C. Genetic Diversity and Stock Identification of Small Abalone (Haliotis diversicolor) in Taiwan and Japan. PLoS ONE 2017, 12, e0179818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Marshall, B.A.; Kennedy, M.; Spencer, H.G. Molluscan Research, The Phylogeny and Taxonomy of New Zealand Notoacmea and Patelloida Species (Mollusca: Patellogastropoda: Lottiidae) Inferred from DNA Sequences. Molluscan Res. 1926, 29, 1880. [Google Scholar]
- Sharina, S.N.; Chernyshev, A.V.; Zaslavskaya, N.I. Genetic Diversity and Phylogeny of Limpets of the Genus Nipponacmea (Patellogastropoda: Lottiidae) Based on Mitochondrial DNA Sequences. Mitochondrial DNA A DNA Mapp. Seq. Anal. 2017, 28, 703–710. [Google Scholar] [CrossRef]
- Young, M.K.; Smith, R.; Pilgrim, K.L.; Schwartz, M.K. Molecular Species Delimitation Refines the Taxonomy of Native and Nonnative Physinine Snails in North America. Sci. Rep. 2021, 11, 21739. [Google Scholar] [CrossRef]
- Ahyong, S.; Boyko, C.B.; Bailly, N.; Bernot, J.; Bieler, R.; Brandão, S.N.; Daly, M.; de Grave, S.; Gofas, S.; Hernandez, F.; et al. World Register of Marine Species (WoRMS) 2022. Available online: https://www.marinespecies.org/ (accessed on 4 December 2022).
- Geiger, D.L. Distribution and Biogeography of the Recent Haliotidae World-Wide (Gastropoda: Vetigastropoda). Boll. Malacol. 1999, 35, 57–118. [Google Scholar]
- Mercer, J.P.; Mai, K.-S.; Donlon, J. Comparative Studies on the Nutrition of Two Species of Abalone, Haliotis Tuberculata Linnaeus and Haliotis discus hannai Ino I. Effects of Algal Diets on Growth and Biochemical Composition. Invertebr. Reprod. Dev. 1993, 23, 75–88. [Google Scholar] [CrossRef]
- Coleman, A.W.; Vacquier, V.D. Exploring the Phylogenetic Utility of ITS Sequences for Animals: A Test Case for Abalone (Haliotis). J. Mol. Evol. 2002, 54, 246–257. [Google Scholar] [CrossRef] [PubMed]
- Geiger, D.L.; Groves, L.T. Review of Fossil Abalone (Gastropoda: Vetigastropoda: Haliotidae) with Comparison to Recent Species. J. Paleontol. 1999, 73, 872–885. [Google Scholar] [CrossRef]
- Ubaldi, R. The Haliotis in the Atlantic-Mediterranean Area. Argonauta 1987, 3, 268–290. [Google Scholar]
- Bilbao, A.; Tuset, V.; Viera, M.; de Vicose, G.C.; Fernández-Palacios, H.; Haroun, R.; Izquierdo, M. Reproduction, Fecundity, and Growth of Abalone (Haliotis tuberculata coccinea, Reeve 1846) in the Canary Islands. J. Shellfish Res. 2010, 29, 959–967. [Google Scholar] [CrossRef] [Green Version]
- Fernández, R.; Lemer, S.; Mcintyre, E.; Giribet, G. Comparative Phylogeography and Population Genetic Structure of Three Widespread Mollusc Species in the Mediterranean and near Atlantic. Mar. Ecol. 2015, 36, 701–715. [Google Scholar] [CrossRef]
- Gruenthal, K.M.; Acheson, L.K.; Burton, R.S. Genetic Structure of Natural Populations of California Red Abalone (Haliotis rufescens) Using Multiple Genetic Markers. Mar. Biol. 2007, 152, 1237–1248. [Google Scholar] [CrossRef]
- Hart, L.C.; Goodman, M.C.; Walter, R.K.; Rogers-be Ett, L.; Shum, P.; Garrett, A.D.; Watanabe, J.M.; Iferk, J.O. Abalone Recruitment in Low Density and Aggregated Populations Facing Climatic Stress. J. Shellfish Res. 2020, 39, 359–373. [Google Scholar] [CrossRef]
- Mercer, J.P. Abalone Cultivation in Ireland: Current Status; B0078; Shellfish Research Laboratory: Savannah, GA, USA, 1998. [Google Scholar]
- Geiger, D.L. Recent Genera and Species of the Family Haliotidae Rafinesque, 1818 (Gastropoda: Vetigastropoda). Nautilus 1998, 111, 85–116. [Google Scholar]
- Geiger, D.L.; Poppe, G.T. A Conchological Iconography: The Family Haliotidae; Conchbooks: Harxheim, Germany, 2000. [Google Scholar]
- Wagner, R.J.L.; Abbott, R.T. Standard Catalogue of Shells; American Malacologists Inc.: Greenville, DE, USA, 1978. [Google Scholar]
- Lindberg, D.R. Evolution, Distribution and Systematics of Haliotidae. In Abalone of the World: Biology, Fisheries and Culture; Shepherd, S.A., Tegner, M.J., Guzman del Proo, S.A., Eds.; Wiley: London, UK, 1992; pp. 3–18. [Google Scholar]
- Mgaya, Y.D.; Mercer, J.P. A Review of the Biology, Ecology, Fisheries and Mariculture of the European Abalone Haliotis tuberculata Linnaeus 1758 (Gastropoda: Haliotidae). In Biology and Environment: Proceedings of the Royal Irish Academy; Royal Irish Academy: Dublin, Ireland, 1994; Volume 94, pp. 285–304. [Google Scholar]
- Gaillard, J.M. Haliotis Tuberculata Linné. Systématique et Distribution. Bull. Lab. Marit. Dinard 1958, 44, 7–11. [Google Scholar]
- Nordsieck, F. Some Trips to the Canary Islands. Conchiglia 1975, 7, 3–22. [Google Scholar]
- Parenzan, P. Carta d’identità Delle Conchiglie Del Mediterraneo. Volume Primo. Gasteropodi; Taras, B., Ed.; Bios Taras Editrice: Taranto, Italy, 1970; Volume 1. [Google Scholar]
- Lee, Y.-H.; Vaequier, V.D. Evolution and Systematics in Haliotidae (Mollusca: Gastropoda): Inferences from DNA Sequences of Sperm Lysin. Mar. Biol. 1995, 124, 267–278. [Google Scholar] [CrossRef]
- Owen, B. The “Buzz” on Abalones. Haliotis Mykonosensis 2004, 26, 129–131. [Google Scholar]
- Owen, B.; Hanavan, S.; Hall, S. A New Species of Abalone (Haliotis) from Greece. Veliger 2001, 44, 301–309. [Google Scholar]
- Bo, M.; al Mabruk, S.A.A.; Balistreri, P.; Bariche, M.; Batjakas, I.E.; Betti, F.; Bilan, M.; Canese, S.; Cattaneo-Vietti, R.; Corsini-Foka, M.; et al. New Records of Rare Species in the Mediterranean Sea (October 2020). Mediterr. Mar. Sci. 2020, 21, 608–630. [Google Scholar] [CrossRef]
- Crocetta, F.; Rismondo, S. Haliotis mykonosensis Owen, Hanavan & Hall, 2001 in the Procida Island (Gulf of Naples) and in the Central Mediterranean Sea, with Notes on the Mediterranean Haliotidae. Mediterr. Mar. Sci. 2009, 10, 139–144. [Google Scholar] [CrossRef] [Green Version]
- Geiger, D.L.; Owen, B. The Identity of Haliotis stomatiaeformis Reeve, 18846, from the Mediterranean Sea. Nautilus 2001, 11, 77–83. [Google Scholar]
- Owen, B. The Buzz on Abalones. The Neglected Haliotis: Haliotis stomatiaeformis Reeve, 1846. Sea Shore 2003, 25, 286–289. [Google Scholar]
- van Wormhoudt, A.; le Bras, Y.; Huchette, S. Haliotis marmorata from Senegal; a Sister Species of Haliotis tuberculata: Morphological and Molecular Evidence. Biochem. Syst. Ecol. 2009, 37, 747–755. [Google Scholar] [CrossRef]
- Oliverio, M.; Mariottini, P. Contrasting Morphological and Molecular Variation in Coralliophila meyendorffii (Muricidae, Coralliophilinae). J. Molluscan Stud. 2001, 67, 243–246. [Google Scholar] [CrossRef] [Green Version]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. Conserved Primers for PCR Amplification of Mitochondrial DNA from Different Invertebrate Phyla. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Colgan, D.J.; Hutchings, P.A.; Brown, S. Phylogenetic Relationships within the Terebellomorpha. J. Mar. Biol. Assoc. 2001, 81, 765–773. [Google Scholar] [CrossRef]
- Leray, M.; Yang, J.Y.; Meyer, C.P.; Mills, S.C.; Agudelo, N.; Ranwez, V.; Boehm, J.T.; Machida, R.J. A New Versatile Primer Set Targeting a Short Fragment of the Mitochondrial COI Region for Metabarcoding Metazoan Diversity: Application for Characterizing Coral Reef Fish Gut Contents. Front. Zool. 2013, 10, 34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puillandre, N.; Brouillet, S.; Achaz, G. ASAP: Assemble Species by Automatic Partitioning. Mol. Ecol. Resour. 2021, 21, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. JModelTest 2: More Models, New Heuristics and Parallel Computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guindon, S.; Gascuel, O. A Simple, Fast, and Accurate Algorithm to Estimate Large Phylogenies by Maximum Likelihood. Syst. Biol. 2003, 52, 696–704. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Rambaut, A.; Drummond, A.J. FigTree: Tree Figure Drawing Tool, v.1.4.4. 2018. Available online: http://tree.bio.ed.ac.uk/software/figtree/ (accessed on 10 December 2022).
- Rambaut, A.; Drummond, A.J.; Xie, D.; Baele, G.; Suchard, M.A. Posterior Summarization in Bayesian Phylogenetics Using Tracer 1.7. Syst. Biol. 2018, 67, 901–904. [Google Scholar] [CrossRef] [Green Version]
- van Wormhoudt, A.; Roussel, V.; Courtois, G.; Huchette, S. Mitochondrial DNA Introgression in the European Abalone Haliotis tuberculata tuberculata: Evidence for Experimental MtDNA Paternal Inheritance and a Natural Hybrid Sequence. Mar. Biotechnol. 2011, 13, 563–574. [Google Scholar] [CrossRef]
- Gaillard, J.M. Gastéropodes. In Fiches DAO D’identification des Espèces pour les Besoins de la Pêche (Révision 1). Méditerranée et mer Noire. Zone de Pêche 37. Volume 1. Végétaux et Invertébrés; Fischer, W., Bauchot, M.L., Schneider, M., Eds.; FAO: Rome, Italy, 1987; Volume 1, pp. 513–632. [Google Scholar]
- Garavelli, C.L. Postille Su Haliotis tuberculata Ed Haliotis lamellosa. Conchiglie 1968, 4, 119–120. [Google Scholar]
- Garavelli, C.L. Le Haliotis Della Provincia Lusitanica. Conchiglie 1968, 4, 93–115. [Google Scholar]
- Gofàs, S.; Moreno, D.; Salas, C. Moluscos Marinos de Andalucía; Universidad de Málaga, Servicio de Publicaciones e Intercambio Científico: Málaga, Sapin, 2011; Volume 1. [Google Scholar]
- Lanza, B.; Manfredi, P.; Moretti, G.; Piersanti, G.; Ruffo, S.; Tortonese, E.; Toschi, A. La Fauna; Touring Club Italiano, Ed.; Conosci: Milano, Italy, 1959; Volume III. [Google Scholar]
- D’Angelo, G.; Gargiullo, S. Guida Alle Conchiglie Mediterranee. Conoscerle Cercarle Collezionarle; Fabbri Editori: Milan, Italy, 1978. [Google Scholar]
- Barash, A.; Danin, Z. Fauna Palestina. Mollusca 1. Annotated List of Mediterranean Molluscs of Israel and Sinai; Israel Academy of Sciences and Humanities: Jerusalem, Israel, 1992; Volume 1. [Google Scholar]
- Roussel, V.; van Wormhoudt, A. The Effect of Pleistocene Climate Fluctuations on Distribution of European Abalone (Haliotis tuberculata), Revealed by Combined Mitochondrial and Nuclear Marker Analyses. Biochem. Genet. 2017, 55, 124–154. [Google Scholar] [CrossRef] [PubMed]
- Clark, N.L.; Findlay, G.D.; Yi, X.; MacCoss, M.J.; Swanson, W.J. Duplication and Selection on Abalone Sperm Lysin in an Allopatric Population. Mol. Biol. Evol. 2007, 24, 2081–2090. [Google Scholar] [CrossRef]
- Giannuzzi-Savelli, R. Atlante Delle Conchiglie Marine Del Mediterraneo: Neogastropoda Parte 1: Muricoidea; Edizioni de “La Conchiglia,”: Port Elgin, ON, Canada, 1994. [Google Scholar]
- Russini, V.; Fassio, G.; Modica, M.V.; deMaintenon, M.J.; Oliverio, M. An Assessment of the Genus Columbella Lamarck, 1799 (Gastropoda: Columbellidae) from Eastern Atlantic. Zoosystema 2017, 39, 197–212. [Google Scholar] [CrossRef] [Green Version]
- Fassio, G.; Bouchet, P.; Oliverio, M.; Strong, E.E. Re-Evaluating the Case for Poecilogony in the Gastropod Planaxis sulcatus (Cerithioidea, Planaxidae). BMC Ecol. Evol. 2022, 22, 13. [Google Scholar] [CrossRef] [PubMed]
- An, H.S.; Jee, Y.J.; Min, K.S.; Kim, B.L.; Han, S.J. Phylogenetic Analysis of Six Species of Pacific Abalone (Haliotidae) Based on DNA Sequences of 16s RRNA and Cytochrome c Oxidase Subunit I Mitochondrial Genes. Mar. Biotechnol. 2005, 7, 373–380. [Google Scholar] [CrossRef]
- Gruenthal, K.M.; Burton, R.S. Genetic Diversity and Species Identification in the Endangered White Abalone (Haliotis sorenseni). Conserv. Genet. 2005, 6, 929–939. [Google Scholar] [CrossRef]
- Aguilera-Muñoz, F.; Lafarga-Cruz, F.; Gallardo-Escárate, C. Molecular Analysis in Chilean Commercial Gastropods Based on 16S RRNA, COI and ITS1-5.8S RDNA-ITS2 Sequences. Gayana 2009, 73, 17–27. [Google Scholar] [CrossRef] [Green Version]
- Peters, H. Haliotis tuberculata. The IUCN Red List of Threatened Species. 2021: e.T78772221A78772628. Available online: https://www.iucnredlist.org/species/78772221/78772628 (accessed on 6 December 2022).
- Geiger, D.L.; Thacker, C.E. Molecular phylogeny of Vetigastropoda reveals non-monophyletic Scissurellidae, Trochoidea, and Fissurelloidea. Molluscan Res. 2005, 25, 47–55. [Google Scholar]
- Aktipis, S.W.; Giribet, G. A phylogeny of Vetigastropoda and other “archaeogastropods”: Re-organizing old gastropod clades. Invertebr. Biol. 2010, 129, 220–240. [Google Scholar] [CrossRef]
- Gruenthal, K.M.; Burton, R.S. Genetic structure of natural populations of the California black abalone (Haliotis cracherodii Leach, 1814), a candidate for endangered species status. J. Exp. Mar. Biol. Ecol. 2008, 355, 47–58. [Google Scholar] [CrossRef]
- Metz, E.C.; Robles-Sikisaka, R.; Vacquier, V.D. Nonsynonymous substitution in abalone sperm fertilization genes exceeds substitution in introns and mitochondrial DNA. Proc. Natl. Acad. Sci. USA 1998, 95, 10676–10681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Y.; Kang, J.; Chen, W.; He, S. DNA barcoding for the identification of common economic aquatic products in Central China and its application for the supervision of the market trade. Food Control 2016, 61, 79–91. [Google Scholar] [CrossRef]
- Wang, Z.Y.; Ho, K.C.; Ke, C.H.; Mak, W.Y.; Chu, K.H. Lack of genetic divergence in nuclear and mitochondrial DNA between subspecies of two Haliotis species. J. Shellfish. Res. 2004, 23, 1143–1147. [Google Scholar]
- Wang, K.J.; Ren, H.L.; Xu, D.D.; Cai, L.; Yang, M. Identification of the up-regulated expression genes in hemocytes of variously colored abalone (Haliotis diversicolor Reeve, 1846) challenged with bacteria. Dev. Comp. Immunol. 2008, 32, 1326–1347. [Google Scholar] [CrossRef] [PubMed]
- Chan, W.H.; Ling, K.H.; Shaw, P.C.; Chiu, S.W.; But, P.P.H. Application of FINS and multiplex PCR for detecting genuine abalone products. Food Control 2012, 23, 137–142. [Google Scholar] [CrossRef]
- Hawk, H.L.; Geller, J.B. DNA entombed in archival seashells reveals low historical mitochondrial genetic diversity of endangered white abalone Haliotis sorenseni. Mar. Freshw. Res. 2018, 70, 359–370. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species ID | Code | Locality | Coordinates | 5′-COI | 5′-COI mb1 | 5′-COI mb2 | 3′-COI |
|---|---|---|---|---|---|---|---|
| Haliotis mykonosensis | BAU 657.3 | Italy, Procida | 40.7691 N, 14.0196 E | X | X | ||
| Owen, Hanavan & Hall 2001 | BAU 657.4 | Italy, Procida | 40.7691 N, 14.0196 E | X | X | ||
| BAU 657.5 | Italy, Procida | 40.7691 N, 14.0196 E | X | X | |||
| BAU 659 | Greece, Mykonos | 37.4088 N, 25.3465 E | X | X | |||
| BAU 4230.2 | Italy, Procida | 40.7691 N, 14.0196 E | X | X | |||
| Haliotis stomatiaeformis | BAU 667 | Sicily, Acireale | 37.6125 N, 15.1661 E | X | |||
| Reeve, 1846 | BAU 669 | Sicily, Acireale | 37.6125 N, 15.1661 E | X | |||
| BAU 673.1 | Sicily, Lampedusa | 35.5035 N, 12.5839 E | X | X | |||
| BAU 673.2 | Sicily, Lampedusa | 35.5035 N, 12.5839 E | X | X | |||
| BAU 680 | Sicily, Lampedusa | 35.5121 N, 12.5556 E | X | X | X | ||
| BAU 4337.1 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 4337.2 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | X | ||
| BAU 4337.3 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | X | ||
| BAU 4337.4 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | X | ||
| Haliotis t. coccinea | BAU 717.2 | Azores, Flores | 39.4581 N, −31.1251 W | X | X | ||
| Reeve, 1846 | BAU 717.3 | Azores, Flores | 39.4581 N, −31.1251 W | X | X | ||
| BAU 733 | Azores, Terceira | 38.8033 N, −27.2564 W | X | X | |||
| Haliotis t. tuberculata | BAU 658.1 | Italy, Zannone Is. | 40.9694 N, 13.0455 E | X | X | ||
| Linnaeus, 1758 | BAU 658.2 | Italy, Zannone Is. | 40.9694N, 13.0455 E | X | X | ||
| BAU 663.1 | Sicily, Acireale | 37.6125 N, 15.1661 E | X | ||||
| BAU 668 | Sicily, Acireale | 37.6125 N, 15.1661 E | X | X | |||
| BAU 670.9 | Italy, Muggia | 45.6056 N, 13.7216 E | X | X | |||
| BAU 670.14 | Italy, Muggia | 45.6056 N, 13.7216 E | X | X | |||
| BAU 670.30 | Italy, Muggia | 45.6056 N, 13.7216 E | X | X | |||
| BAU 674 | Sardinia, Ogliastra | 39.8408 N, 9.6755 E | X | X | |||
| BAU 676.1 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 676.2 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 676.3 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 676.4 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 676.5 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | X | |||
| BAU 677 | Sicily, Pantelleria | 36.7622 N, 11.9436 E | X | ||||
| BAU 678 | Sicily, Pantelleria | 36.7622 N, 11.9436 E | X | X | |||
| BAU 682 | Sicily, Lampedusa | 35.5011 N, 12.6072 E | X | ||||
| BAU 878 | Crete, Ligaria | 35.3990 N, 25.0283 E | X | X | X | ||
| BAU 879.1 | Italy, Porto Pavone | 40.7945 N, 14.1610 E | X | X | X | ||
| BAU 883.1 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | |||
| BAU 883.3 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | |||
| BAU 883.4 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | |||
| BAU 883.5 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | X | ||
| BAU 883.7 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | |||
| BAU 883.9 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | X | ||
| BAU 883.10 | Croatia, Zaboric | 43.6659 N, 15.9388 E | X | X | |||
| BAU 884 | Italy, Leporano | 40.3754 N, 17.3001 E | X | X | |||
| BAU 885.1 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | ||||
| BAU 885.3 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | X | |||
| BAU 885.4 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | ||||
| BAU 885.5 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | X | X | ||
| BAU 885.6 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | X | X | ||
| BAU 885.7 | Sardinia, Bosa | 40.3145 N, 8.4619 E | X | X | |||
| BAU 886.13 | Croatia, Murter Is. | 43.7969 N, 15.6094 E | X | ||||
| BAU 886.19 | Croatia, Murter Is. | 43.7969 N, 15.6094 E | X | ||||
| BAU 1384 | Greece, Astypalea Is. | 36.5869 N, 26.4028 E | X | ||||
| BAU 1386 | Greece, Astypalea Is. | 36.5869 N, 26.4028 E | X | ||||
| BAU 1387 | Greece, Astypalea Is. | 36.5869 N, 26.4028 E | X | ||||
| BAU 1388 | Greece, Astypalea Is. | 36.5647 N, 26.3533 E | X | ||||
| BAU 1389 | Greece, Astypalea Is. | 36.5647 N, 26.3533 E | X | ||||
| BAU 1390 | Greece, Astypalea Is. | 36.5647 N, 26.3533 E | X | ||||
| BAU 1391 | Greece, Astypalea Is. | 36.5759 N, 26.3931 E | X | X | |||
| BAU 1393 | Greece, Astypalea Is. | 36.5759 N, 26.3931 E | X | X | |||
| BAU 1394 | Greece, Astypalea Is. | 36.5759 N, 26.3931 E | X | X | |||
| BAU 1701.1 | Corsica, Tour D’Ancone | 42.0433 N, 8.7208 E | X | X | |||
| BAU 1701.2 | Corsica, Tour D’Ancone | 42.0433 N, 8.7208 E | X | X | |||
| BAU 1701.3 | Corsica, Tour D’Ancone | 42.0433 N, 8.7208 E | X | X | |||
| BAU 1701.4 | Corsica, Tour D’Ancone | 42.0433 N, 8.7208 E | X | X | |||
| BAU 1701.5 | Corsica, Tour D’Ancone | 42.0433 N, 8.7208 E | X | X | |||
| BAU 1703 | Corsica, Sagone | 42.1049 N, 8.6800 E | X | X | |||
| BAU 1703.1 | Corsica, Sagone | 42.1049 N, 8.6800 E | X | X | |||
| BAU 1704 | Sicily, San Gregorio | 38.1586 N, 14.7602 E | X | X | |||
| BAU 1760 | Greece, Foinikounta Is. | 36.8051 N, 21.8146 E | X | X | |||
| BAU 1771.1 | Italy, Sant’Isidoro | 40.2176 N, 17.9212 E | X | X | |||
| BAU 1773 | Greece, Stoupa | 36.8439 N, 22.2573 E | X | X | |||
| BAU 1775 | Greece, Methoni | 36.8153 N, 21.7030 E | X | X | |||
| BAU 1777.1 | Greece, Archangelos | 36.6293 N, 22.8798 E | X | X | |||
| BAU 1777.2 | Greece, Archangelos | 36.6293 N, 22.8798 E | X | X | |||
| BAU 1777.3 | Greece, Archangelos | 36.6293 N, 22.8798 E | X | X | |||
| BAU 1780 | Greece, Cape Matapan | 36.4014 N, 22.4873 E | X | X | |||
| BAU 1946.1 | Sicily, San Vito Lo Capo | 38.1897 N, 12.7347 E | X | X | |||
| BAU 1946.2 | Sicily, San Vito Lo Capo | 38.1897 N, 12.7347 E | X | X | X | ||
| BAU 1946.3 | Sicily, San Vito Lo Capo | 38.1897 N, 12.7347 E | X | X | |||
| BAU 2031.2 | Malta, Blue Grotto | 35.8197 N, 14.4514 E | X | X | X | ||
| BAU 2089.2 | Sicily, Porto Palo di Menfi | 37.5733 N, 12.9002 E | X | X | X | ||
| BAU 2090 | Italy, Sistiana | 45.7687 N, 13.6253 E | X | X | |||
| BAU 2090.1 | Italy, Sistiana | 45.7687 N, 13.6253 E | X | X | |||
| BAU 2090.2 | Italy, Sistiana | 45.7687 N, 13.6253 E | X | ||||
| BAU 2093 | Italy, Muggia | 45.6056 N, 13.7216 E | X | X | |||
| BAU 2615.1 | Croatia, Knin Rogoznica | 43.5376 N, 15.9583 E | X | X | |||
| BAU 2615.3 | Croatia, Knin Rogoznica | 43.5376 N, 15.9583 E | X | X | X | ||
| BAU 2615.4 | Croatia, Knin Rogoznica | 43.5376 N, 15.9583 E | X | X | X | ||
| BAU 2615.6 | Croatia, Knin Rogoznica | 43.5376 N, 15.9583 E | X | ||||
| BAU 2617 | France, Saint Raphael | 43.4112 N, 6.8461 E | X | X |
| H. marmorata | H. stomatiaeformis | H. tuberculata | H. coccinea | ||
| H. marmorata | - | ||||
| H. stomatiaeformis | 8–8.2 | 0.2–0.4 | |||
| H. tuberculata | 6.7–8.2 | 3–4.4 | 0–1.9 | ||
| H. coccinea | 7.4–7.8 | 3.4–3.8 | 1.3–2.8 | 0.2–0.6 | |
| H. crackerodii | H. rufescens | H. walallensis | H. kamtschatkana | H. sorenseni | |
| H. crackerodii | 0.2 | ||||
| H. rufescens | 8.1–8.5 | 0.2–0.7 | |||
| H. walallensis | 7.4–8.2 | 2.2–2.8 | 0.2–0.5 | ||
| H. kamtschatkana | 7.2–7.8 | 2.4–3.4 | 1.5–2.4 | 0.2–0.6 | |
| H. sorenseni | 7.2–7.6 | 2.2–2.8 | 1.3–1.9 | 0.2–0.9 | 0.2 |
| H. corrugata | H. discus | H. madaka | H. gigantea | ||
| H. corrugata | 0.2 | ||||
| H. discus | 8.4–9.3 | 0.7 | |||
| H. madaka | 8.9–9.1 | 0.6 | - | ||
| H. gigantea | 8.9–9.3 | 2.1–3.0 | 2.2–2.8 | 0.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiappa, G.; Fassio, G.; Corso, A.; Crocetta, F.; Modica, M.V.; Oliverio, M. How Many Abalone Species Live in the Mediterranean Sea? Diversity 2022, 14, 1107. https://doi.org/10.3390/d14121107
Chiappa G, Fassio G, Corso A, Crocetta F, Modica MV, Oliverio M. How Many Abalone Species Live in the Mediterranean Sea? Diversity. 2022; 14(12):1107. https://doi.org/10.3390/d14121107
Chicago/Turabian StyleChiappa, Giacomo, Giulia Fassio, Andrea Corso, Fabio Crocetta, Maria Vittoria Modica, and Marco Oliverio. 2022. "How Many Abalone Species Live in the Mediterranean Sea?" Diversity 14, no. 12: 1107. https://doi.org/10.3390/d14121107
APA StyleChiappa, G., Fassio, G., Corso, A., Crocetta, F., Modica, M. V., & Oliverio, M. (2022). How Many Abalone Species Live in the Mediterranean Sea? Diversity, 14(12), 1107. https://doi.org/10.3390/d14121107