
Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species †

 ,
,  , ,
, , 
Abstract
:1. Introduction
2. Materials and Methods
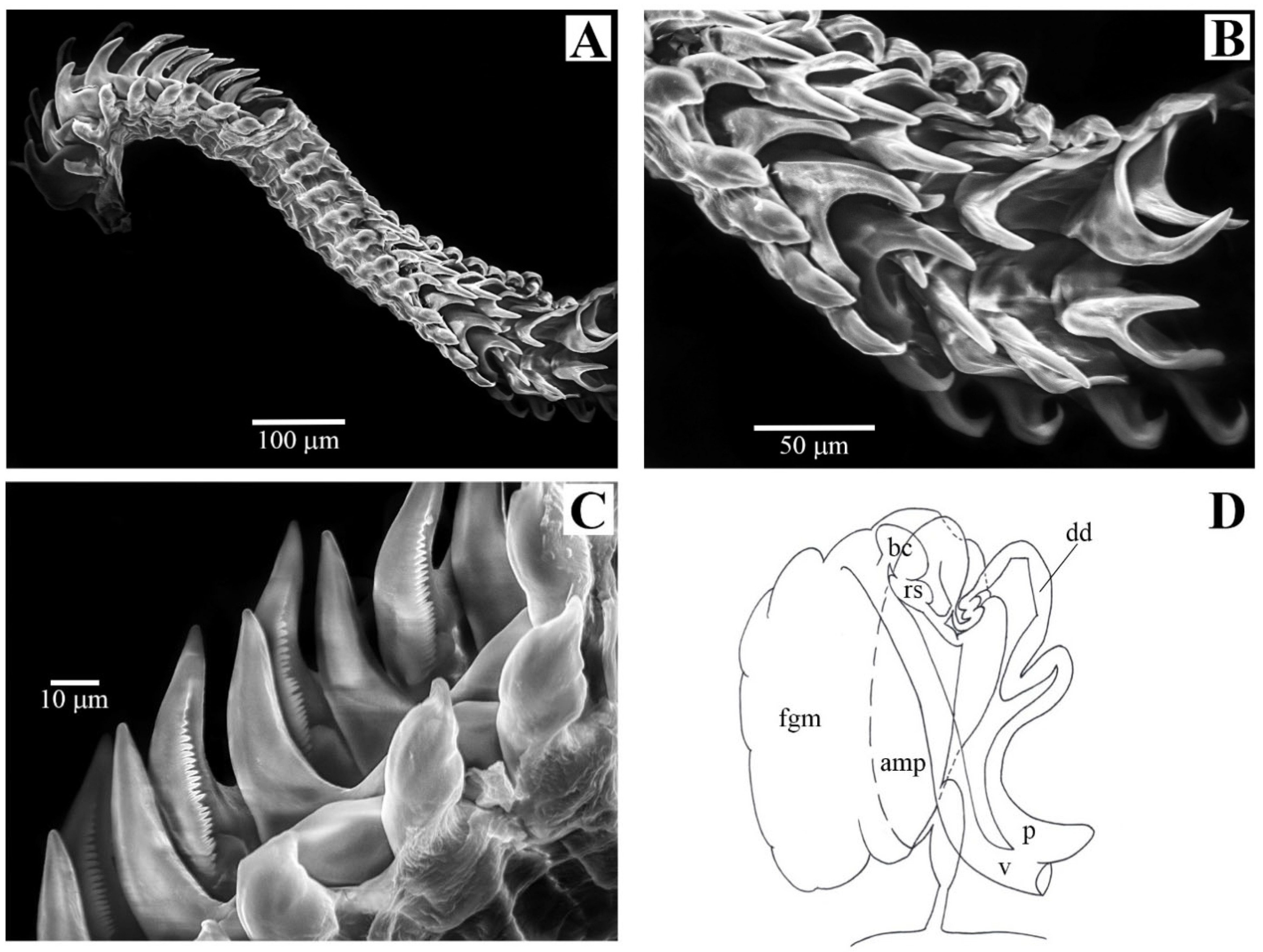
2.1. Morphological Analyses
2.2. Molecular Analyses
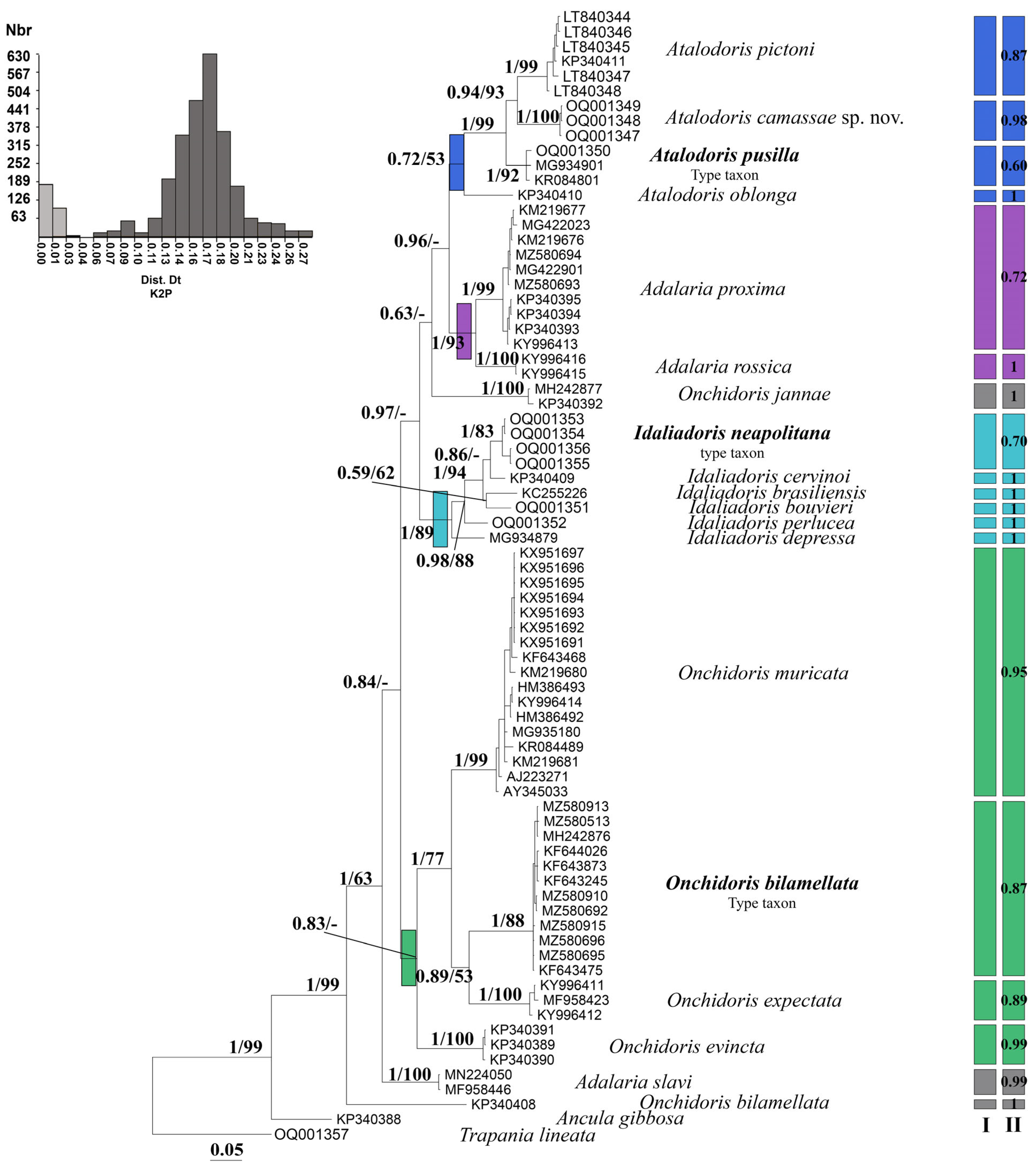
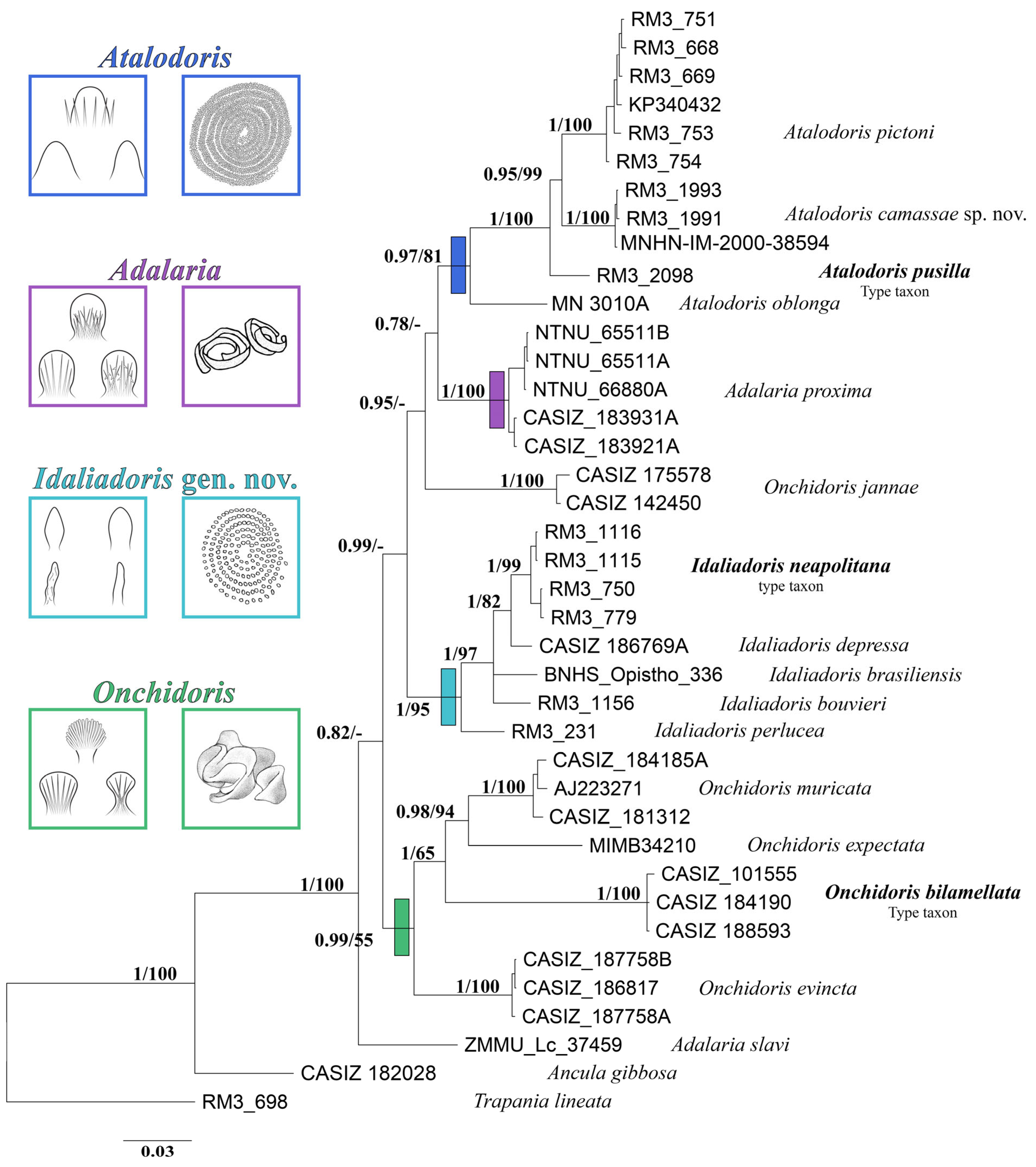
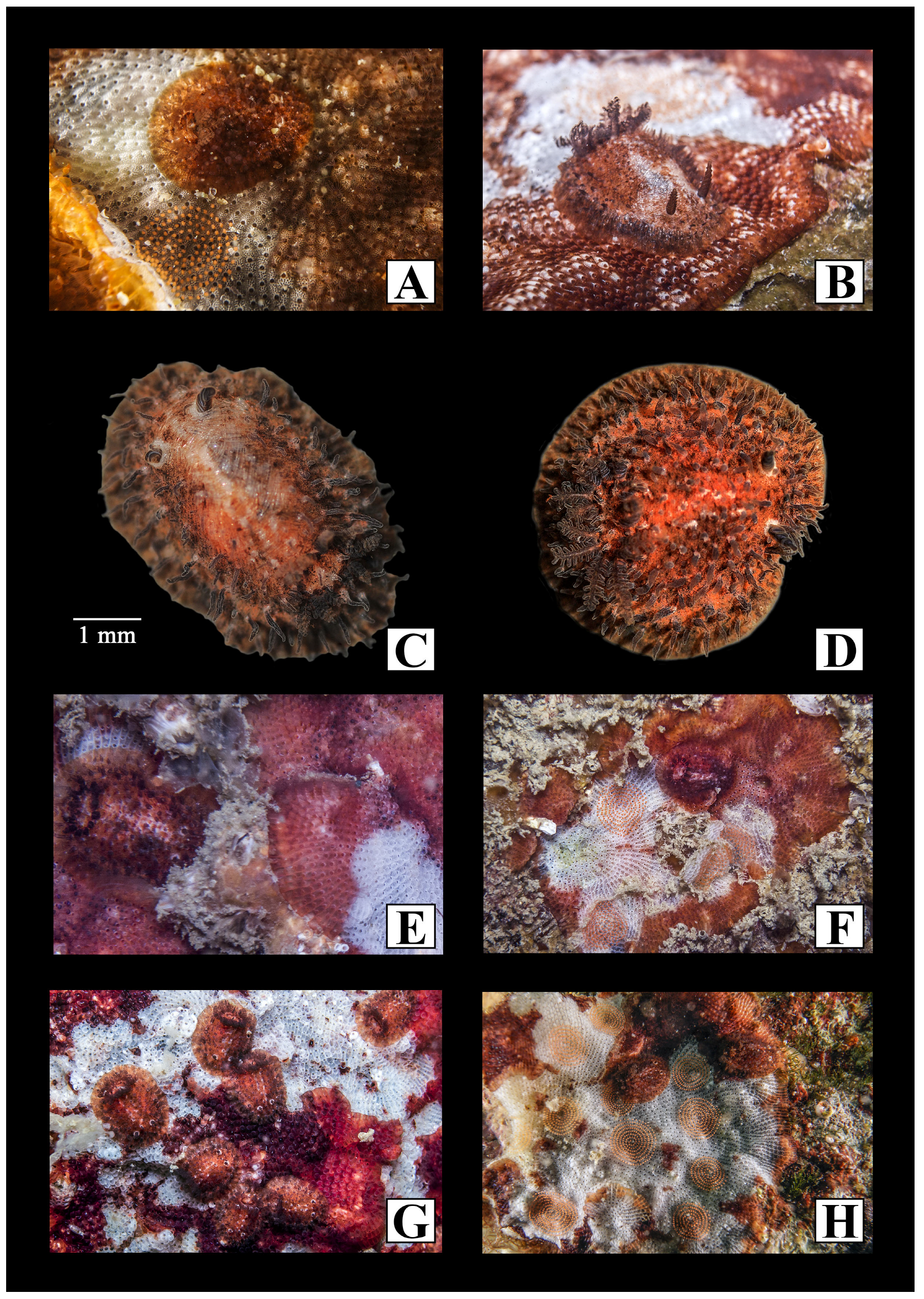
3. Results
Molecular Analyses
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Ben Rais Lasram, F.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Voultsiadou, E.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moullec, F.; Barrier, N.; Drira, S.; Guilhaumon, F.; Marsaleix, P.; Somot, S.; Ulses, C.; Velez, L.; Shin, Y.J. An end-to-end model reveals losers and winners in a warming Mediterranean Sea. Front. Mar. Sci. 2019, 345, 1–19. [Google Scholar] [CrossRef]
- Renda, W.; Amati, B.; Bogi, C.; Bonomolo, G.; Capua, D.; Dell’Angelo, B.; Giulia, F.; Riccardo, G.S.; Rafael, L.P.; Oliverio, M.; et al. The new Checklist of the Italian Fauna: Marine Mollusca. Biogeogr. J. Integr. Biogeogr. 2022, 37, 1–14. [Google Scholar] [CrossRef]
- Strong, E.E.; Gargominy, O.; Ponder, W.F.; Bouchet, P. Global Diversity of Gastropods (Gastropoda; Mollusca) in Freshwater. Hydrobiologia 2008, 595, 149–166. [Google Scholar] [CrossRef]
- Brown, K.M. 10—Mollusca: Gastropoda. In Ecology and Classification of North American Freshwater Invertebrates, 2nd ed.; Thorp, J.H., Covich, A.P., Eds.; Academic Press: San Diego, CA, USA, 2001; pp. 297–329. ISBN 978-0-12-690647-9. [Google Scholar]
- Furfaro, G.; Vitale, F.; Licchelli, C.; Mariottini, P. Two Seas for One Great Diversity: Checklist of the Marine Heterobranchia (Mollusca; Gastropoda) from the Salento Peninsula (South-East Italy). Diversity 2020, 12, 171. [Google Scholar] [CrossRef]
- Chimienti, G.; Angeletti, L.; Furfaro, G.; Canese, S.; Taviani, M. Habitat, morphology and trophism of Tritonia callogorgiae sp. nov., a large nudibranch inhabiting Callogorgia verticillata forests in the Mediterranean Sea. Deep Sea Res. Part I Oceanogr. Res. Pap. 2020, 165, 103364. [Google Scholar] [CrossRef]
- Canessa, M.; Bavestrello, G.; Cattaneo-Vietti, R.; Furfaro, G.; Doneddu, M.; Navone, A.; Trainito, E. Rocky substrate affects benthic heterobranch assemblages and prey/predator relationships. Estuar. Coast. Shelf Sci. 2021, 261, 31. [Google Scholar] [CrossRef]
- Trainito, E.; Fantin, M.; Manganelli, E.; Furfaro, G. What are you doing here? Investigating on an unexpected association in shallow Mediterranean dark caves sheds light on the diet of Marionia blainvillea (Mollusca, Gastropoda, Nudibranchia). Turk. J. Zool. 2021, 45, 550–556. [Google Scholar] [CrossRef]
- Riccardi, A.; Colletti, A.; Virgili, R.; Cerrano, C. Diversity and behavior of sea slugs (Heterobranchia) in the rocky tide pools of Conero Riviera (western Adriatic Sea). Eur. Zool. J. 2022, 89, 856–869. [Google Scholar] [CrossRef]
- MolluscaBase eds. 2022. MolluscaBase. Onchidorididae Gray, 1827. World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=175 (accessed on 24 November 2022).
- MolluscaBase eds. 2022. MolluscaBase. Acanthodoris Gray, 1850. World Register of Marine Species. Available online: https://www.marinespecies.org/aphia.php?p=taxdetails&id=138284 (accessed on 24 November 2022).
- Hallas, J.M.; Simison, W.B.; Gosliner, T.M. Dating and biogeographical patterns in the sea slug genus Acanthodoris Gray, 1850 (Mollusca, Gastropoda, Nudibranchia). Mol. Phylogenetics Evol. 2016, 97, 19–31. [Google Scholar] [CrossRef]
- Hallas, J.M.; Gosliner, T.M. Family matters: The first molecular phylogeny of the Onchidorididae Gray, 1827 (Mollusca, Gastropoda, Nudibranchia). Mol. Phylogenetics Evol. 2015, 88, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Martynov, A.; Sanamyan, N.; Korshunova, T. Review of the opisthobranch mollusc fauna of Russian Far Eastern seas: Pleurobranchomorpha, Doridida and Nudibranchia. Bull. Kamchatka State Tech. Univ. 2015, 34, 62–87. (In Russian) [Google Scholar]
- Iredale, T.; O’Donoghue, C.H. List of British nudibranchiate Mollusca. Proc. Malacol. Soc. Lond. 1923, 15, 195–233. [Google Scholar]
- Ortea, J. Nueva especie del género Atalodoris Iredale & O’Donoghue, 1923 (Mollusca: Nudibranchia) colectada en la Concha de Artedo, Asturias, dedicada al periodista deportivo José María García. Rev. De La Acad. Canar. De Cienc. 2021, 33, 97–106. [Google Scholar]
- Schmekel, L.; Portmann, A. Opisthobranchia des Mittelmeeres; Springer Science & Business Media: Berlin, Germany, 1982. [Google Scholar]
- Furfaro, G.; Mariottini, P. A New Dondice Marcus Er. 1958 (Gastropoda: Nudibranchia) from the Mediterranean Sea Reveals Interesting Insights into the Phylogenetic History of a Group of Facelinidae Taxa. Zootaxa 2020, 4731, 1–22. [Google Scholar] [CrossRef]
- Furfaro, G.; Schreier, C.; Trainito, E.; Pontes, M.; Madrenas, E.; Girard, P.; Mariottini, P. The Sea Slug Doriopsilla areolata Bergh, 1880 (Mollusca, Gastropoda) in the Mediterranean Sea: Another Case of Cryptic Diversity. Diversity 2022, 14, 297. [Google Scholar] [CrossRef]
- Martynov, A.; Korshunova, T. World’s northenmost and rarely observed nudibranchs: Three new onchidoridid species (Gastropoda: Doridida) from Russian seas. Zootaxa 2017, 4299, 391–404. [Google Scholar] [CrossRef]
- Furfaro, G.; Trainito, E. A new species from the Mediterranean Sea and North-Eastern Atlantic Ocean: Knoutsodonta pictoni n. sp. (Gastropoda Heterobranchia Nudibranchia). Biodivers. J. 2017, 8, 725–738. [Google Scholar]
- Aljanabi, S.M.; Martinez, I. Universal and Rapid Salt-Extraction of High Quality Genomic DNA for PCR-Based Techniques. Nucleic Acids Res. 1997, 25, 4692–4693. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA Primers for Amplification of Mitochondrial Cytochrome c Oxidase Subunit I from Diverse Metazoan Invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Palumbi, S.R.; Martin, A.; Romano, S.; McMillan, W.O.; Stice, L.; Grabowski, G. The Simple Fool’s Guide to PCR; Dept. of Zoology and Kewalo Marine Laboratory, University of Hawaii: Honolulu, HI, USA, 2002. [Google Scholar]
- Colgan, D.J.; McLauchlan, A.; Wilson, G.D.F.; Livingston, S.P.; Edgecombe, G.D.; Macaranas, J.; Cassis, G.; Gray, M.R. Histone H3 and U2 SnRNA DNA Sequences and Arthropod Molecular Evolution. Aust. J. Zool. 1999, 46, 419–437. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Posada, D. JModelTest: Phylogenetic Model Averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef]
- Rozas, J.; Ferrer-Mata, A.; Sánchez-DelBarrio, J.C.; Guirao-Rico, S.; Librado, P.; Ramos-Onsins, S.E.; Sánchez-Gracia, A. DnaSP 6: DNA Sequence Polymorphism Analysis of Large Data Sets. Mol. Biol. Evol. 2017, 34, 3299–3302. [Google Scholar] [CrossRef] [PubMed]
- Puillandre, N.; Lambert, A.; Brouillet, S.; Achaz, G. ABGD, Automatic Barcode Gap Discovery for Primary Species Delimitation. Mol. Ecol. 2012, 21, 1864–1877. [Google Scholar] [CrossRef]
- Puillandre, N.; Modica, M.V.; Zhang, Y.; Sirovich, L.; BOISSELIER, M.-C.; Cruaud, C.; Holford, M.; Samadi, S. Large-Scale Species Delimitation Method for Hyperdiverse Groups. Mol. Ecol. 2012, 21, 2671–2691. [Google Scholar] [CrossRef]
- Meier, R.; Shiyang, K.; Vaidya, G.; Ng, P.K.L. DNA Barcoding and Taxonomy in Diptera: A Tale of High Intraspecific Variability and Low Identification Success. Syst. Biol. 2006, 55, 715–728. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Kapli, P.; Pavlidis, P.; Stamatakis, A. A General Species Delimitation Method with Applications to Phylogenetic Placements. Bioinformatics 2013, 29, 2869–2876. [Google Scholar] [CrossRef] [Green Version]
- Ronquist, F.; Teslenko, M.; van der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian Phylogenetic Inference and Model Choice Across a Large Model Space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef] [Green Version]
- Silvestro, D.; Michalak, I. RaxmlGUI: A Graphical Front-End for RAxML. Org. Divers. Evol. 2012, 12, 335–337. [Google Scholar] [CrossRef]
- Stamatakis, A. RAxML Version 8: A Tool for Phylogenetic Analysis and Post-Analysis of Large Phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šarić, T.; Župan, I.; Aceto, S.; Villari, G.; Palić, D.; De Vico, G.; Carella, F. Epidemiology of noble pen shell (Pinna nobilis l. 1758) mass mortality events in Adriatic Sea is characterised with rapid spreading and acute disease progression. Pathogens 2020, 9, 776. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, G.; Salvi, D.; Mancini, E.; Mariottini, P. A Multilocus View on Mediterranean Aeolid Nudibranchs (Mollusca): Systematics and Cryptic Diversity of Flabellinidae and Piseinotecidae. Mol. Phylogenet. Evol. 2018, 118, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Furfaro, G.; Salvi, D.; Trainito, E.; Vitale, F.; Mariottini, P. When Morphology Does Not Match Phylogeny: The Puzzling Case of Two Sibling Nudibranchs (Gastropoda). Zool. Scr. 2021, 50, 439–454. [Google Scholar] [CrossRef]
- Bergh, L.S.R. Malacologische Untersuchungen. In Reisen im Archipel der Philippinen; Semper, C. (1868). II Theil. Wissenschaftliche Resultäte, 1. Available online: https://curiosity.lib.harvard.edu/expeditions-and-discoveries/catalog/38-990041866000203941 (accessed on 25 December 2022).
- Pruvot-Fol, A. Mollusques Opisthobranches. Faune De Fr. Paris 1954, 58, 1–460. [Google Scholar]
- Perrone, A.S. Opistobranchi (Aplysiomorpha, Pleurobrancomorpha, Sacoglossa Nudibranchia) del litorale Salentino (Mare Jonio) (Elenco-contrib. secondo). Thalass. Salentina 1986, 16, 19–42. [Google Scholar]
- Ballesteros, M.; Madrenas, E.; Pontes, M. “Knoutsodonta albonigra“ a OPK-Opistobranquis. 2022. Available online: https://opistobranquis.info/ca/1HCSO (accessed on 25 November 2022).
- Betti, F.; Bava, S.; Cattaneo-Vietti, R. Composition and seasonality of a heterobranch assemblage in a sublittoral, unconsolidated, wave-disturbed community in the Mediterranean Sea. J. Molluscan. Stud. 2017, 83, 325–332. [Google Scholar] [CrossRef]
- Ballesteros, M.; Madrenas, E.; Pontes, M. “Knoutsodonta pusilla” in OPK-Opistobranchs, 2022. Available online: https://opistobranquis.info/ca/JuGvP (accessed on 25 November 2022).
- Alder, J.; Hancock, A. Notice of a new genus and several new species of nudibranchiate Mollusca. Ann. Mag. Nat. Hist. 1845, 16, 311–316. [Google Scholar] [CrossRef] [Green Version]
- Alvim, J.; Padula, V.; Pimenta, A.D. First record of the genus Onchidoris (Gastropoda: Nudibranchia: Onchidorididae) from the South Atlantic Ocean, with the description of a new species from Brazil. J. Mar. Biol. Assoc. UK 2011, 91, 505–511. [Google Scholar] [CrossRef]
- Verrill, A.E. Additions to the fauna of the Bermudas from the Yale Expedition of 1901, with notes on other species. Trans. Conn. Acad. Arts Sci. 1901, 11, 15–62. [Google Scholar]
- Martynov, A.V. Marine and Brackish Water Gastropoda of Russia and Adjacent Countries: An Illustrated Catalogue; Kantor, Y.I., Sysoev, A.V., Eds.; KMK Scientific Press Ltd.: Moscow, Russia, 2006; 372p. [Google Scholar]
- Abraham, P.S. Revision of the anthobranchiate nudibranchiate Mollusca, with descriptions or notices of forty-one hitherto undescribed species. Proc. Zool. Soc. Lond. Year 1877, 196, 27–30. [Google Scholar]
- Gould, A.A. Report on the Invertebrata of Massachusetts, 2nd ed.; Published Agreeably to an Order of the Legislature; Wright and Potter: Cambridge, UK, 1870; pp. 225–253, pls. XVI–XXII. [Google Scholar]
- McDonald, G. Bibliographia Nudibranchia. Third online edition, 2021. Available online: eScholarship.org (accessed on 25 December 2022).
- Balch, F.N. Rectification of the nomenclature of the Nudibranchiata in Lermond’s “Shells of Maine”. Nautica 1910, 23, 100–102. [Google Scholar]
- Trott, T.J. Cobscook Bay inventory: A historical checklist of marine invertebrates spanning 162 years. Northeast. Nat. 2004, 11, 261–324. [Google Scholar] [CrossRef]
- Prkić, J.; Furfaro, G.; Mariottini, P.; Carmona, L.; Cervera, J.L.; Modica, M.V.; Oliverio, M. First record of Calma gobioophaga Calado and Urgorri, 2002 (Gastropoda: Nudibranchia) in the Mediterranean Sea. Mediterr. Mar. Sci. 2014, 15, 423–428. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
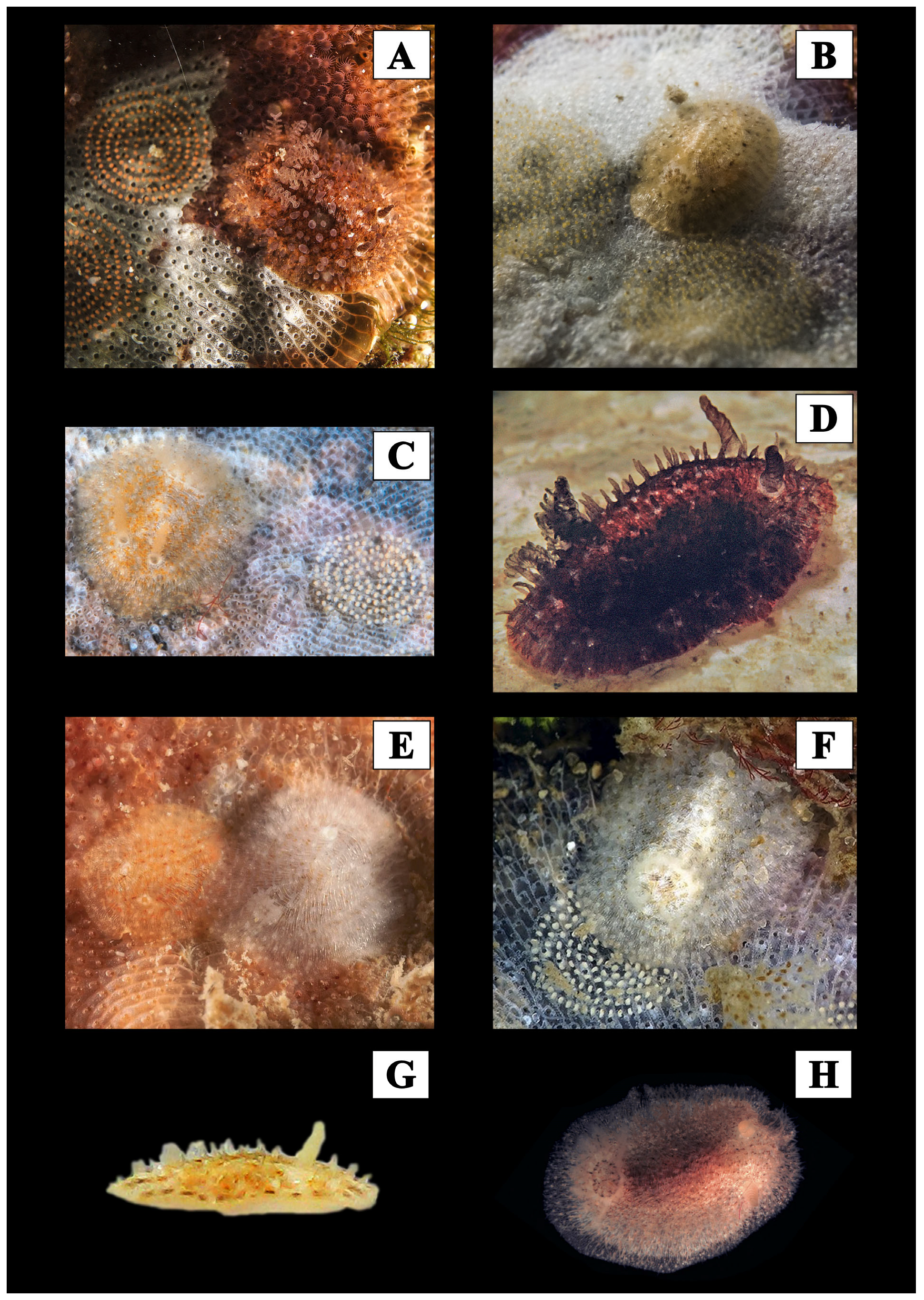{kind=link}
| Species | Voucher | Locality | COI | 16S | H3 |
|---|---|---|---|---|---|
| Adalaria proxima | CCDB22758-H04 | Canada, British Columbia, Howe Sound | MG422023 | ------------ | ------------ |
| USNM:IZ:1523836 | USA, Dutch Harbor, City Spit Dock | MZ580694 | ------------ | ------------ | |
| CCDB22758-H05 | Canada, British Columbia, Howe Sound | MG422901 | ------------ | ------------ | |
| USNM:IZ:1523835 | USA, Dutch Harbor, City Spit Dock | MZ580693 | ------------ | ------------ | |
| ZMMU:Op-596 | Russia | KY996413 | ------------ | ------------ | |
| (as Onchidoris proxima) | CASIZ 183931A * | Passamaquody Bay Eastport: Washington Co., Maine | KM219677 | KJ653674 | KM225827 |
| (as Onchidoris proxima) | CASIZ 183921A * | Passamaquody Bay Eastport: Washington Co., Maine | KM219676 | KJ653673 | KM225826 |
| (as Onchidoris loveni) | NTNU 65511B * | Norway, Mausund | KP340395 | KP340300 | ------------ |
| (as Onchidoris loveni) | NTNU 65511A * | Norway, Mausund | KP340394 | KP340299 | ------------ |
| (as Onchidoris loveni) | NTNU 66880A * | Norway, Gløssvika | KP340393 | KP340297 | ------------ |
| Adalaria rossica | ZMMU:Op-548 | Russia | KY996416 | ------------ | ------------ |
| ZMMU:Op-550 | Russia | KY996415 | ------------ | ------------ | |
| Adalaria slavi | ZMMU:Lc-37459 * | Russia | MN224050 | MN224074 | ------------ |
| MIMB34211 | Russia, Avacha Bay, Kamchatka | MF958446 | ------------ | ------------ | |
| Atalodoris camassae sp. nov. | MNHN-IM-2000-38594 * | Slovenia, Fiesa | OQ001347 | OP965547 | OQ096455 |
| RM3_1993 * | Slovenia, Fiesa | OQ001348 | OP965548 | OQ096456 | |
| RM3_1991 * | Slovenia, Fiesa | OQ001349 | OP965549 | OQ096457 | |
| Atalodoris oblonga (as Knoutsodonta oblonga) | MN 3010A * | Mewstone, Skomer | KP340410 | ------------ | KP340430 |
| Atalodoris pictoni (as Knoutsodonta pictoni) | BAU02982 * | Ireland, South of Inishgalloon, Purteen, Keel, Achill Island | LT840347 | OP965550 | OQ096458 |
| (as Knoutsodonta pictoni) | BAU02983 * | Ireland, South of Inishgalloon, Purteen, Keel, Achill Island | LT840348 | OP965551 | OQ096459 |
| (as Knoutsodonta pictoni) | BAU02985 * | Italy, Trieste, Sistiana | LT840346 | OP965552 | OQ096460 |
| (as Knoutsodonta pictoni) | BAU02986 * | Italy, Sardinia, Porto San Paolo | LT840344 | OP965553 | ------------ |
| (as Knoutsodonta pictoni) | BAU02987 * | Italy, Sardinia, Porto San Paolo | LT840345 | OP965554 | ------------ |
| (as Knoutsodonta pictoni) | CASIZ 208194 * | Scotland | KP340411 | KP340316 | KP340432 |
| Atalodoris pusilla | RM3_2098 * | Spain, Costa Brava, Girona, Punta del Ferro | OQ001350 | OP965555 | OQ096461 |
| (as Knoutsodonta pusilla) | Gastr 8972V | Sweden, Skagerrak | MG934901 | ------------ | ------------ |
| (as Onchidoris bilamellata) | MT09252 | North Sea | KR084801 | ------------ | ------------ |
| Idaliadoris brasiliensis (as Onchidoris brasiliensis) | BNHS-Opistho-336 * | ------------------------------------------ | KC255226 | KC255225 | ------------ |
| Idaliadoris bouvieri | RM3_1156 * | Croatia, Rijeka | OQ001351 | ------------ | OQ096462 |
| Idaliadoris cervinoi (as Knoutsodonta depressa) | CASIZ 186769A * | Spain, Huelva | KP340409 | KP340315 | KP340428 |
| Idaliadoris depressa (as Knoutsodonta depressa) | Gastr 8969V | Sweden, Skagerrak | MG934879 | ------------ | ------------ |
| Idaliadoris neapolitana | RM3_1116 * | Italy, Sardinia, Gulf of Olbia, Lu Carragioni | OQ001353 | OP965557 | OQ096464 |
| RM3_1115 * | Italy, Sardinia, Gulf of Olbia, Lu Carragioni | OQ001354 | ------------ | OQ096465 | |
| RM3_750 * | Italy, Trieste, Sistiana | OQ001355 | OP965558 | ------------ | |
| RM3_779 * | Italy, Sardinia, Porto San Paolo | OQ001356 | OP965559 | OQ096466 | |
| Idaliadoris perlucea | RM3_231 * | Italy, Latium, M.P.A. Secche di Tor Paterno | OQ001352 | OP965556 | OQ096463 |
| Onchidoris bilamellata | USNM:IZ:1503309 | USA, Dutch Harbor; City Spit Dock | MZ580513 | ------------ | ------------ |
| BMBM-0089 | USA, Washington, San Juan County, San Juan Island, Snug Harbor | MH242876 | ------------ | ------------ | |
| USNM:IZ:1529241 | USA, Dutch Harbor | MZ580910 | ------------ | ------------ | |
| USNM:IZ:1523834 | USA, Captains Bay; North Pacific Fuel Dock | MZ580692 | ------------ | ------------ | |
| USNM:IZ:1529283 | USA, Dutch Harbor | MZ580915 | ------------ | ------------ | |
| USNM:IZ:1523838 | USA, Captains Bay; Little South America | MZ580696 | ------------ | ------------ | |
| USNM:IZ:1523837 | USA, Dutch Harbor; City Spit Dock | MZ580695 | ------------ | ------------ | |
| (as Onchidoris muricata) | USNM:IZ:1529244 | USA, Dutch Harbor | MZ580913 | ------------ | ------------ |
| CASIZ_101555 * | California, Peninsula Point, Marin Co. | KP340408 | KP340312 | ------------ | |
| CASIZ 184190 * | New Hampshire, Rockingham Co. | ------------ | KP340313 | KP340426 | |
| 10NBMOL-10020 | Canada, New Brunswick, St. Andrews, Passamaquoddy Bay | KF644026 | ------------ | ------------ | |
| 10NBMOL-10019 | Canada, New Brunswick, St. Andrews, Passamaquoddy Bay | KF643873 | ------------ | ------------ | |
| 10BCMOL-00203 | Canada, British Columbia, Bamfield, Wizard Islet | KF643475 | ------------ | ------------ | |
| 10NBMOL-10018 | Canada, New Brunswick, St. Andrews, Passamaquoddy Bay | KF643245 | ------------ | ------------ | |
| CASIZ 188593 * | Washington, Kitsap Co., Puget Sound | ------------ | KP340314 | KP340427 | |
| Onchidoris evincta | CASIZ 187758B* | Washington, Kitsap Co., Puget Sound | KP340391 | KP340294 | ------------ |
| CASIZ 186817 * | Washington, Kitsap Co., Puget Sound | KP340389 | KP340292 | ------------ | |
| CASIZ 187758A* | USA, Washington, Kitsap County, Puget Sound | KP340390 | KP340293 | ------------ | |
| Onchidoris expectata | ZMMU:Op-595 | Russia | KY996411 | ------------ | ------------ |
| ZMMU:Op-593 | Russia | KY996412 | ------------ | ------------ | |
| (as Onchidoris macropompa) | MIMB34210 * | Russia, Avacha Bay, Kamchatka | MF958423 | MF958292 | ------------ |
| Onchidoris jannae (as Knoutsodonta jannae) | CASIZ 175578 * | California, San Mateo Co., Pillar Point | KP340392 | KP340296 | KP340415 |
| (as Knoutsodonta jannae) | CASIZ 142450 * | Alaska, Prince William Sound | ------------ | KP340295 | KP340414 |
| Onchidoris muricata | BMBM-0550 | USA, Washington, San Juan County, San Juan Island, FHL Dock | MH242877 | ------------ | ------------ |
| AC19-30 | Russia, Nevelsk, Sakhalin Is., Sea of Japan | KX951697 | ------------ | ------------ | |
| AC19-32 | Russia, Kievka Bay, Sea of Japan | KX951696 | ------------ | ------------ | |
| AC19-31 | Russia, Kievka Bay, Sea of Japan | KX951695 | ------------ | ------------ | |
| AC17-40 | Russia, Rudnaya Bay, Sea of Japan | KX951694 | ------------ | ------------ | |
| AC17-24 | Russia, Rudnaya Bay, Sea of Japan | KX951693 | ------------ | ------------ | |
| AC16-19 | Russia, Rudnaya Bay, Sea of Japan | KX951692 | ------------ | ------------ | |
| AC16-18 | Russia, Rudnaya Bay, Sea of Japan | KX951691 | ------------ | ------------ | |
| BIOUG < CAN >: WS130 | Russia | HM386493 | ------------ | ------------ | |
| ZMMU:Op-597 | Russia | KY996414 | ------------ | ------------ | |
| BIOUG < CAN >: WS125 | Russia | HM386492 | ------------ | ------------ | |
| Gastr 8524V | Sweden, Kattegatt | MG935180 | ------------ | ------------ | |
| ------------ | Scotland, Clachan Seil | AY345033 | ------------ | ------------ | |
| MT07703 | North Sea | KR084489 | ------------ | ------------ | |
| CASIZ 184185A * | New Hampshire, Rockingham Co., New Castle Portsmouth Bay | KM219681 | KJ653677 | KM225831 | |
| CASIZ 181312 * | California, Monterey Co., Asilomar | KM219680 | KJ653676 | KM225830 | |
| 10BCMOL-00318 | Canada, British Columbia, Haida Gwaii, Juskalta Narrows | KF643468 | ------------ | ------------ | |
| ------------ * | Sweden, Bohuslan, Kristineberg | AJ223271 | AJ225196 | ------------ | |
| Ancula gibbosa | CASIZ 182028 * | USA, Cumberland County, Maine | KP340388 | KP340291 | KP340413 |
| Trapania lineata | RM3_698 * | Italy, Sardinia, Secca La Mandria | OQ001357 | OP965560 | OQ096467 |
| Species Attributed to the Genus Adalaria Bergh 1878 | ||||||
|---|---|---|---|---|---|---|
| Species | Body | Rhinophores | Gills | Radular Formula | Distribution | Ontogenesis N Capsules |
| Adalaria loveni (Alder & Hancock, 1862) | White, sometimes pale yellow | White | White | 2–46 × 12–13.1.1.1.13–12 | Great Britain, W. Scotland, to Norway | ? |
| Adalaria proxima (Alder & Hancock, 1854) | White to yellow orange | Same as body | Same as body | 39–50 × 13–9. 1.1.1.9–13 | Amphiboreal: North Atlantic, North Pacific | P 2470 |
| Adalaria olgae Martynov, Korshunova, Sanamyan & Sanamyan, 2009 | Intense lemon yellow, invariable | Semitransparent white | Semitransparent white | 30-31 × 3–4.1.1.1.4–3 | Northwest Pacific | ? |
| Adalaria rossica Martynov & Korshunova, 2017 | Yellowish white, inconspicuous white dots | Yellowish | Yellowish | 46 × 1–10.1.1.1.10–1 | Arctic Ocean | ? |
| Adalaria tschuktschica Krause, 1885 | Yellowish white when preserved | Yellowish white when preserved | Yellowish white when preserved | 30–32 × 6–8.1.1.1.8–6 | Arctic Ocean to Northern Bering Sea, NW Pacific | ? |
| Adalaria ultima Martynov & Korshunova, 2017 | Yellowish white, inconspicuous white dots | Yellowish | Yellowish | 37 × 1–10.1.1.1.10–1 | Arctic Ocean, Okhotsk Sea | ? |
| Species Attributed to the Genus Atalodoris | ||||||
|---|---|---|---|---|---|---|
| Species | Body | Rhinophores | Gills | Radular Formula | Distribution | Ontogenesis N° Capsules |
| Atalodoris camassae sp. nov. | Hyaline off-white with a thick covering of dark irregular brown spots (salt and pepper) with a lighter brown halo. Central area darker in correspondence with internal organs, ending around the gills and with a pointed end between the rhinophores | Hyaline spine with dark lamellae | Hyaline in the internal part, darker and spotted in the external portion | ? × 1.1.0.1.1 | Mediterranean endemic? | P 1400 |
| Atalodoris butanito Ortea, 2021 | Orange with reddish brown speckles that are less sparse in the central region; the entire edge of the mantle is blotched brown, with a continuous series of spicules running perpendicular to the edge | Yellowish with dark leaflets | Yellowish | 48 × 1.1.0.1.1 | Northeast Atlantic | P 3000 eggs × cm |
| Atalodoris inconspicua (Alder & Hancock, 1851) | White or pale brown, often tinged with a purple hue, covered by small specks of brown pigment | Pale yellow | White | 29 × 1.1.0.1.1 | Northeast Atlantic | P |
| Atalodoris oblonga (Alder & Hancock,1845) | Grey with a few darker blotches on the back | Yellowish | Yellowish white | 28 × 1.1.0.1.1 | Northeast Atlantic | P |
| Atalodoris pictoni (Furfaro & Trainito, 2017) | Uniform dark brown, almost black, with small bluish white speckles scattered along the mantle | White | Dark | 25–28 × 1.1.0.1.1 | Northeast Atlantic, Mediterranean | p > 9000 |
| Atalodoris pusilla (Alder & Hancock,1845) | Dense dark brown pigment spots responsible for dark appearance | Transparent | Transparent | 21–29 × 1.1.0.1.1 | Northeast Atlantic, Mediterranean | P |
| Atalodoris reticulata (Ortea,1979) | Whitish, intensely pigmented in green at center, fading at borders; whitish longitudinal bands forming a net with bands running from tubercles surrounding brown and green patches | Large brown stains | Pigmented dirt cream | 51–54 × 1.1.0.1.1 | Northeast Atlantic | P |
| Atalodoris sparsa (Alder & Hancock,1846) | Pale brown mantle with regularly spaced, darker blotches forming patches | Blotched with olive-brown | Colorless | 32–36 × 1.1.0.1.1 | Northeast Atlantic, Mediterranean | P |
| Species Attributed to the Genus Idaliadoris gen. nov. | ||||||
|---|---|---|---|---|---|---|
| Species | Body | Rhinophores | Gills | Radular Formula | Distribution | Ontogenesis N° Capsules |
| Idaliadoris bouvieri comb. nov. (Vayssieère,1919) | Pale translucent pink with scattered red brown patches distributed in three longitudinal bands | Yellowish pink | Yellowish pink | 40 × 1.1.0.1.1 | Med. endemic? | L 240 |
| Idaliadoris brasiliensis comb. nov. (Alvim, Padula & Pimenta, 2011) | Greyish white or yellowish orange, both translucent, with a pattern of scattered dark brown (sometimes orange) tiny spots, tending towards the mid-line | Translucent white | Translucent white | 16–17 × 1.1.0.1.1 | Southeast Atlantic | L 116 |
| Idaliadoris cervinoi comb. nov. (Ortea & Urgorri, 1979) | Orange, centrally intense, fading at the borders; some specimens’ borders almost whitish | Orange with dark brown lamellae | Dark brown circled by a light area | ? × 1.0.1 | Northeast Atlantic | L |
| Idaliadoris depressa comb. nov. (Alder & Hancock, 1842) | Pale or translucent brown with scattered orange or purple-brown speckles | Translucent | Translucent | 33–34 × 1.1.0.1.1 | Northeast Atlantic, Mediterranean | L 354 |
| Idaliadoris maugeansis comb. nov. (Burn, 1958) | Pale pattern with shades or colors of yellow and/or orange | Yellowish | Yellowish | 22 rows | Southwest Pacific | L 129 |
| Idaliadoris neapolitana comb. nov. (Delle Chiaie, 1841) | Pale reddish or pale brown covered with dense and intense red or purple/brown; at the notal edge, pigment becomes linear | Dark | Dark | 24 × 1.1.0.1.1 | Mediterranean endemic | L 212 |
| Idaliadoris perlucea comb. nov. (Ortea & Moro, 2014) | Transparent with brown speckles | Translucent white | Translucent | ? | Northeast Atlantic, Mediterranean | L 354 |
| Idaliadoris tridactila comb. nov. (Ortea & Ballesteros, 1982) | Whitish with orange or dark reddish spots; in between rhinophores and gills, up to six irregular lines formed by this pigment | Yellow or light pink | Yellow | 12 × 1-1-0-1-1 | Northeast Atlantic | L 354 |
| Species Attributed to the Genus Onchidoris | ||||||
|---|---|---|---|---|---|---|
| Species | Body | Rhinophores | Gills | Radular Formula | Distribution | Ontogenesis N° Capsules |
| Onchidoris bilamellata (L., 1767) | Light brown or yellowish grounds with numerous dark brown spots | Dark at the base lighter on top | Dark at the base lighter on top | 23–30 × 1.1.1.1.1 | Amphiboreal | P 60.000 |
| Onchidoris evincta Millen, 2006 | White, rarely yellowish | White or yellowish | White or yellowish | 4–39 × 3–6.1.1.1.6–3 | Northeast Pacific, British Columbia | ? |
| Onchidoris expectata Martynov & Korshunova, 2017 | Opaque white | White | White | 35–38 × 1.1.1.1.1 | Northwest Pacific | ? |
| Onchidoris hystricina (Bergh, 1878) | Uniformly whitish shining spicules | Whitish | Whitish | 40 × | East Pacific (Alaska + California) | ? |
| Onchidoris macropompa Martynov, Korshunova, Sanamyan & Sanamyan, 2009 | Off-white transparent | White | White | 35–38 × 1.1.1.1.1 | Northern Pacific | ? |
| Onchidoris muricata (Müller, 1776) | White to yellow | White | White | 20–36 × 1.1.1.1.1 | Amphiboreal | P 2500 |
| List of Nomina Dubia | |||||
|---|---|---|---|---|---|
| Species | Original Genus | Status | Synonymy | Distribution | References |
| Onchidoris aureopuncta (Verrill, 1901) | Lamellidoris | Nomen dubium | Possible synonym of Cadlina laevis | Central West Atlantic | / |
| Onchidoris lactea (Verrill, 1900) | Lamellidoris | Nomen dubium | ND | Central West Atlantic | [48] |
| Onchidoris miniata (Verrill, 1901) | Lamellidoris | Nomen dubium | ND | Central West Atlantic | / |
| Onchidoris olivacea (Verrill, 1900) | Lamellidoris | Nomen dubium | ND | Central West Atlantic | [48] |
| Onchidoris quadrimaculata (Verrill, 1900) | Lamellidoris | Nomen dubium | ND | Central West Atlantic (Bermuda) | / |
| Onchidoris derjugini (Volodchemko, 1941) | Adalaria | Nomen dubium | Possible senior synonym of Adalaria jannae | Sea of Japan | [49] |
| Onchidoris diademata (Agassiz, 1850) | Lamellidoris, Doris | Nomen dubium | Archidoris tuberculata | Northwest Atlantic | [50,51,52] |
| Onchidoris grisea (Stimpson, MS, Gould, 1870) | Lamellidoris | Nomen dubium | ND | Northwest Atlantic | [53] |
| Onchidoris tenella (Agassiz, 1850) (Gould, 1870) | Lamellidoris, Doris | Nomen dubium | Doridacea, according to Trott [54] | Northwest Atlantic | [53] |
| Onchidoris beringi (Volodchemko, 1941) | Lamellidoris, Adalaria | Nomen dubium | ND | North Pacific | [49] |
| Onchidoris spiculoides (Volodchemko, 1941) | Lamellidoris, Adalaria | Nomen dubium | Possible synonym of Adalaria tschuktschica | North Pacific | [49] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Furfaro, G.; Trainito, E.; Fantin, M.; D’Elia, M.; Madrenas, E.; Mariottini, P. Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species. Diversity 2023, 15, 38. https://doi.org/10.3390/d15010038
Furfaro G, Trainito E, Fantin M, D’Elia M, Madrenas E, Mariottini P. Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species. Diversity. 2023; 15(1):38. https://doi.org/10.3390/d15010038
Chicago/Turabian StyleFurfaro, Giulia, Egidio Trainito, Marco Fantin, Marcella D’Elia, Enric Madrenas, and Paolo Mariottini. 2023. "Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species" Diversity 15, no. 1: 38. https://doi.org/10.3390/d15010038
APA StyleFurfaro, G., Trainito, E., Fantin, M., D’Elia, M., Madrenas, E., & Mariottini, P. (2023). Mediterranean Matters: Revision of the Family Onchidorididae (Mollusca, Nudibranchia) with the Description of a New Genus and a New Species. Diversity, 15(1), 38. https://doi.org/10.3390/d15010038