
A Solemys Skull from the Late Cretaceous of Southern France


 ,
, 

Abstract
:1. Introduction
2. Geological Setting
3. Systematic Palaeontology
3.1. Description and Comparisons
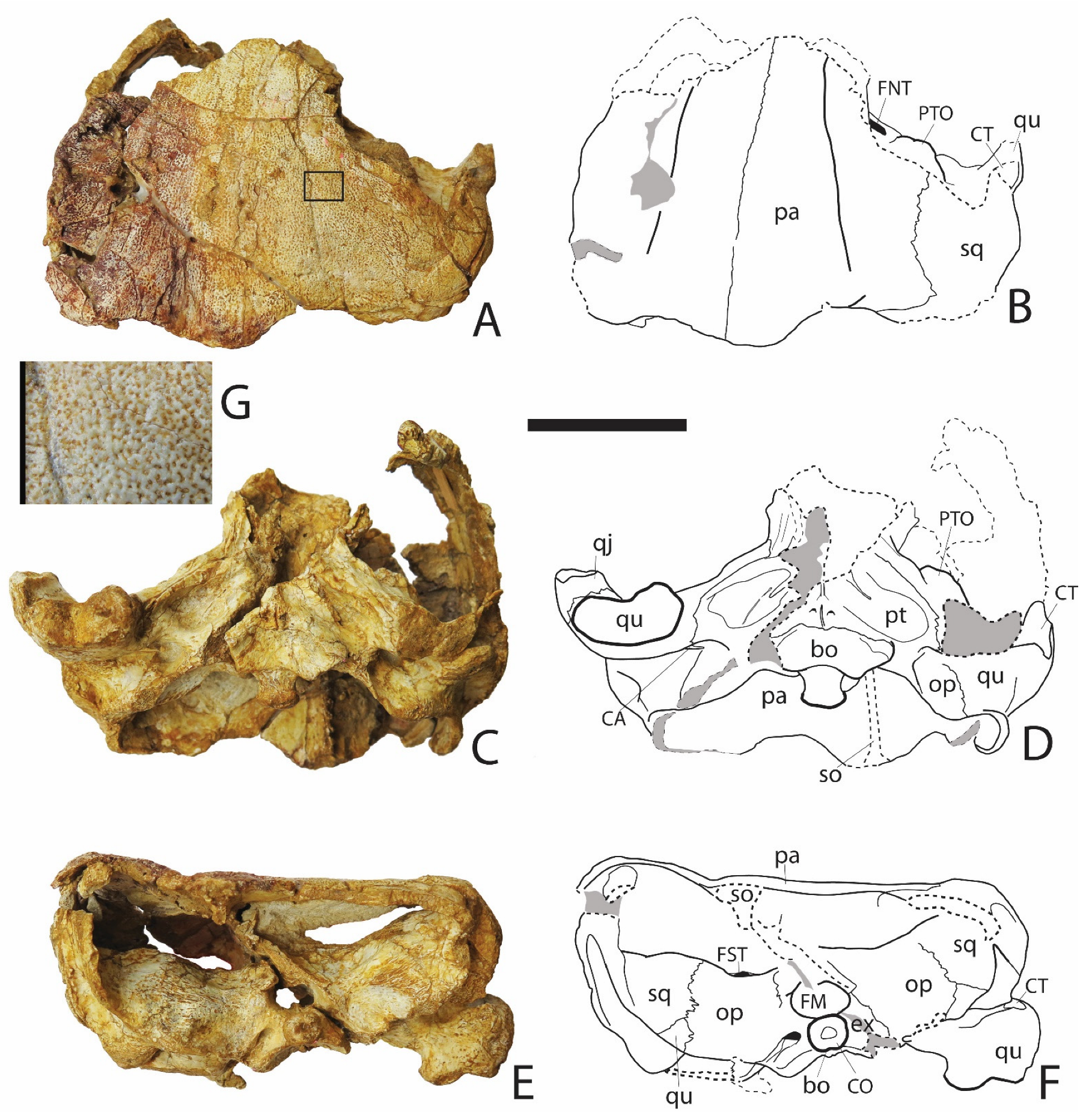
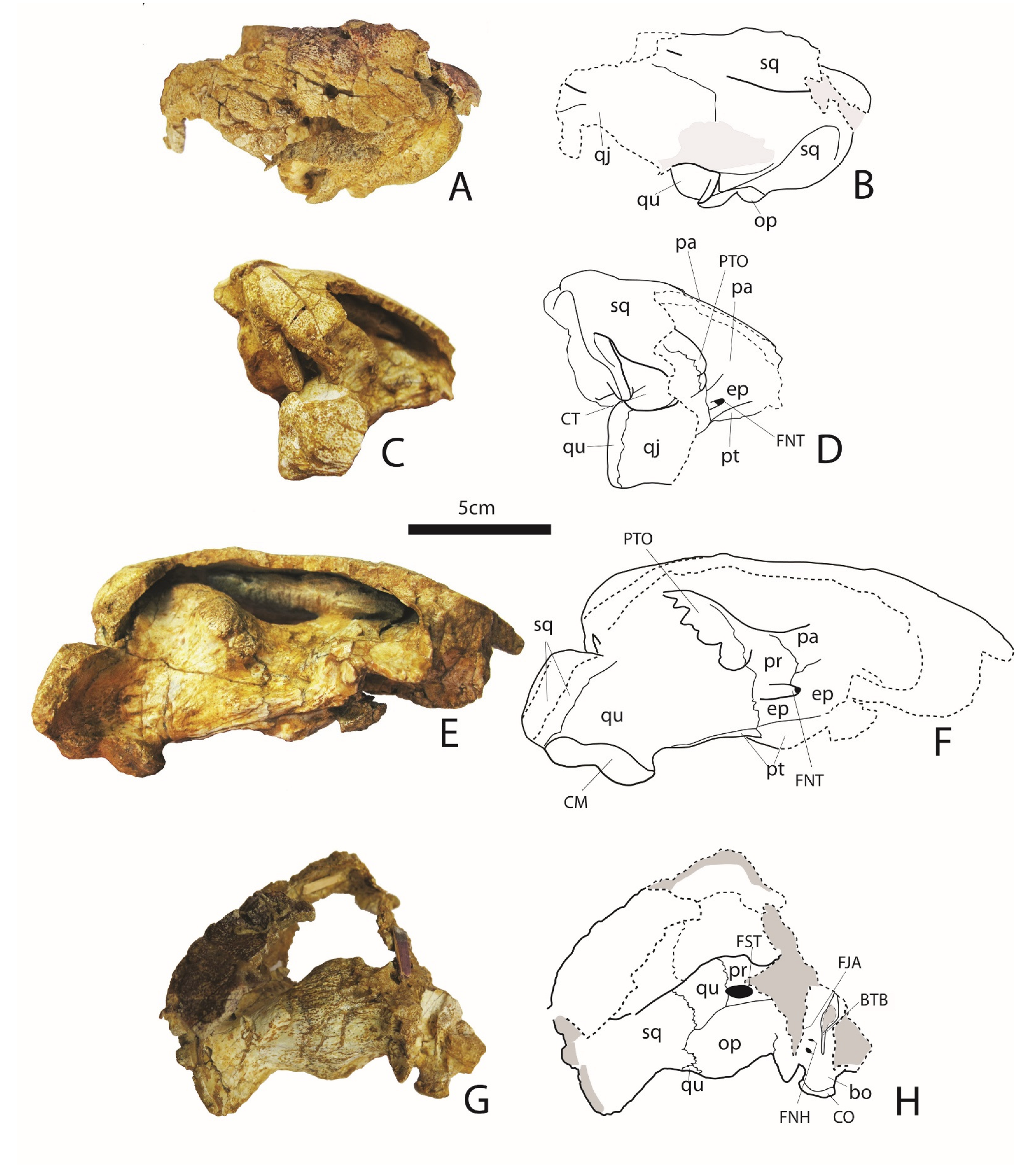
3.1.1. Skull (Figure 1, Figure 2 and Figure 3)
3.1.2. Shell (Figure 4)
4. Phylogenetic Analyses
5. Discussion
5.1. Systematic Assignment
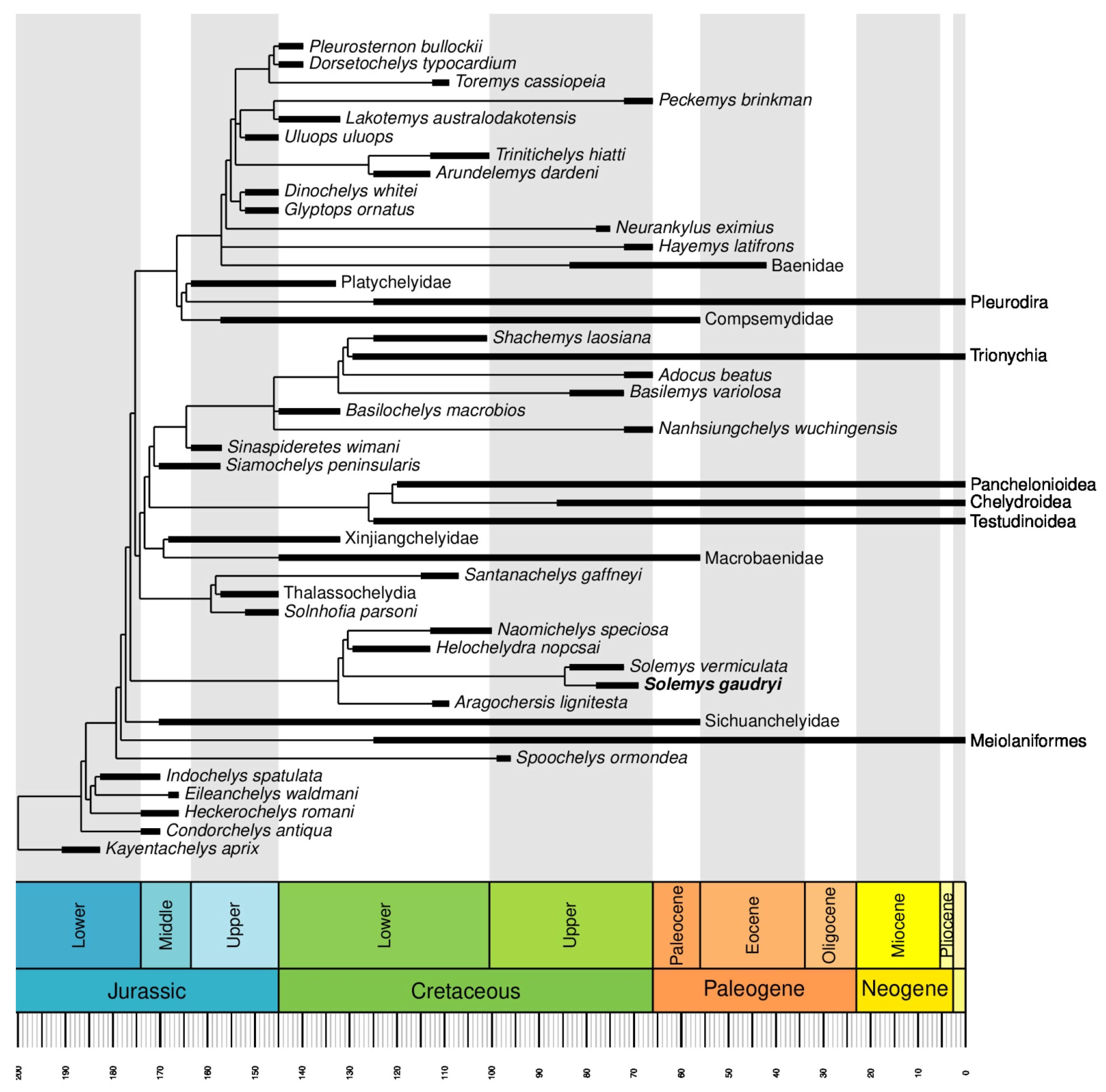
5.2. Phylogenetic Relationships of Solemys
5.3. Relationships of Helochelydridae with Other Clades and Higher Relationships
6. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joyce, W.G. A Review of the Fossil Record of Basal Mesozoic Turtles. Bull. Peabody Mus. Nat. Hist. 2017, 58, 65–113. [Google Scholar] [CrossRef] [Green Version]
- Joyce, W.G. A Review of Helochelydrid Shell Material from Late Albian to Early Cenomanian Greensands of Southern England, United Kingdom. Anat. Rec. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Pérez-García, A.; Espílez, E.; Mampel, L.; Alcalá, L. A New Basal Turtle Represented by the Two Most Complete Skeletons of Helochelydridae in Europe. Cretac. Res. 2020, 107, 104291. [Google Scholar] [CrossRef]
- Joyce, W.G.; Chapman, S.D.; Moody, R.T.; Walker, C.A. The Skull of the Solemydid Turtle Helochelydra nopcsai from the Early Cretaceous of the Isle of Wight (UK) and a Review of Solemydidae. Spec. Pap. Palaeontol. 2011, 86, 75–97. [Google Scholar]
- Joyce, W.G.; Sterli, J.; Chapman, S.D. The Skeletal Morphology of the Solemydid Turtle Naomichelys speciosa from the Early Cretaceous of Texas. J. Paleontol. 2014, 88, 1257–1287. [Google Scholar] [CrossRef] [Green Version]
- Paulina-Carabajal, A.; Sterli, J.; Werneburg, I. The Endocranial Anatomy of the Stem Turtle Naomichelys speciosa from the Early Cretaceous of North America. Acta Palaeontol. Pol. 2019, 64, 711–716. [Google Scholar] [CrossRef]
- Tortosa, T. Vertébrés Continentaux Du Crétacé Supérieur de Provence (Sud-Est de La France). PhD Thesis, Paris 6, Paris, France, 2014. [Google Scholar]
- Buffetaut, E.; Le Loeuff, J.; Mechin, P.; Mechin-Salessy, A. A Large French Cretaceous Bird. Nature 1995, 377, 110. [Google Scholar] [CrossRef]
- Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. An Archaic Bird (Enantiornithes) from the Upper Cretaceous of Provence (Southern France). Comptes Rendus Académie Sci. Ser. IIA-Earth Planet. Sci. 2000, 331, 557–561. [Google Scholar] [CrossRef]
- Buffetaut, E.; Mechin, P.; Mechin-Salessy, A. An Azhdarchid Pterosaur from the Upper Cretaceous of Provence (Southern France). In Mesozoic and Cenozoic vertebrates and paleoenvironments, tributes the career of Professor Dan Grigorescu; Editura Ars Docendi: Bucharest, Romania, 2006; pp. 95–100. [Google Scholar]
- Buffetaut, E.; Angst, D.; Mechin, P.; Mechin-Salessy, A. New Remains of the Giant Bird Gargantuavis philoinos from the Late Cretaceous of Provence (South-Eastern France). Palaeovertebrata 2015, 39, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Buffetaut, E.; Angst, D.; Mechin, P.; Mechin-Salessy, A. A Femur of the Giant Bird Gargantuavis from the Late Cretaceous of Var (South-Eastern France). Carnets Nat. 2019, 6, 47–52. [Google Scholar]
- Le Loeuff, J.; Buffetaut, E. A New Dromaeosaurid Theropod from the Upper Cretaceous of Southern France. Oryctos 1998, 1, 105–112. [Google Scholar]
- Tong, H.; Gaffney, E.S.; Buffetaut, E. Foxemys, a New Side-Necked Turtle (Bothremydidae, Pelomedusoides) from the Late Cretaceous of France. Am. Mus. Novit. 1998, 3251. [Google Scholar]
- Martin, J.E. New Material of the Late Cretaceous Globidontan Acynodon iberoccitanus (Crocodylia) from Southern France. J. Vertebr. Paleontol. 2007, 27, 362–372. [Google Scholar] [CrossRef]
- Cavin, L.; Buffetaut, E.; Dutour, Y.; Garcia, G.; Le Loeuff, J.; Méchin, A.; Méchin, P.; Tong, H.; Tortosa, T.; Turini, E. The Last Known Freshwater Coelacanths: New Late Cretaceous Mawsoniid Remains (Osteichthyes: Actinistia) from Southern France. PLoS ONE 2020, 15, e0234183. [Google Scholar] [CrossRef] [PubMed]
- Sterli, J.; Joyce, W.G. The Cranial Anatomy of the Early Jurassic Turtle Kayentachelys aprix. Acta Palaeontol. Pol. 2007, 52, 675–694. [Google Scholar]
- Gaffney, E.S. Comparative Cranial Morphology of Recent and Fossil Turtles. Bull. Am. Mus. Nat. Hist. 1979, 164, 65–376. [Google Scholar]
- Gaffney, E.S. The Jurassic Turtles of North America. Bull. Am. Mus. Nat. Hist. 1979, 162, 91–136. [Google Scholar]
- Albrecht, P.W. The Cranial Arteries and Cranial Arterial Foramina of the Turtle Genera Chrysemys, Sternotherus, and Trionyx: A Comparative Study with Analysis of Possible Evolutionary Implications. Tulane Stud. Zool. 1967, 14, 81–99. [Google Scholar]
- Albrecht, P.W. The Cranial Arteries of Turtles and Their Evolutionary Significance. J. Morphol. 1976, 149, 159–182. [Google Scholar] [CrossRef]
- Rollot, Y.; Lyson, T.R.; Joyce, W.G. A Description of the Skull of Eubaena cephalica (Hay, 1904) and New Insights into the Cranial Circulation and Innervation of Baenid Turtles. J. Vertebr. Paleontol. 2018, 38, e1474886. [Google Scholar] [CrossRef] [Green Version]
- Tong, H.; Chanthasit, P.; Naksri, W.; Ditbanjong, P.; Suteethorn, S.; Buffetaut, E.; Suteethorn, V.; Wongko, K.; Deesri, U.; Claude, J. Yakemys multiporcata n. g. n. Sp., a Large Macrobaenid Turtle from the Basal Cretaceous of Thailand, with a Review of the Turtle Fauna from the Phu Kradung Formation and Its Stratigraphical Implications. Diversity 2021, 13, 630. [Google Scholar] [CrossRef]
- Tong, H.; Tortosa, T.; Buffetaut, E.; Dutour, Y.; Turini, E.; Claude, J. A Compsemydid Turtle from the Upper Cretaceous of Var, Southern France. Ann. Paléontologie 2022. [Google Scholar] [CrossRef]
- Rollot, Y.; Evers, S.W.; Joyce, W.G. A Redescription of the Late Jurassic (Tithonian) Turtle Uluops uluops and a New Phylogenetic Hypothesis of Paracryptodira. Swiss J. Palaeontol. 2021, 140, 1–30. [Google Scholar] [CrossRef] [PubMed]
- Tong, H.; Li, L. A Revision of the Holotype of Nanhsiungchelys wuchingensis, Ye, 1966 (Testudines: Cryptodira: Trionychoidae: Nanhsiungchelyidae). Cretac. Res. 2019, 95, 151–163. [Google Scholar] [CrossRef]
- Swofford, D.L. Phylogenetic Analysis Using Parsimony (*and Other Methods), Version 4.0; Sinauer: Sunderland, MA, USA, 1998. [Google Scholar]
- Thomson, R.C.; Spinks, P.Q.; Shaffer, H.B. A Global Phylogeny of Turtles Reveals a Burst of Climate-Associated Diversification on Continental Margins. Proc. Natl. Acad. Sci. USA 2021, 118, 1–10. [Google Scholar] [CrossRef]
- Meyer, H.v. Helochelys danubina, Aus Dem Grünsande von Kelheim in Bayern. Palaeontographica (1846–1933). 1855; 96–105. [Google Scholar]
- Matheron, P. Mém. Acad. Imp. Sc. B.-L. Arts Marseille. 1869; 39. [Google Scholar]
- Lapparent de Broin, F.d.; Murelaga, X. Une Nouvelle Faune de Chéloniens Dans Le Crétacé Supérieur Européen. Comptes rendus de l’Académie des sciences. Série 2. Sci. De La Terre Et Des Planètes 1996, 323, 729–735. [Google Scholar]
- Lapparent de Broin, F.d.; Murelaga, X. Turtles from the Upper Cretaceous of Laño (Iberian Peninsula). Estud. Del Mus. De Cienc. Nat. De Alava 1999, 14, 135–211. [Google Scholar]
- Rollot, Y.; Evers, S.W.; Pierce, S.E.; Joyce, W.G. Cranial Osteology, Taxonomic Reassessment, and Phylogenetic Relationships of the Early Cretaceous (Aptian-Albian) Turtle Trinitichelys hiatti (Paracryptodira). PeerJ 2022, 10, e14138. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characters/Taxon | Solemys gaudryi | Helochelydra nopcsai | Naomichelys speciosa | Aragochersis lignitesta |
|---|---|---|---|---|
| Skull roof surface ornamentation | Present, fine tubercles and vermiculated ridges that are coalesced and not dislocated | Present, isolated pustules that are not coalesced but easily dislocated | Present, isolated pus- tules that are not coalesced but easily dislocated | Present, short, broad and blunt irregular tubercles, in contact with each other, generating a roughsurface |
| Skull surface sculpture matching that of the shell | Yes | Yes | Yes | No? |
| Skull roof scales | Present | Absent | Present | Absent |
| Temporal emargination | Small | Absent | Wider than in Solemys | Small |
| Supraoccipital exposure on skull roof | Very small or absent | Large, between the parietals | Small, posterior to the parietals | Very small or absent |
| Squamosal expanded medially behind the parietal | Yes | Yes | Yes | ? |
| Contribution of the parietal to the temporal margin | Large, greater than that of squamosal | Smaller than that of squamosal | Smaller than that of squamosal | Large, greater than that of squamosal |
| Parietal excluded from foramen nervi trigemini edge by epipterygoid/prootic contact | Yes | Yes | ? | ? |
| Quadrate exposed between processus paroccipitalis of the opisthotic and squamosal dorsally | Yes, ventral to squamosal/opithotic suture | Yes, dorsal to squamosal/opithotic suture | No | ? |
| Quadratojugal fully covers the processus articularis laterally | Yes | Yes | Yes | Yes |
| Pterygoids fully cover parabasisphenoid ventrally | Yes | Yes | No | Yes? |
| Tubercula basioccipitale | Two pairs Not posterior directed Posterior pair less developed than anterior | Two pairs Posteriorly directed Two pairs similar in size | Two pairs Posteriorly directed Two pairs similar in size | Two pairs Posteriorly directed Two pairs similar in size |
| Anterior pair of tubercula basioccipitale | Formed by pterygoids and basioccipital | Formed entirely by pterygoids | Formed entirely by pterygoids | Formed entirely by pterygoids |
| Position of condylus occipitalis relative to condylus mandibularis | Very close to one another | Close to one another | Distant from one another | Distant from one another |
| Foramen pro ramo vidiani exposed on ventral surface | Yes | Yes? | Yes | ? |
| Osseous subdivision between the inner and middle ear cavities | Present | Absent | Absent | ? |
| Processus interfenestralis of the opisthotic | Present | Absent | Absent | ? |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tong, H.; Buffetaut, E.; Méchin, P.; Méchin-Salessy, A.; Claude, J. A Solemys Skull from the Late Cretaceous of Southern France. Diversity 2023, 15, 58. https://doi.org/10.3390/d15010058
Tong H, Buffetaut E, Méchin P, Méchin-Salessy A, Claude J. A Solemys Skull from the Late Cretaceous of Southern France. Diversity. 2023; 15(1):58. https://doi.org/10.3390/d15010058
Chicago/Turabian StyleTong, Haiyan, Eric Buffetaut, Patrick Méchin, Annie Méchin-Salessy, and Julien Claude. 2023. "A Solemys Skull from the Late Cretaceous of Southern France" Diversity 15, no. 1: 58. https://doi.org/10.3390/d15010058
APA StyleTong, H., Buffetaut, E., Méchin, P., Méchin-Salessy, A., & Claude, J. (2023). A Solemys Skull from the Late Cretaceous of Southern France. Diversity, 15(1), 58. https://doi.org/10.3390/d15010058