




Catalysis before Enzymes: Thiol-Rich Peptides as Molecular Diversity Providers on the Early Earth




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
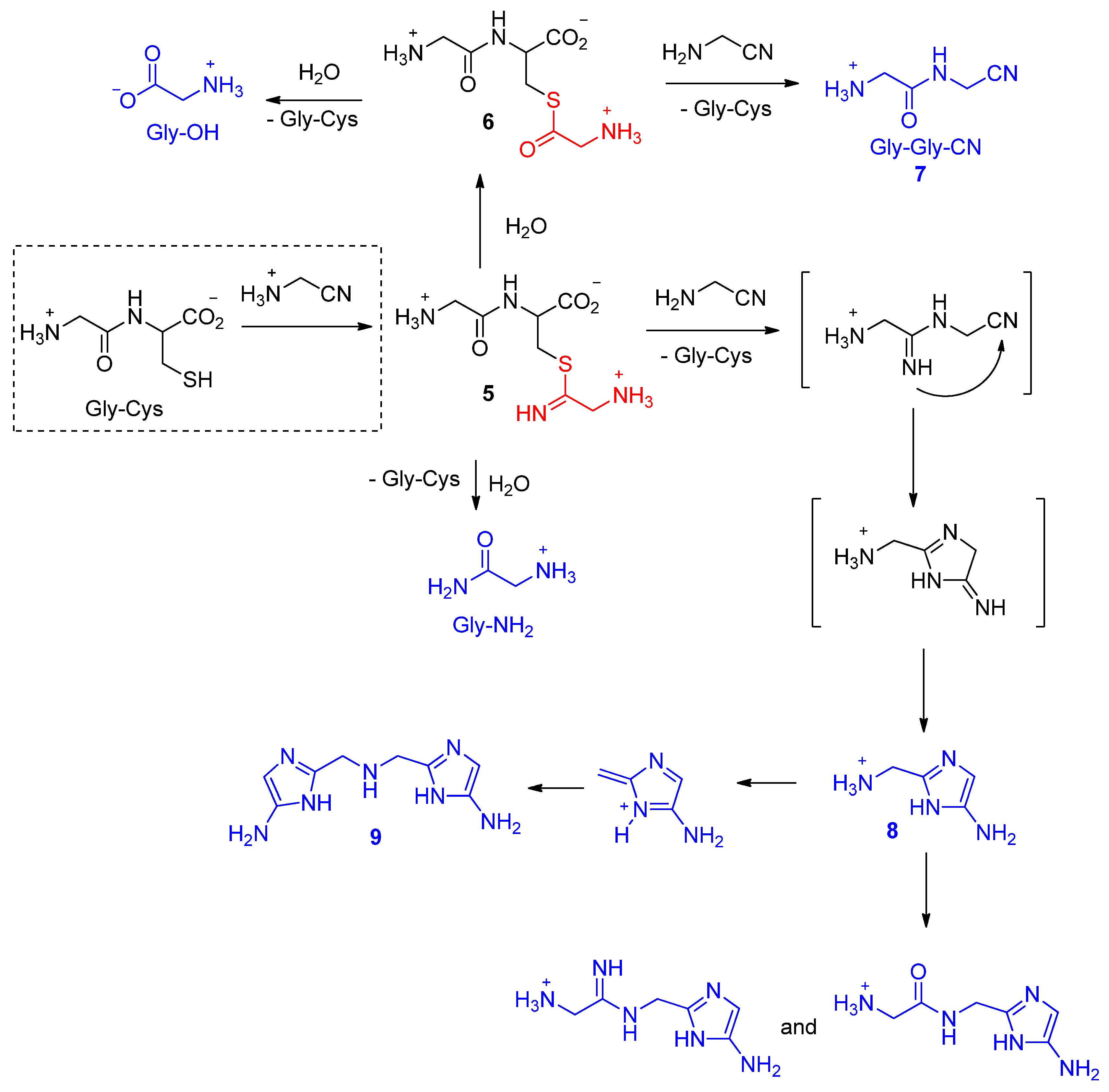
:1. Introduction
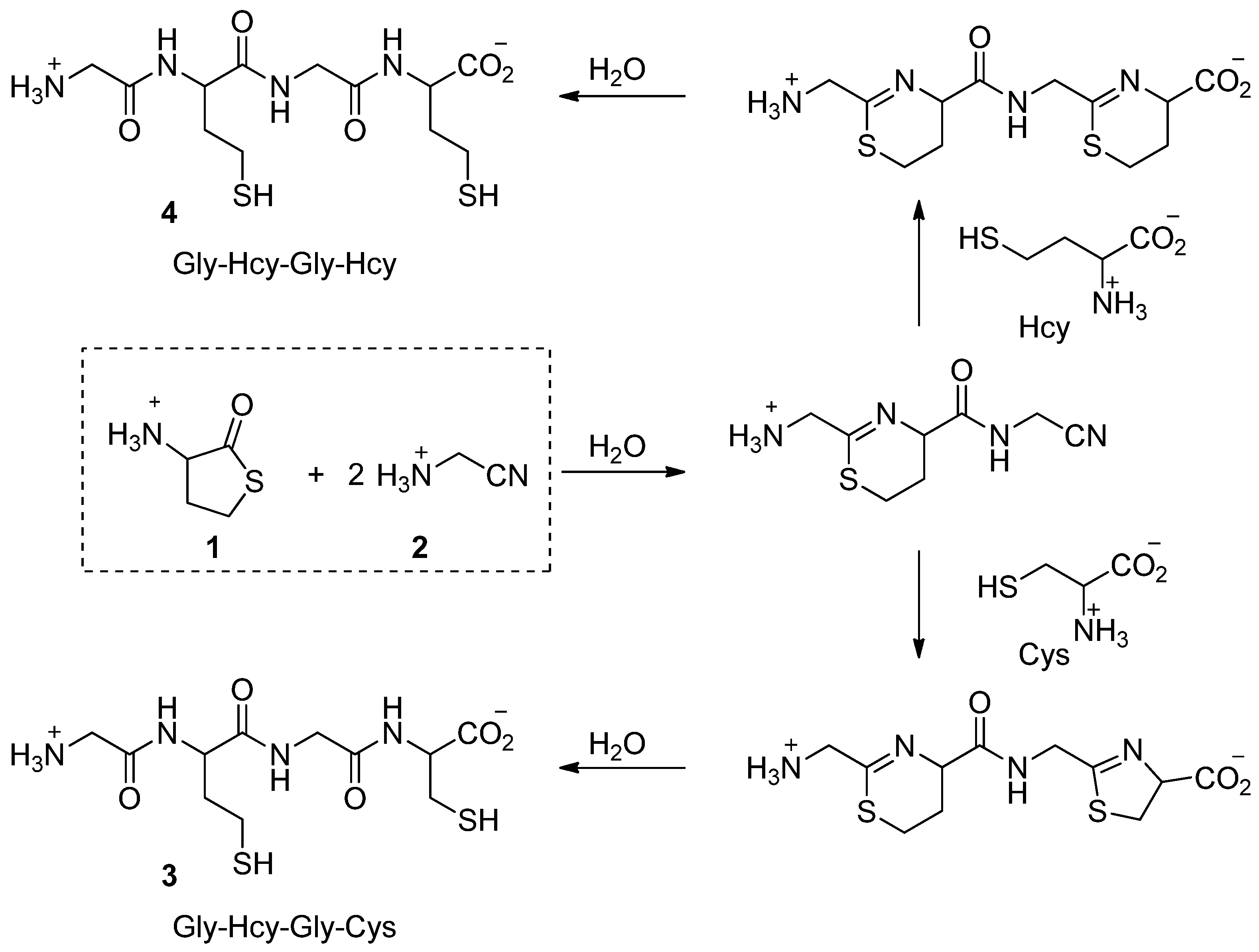
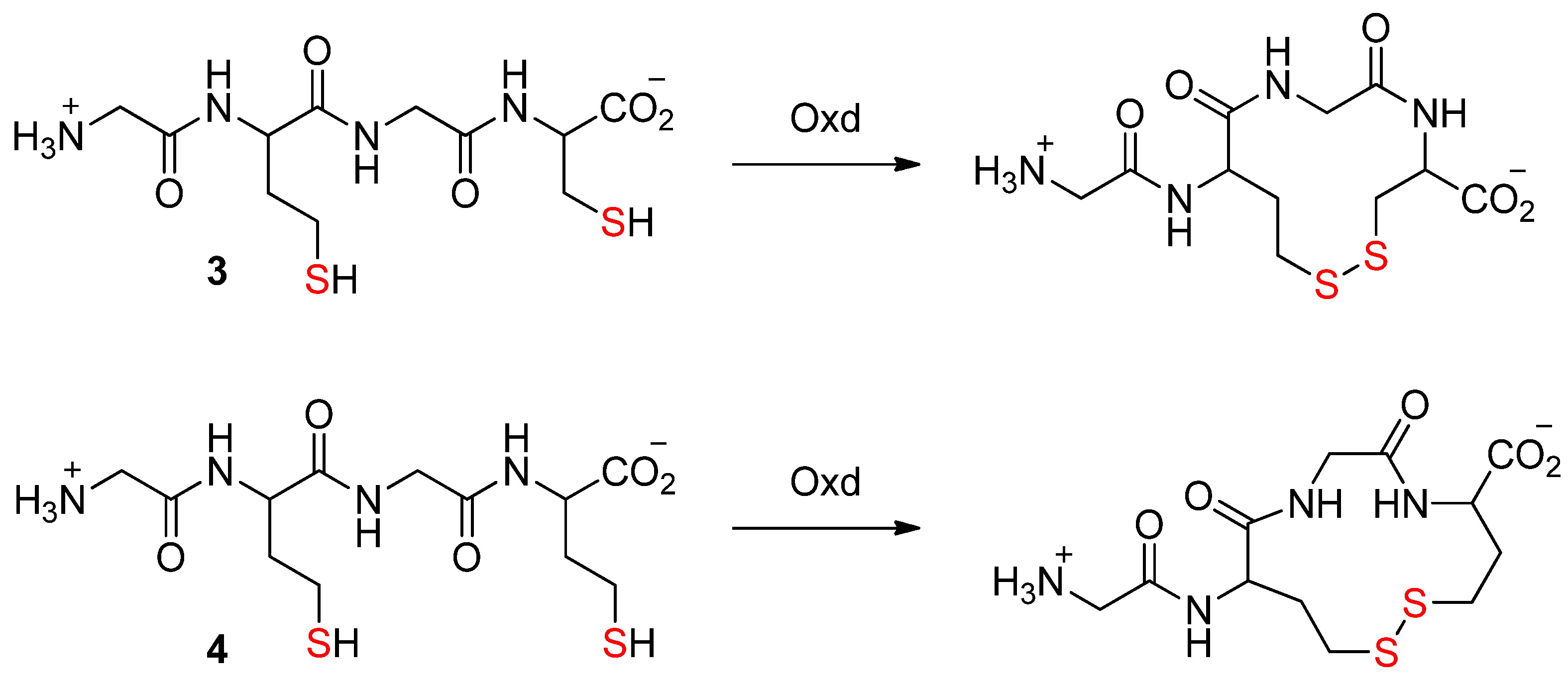
2. Thiols in the Making of Small Peptides and Nitrogen Heterocycles
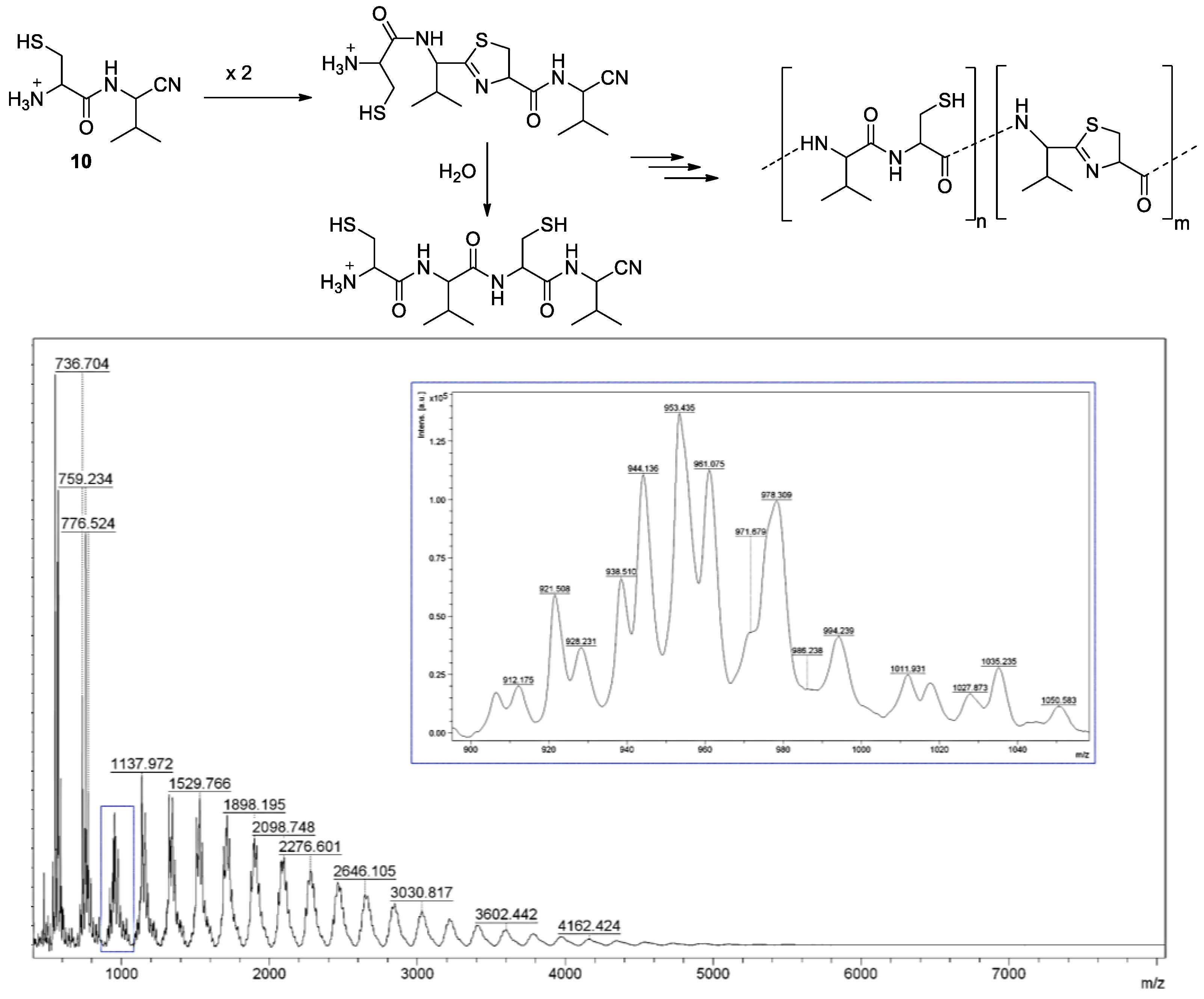
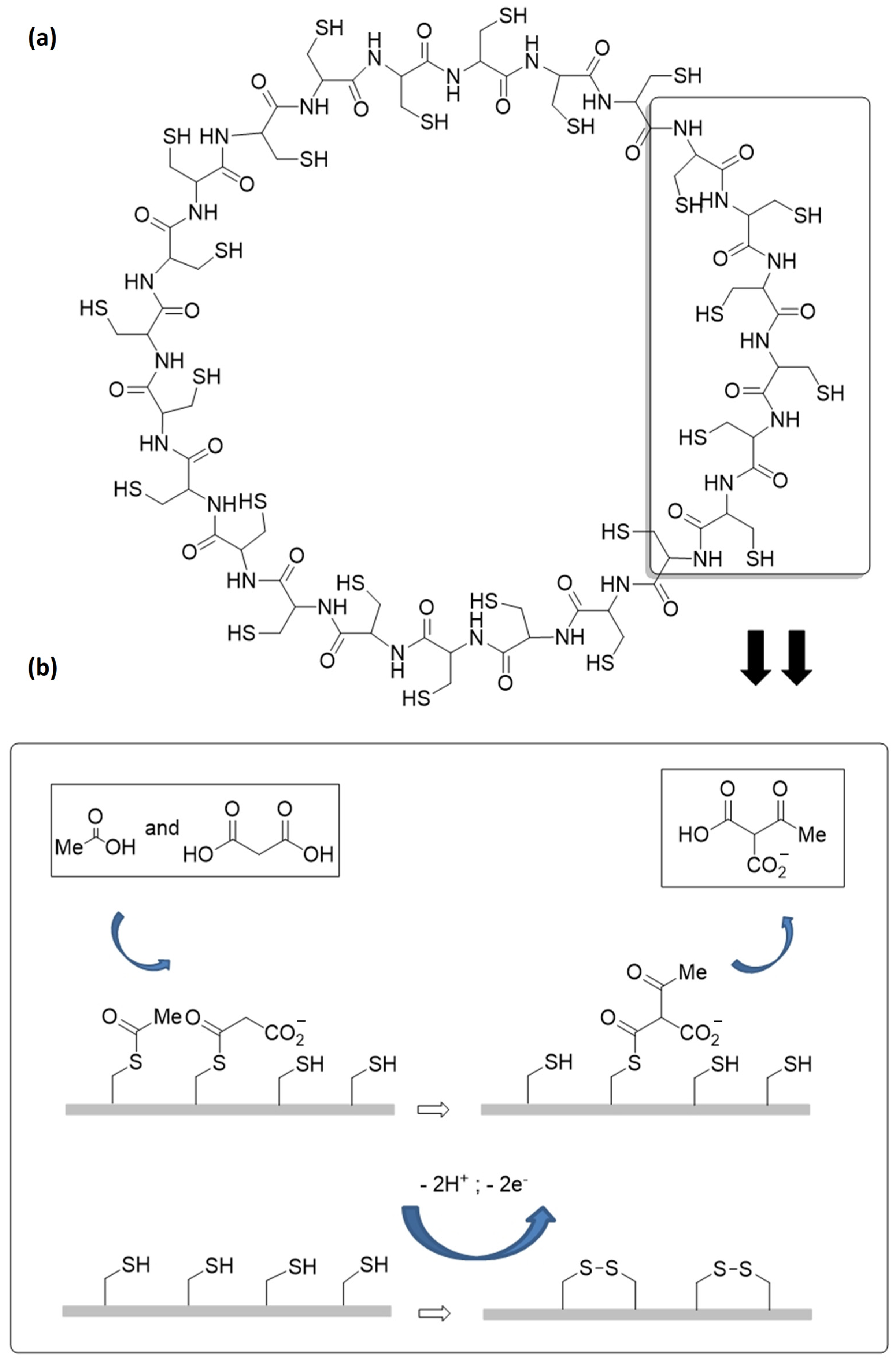
3. Thiols in the Making of Polypeptides
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Glansdorff, N.; Xu, Y.; Labedan, B. The Last Universal Common Ancestor: Emergence, Constitution and Genetic Legacy of an Elusive Forerunner. Biol. Direct 2008, 3, 29. [Google Scholar] [CrossRef]
- Muchowska, K.B.; Varma, S.J.; Moran, J. Nonenzymatic Metabolic Reactions and Life’s Origins. Chem. Rev. 2020, 120, 7708–7744. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Chieffo, C.; Lopez, A.; Fayolle, D.; Ruiz, J.; Soulère, L.; Oger, P.; Altamura, E.; Popowycz, F.; Buchet, R. Synthesis of Phospholipids Under Plausible Prebiotic Conditions and Analogies with Phospholipid Biochemistry for Origin of Life Studies. Astrobiology 2022, 22, 598–627. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F. Early Evolution without a Tree of Life. Biol. Direct 2011, 6, 36. [Google Scholar] [CrossRef]
- Wolfenden, R. Primordial Chemistry and Enzyme Evolution in a Hot Environment. Cell. Mol. Life Sci. 2014, 71, 2909–2915. [Google Scholar] [CrossRef]
- Kurland, C.G. The RNA Dreamtime. BioEssays 2010, 32, 866–871. [Google Scholar] [CrossRef]
- Ulusu, N.N. Evolution of Enzyme Kinetic Mechanisms. J. Mol. Evol. 2015, 80, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Kaschke, M.; Russell, M.J.; Cole, W.J. [FeS/FeS2]. A Redox System for the Origin of Life. Orig. Life Evol. Biosph. 1994, 24, 43–56. [Google Scholar] [CrossRef]
- Soranzo, T.; ben Tahar, A.; Chmayssem, A.; Zelsmann, M.; Vadgama, P.; Lenormand, J.-L.; Cinquin, P.; Martin, D.K.; Zebda, A. Electrochemical Biosensing of Glucose Based on the Enzymatic Reduction of Glucose. Sensors 2022, 22, 7105. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.V.; Flück, C.E. NADPH P450 Oxidoreductase: Structure, Function, and Pathology of Diseases. Pharmacol. Ther. 2013, 138, 229–254. [Google Scholar] [CrossRef]
- Pedone, E.; Ren, B.; Ladenstein, R.; Rossi, M.; Bartolucci, S. Functional Properties of the Protein Disulfide Oxidoreductase from the Archaeon Pyrococcus Furiosus. Eur. J. Biochem. 2004, 271, 3437–3448. [Google Scholar] [CrossRef]
- Kozlowski, L.P. Proteome-PI: Proteome Isoelectric Point Database. Nucleic Acids Res. 2017, 45, D1112–D1116. [Google Scholar] [CrossRef]
- Kua, J.; Miller, N.A. Preliminary Free Energy Map of Prebiotic Compounds Formed from CO2, H2 and H2S. Life 2022, 12, 1763. [Google Scholar] [CrossRef]
- Youssef-Saliba, S.; Vallée, Y. Sulfur Amino Acids: From Prebiotic Chemistry to Biology and Vice Versa. Synthesis 2021, 53, 2798–2808. [Google Scholar]
- Shalayel, I.; Vallée, Y. Chemistry of Homocysteine Thiolactone in a Prebiotic Perspective. Life 2019, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Shalayel, I.; Coulibaly, S.; Ly, K.; Milet, A.; Vallée, Y. The Reaction of Aminonitriles with Aminothiols: A Way to Thiol-Containing Peptides and Nitrogen Heterocycles in the Primitive Earth Ocean. Life 2018, 8, 47. [Google Scholar] [CrossRef] [PubMed]
- Vallee, Y.; Shalayel, I.; Ly, K.-D.; Rao, K.V.R.; de Paëpe, G.; Märker, K.; Milet, A. At the Very Beginning of Life on Earth: The Thiol-Rich Peptide (TRP) World Hypothesis. Int. J. Dev. Biol. 2017, 61, 471–478. [Google Scholar] [CrossRef]
- Shalayel, I. A Plausible Prebiotic Synthesis of Thiol-Rich Peptides: The Reaction of Aminothiols with Aminonitriles. Ph.D. Thesis, University Grenoble Alpes, Grenoble, France, 2018. [Google Scholar]
- Buchanan, B.B.; Holmgren, A.; Jacquot, J.-P.; Scheibe, R. Fifty Years in the Thioredoxin Field and a Bountiful Harvest. Biochim. Biophys. Acta Gen. Subj. 2012, 1820, 1822–1829. [Google Scholar] [CrossRef] [PubMed]
- Leqraa, N.; Nicolet, Y.; Milet, A.; Vallée, Y. A Way to Thioacetate Esters Compatible with Non-Oxidative Prebiotic Conditions. Sci. Rep. 2020, 10, 14488. [Google Scholar] [CrossRef]
- de Duve, C. A Research Proposal on the Origin of Life. Orig. Life Evol. Biosph. 2003, 33, 559–574. [Google Scholar] [CrossRef]
- Shalayel, I.; Youssef-Saliba, S.; Vazart, F.; Ceccarelli, C.; Bridoux, M.; Vallée, Y. Cysteine Chemistry in Connection with Abiogenesis. Eur. J. Org. Chem. 2020, 2020, 3019–3023. [Google Scholar] [CrossRef]
- Yi, J.; Kaur, H.; Kazöne, W.; Rauscher, S.A.; Gravillier, L.; Muchowska, K.B.; Moran, J. A Nonenzymatic Analog of Pyrimidine Nucleobase Biosynthesis. Angew. Chem. Int. Ed. 2022, 61, e202117211. [Google Scholar] [CrossRef]
- Rossetto, D.; Mansy, S.S. Metals Are Integral to Life as We Know It. Front. Cell Dev. Biol. 2022, 10, 864830. [Google Scholar] [CrossRef]
- Ludlow, R.F.; Otto, S. Systems Chemistry. Chem. Soc. Rev. 2008, 37, 101–108. [Google Scholar] [CrossRef]
- Ruiz-Mirazo, K.; Briones, C.; De la Escosura, A. Prebiotic Systems Chemistry: New Perspectives for the Origins of Life. Chem. Rev. 2014, 114, 285–366. [Google Scholar] [CrossRef]
- Lopez, A.; Fayolle, D.; Fiore, M.; Strazewski, P. Chemical Analysis of Lipid Boundaries after Consecutive Growth and Division of Supported Giant Vesicles. iScience 2020, 23, 101677. [Google Scholar] [CrossRef] [PubMed]
- Fiore, M.; Maniti, O.; Girard-Egrot, A.; Monnard, P.-A.; Strazewski, P. Glass Microsphere-Supported Giant Vesicles for the Observation of Self-Reproduction of Lipid Boundaries. Angew. Chem. Int. Ed. 2018, 57, 282–286. [Google Scholar] [CrossRef] [PubMed]
- Royal Society of Chemistry. Prebiotic Chemistry and Life’s Origin; Fiore, M., Ed.; Chemical Biology; Royal Society of Chemistry: Cambridge, UK, 2022; ISBN 978-1-78801-749-7. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shalayel, I.; Leqraa, N.; Blandin, V.; Vallée, Y. Catalysis before Enzymes: Thiol-Rich Peptides as Molecular Diversity Providers on the Early Earth. Diversity 2023, 15, 256. https://doi.org/10.3390/d15020256
Shalayel I, Leqraa N, Blandin V, Vallée Y. Catalysis before Enzymes: Thiol-Rich Peptides as Molecular Diversity Providers on the Early Earth. Diversity. 2023; 15(2):256. https://doi.org/10.3390/d15020256
Chicago/Turabian StyleShalayel, Ibrahim, Naoual Leqraa, Veronique Blandin, and Yannick Vallée. 2023. "Catalysis before Enzymes: Thiol-Rich Peptides as Molecular Diversity Providers on the Early Earth" Diversity 15, no. 2: 256. https://doi.org/10.3390/d15020256
APA StyleShalayel, I., Leqraa, N., Blandin, V., & Vallée, Y. (2023). Catalysis before Enzymes: Thiol-Rich Peptides as Molecular Diversity Providers on the Early Earth. Diversity, 15(2), 256. https://doi.org/10.3390/d15020256