
Climate Change Impacts on Species Composition and Floristic Regions in Thailand



Abstract
:1. Introduction
2. Methods
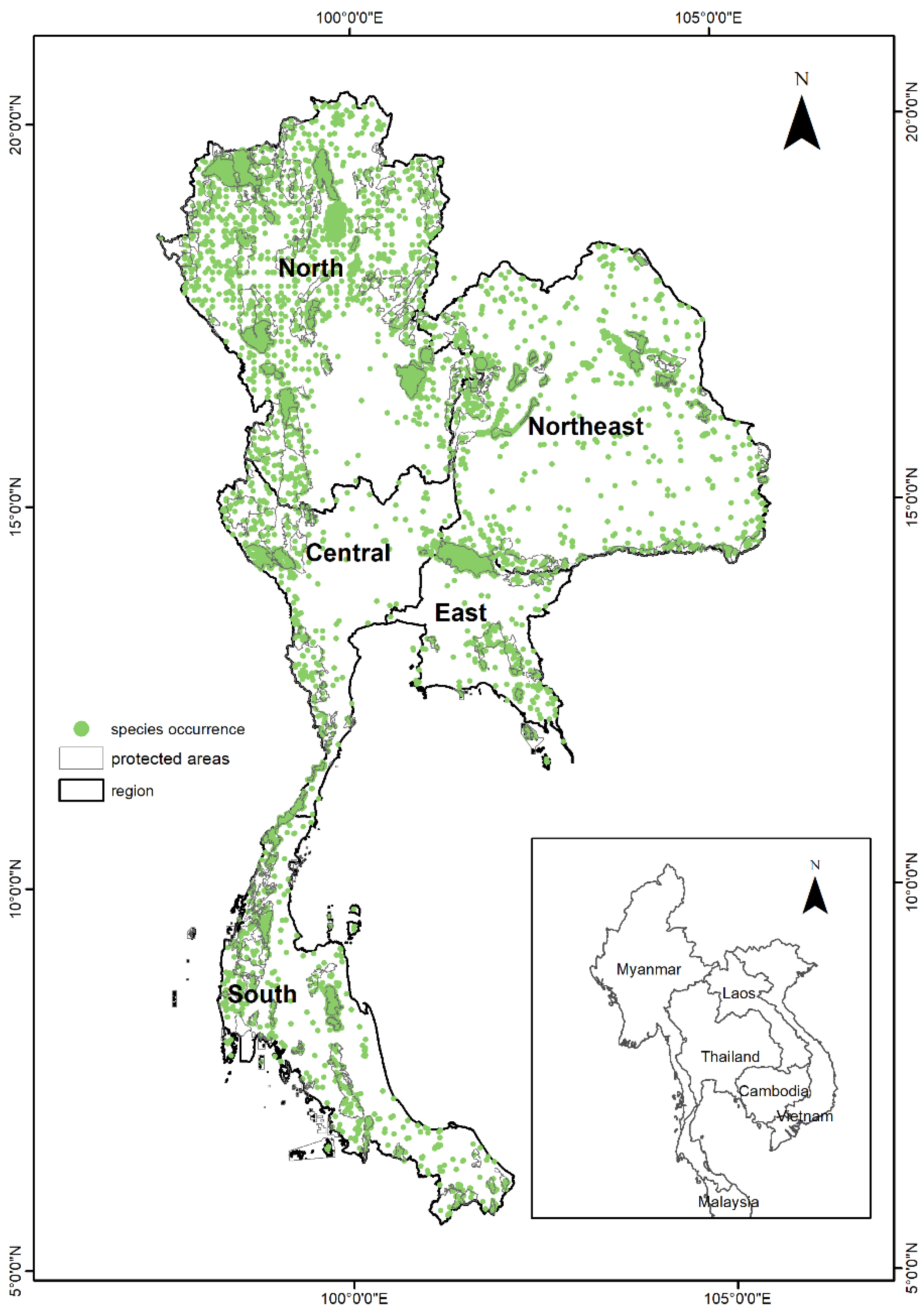
2.1. Study Area
2.2. Species Selection
2.3. Environmental Variables
2.4. Generation of Species Distribution
2.5. Clustering Floristic Regions Based on SDMs
3. Results
3.1. Candidate Tree Species for Modeling
3.2. Selected Environmental Variables and Their Contributions
3.3. Projected Species Ranges
3.4. Changes in Suitable Range for the Modeled Species
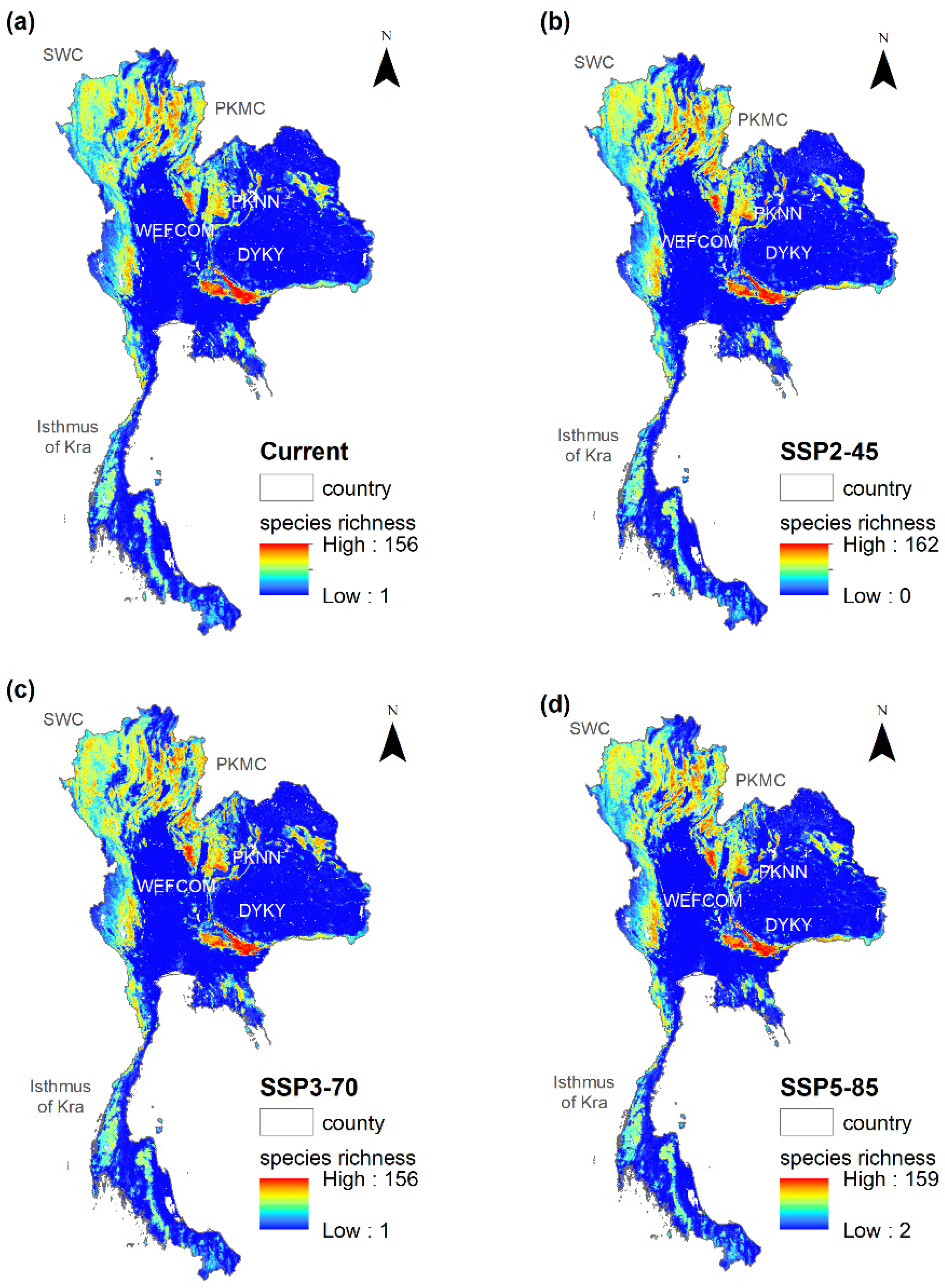
3.5. Potential Floristic Regions
3.6. Changes in Habitat of Individual Potential Floristic Regions
4. Discussion
4.1. How Do Tropical Tree Species in Thailand Spatially Response to Future Climate Change?
4.2. How Many Optimum Floristic Regions Can Be Delineated in Thailand Based on the Current Species Distribution?
4.3. Which Floristic Regions Are Vulnerable to Climate Change?
4.4. Conservation Implications
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murphy, B.P.; Bowman, D. What controls the distribution of tropical forest and savanna? Ecol. Lett. 2012, 15, 748–758. [Google Scholar] [CrossRef]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Da Fonseca, G.A.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef] [PubMed]
- Mitchard, E.T.A. The tropical forest carbon cycle and climate change. Nature 2018, 559, 527–534. [Google Scholar] [CrossRef] [PubMed]
- Qie, L.; Lewis, S.L.; Sullivan, M.J.; Lopez-Gonzalez, G.; Pickavance, G.C.; Sunderland, T.; Ashton, P.; Hubau, W.; Abu Salim, K.; Aiba, S.I.; et al. Long-term carbon sink in Borneo’s forests halted by drought and vulnerable to edge effects. Nat. Commun. 2017, 8, 1966. [Google Scholar] [CrossRef] [PubMed]
- Law, E.A.; Bryan, B.A.; Meijaard, E.; Mallawaarachchi, T.; Struebig, M.J.; Watts, M.E.; Wilson, K.A. Mixed policies give more options in multifunctional tropical forest landscapes. J. Appl. Ecol. 2017, 54, 51–60. [Google Scholar] [CrossRef]
- Curtis, P.G.; Slay, C.M.; Harris, N.L.; Tyukavina, A.; Hansen, M.C. Classifying drivers of global forest loss. Science 2018, 361, 1108–1111. [Google Scholar] [CrossRef] [PubMed]
- Hosonuma, N.; Herold, M.; De Sy, V.; De Fries, R.S.; Brockhaus, M.; Verchot, L.; Romijn, E. An assessment of deforestation and forest degradation drivers in developing countries. Environ. Res. Lett. 2012, 7, 044009. [Google Scholar] [CrossRef]
- Seymour, F.; Harris, N.L. Reducing tropical deforestation. Science 2019, 365, 756–757. [Google Scholar] [CrossRef]
- FAO; UNEP. The State of the World’s Forests 2020; FAO: Rome, Italy; UNEP: Rome, Italy, 2020. [Google Scholar]
- RFD. Forest Statistic Report 2019; Royal Forest Department, Bangkok: Bangkok, Thailand, 2019. [Google Scholar]
- Brando, P.M.; Paolucci, L.; Ummenhofer, C.C.; Ordway, E.M.; Hartmann, H.; Cattau, M.E.; Rattis, L.; Medjibe, V.; Coe, M.T.; Balch, J. Droughts, wildfires, and forest carbon cycling: A pantropical synthesis. Annu. Rev. Earth Planet. Sci. 2019, 47, 555–581. [Google Scholar] [CrossRef]
- Malhi, Y.; Gardner, T.A.; Goldsmith, G.R.; Silman, M.R.; Zelazowski, P. Tropical forests in the Anthropocene. Annu. Rev. Environ. Resour. 2014, 39, 125–159. [Google Scholar] [CrossRef]
- Von Rintelen, K.; Arida, E.; Häuser, C. A review of biodiversity-related issues and challenges in megadiverse Indonesia and other Southeast Asian countries. Res. Ideas Outcomes 2017, 3, e20860. [Google Scholar] [CrossRef]
- Zelazowski, P.; Malhi, Y.; Huntingford, C.; Sitch, S.; Fisher, J.B. Changes in the potential distribution of humid tropical forests on a warmer planet. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2011, 369, 137–160. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014 Synthesis Report; Intergovernmental Panel on Climate Change (IPCC): Geneva, Switzerland, 2014. [Google Scholar]
- Zebiak, S.E.; Orlove, B.; Muñoz, Á.G.; Vaughan, C.; Hansen, J.; Troy, T.; Thomson, M.C.; Lustig, A.; Garvin, S. Investigating El Niño-Southern Oscillation and society relationships. Wiley Interdisciplinary Reviews. Clim. Chang. 2015, 6, 17–34. [Google Scholar]
- IPCC. Summary for policymakers. In Climate Chang. 2021: The Physical Science Basis; Masson-Delmotte, V., Zhai, P., Pirani, A., Connors, S.L., Péan, C., Berger, S., Caud, N., Chen, Y., Goldfarb, L., Gomis, M.I., et al., Eds.; Contribution of Working Group I to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Liu, H.; Mi, Z.; Lin, L.I.; Wang, Y.; Zhang, Z.; Zhang, F.; Wang, H.; Liu, L.; Zhu, B.; Cao, G.; et al. Shifting plant species composition in response to climate change stabilizes grassland primary production. Proc. Natl. Acad. Sci. USA 2018, 115, 4051–4056. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.Q.; Xie, S.P. Effects of climatological model biases on the projection of tropical climate change. J. Clim. 2015, 28, 9909–9917. [Google Scholar] [CrossRef]
- Aguirre-Gutiérrez, J.; Oliveras, I.; Rifai, S.; Fauset, S.; Adu-Bredu, S.; Affum-Baffoe, K.; Baker, T.R.; Feldpausch, T.R.; Gvozdevaite, A.; Hubau, W.; et al. Drier tropical forests are susceptible to functional changes in response to a long-term drought. Ecol. Lett. 2019, 22, 855–865. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, M.K.; Detto, M.; Pacala, S.W. Predicting shifts in the functional composition of tropical forests under increased drought and CO 2 from trade-offs among plant hydraulic traits. Ecol. Lett. 2019, 22, 67–77. [Google Scholar] [CrossRef]
- Corlett, R.T. The impacts of droughts in tropical forests. Trends Plant Sci. 2016, 21, 584–593. [Google Scholar] [CrossRef] [PubMed]
- Fontes, C.G.; Dawson, T.E.; Jardine, K.; McDowell, N.; Gimenez, B.O.; Anderegg, L.; Negrón-Juárez, R.; Higuchi, N.; Fine, P.V.; Araújo, A.C.; et al. Dry and hot: The hydraulic consequences of a climate change–type drought for Amazonian trees. Philos. Trans. R. Soc. B Biol. Sci. 2018, 373, 20180209. [Google Scholar] [CrossRef] [PubMed]
- Johnston, J.M.; Zomer, R.J.; Wang, M.C. Predicting future vegetated landscapes under climate change: Application of the environmental stratification methodology to protected areas in the Lower Mekong Basin. In Ecosystem-Based Management, Ecosystem Services and Aquatic Biodiversity; O’Higgins, T., Lago, M., DeWitt, T., Eds.; Springer: Cham, Switzerland, 2020. [Google Scholar] [CrossRef]
- Pomoim, N.; Hughes, A.C.; Trisurat, Y.; Corlett, R.T. Vulnerability to climate change of species in protected areas in Thailand. Sci. Rep. 2022, 12, 5705. [Google Scholar] [CrossRef]
- Zomer, R.J.; Trabucco, A.; Wang, M.; Lang, R.; Chen, H.; Metzger, M.J.; Smajgl, A.; Beckschäfer, P.; Xu, J. Environmental stratification to model climate change impacts on biodiversity and rubber production in Xishuangbanna, Yunnan, China. Biol. Conserv. 2014, 170, 264–273. [Google Scholar] [CrossRef]
- Brodie, J.F. Synergistic effects of climate change and agricultural land use on mammals. Front. Ecol. Environ. 2016, 14, 20–26. [Google Scholar] [CrossRef]
- Anadón, J.D.; Sala, O.E.; Maestre, F.T. Climate change will increase savannas at the expense of forests and treeless vegetation in tropical and subtropical Americas. J. Ecol. 2014, 102, 1363–1373. [Google Scholar] [CrossRef]
- Rashid, I.; Romshoo, S.A.; Chaturvedi, R.K.; Ravindranath, N.H.; Sukumar, R.; Jayaraman, M.; Lakshmi, T.V.; Sharma, J. Projected climate change impacts on vegetation distribution over Kashmir Himalayas. Clim. Chang. 2015, 132, 601–613. [Google Scholar] [CrossRef]
- 30 Rehm, E.M.; Olivas, P.; Stroud, J.; Feeley, K.J. Losing your edge: Climate change and the conservation value of range-edge populations. Ecol. Evol. 2015, 5, 4315–4326. [Google Scholar] [CrossRef] [PubMed]
- Fadrique, B.; Báez, S.; Duque, Á.; Malizia, A.; Blundo, C.; Carilla, J.; Osinaga-Acosta, O.; Malizia, L.; Silman, M.; Farfán-Ríos, W.; et al. Widespread but heterogeneous responses of Andean forests to climate change. Nature 2018, 564, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.C.; Clark, T.D.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity redistribution under climate change: Impacts on ecosystems and human well-being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef] [PubMed]
- Krupnick, G.A. Conservation of tropical plant biodiversity: What have we done, where are we going? Biotropica 2013, 45, 693–708. [Google Scholar] [CrossRef]
- Nicotra, A.B.; Atkin, O.K.; Bonser, S.P.; Davidson, A.M.; Finnegan, E.J.; Mathesius, U.; Poot, P.; Purugganan, M.D.; Richards, C.L.; Valladares, F.; et al. Plant phenotypic plasticity in a changing climate. Trends Plant Sci. 2010, 15, 684–692. [Google Scholar] [CrossRef] [PubMed]
- Trisurat, Y.; Shrestha, R.P.; Kjelgren, R. Plant species vulnerability to climate change in Peninsular Thailand. Appl. Geogr. 2011, 31, 1106–1114. [Google Scholar] [CrossRef]
- Normah, M.N.; Sulong, N.; Reed, B.M. Cryopreservation of shoot tips of recalcitrant and tropical species: Advances and strategies. Cryobiology 2019, 87, 1–14. [Google Scholar] [CrossRef]
- Fernández, A.; León-Lobos, P.; Contreras, S.; Ovalle, J.F.; van der Walt, K.; Ballesteros, D. The potential impacts of climate change on ex situ conservation options for recalcitrant-seeded species. Front. For. Glob. Chang. 2023, 6, 1110431. [Google Scholar] [CrossRef]
- Trisurat, Y.; Alkemade, J.R.M.; Arets, E.J.M.M. Projecting forest tree distributions and adaptation to climate change in northern Thailand. J. Ecol. Nat. Environ. 2009, 1, 55–63. [Google Scholar]
- Van Zonneveld, M.; Koskela, J.; Vinceti, B.; Jarvis, A. Impact of climate change on the distribution of tropical pines in Southeast Asia. Unasylva 2009, 60, 24–28. [Google Scholar]
- Wang, T.; Wang, G.; Innes, J.; Nitschke, C.; Kang, H. Climatic niche models and their consensus projections for future climates for four major forest tree species in the Asia–Pacific region. For. Ecol. Manag. 2016, 360, 357–366. [Google Scholar] [CrossRef]
- Hiura, T.; Go, S.; Iijima, H. Long-term forest dynamics in response to climate change in northern mixed forests in Japan: A 38-year individual-based approach. For. Ecol. Manag. 2019, 449, 117469. [Google Scholar] [CrossRef]
- Takhtadzhi︠a︡n, A.L.; Takhtadzhi︠a︡n, L.A.; Takhtajan, A.; Crovello, T.J. Floristic Regions of the World; University of California Press: Berkeley, CA, USA, 1986. [Google Scholar]
- Middleton, D.J. Progress on the Flora of Thailand. Telopea 2003, 10, 33–42. [Google Scholar] [CrossRef]
- Parnell, J.A.N.; Simpson, D.A.; Moat, J.; Kirkup, D.W.; Chantaranothai, P.; Boyce, P.C.; Bygrave, P.; Dransfield, S.; Jebb, M.H.P.; Macklin, J.; et al. Plant collecting spread and densities: Their potential impact on biogeographical studies in Thailand. J. Biogeogr. 2003, 30, 193–209. [Google Scholar] [CrossRef]
- Santisuk, T.; Smitinand, T.; Hoamuangkaew, W.; Ashton, P.; Sohmer, S.H.; Vincent, J.R. Plants for Our Future: Botanical Research and Conservation Needs in Thailand; Royal Forest Department: Bangkok, Thailand, 1991. [Google Scholar]
- Smitinand, T. The genus Dipterocarpus Gaertn. f. in Thailand. Thai For. Bull. 1980, 4, 1–26. [Google Scholar]
- Dinerstein, E.; Olson, D.; Joshi, A.; Vynne, C.; Burgess, N.D.; Wikramanayake, E.; Hahn, N.; Palminteri, S.; Hedao, P.; Noss, R.; et al. An ecoregion-based approach to protecting half the terrestrial realm. BioScience 2017, 67, 534–545. [Google Scholar] [CrossRef] [PubMed]
- ONEP. Master Plan for Integrated Biodiversity Management B.E. 2558–2564 (2015-2021); Office of Natural Resources and Environmental Policy and Planning, Ministry of Natural Resources and Environment: Bangkok, Thailand, 2016. [Google Scholar]
- Trisurat, Y.; Eiadthong, W.; Khunrattanasiri, W.; Saengnin, S.; Chitechote, A.; Maneerat, S. Systematic forest inventory plots and their contribution to plant distribution and climate change impact studies in Thailand. Ecol. Res. 2020, 35, 724–732. [Google Scholar] [CrossRef]
- Brown, J.L.; Bennett, J.R.; French, C.M. SDMtoolbox 2.0: The next generation Python-based GIS toolkit for landscape genetic, biogeographic and species distribution model analyses. PeerJ 2017, 5, e4095. [Google Scholar] [CrossRef]
- Boria, R.A.; Olson, L.E.; Goodman, S.M.; Anderson, R.P. Spatial filtering to reduce sampling bias can improve the performance of ecological niche models. Ecol. Model. 2014, 275, 73–77. [Google Scholar] [CrossRef]
- Moudrý, V.; Šímová, P. Influence of positional accuracy, sample size and scale on modelling species distributions: A review. Int. J. Geogr. Inf. Sci. 2012, 26, 2083–2095. [Google Scholar] [CrossRef]
- Santini, L.; Benítez-López, A.; Maiorano, L.; Čengić, M.; Huijbregts, M.A. Assessing the reliability of species distribution projections in climate change research. Div. Dist. 2021, 27, 1035–1050. [Google Scholar] [CrossRef]
- .Wisz, M.S.; Guisan, A. Do pseudo-absence selection strategies influence species distribution models and their predictions? An information-theoretic approach based on simulated data. BMC Ecol. 2009, 9, 8. [Google Scholar] [CrossRef] [PubMed]
- FAO. Harmonized World Soil Database, Version 1.2; FAO/IIASA/ISRIC/ISS-CAS/JRC; FAO: Rome, Italy, 2012. [Google Scholar]
- Swart, N.C.; Cole, J.N.S.; Kharin, V.V.; Lazare, M.; Scinocca, J.F.; Gillett, N.P.; Anstey, J.; Arora, V.; Christian, J.R.; Hanna, S.; et al. The Canadian earth system model version 5 (CanESM5.0.3). Geosci. Model Dev. 2019, 12, 4823–4873. [Google Scholar] [CrossRef]
- Yan, Y.; Zhu, C.; Liu, B.; Jiang, S. Annual cycle of East Asian precipitation simulated by CMIP6 Models. Atmosphere 2021, 12, 24. [Google Scholar] [CrossRef]
- Hosmer, D.W.; Lemeshow, S.; Sturdivant, R.X. Introduction to the logistic regression model. In Applied Logistic Regression; Hosmer, D.M., Lemesbhow, S., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2000; pp. 1–30. [Google Scholar]
- Phillips, S.J.; Anderson, R.P.; Schapire, R.E. Maximum entropy modeling of species geographic distributions. Ecol. Model. 2006, 190, 231–259. [Google Scholar] [CrossRef]
- Elith, J.; Phillips, S.P.; Hastie, T.; Dudik, M.; Chee, Y.E.; Yates, C.J. A statistical explanation to MaxEnt for ecologists. Divers. Distrib. 2010, 17, 43–57. [Google Scholar] [CrossRef]
- Manel, S.; Dias, J.-M.; Ormerod, S.J. Comparing discriminant analysis, neural networks and logistic regression for predicting species distributions: A case study with a Himalayan River bird. Ecol. Model. 1999, 120, 337–347. [Google Scholar] [CrossRef]
- Liu, C.; Berry, P.M.; Dawson, T.P.; Pearson, R.G. Selecting thresholds of occurrence in the prediction of species distributions. Ecography 2005, 28, 385–393. [Google Scholar] [CrossRef]
- Hair, J.F.; Anderson, R.E.; Tatham, R.L.; Black, W.C. Multivariate Data Analysis With Eeadings, 3rd ed.; Macmillan Polishing Company: New York, NY, USA, 1992. [Google Scholar]
- Smitinand, T. Thai Plant Names. Bangkok Forest Herbarium, Department of National Parks; Wildlife and Plant Conservation: Bangkok, Thailand, 2014. [Google Scholar]
- Sterck, F.; Anten, N.P.; Schieving, F.; Zuidema, P.A. Trait acclimation mitigates mortality risks of tropical canopy trees under global warming. Front. Plant Sci. 2016, 7, 607. [Google Scholar] [CrossRef] [PubMed]
- Schleussner, C.F.; Rogelj, J.; Schaeffer, M.; Lissner, T.; Licker, R.; Fischer, E.M.; Knutti, R.; Levermann, A.; Frieler, K.; Hare, W. Science and policy characteristics of the Paris Agreement temperature goal. Nat. Clim. Chang. 2016, 6, 827–835. [Google Scholar] [CrossRef]
- Chong, K.Y.; Chong, R.; Tan, L.W.; Yee, A.T.; Chua, M.A.; Wong, K.M.; Tan, H.T. Seed production and survival of four dipterocarp species in degraded forests in Singapore. Plant Ecol. Divers. 2016, 9, 483–490. [Google Scholar] [CrossRef]
- O’Brien, M.J.; Reynolds, G.; Ong, R.; Hector, A. Resistance of tropical seedlings to drought is mediated by neighbourhood diversity. Nat. Ecol. Evol. 2017, 1, 1643–1648. [Google Scholar] [CrossRef] [PubMed]
- Pulla, S.; Ramaswami, G.; Mondal, N.; Chitra-Tarak, R.; Suresh, H.S.; Dattaraja, H.S.; Vivek, P.; Parthasarathy, N.; Ramesh, B.R.; Sukumar, R. Assessing the resilience of global seasonally dry tropical forests. Int. For. Rev. 2015, 17, 91–113. [Google Scholar] [CrossRef]
- Umarani, R.; Aadhavan, E.K.; Faisal, M.M. Understanding poor storage potential of recalcitrant seeds. Curr. Sci. 2015, 108, 2023–2034. [Google Scholar]
- Tsioumani, E. Convention on Biological Diversity: A Review of the Post-2020 Global Biodiversity Framework Working Group Negotiations. Environ. Policy Law 2020, 50, 55–59. [Google Scholar] [CrossRef]
- CBD Secretariat. The Kunming-Montreal Global Biodiversity Framework of the Conference of Parties to the UN Convention on Biological Diversity; CBD/COP/15/L25; Convention in Biological Diversity: Montreal, QC, Canada, 2022. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Description | Baseline | Scenarios | Change from Baseline | ||||
|---|---|---|---|---|---|---|---|---|
| SSP2-4.5 | SSP3-7.0 | SSP5-8.5 | 2000—SSP2-4.5 | 2000—SSP3-7.0 | 2000—SSP5-8.5 | |||
| BIO1 | Annual mean temperature | 26.1 | 26.4 | 27.5 | 28.8 | 0.3 | 1.4 | 2.7 |
| BIO2 | Mean diurnal range | 10.6 | 10.1 | 9.97 | 10.2 | −0.5 | −0.63 | −0.4 |
| BIO3 | Isothermality | 56.7 | 55.8 | 56.6 | 55.2 | −0.9 | −0.1 | −1.5 |
| BIO4 | Temperature seasonality | 187.5 | 203.4 | 200.7 | 213.1 | 15.9 | 13.2 | 25.6 |
| BIO5 | Maximum temperature of warmest month | 34.9 | 37.3 | 36,1 | 38.3 | 2.4 | 1.2 | 3.4 |
| BIO6 | Minimum temperature of coldest month | 15.9 | 18.7 | 18.1 | 19.4 | 2.8 | 2.2 | 3.5 |
| BIO7 | Temperature annual range | 19.0 | 18.6 | 18.0 | 19.0 | −0.4 | −1 | 0 |
| BIO8 | Mean temperature of wettest quarter | 26.6 | 28.4 | 27.8 | 29.0 | 1.8 | 1.2 | 2.4 |
| BIO9 | Mean temperature of driest quarter | 23.8 | 25.8 | 25.4 | 26.4 | 2 | 1.6 | 2.6 |
| BIO10 | Mean temperature of warmest quarter | 28.2 | 30.5 | 29.8 | 31.4 | 2.3 | 1.6 | 3.2 |
| BIO11 | Mean temperature of coldest quarter | 23.4 | 25.5 | 24.9 | 26.1 | 2.1 | 1.5 | 2.7 |
| BIO12 | Annual precipitation | 1495.2 | 1618.8 | 1530.4 | 1615.9 | 123.6 | 35.2 | 120.7 |
| BIO13 | Precipitation of wettest month | 299.7 | 323.5 | 305.6 | 328.7 | 23.8 | 5.9 | 29 |
| BIO14 | Precipitation of driest month | 10.2 | 8.34 | 8.0 | 8.2 | −1.86 | −2.2 | −2 |
| BIO15 | Precipitation seasonality | 79.6 | 85.7 | 86.6 | 87.4 | 6.1 | 7 | 7.8 |
| BIO16 | Precipitation of wettest quarter | 756.8 | 841.9 | 802.2 | 857.4 | 85.1 | 45.4 | 100.6 |
| BIO17 | Precipitation of driest quarter | 48.8 | 48.9 | 41.5 | 41.9 | 0.1 | −7.3 | −6.9 |
| BIO18 | Precipitation of warmest quarter | 372.0 | 378.8 | 390.4 | 346.8 | 6.8 | 18.4 | −25.2 |
| BIO19 | Precipitation of coldest quarter | 120.7 | 126.1 | 130.3 | 124.2 | 5.4 | 9.6 | 3.5 |
| Extent at Present | SSP2-4.5 in 2050 | SSP3-7.0 in 2050 | SSP5-8.5 in 2050 | % Found in Protected Areas | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Cluster | Total (%) | Total (%) | Change (%) | Total (%) | Change (%) | Total (%) | Change (%) | 2000 | SSP2-45 | SSP3-70 | SSP5-85 |
| 0 1 | 67.61 | 67.61 | 0.00 | 67.61 | 0.00 | 67.61 | 0.00 | 4.41 | 4.41 | 4.41 | 4.41 |
| 1/3 2 | 10.42 | 10.15 | −2.63 | 10.13 | −2.82 | 10.08 | −3.26 | 49.84 | 49.16 | 50.97 | 47.47 |
| 2 | 4.50 | 4.61 | 2.57 | 4.55 | 1.30 | 4.16 | −7.51 | 72.15 | 71.90 | 68.72 | 72.38 |
| 4 | 6.98 | 6.45 | −7.69 | 6.82 | −2.39 | 6.83 | −2.27 | 55.07 | 57.03 | 54.30 | 57.98 |
| 5 | 1.15 | 1.37 | 19.05 | 1.44 | 25.40 | 1.33 | 16.03 | 77.01 | 79.86 | 75.43 | 96.67 |
| 6 | 1.85 | 2.08 | 12.18 | 1.67 | −9.70 | 1.81 | −2.07 | 42.31 | 40.01 | 41.97 | 36.92 |
| 7 | 0.88 | 0.79 | −11.20 | 0.51 | −42.40 | 0.78 | −11.81 | 96.96 | 95.40 | 92.87 | 92.09 |
| 8 | 0.89 | 0.79 | −11.60 | 0.99 | 11.01 | 1.08 | 21.74 | 99.22 | 102.34 | 95.56 | 101.62 |
| 9 | 0.54 | 0.43 | −21.40 | 0.62 | 14.19 | 0.54 | 0.37 | 90.26 | 94.03 | 89.96 | 92.42 |
| 10 | 1.32 | 1.42 | 7.24 | 1.56 | 18.64 | 1.69 | 28.15 | 96.23 | 92.79 | 91.66 | 88.83 |
| 11 | 1.98 | 2.25 | 13.59 | 1.96 | −1.14 | 1.94 | −2.06 | 75.94 | 71.34 | 74.53 | 72.69 |
| 12 | 1.87 | 2.07 | 10.95 | 2.14 | 14.28 | 2.14 | 14.37 | 87.21 | 87.23 | 90.64 | 87.60 |
| 100.00 | 100.00 | 100.00 | 100.00 | ||||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trisurat, Y.; Sutummawong, N.; Roehrdanz, P.R.; Chitechote, A. Climate Change Impacts on Species Composition and Floristic Regions in Thailand. Diversity 2023, 15, 1087. https://doi.org/10.3390/d15101087
Trisurat Y, Sutummawong N, Roehrdanz PR, Chitechote A. Climate Change Impacts on Species Composition and Floristic Regions in Thailand. Diversity. 2023; 15(10):1087. https://doi.org/10.3390/d15101087
Chicago/Turabian StyleTrisurat, Yongyut, Nantida Sutummawong, Patrick R. Roehrdanz, and Auschada Chitechote. 2023. "Climate Change Impacts on Species Composition and Floristic Regions in Thailand" Diversity 15, no. 10: 1087. https://doi.org/10.3390/d15101087
APA StyleTrisurat, Y., Sutummawong, N., Roehrdanz, P. R., & Chitechote, A. (2023). Climate Change Impacts on Species Composition and Floristic Regions in Thailand. Diversity, 15(10), 1087. https://doi.org/10.3390/d15101087