
Invasion of Bythotrephes longimanus and Cercopagis pengoi in Lake Champlain: Impacts on the Native Zooplankton Community


Abstract
:1. Introduction
Potential Impact of the Cercopagidae Invasion
2. Materials and Methods
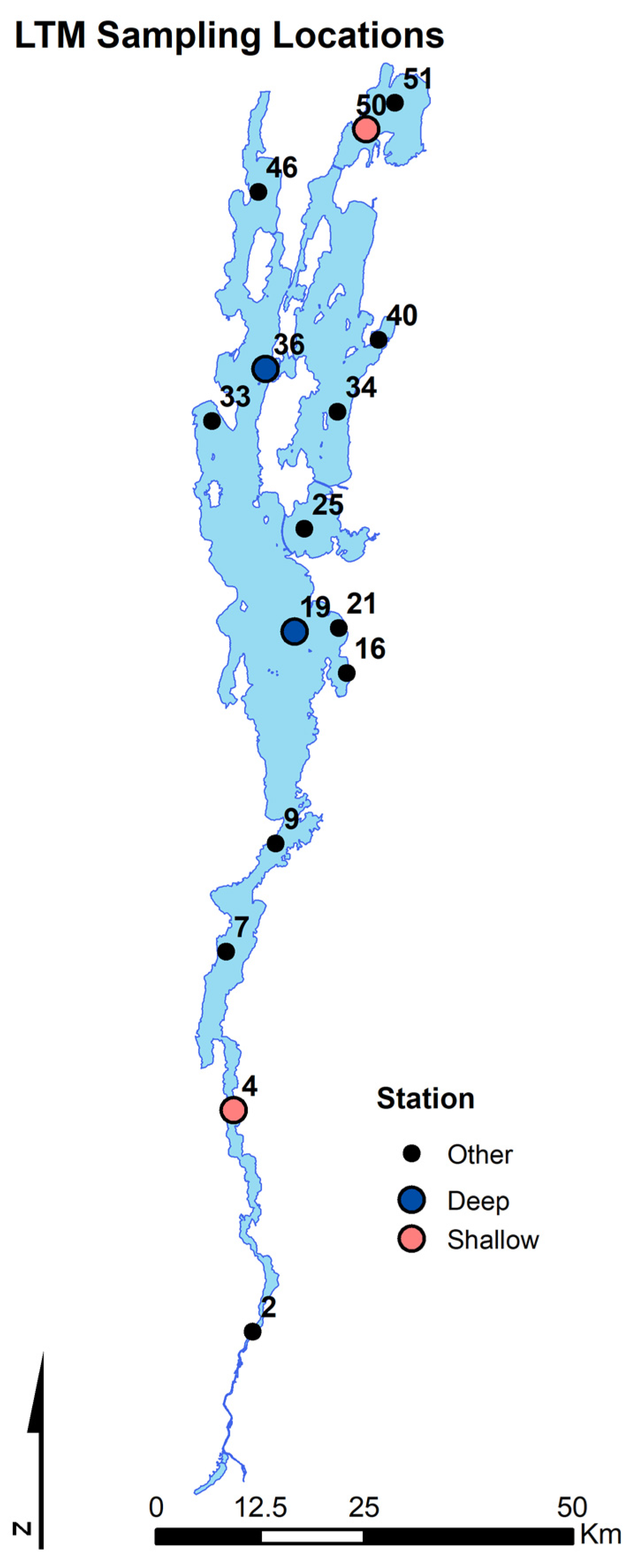
2.1. Zooplankton Community Sampling
2.2. Data Analysis
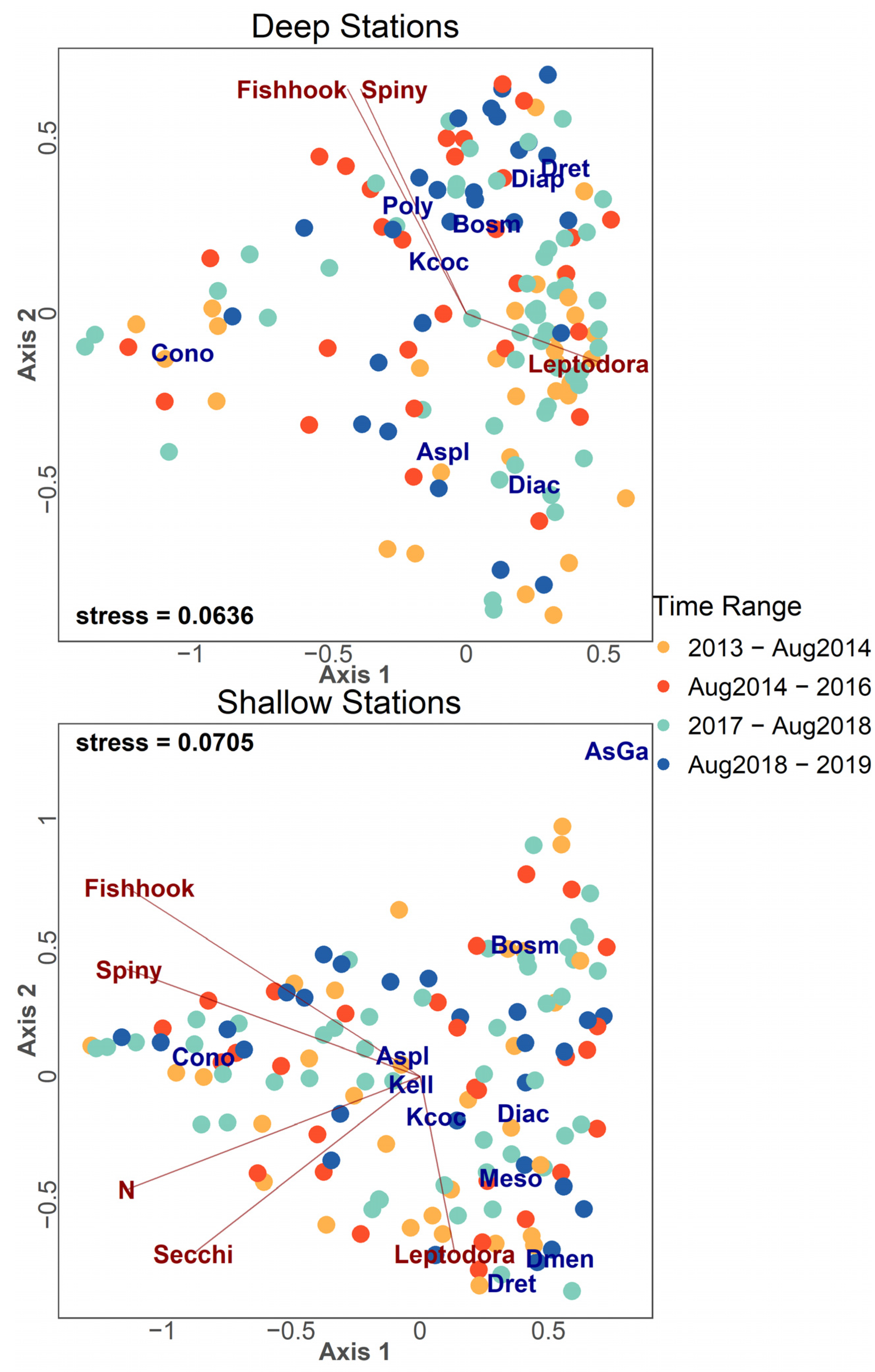
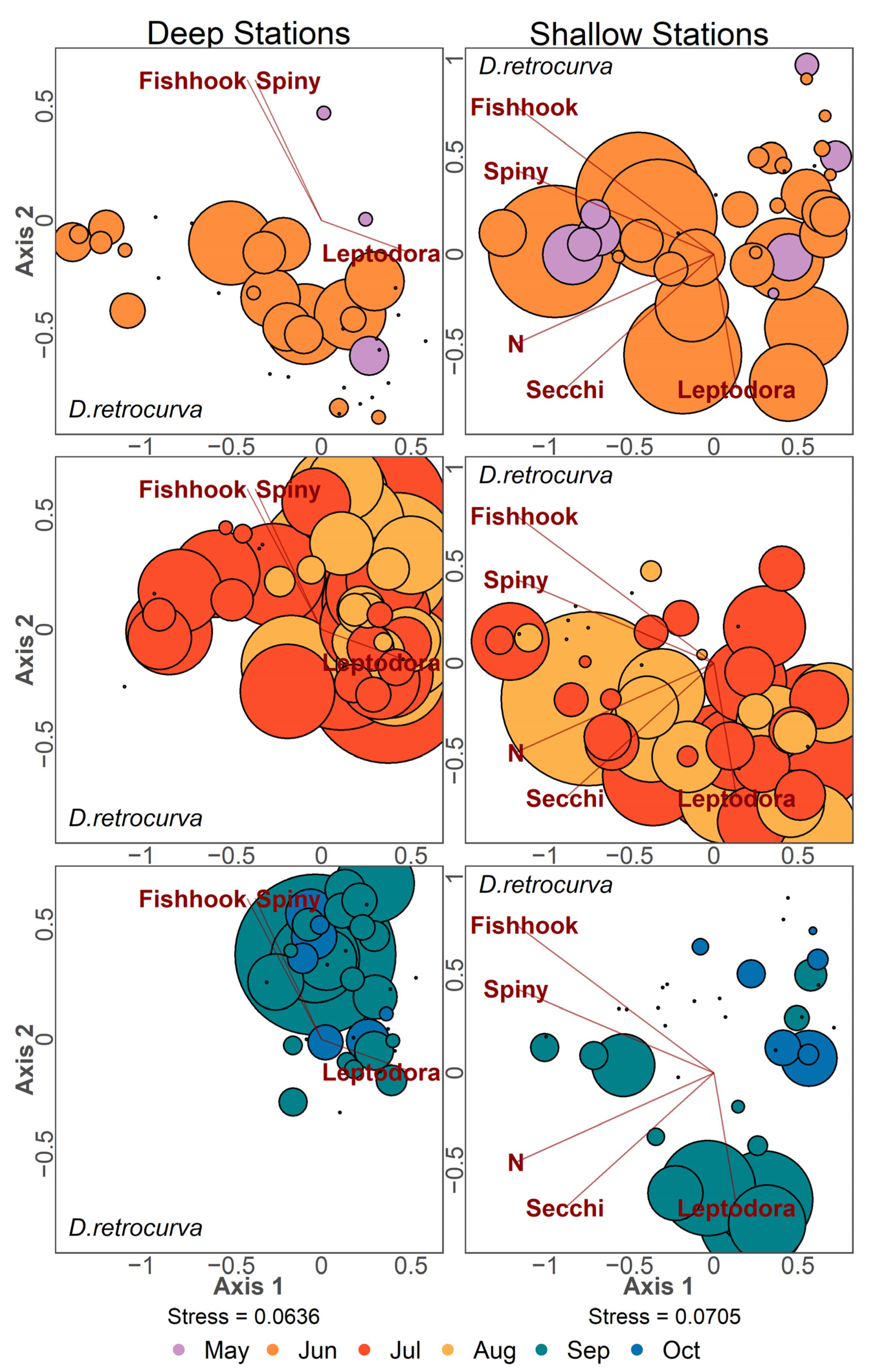
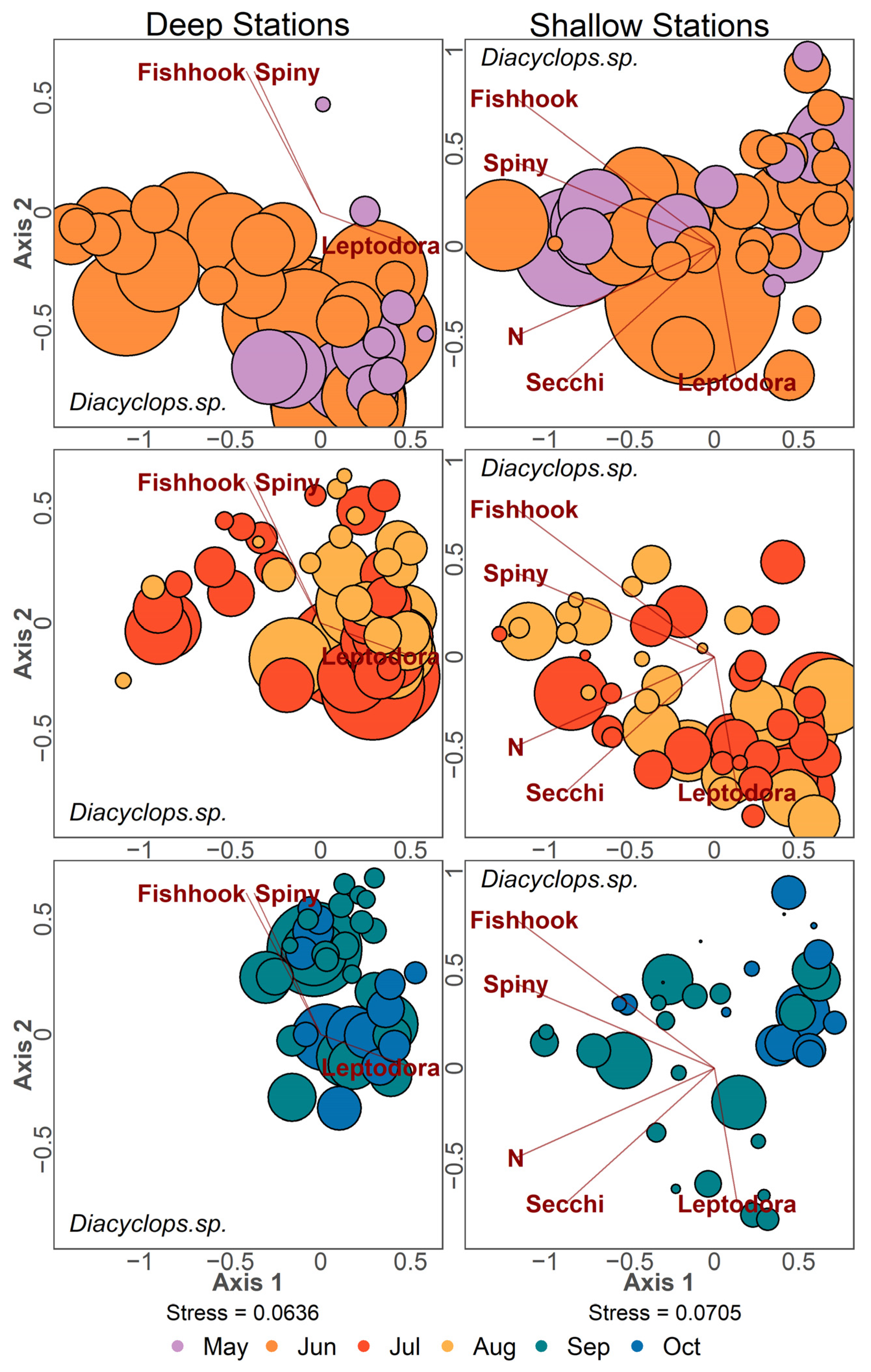
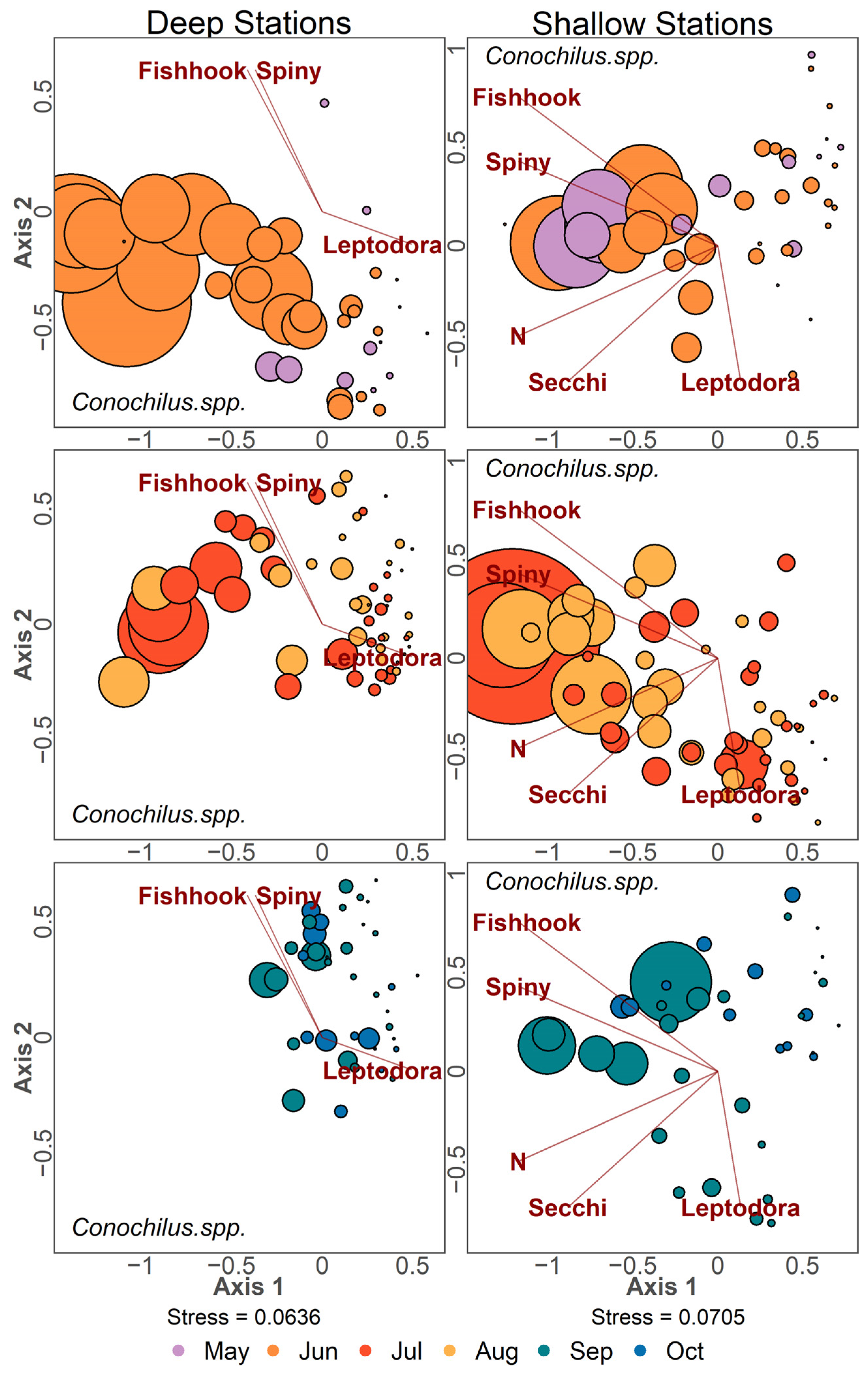
2.2.1. Ordination Analysis
2.2.2. Community Structure Analysis
3. Results
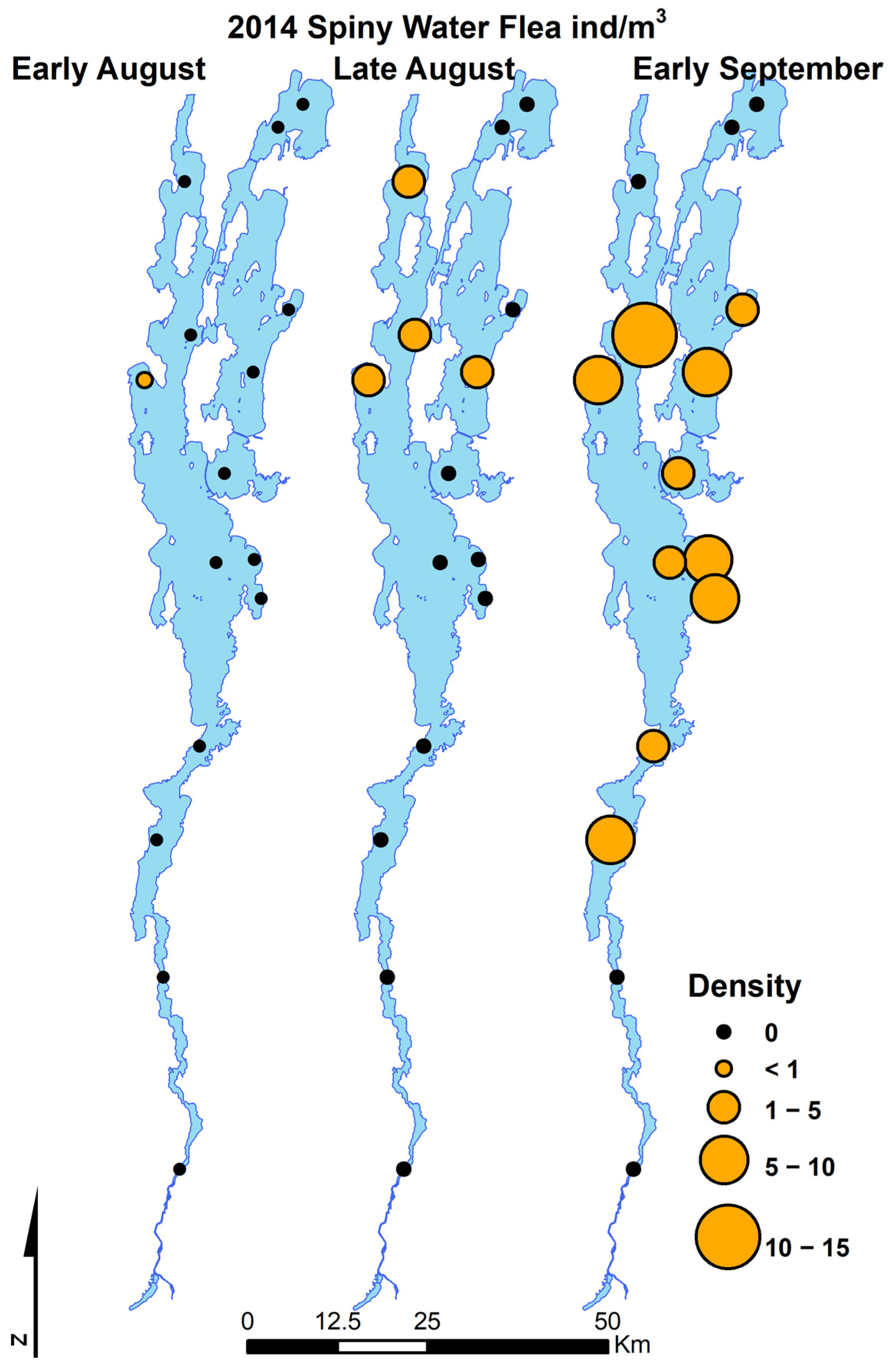
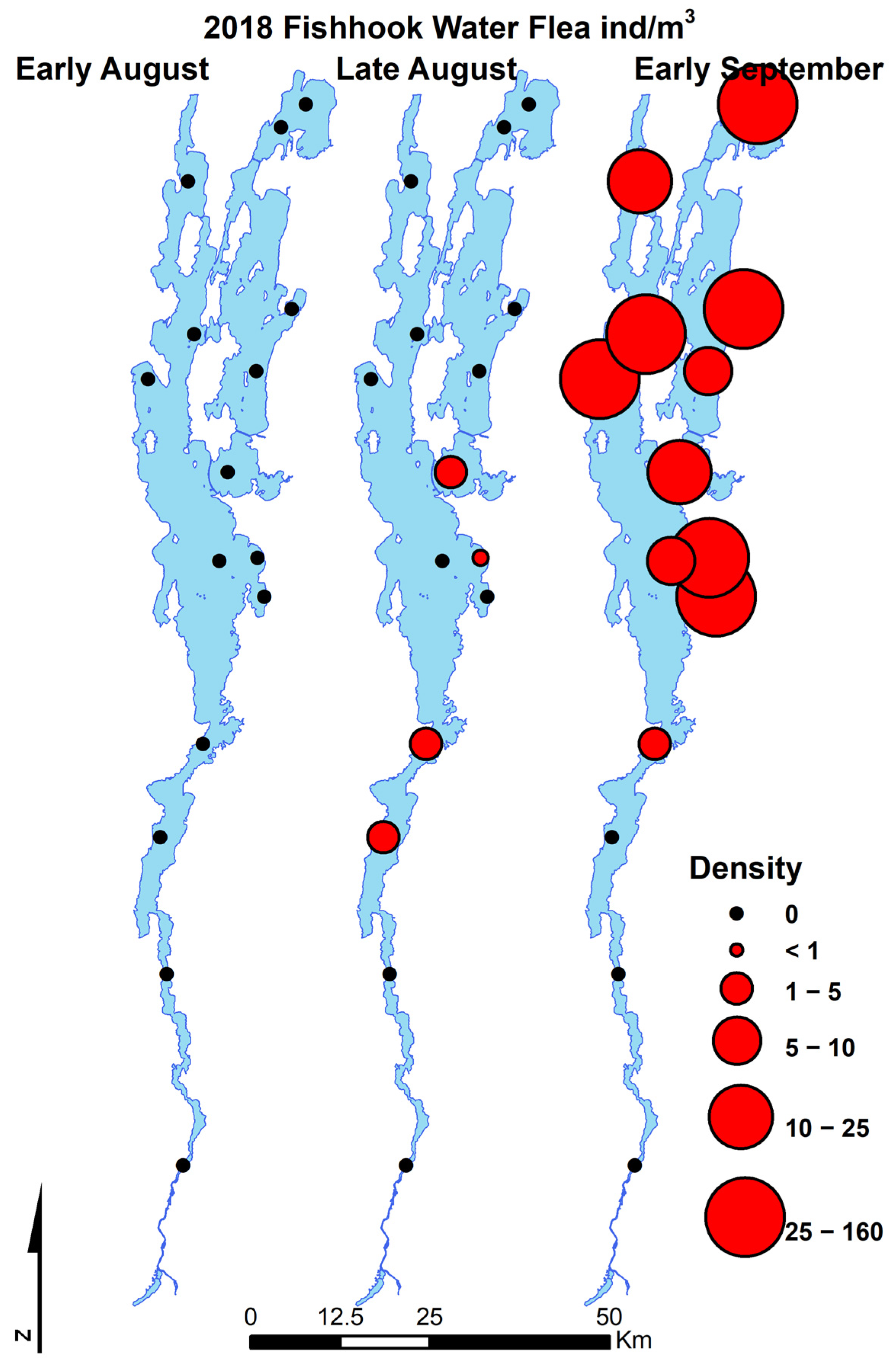
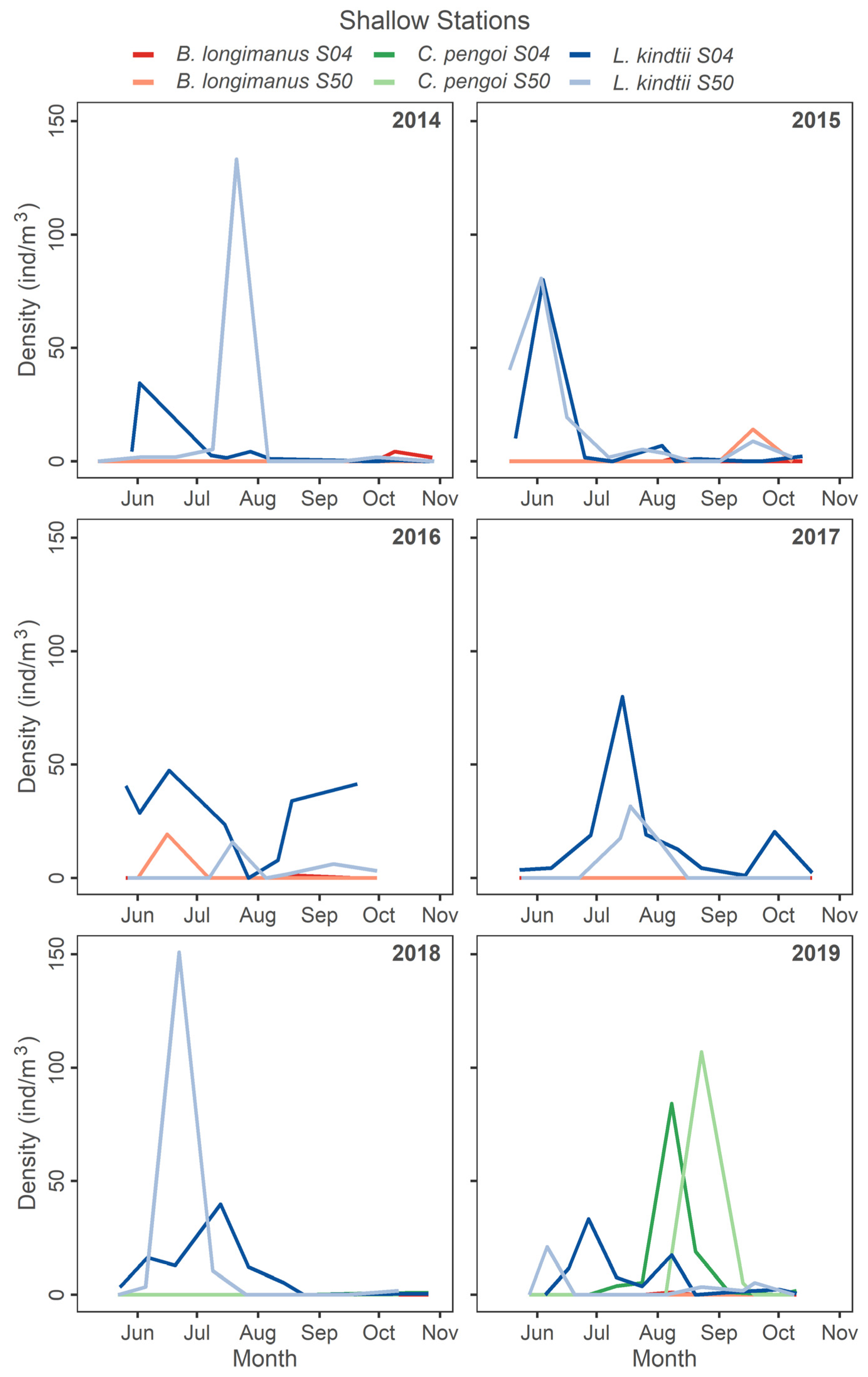
3.1. Invasion Dynamics in Lake Champlain
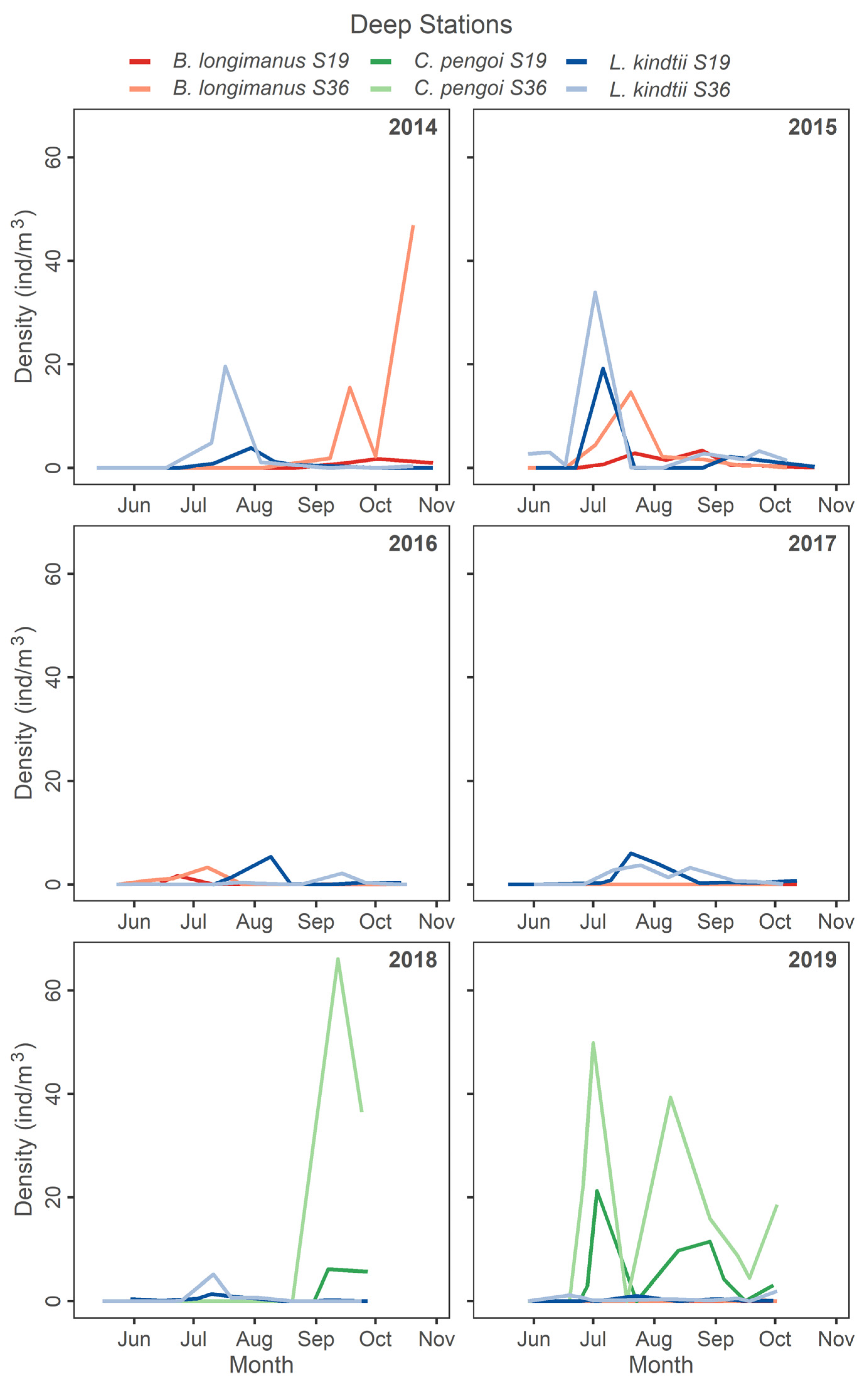
3.2. Response of Large Bodied Native L. kindtii
3.3. Zooplankton Abundance Patterns Following Invasion
3.4. Zooplankton Community Structure Following Invasion
3.5. Community Patterns Pre- and Post-Invasion
4. Discussion
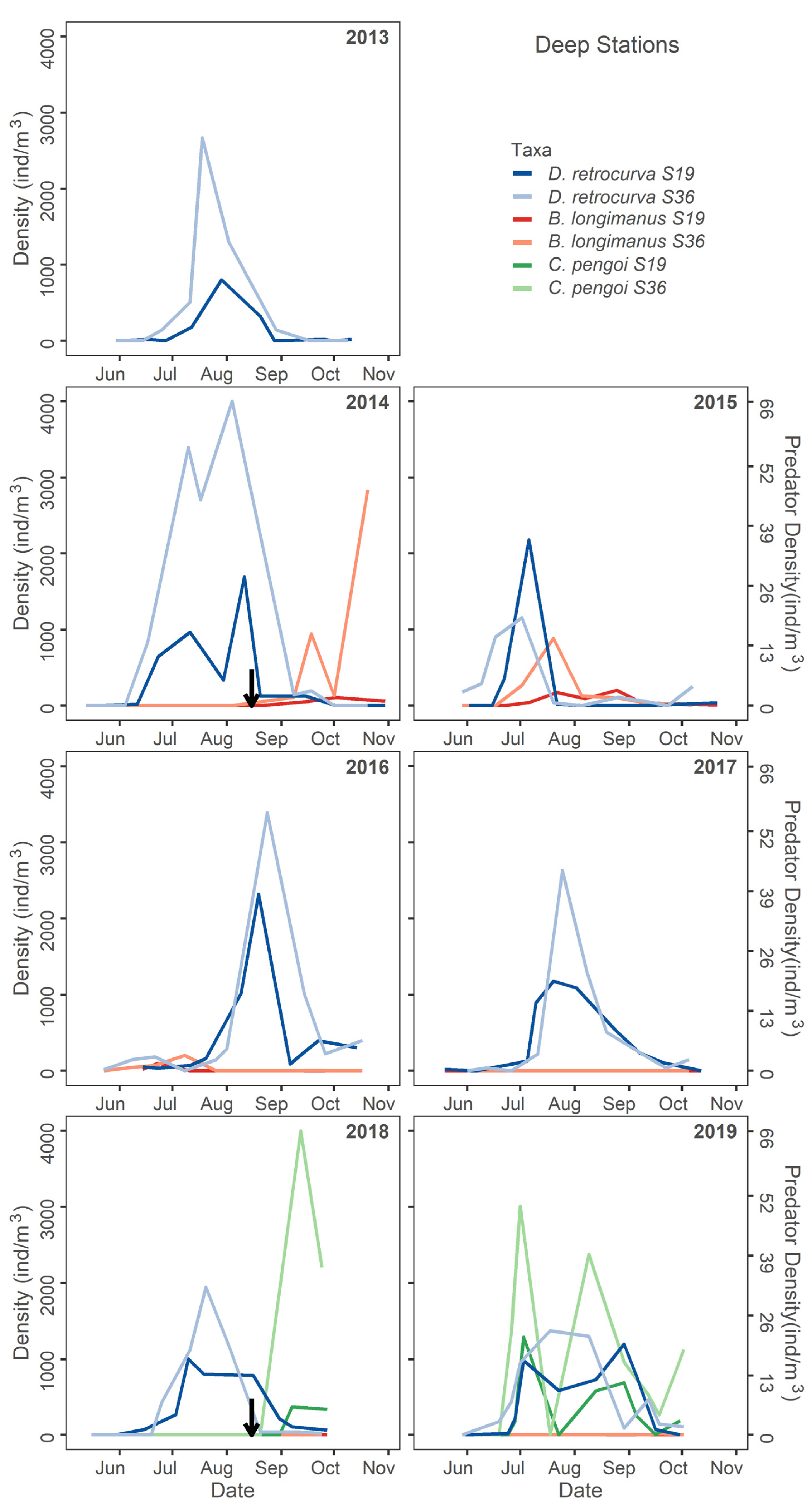
4.1. Large Predatory Cladoceran Patterns
4.2. Zooplankton Invasion Response Trends
4.3. Filter-Feeding Cladocerans
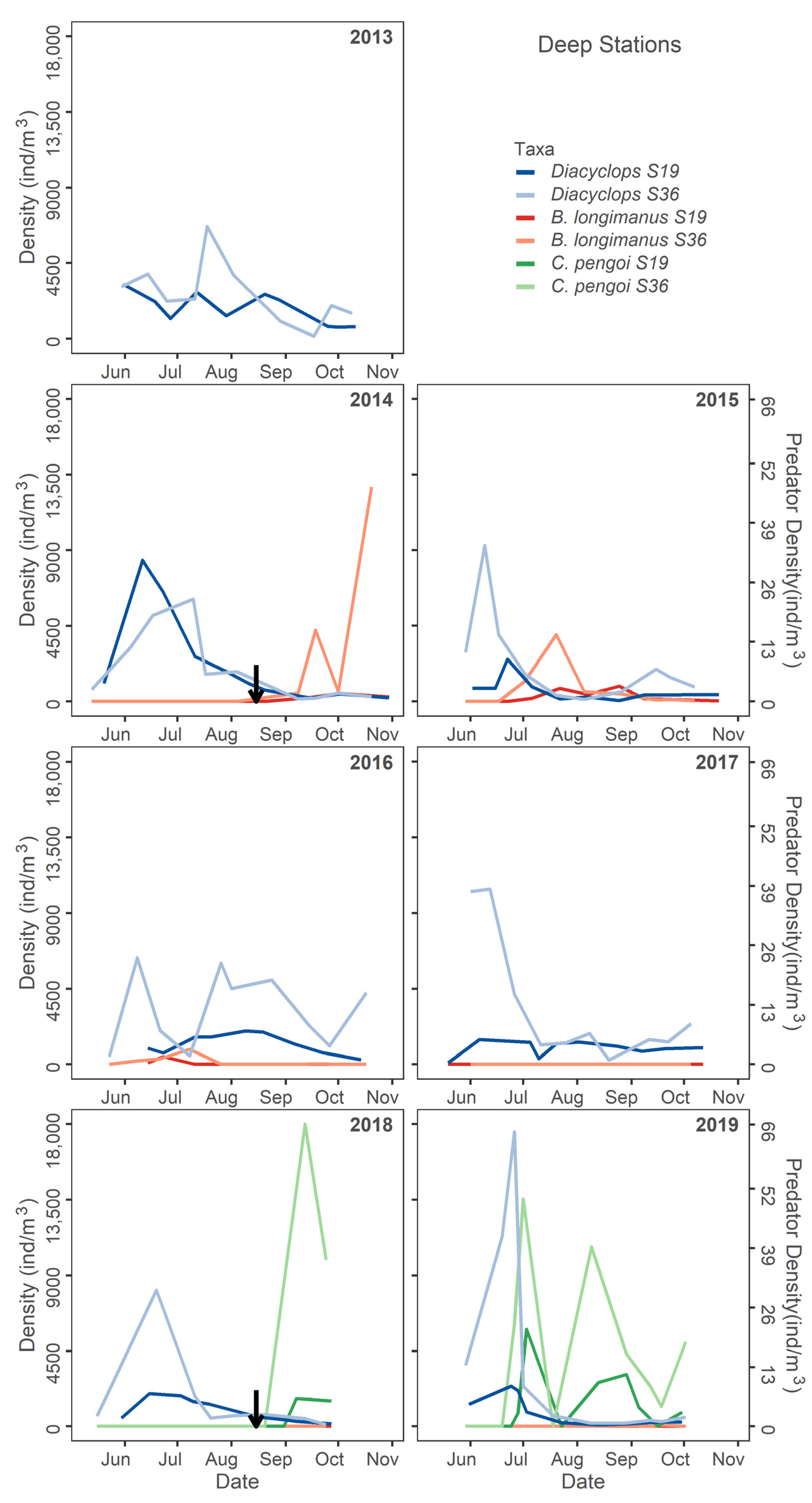
4.4. Copepods
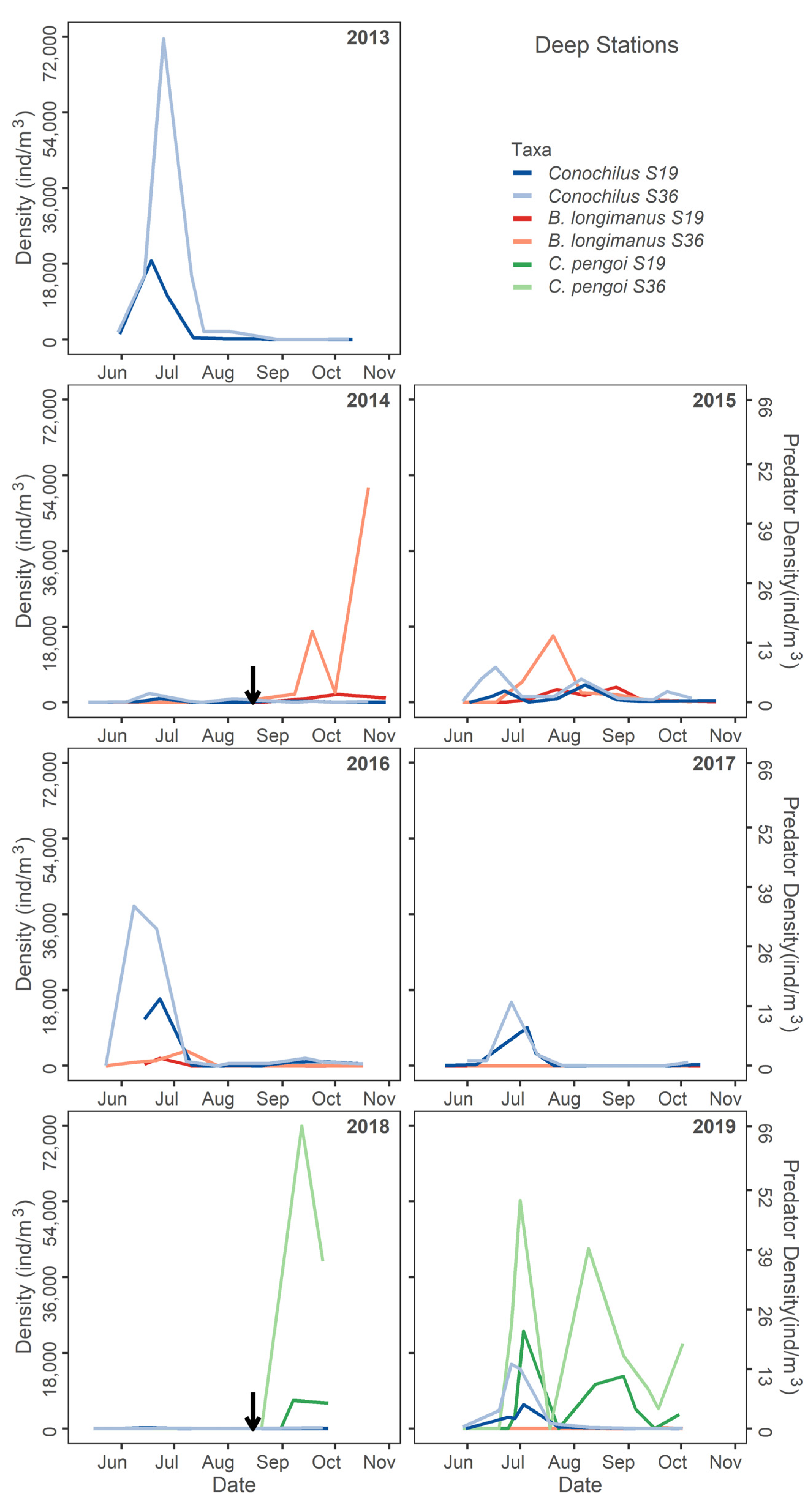
4.5. Rotifers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Escobar, L.; Mallez, S.; McCartney, M.; Lee, C.; Lielinski, D.; Ghosal, R.; Bajer, P.; Wagner, C.; Nash, B.; Tomamichel, M.; et al. Aquatic invasive species in the Great Lakes region: An overview. Rev. Fish. Sci. Aquac. 2017, 26, 121–138. [Google Scholar] [CrossRef]
- Sturtevant, R.; Mason, D.; Rutherford, E.; Elgin, A.; Lower, E.; Martinez, F. Recent history of nonindigenous species in the Laurentian Great Lakes; An update to Mills et al., 1993 (25 years later). J. Great Lakes Res. 2019, 45, 1011–1035. [Google Scholar] [CrossRef]
- Marsden, J.E.; Hauser, M. Exotic species in Lake Champlain. J. Great Lakes Res. 2009, 35, 250–265. [Google Scholar] [CrossRef]
- Mihuc, T.; Dunlap, F.; Binggeli, C.; Myers, L.; Pershyn, C.; Groves, A.; Waring, A. Long-term patterns in Lake Champlain’s zooplankton: 1992–2010. J. Great Lakes Res. 2012, 38, 49–57. [Google Scholar] [CrossRef]
- Mihuc, T.B.; Recknagel, F. Lessons from Bioinvasion of Lake Champlain, U.S.A. In Ecological Informatics; Recknagel, F., Michener, W., Eds.; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Benoit, H.; Johannsson, O.; Warner, D.; Sprules, W.; Rudstam, L. Assessing the impact of a recent predatory invader: The population dynamics, vertical distribution, and potential prey of Cercopagis pengoi in Lake Ontario. Limnol. Oceanogr. 2002, 47, 626–635. [Google Scholar] [CrossRef]
- Boudreau, S.; Yan, N. The differing crustacean zooplankton communities of Canadian Shield lakes with and without the nonindigenous zooplanktivore Bythotrephes longimanus. Can. J. Aquat. Sci. 2003, 60, 1307–1313. [Google Scholar] [CrossRef]
- Kim, N.; Yan, N. Food limitation impacts life history of the predatory cladoceran Bythotrephes longimanus, an invader to North America. Hydrobiologia 2013, 715, 213–224. [Google Scholar] [CrossRef]
- Pawlowski, M.; Branstrator, D.; Hrabik, T.; Sterner, R. Changes in the cladoceran community of Lake Superior and the role of Bythotrephes longimanus. J. Great Lakes Res. 2017, 43, 1101–1110. [Google Scholar] [CrossRef]
- Cristescu, M.; Hebert, P.; Witt, J.; MacIsaac, H.; Grigorovich, I. An invasion history for Cercopagis pengoi based on mitochondrial gene sequences. Limnol. Oceanogr. 2001, 46, 224–229. [Google Scholar] [CrossRef]
- Muirhead, J.; Sprules, G. Reaction distance of Bythotrephes longimanus, encounter rate and index of prey risk for Harp Lake, Ontario. Freshw. Biol. 2003, 48, 135–146. [Google Scholar] [CrossRef]
- Pichlova-Ptacnikova, R.; Vanderploeg, H. The invasive cladoceran Cercopagis pengoi is a generalist predator capable of feeding on a variety of prey species of different sizes and escape abilities. Fundam. Appl. Limnol. Arch. Fur Hydrobiol. 2009, 173, 267–279. [Google Scholar] [CrossRef]
- Grigorovich, I.; Pashkova, O.; Gromova, Y.; Overdijk, C. Bythotrephes longimanus in the Commonwealth of Independent States: Variability, distribution and ecology. Hydrobiologia 1998, 379, 183–198. [Google Scholar] [CrossRef]
- Jokela, A.; Arnott, S.; Beisner, B. Patterns of Bythotrephes longimanus distribution relative to native macroinvertebrates and zooplankton prey. Biol. Invasions 2011, 13, 2573–2594. [Google Scholar] [CrossRef]
- Lehman, J.; Caceres, C. Food-web responses to species invasion by a predatory invertebrate: Bythotrephes in Lake Michigan. Limnol. Oceanogr. 1993, 38, 879–891. [Google Scholar] [CrossRef]
- Ojaveer, H.; Kuhns, L.; Barbiero, R.; Tuchman, M. Distribution and population characteristics of Cercopagis pengoi in Lake Ontario. J. Great Lakes Res. 2001, 27, 10–18. [Google Scholar] [CrossRef]
- Pichlova-Ptacnikova, R.; Vanderploeg, H.; Cavaletto, J. Big versus small: Does Bythotrephes longimanus predation regulate spatial distribution of another invasive predatory cladoceran, Cercopagis pengoi? J. Great Lakes Res. 2015, 41, 143–149. [Google Scholar] [CrossRef]
- VTDEC; NYDEC. Lake Champlain Long-Term Water Quality and Biological Monitoring Program: Summary of Program Activities During 2012. Lake Champlain Basin Program. 2012. Available online: https://dec.vermont.gov/sites/dec/files/wsm/lakes/docs/lp_2012lcmannualreport.pdf (accessed on 23 October 2023).
- Barbiero, R.; Tuchman, M. Changes in the crustacean communities of Lakes Michigan, Huron, and Erie following the invasion of the predatory cladoceran Bythotrephes longimanus. Can. J. Fish. Aquat. Sci. 2004, 61, 2111–2125. [Google Scholar] [CrossRef]
- Yan, N.; Leung, B.; Lewis, M.; Peacor, S. The spread, establishment and impacts of the spiny water flea, Bythotrephes longimanus, in temperate North America: A synopsis of the special issue. Biol. Invasions 2011, 13, 2423–2432. [Google Scholar] [CrossRef]
- Barbiero, R.; Warren, G. Rotifer communities in the Laurentian Great Lakes. 1983–2006 and the factors affecting their composition. J. Great Lake Res. 2011, 37, 528–540. [Google Scholar] [CrossRef]
- Laxson, C.; McPhedran, K.; Makarewicz, J.; Telesh, I.; MacIsaac, H. Effects of the non-indigenous cladoceran Cercopagis pengoi on the lower food web of Lake Ontario. Freshw. Biol. 2003, 48, 2094–2106. [Google Scholar] [CrossRef]
- Facey, D.; Marsden, E.; Mihuc, T.; Howe, E. Lake Champlain 2010: A summary of recent research and monitoring initiatives. J. Great Lakes Res. 2012, 38, 1–5. [Google Scholar] [CrossRef]
- Young, J.; Strecker, A.; Yan, N. Increased abundance of the non-indigenous zooplanktivore, Bythotrephes longimanus, is strongly correlated with greater spring prey availability in Canadian Shield lakes. Biol. Invasions 2011, 13, 2605–2619. [Google Scholar] [CrossRef]
- Bowen, K.; Johannsson, O. Changes in zooplankton biomass in the Bay of Quinte with the arrival of the mussels, Dreissena polymorpha and D. rostiformis bugensis, and the predatory cladoceran, Cercopagis pengoi: 1975 to 2008. Aquat. Ecosyst. Health Manag. 2011, 14, 44–55. [Google Scholar] [CrossRef]
- Pichlova-Ptacnikova, R.; Vanderploeg, H. The quick and the dead: Might differences in escape rates explain the changes in the zooplankton community composition of Lake Michigan after invasion by Bythotrephes? Biol. Invasions 2011, 13, 2595–2604. [Google Scholar] [CrossRef]
- Branstrator, D.; Lehman, J. Invertebrate predation in Lake Michigan: Regulation of Bosmina longirostris by Leptodora kindtii. Limnol. Oceanogr. 1991, 36, 483–495. [Google Scholar] [CrossRef]
- LaMay, M. Diel Vertical Migration of Zooplankton in a Lake Containing the Invasive Bythotrephes longimanus (Lake George) Compared to a Non-Invaded Lake (Lake Champlain). Master’s Thesis, State University of New York at Plattsburgh, Plattsburgh, NY, USA, 2014; p. 157. [Google Scholar]
- Witt, A.; Dettmers, J.; Caceres, C. Cercopagis pengoi in Southwestern Lake Michigan in four years following invasion. J. Great Lakes Res. 2005, 31, 245–252. [Google Scholar] [CrossRef]
- Gorokhova, E. Zooplankton spatial distribution and potential predation by invertebrate zooplanktivores. In Proceedings of the Second BAYSYS Annual Science Conference, Stockholm, Sweden, 23–25 September 1998. Paper Abstracts: 7. [Google Scholar]
- Makarewicz, J.; Lewis, T. Long-term changes in Lake Ontario rotifer abundance and composition: A response to Cercopagis predation? J. Great Lakes Res. 2015, 41, 192–199. [Google Scholar] [CrossRef]
- Hovius, J.; Beisner, B.; McCann, K. Epilimnetic rotifer community responses to Bythotrephes longimanus invasion in Canadian Shield lakes. Limnol. Oceanogr. 2006, 51, 1004–1012. [Google Scholar] [CrossRef]
- Cavaletto, J.; Vanderploeg, H.; Ptacnikova, R.; Pothoven, S.; Liebig, J.; Fahnenstiel, G. Temporal and spatial separation allow coexistence of predatory cladocerans: Leptodora kindtii. Bythotrephes longimanus and Cercopagis pengoi in southeastern Lake Michigan. J. Great Lakes Res. 2010, 36, 65–73. [Google Scholar] [CrossRef]
- Enz, C.; Heller, C.; Muller, R.; Burgi, H. Investigations on fecundity of Bythotrephes longimanus in Lake Lucerne (Switzerland) and on niche segregation of Leptodora kindtii and Bythotrephes longimanus in Swiss lakes. Hytrobiologia 2001, 464, 143–151. [Google Scholar] [CrossRef]
- Kane, D.; Haas, E.; Culver, D. The characteristics and potential ecological effects of the exotic crustacean zooplankter Cercopagis pengoi (Cladocera: Cercopagidae), a recent invader of Lake Erie. Ohio J. Sci. 2003, 103, 79–83. [Google Scholar]
- Branstrator, D. Ecological interactions between Bythotrephes longimanus and Leptodora kindtii and the implications for species replacement in Lake Michigan. J. Great Lakes Res. 1995, 21, 670–679. [Google Scholar] [CrossRef]
- Johannsson, O.; Bowen, K. Zooplankton production in the Bay of Quinte 1975–2008: Relationships with primary production.habitat. planktivory. and aquatic invasive species (Dreissena spp. and Cercopagis pengoi). Can. J. Fish. Aquat. Sci. 2012, 69, 2046–2063. [Google Scholar] [CrossRef]
- Lake Champlain Basin Program. Long-term water quality and biological monitoring project for Lake Champlain. In Proceedings of the 2018–2023 Quality Assurance Project Plan/Workplan, Grand Isle, VT, USA, 5 June 2020; Vermont/New York Department of Environmental Conservation Watershed Management Division/Division of Water, Lake Champlain Basin Program (Vermont). Lake Champlain Basin Program, 54 West Shore Road Grand Isle, Vermont. p. 68. [Google Scholar]
- Hayes-Pontius, E.M. Zooplankton Community Dynamics Following Extreme Flood Events in Lake Champlain. Master’s Thesis, State University of New York at Plattsburgh, Plattsburgh, NY, USA, 2013; p. 103. [Google Scholar]
- LaMay, M.; Hayes-Pontius, E.; Ater, I.; Mihuc, T. A revised key to the zooplankton of Lake Champlain. State Univ. N. Y. Plattsbg. 2013, 6, 1–41. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing. Vienna, Austria. 2023. Available online: https://www.R-project.org/ (accessed on 23 October 2023).
- Oksanen, J.; Simpson, G.; Blanchet, F.; Kindt, R.; Legendre, P.; Minchin, P.; O’Hara, R.; Solymos, P.; Stevens, M.; Szoecs, E.; et al. _Vegan: Community Ecology Package_. R Package Version 2.6-4. 2022. Available online: https://CRAN.R-project.org/package=vegan (accessed on 23 October 2023).
- Clarke, K.; Warwick, R. Quantifying structural redundancy in ecological communities. Oecologia 1998, 113, 278–289. [Google Scholar] [CrossRef]
- Taylor, M. _sinkr: Collection of Functions with Emphasis in Multivariate Data Analysis_. R Package Version 0.7. 2022. Available online: https://github.com/marchtaylor/sinkr (accessed on 23 October 2023).
- Vanderploeg, H.; Pothoven, S.; Fahnestiel, G.; Cavaletto, J.; Liebig, J.; Stow, C.; Nalepa, T.; Madenjian, C.; Bunnell, D. Seasonal zooplankton dynamics in Lake Michigan: Disentangling impacts of resource limitation, ecosystem engineering, and predation during a critical ecosystem transition. J. Great Lakes Res. 2012, 38, 336–352. [Google Scholar] [CrossRef]
- Berg, D.; Garton, D. Seasonal abundance of the exotic predatory cladoceran Bythotrephes longimanus, in western Lake Erie. J. Great Lakes Res. 1988, 14, 479–488. [Google Scholar] [CrossRef]
- Branstrator, D. Contrasting life histories of the predatory cladocerans Leptodora kindtii and Bythotrephes longimanus. J. Plankton Res. 2005, 27, 569–585. [Google Scholar] [CrossRef]
- Mills, E.; Gorman, R.; DeGisi, J.; Heberger, R.; House, R. Food of the alewife (Alosa pseudoharengus) in Lake Ontario before and after the establishment of Bythtrephes longimanus. Can. J. Fish Aquat. Sci. 1992, 49, 2009–2019. [Google Scholar] [CrossRef]
- Martin, B.; Mrnak, J.; Vander Zanden, J. Evaluating the potential role of predation by native fish regulating the abundance of invasive spiny water flea. J. Freshw. Ecol. 2023, 38, 2187470. [Google Scholar] [CrossRef]
- Pothoven, S.; Vanderploeg, H.; Cavaletto, J.; Krueger, D.; Mason, D.; Brandt, S. Alewife planktivory controls the abundance of two invasive predatory cladocerans in Lake Michigan. Freshw. Biol. 2007, 52, 561–573. [Google Scholar] [CrossRef]
- Makarewicz, J.; Damaske, E.; Laxson, C. Seasonal and Vertical Distribution, Food Web Dynamics and Contaminant Biomagnification of Cercopagis pengoi in Lake Ontario. SUNY Open Access Repository (SOAR). Brockport Technical Reports (Water Resources). 2002. Available online: https://soar.suny.edu/handle/20.500.12648/4437 (accessed on 23 October 2023).
- Sorenson, M.; Branstrator, D. The North American invasive zooplanktivore Bythotrephes longimanus is less hypoxia-tolerant than the native Leptodora kindtii. Can. J. Fish Aquat. Sci. 2016, 74, 824–832. [Google Scholar] [CrossRef]
- Yan, N.; Pawson, T. Changes in the crustacean zooplankton community of Harp Lake, Canada, following invasion by Bythotrephes longimanus. Freshw. Biol. 1997, 37, 409–425. [Google Scholar] [CrossRef]
- Barbiero, R.; Rudstam, L.; Watkins, J.; Lesht, B. A cross-lake comparison of crustacean zooplankton communities in the Laurentian Great Lakes, 1997–2016. J. Great Lakes Res. 2019, 45, 672–690. [Google Scholar] [CrossRef]
- Bunnell, D.; Keeler, K.; Puchala, E.; Davis, B.; Pothoven, S. Comparing seasonal dynamics of the Lake Huron zooplankton community between 1983–1984 and 2007 and revisiting the impact of Bythotrephes planktivory. J. Great Lakes Res. 2012, 38, 451–462. [Google Scholar] [CrossRef]
- Yan, N.; Blukacz, A.; Sprules, G.; Kindy, P.; Hackett, D.; Girard, R.; Clark, B. Changes in zooplankton and the phenology of the spiny water flea, Bythotrephes, following its invasion of Harp Lake, Ontario, Canada. Can. J. Fish. Aquat. Sci. 2001, 58, 2341–2350. [Google Scholar] [CrossRef]
- Kerfoot, W.; Hobmeier, M.; Yousef, F.; Lafrancois, B.; Maki, R.; Hirsch, J. A plague of waterfleas (Bythotrephes): Impacts on microcrustacean community structure, seasonal biomass, and secondary production in a large inland-lake complex. Biol. Invasions 2016, 18, 1121–1145. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Station | Month | Fishhook Impact | Spiny Impact |
|---|---|---|---|
| 4 | July | 0.672 | 0.965 |
| August | 0.029 | 0.242 | |
| September | 0.854 | 0.107 | |
| 19 | July | 0.313 | 0.864 |
| August | 0.031 | 0.934 | |
| September | 0.134 | 0.016 | |
| 36 | July | 0.429 | 0.168 |
| August | 0.902 | 0.221 | |
| September | 0.022 | 0.302 | |
| 50 | July | 0.051 | 0.281 |
| August | 0.012 | 0.008 | |
| September | 0.082 | 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cutter, Z.A.; Mihuc, T.B.; Myers, L.W. Invasion of Bythotrephes longimanus and Cercopagis pengoi in Lake Champlain: Impacts on the Native Zooplankton Community. Diversity 2023, 15, 1112. https://doi.org/10.3390/d15111112
Cutter ZA, Mihuc TB, Myers LW. Invasion of Bythotrephes longimanus and Cercopagis pengoi in Lake Champlain: Impacts on the Native Zooplankton Community. Diversity. 2023; 15(11):1112. https://doi.org/10.3390/d15111112
Chicago/Turabian StyleCutter, Zachary A., Timothy B. Mihuc, and Luke W. Myers. 2023. "Invasion of Bythotrephes longimanus and Cercopagis pengoi in Lake Champlain: Impacts on the Native Zooplankton Community" Diversity 15, no. 11: 1112. https://doi.org/10.3390/d15111112
APA StyleCutter, Z. A., Mihuc, T. B., & Myers, L. W. (2023). Invasion of Bythotrephes longimanus and Cercopagis pengoi in Lake Champlain: Impacts on the Native Zooplankton Community. Diversity, 15(11), 1112. https://doi.org/10.3390/d15111112