
Responses of Phytoplankton Communities to Flow Regulation in Northeastern Riverine Wetlands of China


Abstract
:1. Introduction
2. Study Design
2.1. Study Area
2.2. Sampling Design
2.3. Sample Handling
2.4. Functional Traits
2.5. Data Analysis
3. Results
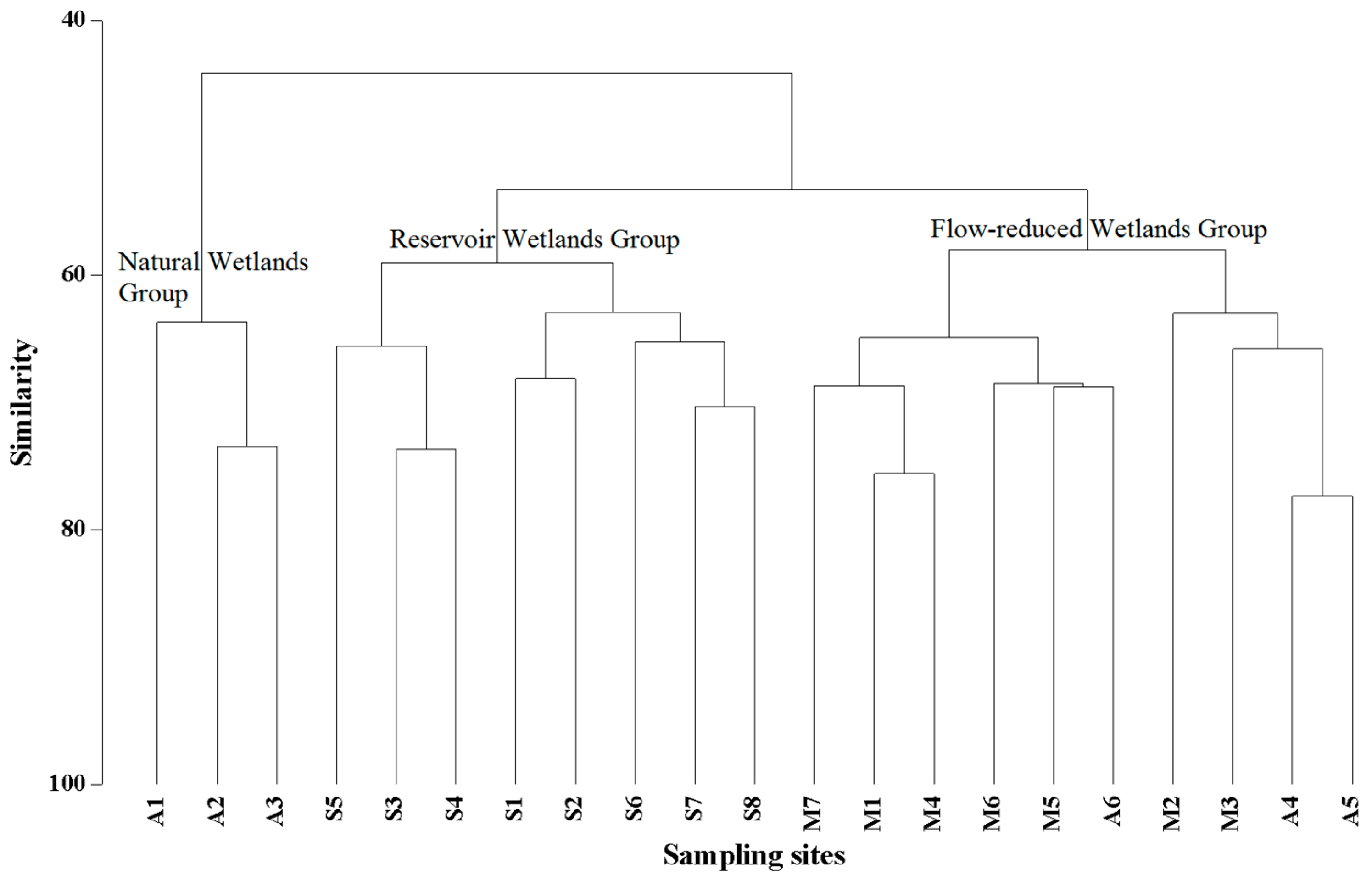
3.1. Phytoplankton Community Assemblage Structure
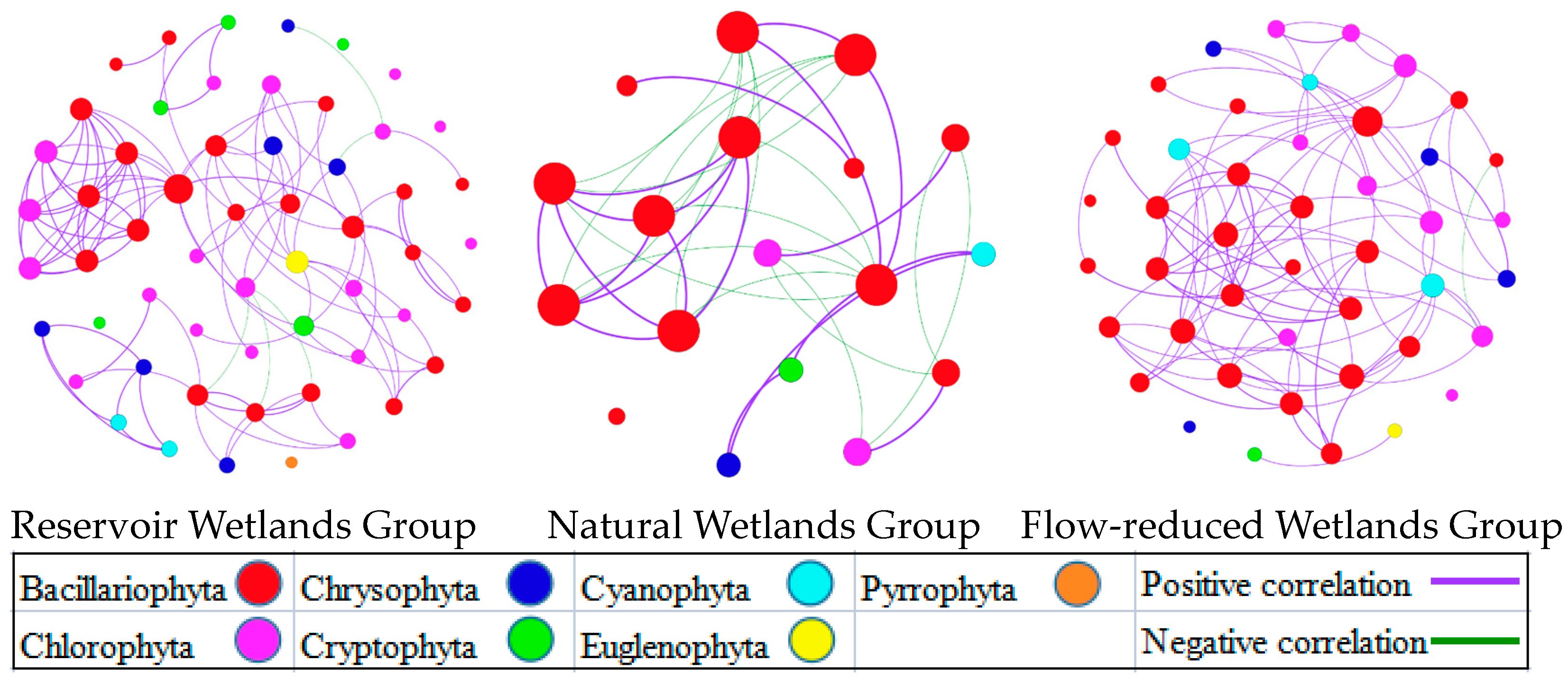
3.2. Phytoplankton Community Functional Group
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muhammad, B.L.; Kim, T.; Ki, J.S. 18S rRNA analysis reveals high diversity of phytoplankton with emphasis on a naked Dinoflflagellate Gymnodinium sp. at the Han River (Korea). Diversity 2021, 13, 73. [Google Scholar] [CrossRef]
- Guo, J.; Zheng, Y.Y.; Teng, J.H.; Wang, X.M.; Song, J.X. Characteristics of spatial distribution for microbial ecology inside and outside source water reservoir. J. Clean Prod. 2021, 311, 127697. [Google Scholar] [CrossRef]
- Rui, P.R.; Feio, M.J.; Almeida, S.F.P.; Vieira, C.; Aguiar, F.C. Multi-biologic group analysis for an ecosystem response to longitudinal river regulation gradients. Sci. Total Eviron. 2021, 767, 144327. [Google Scholar] [CrossRef]
- Kim, D.Y.; Ji, R.B.; Feng, Z.X.; Jang, J.B.; Lee, D.I.; Lee, W.C.; Kang, C.K. Estuarine dam water discharge enhances summertime primary productivity near the southwestern Korean coast. Mar. Pollut. Bull. 2023, 191, 114971. [Google Scholar] [CrossRef]
- Meng, Y.; Wu, H.T.; Guan, Q.; Lu, K.L.; Kang, Y.J.; Dai, G.H. Responses of riparian insect communities to flow regulation in riverine wetlands of Northeastern China. Hydrobiologia 2022, 850, 3865–3877. [Google Scholar] [CrossRef]
- Korneva, L.G.; Solovyeva, V.V.; Sakharova, E.G. On the distribution of Peridiniopsis kevei Grigor. et Vasas (Dinophyta) in the upper Volga reservoirs. Inland Water Biol. 2015, 8, 414–416. [Google Scholar] [CrossRef]
- Wu, Q.; Li, Q.H.; Luo, H.; Chen, Q.; Chen, H.X.; Dong, Y.J.; Li, S.H. Comparison in phytoplankton diversity-productivity-community stability between river-type reservoir and lake-type reservoir. J. Oceanol Limnol. 2022, 40, 1485–1507. [Google Scholar] [CrossRef]
- dos Santos, N.C.L.; Garcia, B.E.; Dias, J.D.; Lopes, T.M.; Affonso, I.D.; Severi, W.; Gomes, L.C.; Agostinho, A.A. Cumulative ecological effects of a Neotropical reservoir cascade across multiple assemblages. Hydrobiologia 2018, 819, 77–91. [Google Scholar] [CrossRef]
- Yue, Y.H.; Fu, Z.W.; Chen, X.P.; Yang, M.; Wang, B.L.; Wang, F.S. Community structure and diversity phytoplankton in the Wujiang River Basin reservoir. J. Shanghai University (Nat. Sci. Edi.) 2021, 27, 97–105. [Google Scholar]
- Zhang, H.X.; Huo, S.L.; Cao, X.H.; Ma, C.Z.; Zhang, J.T.; Wu, F.C. Homogenization of reservoir eukaryotic algal and cyanobacterial communities is accelerated by dam construction and eutrophication. J. Hydrol. 2021, 603, 126842. [Google Scholar] [CrossRef]
- Kamidis, N.; Koutrakis, E.; Sapounidis, A.; Sylaios, G. Impact of river damming on downstream hydrology and hydrochemistry: The case of lower Nestos River catchment (NE. Greece). Water 2021, 13, 2832. [Google Scholar] [CrossRef]
- Korneva, L.G.; Solovyeva, V.V. Dynamics of morphofuncitonal groups of phytoplankton in the Rybinsk Reservoir and assessment of the Reservoir water quality by the community index. Water Resour. 2021, 48, 65–72. [Google Scholar] [CrossRef]
- Sirunda, J.; Oberholster, P.; Wolfaardt, G.; Truter, C.; van der Merwe, S. Long-term study of the drought impact on the phytoplankton concentration and assemblages in two water supply reservoirs in Namibia. Afr. J. Aquat. Sci. 2023, 48, 152–165. [Google Scholar] [CrossRef]
- Kumar, J.; Alam, A.; Borah, S.; Gogoi, P.; Das, B.K. Influence of barrages on hydrological parameters and functional groups of phytoplankton of a subtropical river Ganga, India. Environ. Sci. Pollut. R. 2023, 30, 77469–77487. [Google Scholar] [CrossRef] [PubMed]
- Caio, G.R.; Janne, S.; Gilsineia, C.; Felipe, S.P.; Marcela, M.; Patricia, D.; Marcelo, M.M. Functional rather than taxonomic diversity reveals changes in the phytoplankton community of a large dammed river. Ecol. Indic. 2021, 121, 107048. [Google Scholar] [CrossRef]
- Adamovich, B.V.; Medvinsky, A.B. Human activity as a factor causing the biomass, structure and seasonal dynamics of phytoplankton in adjacent fishponds. Rend. Lincei.-Sci. Fis. 2023, 34, 605–633. [Google Scholar] [CrossRef]
- Leibowitz, S.G.; Hill, R.A.; Creed, I.F.; Compton, J.E.; Golden, H.E.; Weber, M.H.; Rains, M.C.; Jones, C.E.; Lee, E.H.; Christensen, J.R.; et al. National hydrologic connectivity classification links wetlands with stream water quality. Nat. Water 2023, 1, 370–380. [Google Scholar] [CrossRef] [PubMed]
- Nagata, Y.; Ishiyama, N.; Nakamura, F.; Shibata, H.; Fukuzawa, K.; Morimoto, J. Contribution of Hydrological Connectivity in Maintaining Aquatic Plant Communities in Remnant Floodplain Ponds in Agricultural Landscapes. Wetlands 2023, 43, 38. [Google Scholar] [CrossRef]
- Esther, L.; Joshua, M.E.; Matthew, J.C. Patterns of Wetland Hydrologic Connectivity Across Coastal-Plain Wetlandscapes. Water Resour. 2023, 59, e2023WR034553. [Google Scholar] [CrossRef]
- Pénélope, B.D.; Guillaume, G.; Michael, P.; Joseph, C.; Milla, R. Integrating hydrological connectivity and zooplankton composition in Arctic ponds and lakes. Freshw. Biol. 2023, 68, 2131–2150. [Google Scholar]
- Ma, J.S.; Chen, W.Q.; Chen, M.J.; Zhong, K.; Yao, N.; Zhang, X.M.; Zhang, H.; Jeppesen, E.; Zhou, Q. Water level fluctuations associated with hydrological connectivity consolidate the food web stability of the largest Chinese freshwater lake via mediating trophodynamics and trophic structure. Ecol. Indic. 2023, 153, 110372. [Google Scholar] [CrossRef]
- Stepchinski, L.M.; Rains, M.C.; Lee, L.C.; Lis, R.A.; Nutter, W.L.; Rains, K.C.; Stewart, S.R. Hydrologic Connectivity and Flow Generation from California Vernal Pool, Swale, and Headwater Stream Complexes to Downstream Waters. Wetlands 2023, 43, 34. [Google Scholar] [CrossRef]
- Hu, H.J.; Wei, Y.X. The Freshwater Algae of China, 1st ed.; Science Press: Beijing, China, 2006; pp. 1–1023. [Google Scholar]
- Zhou, F.X.; Chen, J.H. Freshwater Micro-Organisms Atlas, 1st ed.; Chemical Industry Press: Beijing, China, 2005; pp. 1–367. [Google Scholar]
- Reynolds, C.S. Phytoplankton assemblages and their periodicity in stratifying lake systems. Ecography 1980, 3, 141–159. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Merritt, R.W.; Cummins, K.W. An introduction to the aquatic insects of North America. Freshwater Sci. 1996, 15, 593–595. [Google Scholar]
- Li, X.Z.; Liu, L.S.; Li, B.Q. Research and Practice of Marine Macroinvertebrate in China, 1st ed.; Maritime Press: Beijing, China, 2010; pp. 55–57. [Google Scholar]
- Reynolds, C.S.; Huszar, V.; Kruk, C.; Naselli, F.L.; Melo, S. Towards a functional classification of the freshwater phytoplankton. J. Plankton Res. 2002, 24, 417–428. [Google Scholar] [CrossRef]
- Reynolds, C.S. Phytoplankton periodicity: The interactions of form, function and environmental variability. Freshwater Biol. 1984, 14, 111–142. [Google Scholar] [CrossRef]
- Reynolds, C.S. Vegetation Processes in the Pelagic: A Model for Ecosystem Theory; Kinne, O., Ed.; Excellence in Ecology; Ecology Institute: Oldendorf/Luhe, Germany, 1997; pp. 70–71. [Google Scholar]
- Padisak, J.; Crossetti, L.O.; Naselli, F.L. Use and misuse in the application of the phytoplankton functional classification: A critical review with updates. Hydrobiologia 2009, 621, 1–19. [Google Scholar] [CrossRef]
- Bazhenova, O.P.; Mikhailov, V.V. Phytoplankton as an indicator of the modern ecological state of the Novosibirsk Reservoir. Inland Water Biol. 2021, 14, 670–678. [Google Scholar] [CrossRef]
- Huang, G.J.; Wang, X.Q.; Chen, Y.S.; Deng, L.; Xu, D.J. Survival strategies of phytoplankton functional groups to environmental factors in a drinking water reservoir, central China. Ann. Limnol.-Iny. J. Lim. 2021, 57, 18. [Google Scholar] [CrossRef]
- Wang, Y.B.; Liu, P.; Wu, C.; Li, X.; An, R.H.; Xie, K. Reservoir ecological operation by quantifying outflow disturbance to aquatic community dynamics. Eviron. Res. Lett. 2021, 16, 074005. [Google Scholar] [CrossRef]
- Wang, B.L.; Yang, X.Y.; Li, S.L.; Liang, X.; Li, X.D.; Wang, F.S.; Yang, M.L.; Liu, C.Q. Anthropogenic regulation governs nutrient cycling and biological succession in hydropower reservoirs. Sci. Total Eviron. 2022, 834, 155392. [Google Scholar] [CrossRef]
- Datsenko, Y.S.; Puklakov, V.V. Forecasting Phytoplankton Development in a Designed Low-Head Reservoir on the Don River. Water Resour. 2020, 47, 103–112. [Google Scholar] [CrossRef]
- Abdela, K.A.; Fantabil, A.; Muleta, D.; Yohannes, T.; Jonah, K. Bibliographic review on drought and water level articles. Discover Water 2023, 3, 17. [Google Scholar] [CrossRef]
- Trindade, R.M.L.; dos Santos, S.M.; de Souza, C.A.; dos Santos, C.R.A.; Bortolini, J.C. Using morphofunctional characteristics as a model of phytoplankton dynamics in a tropical reservoir. Braz. J. Bot. 2021, 44, 467–477. [Google Scholar] [CrossRef]
- Almeida, R.; Formiga, N.E.; Sousa, P.I.; Antunes, S.C. Contribution of zooplankton as a biological element in the assessment of reservoir water quality. Limnetical 2020, 39, 245–261. [Google Scholar] [CrossRef]
- Teixeira, A.P.; Bortolini, J.C.; Carneiro, F.M. Taxonomic and functional spatial distribution model of phytoplankton in tropical cascading reservoirs. Braz. J. Bot. 2022, 45, 791–805. [Google Scholar] [CrossRef]
- Mohanty, T.R.; Tiwari, N.K.; Das, B.K.; Swain, H.S.; Jhonson, C.; Banerjee, T. Riverine connectivity influences the phytoplankton ecology in the open floodplain wetland of the lower river Ganga. Environ. Monit. Assess. 2023, 195, 1403. [Google Scholar] [CrossRef] [PubMed]
- Abobi, S.M.; Kluger, L.C.; Wolff, M. Comparative assessment of food web structure and fisheries productivity of three reservoirs in Ghana. Fisheries Manag. Ecol. 2021, 28, 573–591. [Google Scholar] [CrossRef]
- Cazzanelli, M.; Castillo, M.M.; Soria, B.M.; Ochoa, G.S.; Sepúlveda, L.A.; Patiño, E.S.G.; Jiménez, P.N.C.; Rodiles, H.R. Epiphytes support aquatic consumers of a large floodplain lake ecosystem in the southern Gulf of Mexico. Aquat. Sci. 2023, 85, 114. [Google Scholar] [CrossRef]
- Han, L.B.; Li, Q.H.; Chen, W.S.; Wang, X.; Zhou, S.H.; Han, M.S.; Brancelj, A. The key environmental factors driving the succession of phytoplankton functional groups in Hongfeng Reservoir, southwest China. J. Oceanol. Limnol. 2022, 40, 1472–1484. [Google Scholar] [CrossRef]
- Sharip, Z. Changes in phytoplankton and zooplankton abundance and diversity in macrophyte-dominated and open pelagic ecosystem of shallow reservoirs. Inland Water Biol. 2021, 14, 427–437. [Google Scholar] [CrossRef]
- Meng, Y.; Wu, H.T.; Guan, Q.; Lu, K.L. The ecological response of Odonata larvae to hydrologic connection blocking in riverside wetlands of the Wusuli River. Chinese J. Ecol. 2021, 40, 453–459. [Google Scholar]
- Li, R.H.; Wu, Z.Q.; Huang, X.S.; Li, L.; Huang, L.; Liu, W.; Li, Y.P.; Wu, J.Q. Correlation analysis of water quality and aquatic communities in Dawangtan Reservoir, China. J. Coastal Res. 2020, 104, 575–583. [Google Scholar] [CrossRef]
- Sarma, V.V.S.S.; Krishna, M.S.; Srinivas, T.N.R. Long-term changes in Nutrient Concentration and Fluxes from the Godavari Estuary: Role of River Discharge and Fertilizer Inputs. Estuar. Coast. 2023, 46, 959–973. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phylum | Species | NW | RW | FW | FG |
|---|---|---|---|---|---|
| Bacillariophyta | Amphora ovalis | 1 | 2 | L0 | |
| Asterionella formosa | 2 | 1 | C | ||
| Ceratoneis arcus | 1 | 1 | MP | ||
| Cocconeis placentula | 1 | 1 | MP | ||
| Cyclotella meneghiniana | 2 | 3 | 2 | C | |
| Cymatopleura solea | 1 | MP | |||
| Cymbella ventricosa | 1 | 2 | 2 | MP | |
| Diatoma vulgare | 1 | 1 | 2 | MP | |
| Eunotia ehrenberg | 1 | MP | |||
| Fragilaria brevistriata | 2 | 3 | 3 | P | |
| Fragilaria ca pucina | 1 | 1 | 2 | P | |
| Fragilaria virescens | 1 | 1 | 1 | P | |
| Gyrosigma acuminatum | 1 | L0 | |||
| Gomphonema constrictum | 1 | 1 | MP | ||
| Gomphonema constrictum var.capitatum | 2 | 1 | 1 | MP | |
| Melosira granulata var. angustissima | 1 | 1 | 1 | P | |
| Melosira granulata var. angustissima f. spiralis | 1 | P | |||
| Melosira varians | 1 | 1 | 2 | TB | |
| Meridion circulare | 2 | 1 | 2 | MP | |
| Navicula anglica | 1 | 2 | 2 | MP | |
| Navicula dicephala | 1 | MP | |||
| Navicula exigua | 2 | 2 | 2 | MP | |
| Navicula radiosa | 2 | 1 | 2 | MP | |
| Pinnularia major | 1 | 1 | MP | ||
| Surirella angustata | 1 | MP | |||
| Synedra acus | 2 | 2 | 2 | D | |
| Synedra ulna | 1 | 1 | D | ||
| Synedra tabulata | 1 | 1 | 1 | D | |
| Chlorophyta | Actinastrum fluviatile | J | |||
| Ankistrodesmus acicularis | 1 | X1 | |||
| Ankistrodesmus angustus | 2 | 1 | X1 | ||
| Chlamydomonas globosa | 1 | 2 | 1 | X2 | |
| Chlamydomonas ovalis | 1 | 3 | 1 | X2 | |
| Chodatella quadriseta | 1 | 1 | J | ||
| Cosmarium obtusatum | 1 | N | |||
| Crucigenia apiculata | J | ||||
| Crucigenia tetrapedia | J | ||||
| Dictyosphaerium pulchellum | 2 | 1 | F | ||
| Kirchneriella lunaris | 1 | F | |||
| Oocystis elliptica | 1 | 1 | F | ||
| Pandorina morum | 2 | G | |||
| Scenedesmus platydiscus | 2 | J | |||
| Scenedesmus quadricauda | 1 | 2 | J | ||
| Selenastrum gracile | 1 | 1 | F | ||
| Stichococcus bacillaris | 1 | 1 | F | ||
| Tetrastrum elegans | 1 | J | |||
| Tetraëdron pusillum | 1 | J | |||
| Tetraëdron trilobulatum | 1 | J | |||
| Treubaria crassispina | 1 | 1 | F | ||
| Ulothris variabilis | MP | ||||
| Westella botryoides | 1 | F | |||
| Westellopsis linearis | 1 | F | |||
| Chrysophyta | Chromulina elegans | 2 | 3 | 2 | X3 |
| Chromulina globosa | 1 | 1 | X3 | ||
| Dinobryon divergens | 1 | E | |||
| Kephyrion planctonicum | 1 | 1 | X3 | ||
| Mallomonas Perty | 1 | Ws | |||
| Synura Ehrenberg | 1 | Ws | |||
| Cryptophyta | Chroomonas acuta | 1 | X2 | ||
| Cryptomonas ovata | 1 | 2 | 1 | Y | |
| Cyanophyta | Aphanizomenon flos-aquae | 1 | H1 | ||
| Chroococcus minutus | 1 | L0 | |||
| Merismopedia marssonii | 2 | 2 | L0 | ||
| Merismopedia minima | 4 | 4 | 4 | L0 | |
| Phormidium allorgei | 2 | 1 | S1 | ||
| Phormidium lismorense | 2 | S1 | |||
| Euglenophyta | Euglena oxyuris | 1 | 1 | W1 | |
| Pyrrophyta | Glenodinium pulvisculus | 1 | Y |
| Phylum | Species | Frequency | Proportion | Dominance |
|---|---|---|---|---|
| Bacillariophyta | Cyclotella meneghiniana Kiits | 1.0000 | 0.1051 | 0.1051 |
| Cymbella ventricosa Kutz. | 0.8333 | 0.0283 | 0.0236 | |
| Fragilaria brevistriata Grun | 1.0000 | 0.1459 | 0.1459 | |
| Navicula exigua Greg | 1.0000 | 0.0912 | 0.0912 | |
| Synedra acus Kutz. | 0.8333 | 0.0422 | 0.0352 | |
| Chlorophyta | Chlamydomonas ovalis Zietz | 0.6667 | 0.2136 | 0.1424 |
| Chrysophyta | Chromulina elegans Doflein | 1.0000 | 0.0576 | 0.0576 |
| Cryptophyta | Cryptomonas ovata Ehrenberg | 0.5000 | 0.0432 | 0.0216 |
| Cyanophyta | Merismopedia minima G.Beck | 0.9444 | 0.0907 | 0.0857 |
| Group | Indicator Species | Functional Group | Indicator Value | p |
|---|---|---|---|---|
| Reservoir Wetlands Group | Chlamydomonas ovalis Zietz | X2 | 99.0 | 0.0002 |
| Synedra acus Kutz. | D | 70.2 | 0.0002 | |
| Chlamydomonas globosa Snow. | X2 | 84.5 | 0.0004 | |
| Ankistrodesmus angustus Bernard | X1 | 80.5 | 0.0004 | |
| Cyclotella meneghiniana Ktz. | C | 67.7 | 0.0034 | |
| Chromulina elegans Doflein | X3 | 65.0 | 0.0044 | |
| Kirchneriella lunaris Krich. | F | 62.5 | 0.0102 | |
| Asterionella formosa Hassall | C | 60.5 | 0.0268 | |
| Flow-reduced Wetlands Group | Diatoma vulgare Bory | MP | 84.6 | 0.0030 |
| Fragilaria capucina Desm. | P | 68.9 | 0.0124 | |
| Meridion circulare Ag. | MP | 73.1 | 0.0224 | |
| Natural Wetlands Group | Ceratoneis arcus Ktz. | MP | 89.9 | 0.0018 |
| Treubaria crassispina Smith | F | 61.0 | 0.0134 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meng, Y.; Wu, H. Responses of Phytoplankton Communities to Flow Regulation in Northeastern Riverine Wetlands of China. Diversity 2023, 15, 1191. https://doi.org/10.3390/d15121191
Meng Y, Wu H. Responses of Phytoplankton Communities to Flow Regulation in Northeastern Riverine Wetlands of China. Diversity. 2023; 15(12):1191. https://doi.org/10.3390/d15121191
Chicago/Turabian StyleMeng, Yao, and Haitao Wu. 2023. "Responses of Phytoplankton Communities to Flow Regulation in Northeastern Riverine Wetlands of China" Diversity 15, no. 12: 1191. https://doi.org/10.3390/d15121191
APA StyleMeng, Y., & Wu, H. (2023). Responses of Phytoplankton Communities to Flow Regulation in Northeastern Riverine Wetlands of China. Diversity, 15(12), 1191. https://doi.org/10.3390/d15121191