

Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Sample Survey, Collection, and Fungal Isolation
2.2. DNA Extraction, PCR Amplification, and Sequencing
2.3. Phylogenetic Analyses
2.4. Pathogenicity Test
2.5. Determination of Colony Morphologies and Growth Rates
2.6. Analysis of Culture Characteristics
2.7. Statistical Analysis
3. Results
3.1. Incidence and Fungal Isolation
3.2. Phylogenetic Analyses
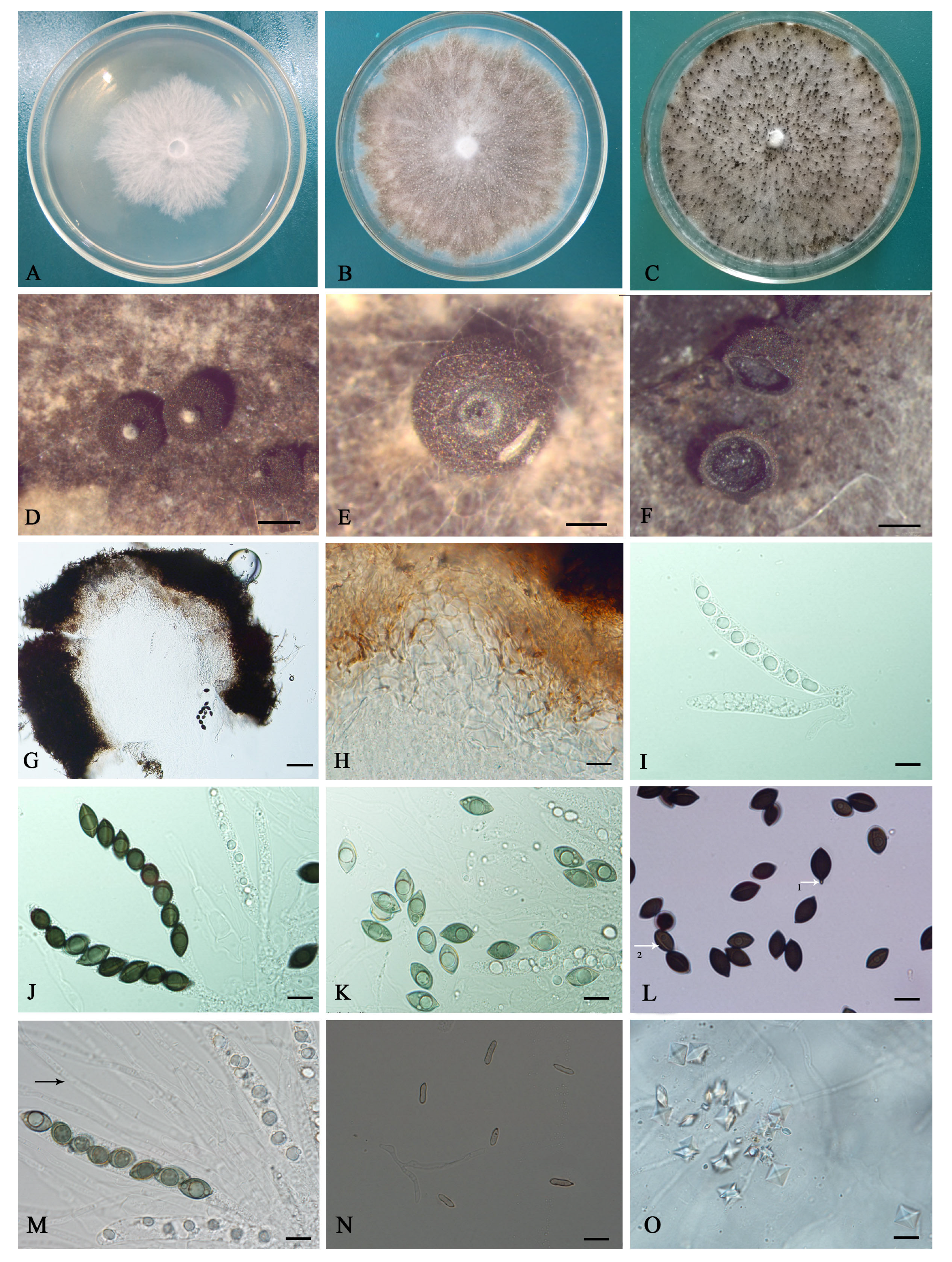
3.3. Description of Coniolariella gamsii
3.4. Pathogenicity Test
3.5. Cultural Characteristics of the Pathogen
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wiehle, M.; Vornam, B.; Wesche, K.; Goenster, S.; Buerkert, A. Population structure and genetic diversity of Populus laurifolia in fragmented riparian gallery forests of the Mongolian Altai Mountains. Flora 2016, 224, 112–122. [Google Scholar] [CrossRef]
- Zheng, S.; Zhang, J.G.; He, C.Y.; Bao, J.; Duan, A.G.; Zeng, Y.F.; Sai, L. Genetic diversity of Populus laurifolia and Populus nigra along Erqis River. For. Res. 2014, 27, 295–301. [Google Scholar]
- Checa, J.; Arenal, F.; Blanco, N.; Rogers, J.D. Coniolariella hispanica sp. nov. and other additions to Coniolariella. Mycol. Res. 2008, 112, 795–801. [Google Scholar] [CrossRef] [PubMed]
- Zare, R.; Asgari, B.; Gams, W. The species of Coniolariella. Mycologia 2010, 102, 1383–1388. [Google Scholar] [CrossRef] [PubMed]
- García, D.; Stchigel, A.M.; Cano, J.; Calduch, M.; Hawksworth, D.L.; Guarro, J. Molecular phylogeny of Coniochaetales. Mycol. Res. 2006, 110, 1271–1289. [Google Scholar] [CrossRef] [PubMed]
- Zhuang, J.Y. Rust fungi from the desert of northern Xinjiang. Acta Mycol. Sin. 1989, 8, 259–269. [Google Scholar]
- Zhuang, W.Y. Fungi of Northwestern China; Mycotaxon, Ltd.: Ithaca, NY, USA, 2005; p. 430. [Google Scholar]
- Jiang, N.; Voglmayr, H.; Xue, H.; Piao, C.G.; Li, Y. Morphology and Phylogeny of Pestalotiopsis (Sporocadaceae, Amphisphaeriales) from Fagaceae Leaves in China. Microbiol Spectr. 2022, 10, e03272-22. [Google Scholar] [CrossRef]
- Zhang, Q.T.; He, M. Canker on bark of Populus spp. caused by Cytospora tritici, a new disease in China. Plant Dis. 2012, 96, 1578. [Google Scholar] [CrossRef]
- Huang, L.; Zhu, Y.N.; Yang, J.Y.; Li, D.W.; Li, Y.; Bian, L.M.; Ye, J.R. Shoot blight on Chinese fir (Cunninghamia lanceolata) is caused by Bipolaris oryzae. Plant Dis. 2017, 102, 500–506. [Google Scholar] [CrossRef]
- Solarte, F.A.; Muñoz, C.G.; Maharachchikumbura, S.; Álvarez, E. Diversity of Neopestalotiopsis and Pestalotiopsis spp., causal agents of guava scab in Colombia. Plant Dis. 2017, 102, 49–59. [Google Scholar] [CrossRef]
- Ma, R.; Ye, S.H.; Zhao, Y.; Michailides, T.J.; Tian, C.M. New leaf and fruit disease of Juglans regia caused by Juglanconis juglandina in Xinjiang, China. For. Pathol. 2019, 49, e12537. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. Guide Methods Appl. 1990, 18, 315–322. [Google Scholar]
- O'Donnell, K.; Cigelnik, E. Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus fusariumare nonorthologous. Molecul Phylogenet Evol. 1997, 7, 103–116. [Google Scholar] [CrossRef]
- Jiang, N.; Voglmayr, H.; Ma, C.Y.; Xue, H.; Piao, C.G.; Li, Y. A new Arthrinium-like genus of Amphisphaeriales in China. MycoKeys 2022, 92, 27–43. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Toh, H. Parallelization of the MAFFT multiple sequence alignment program. Bioinformatics 2010, 26, 1899–1900. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Pfeiffer, W.; Schwartz, T. Creating the CIPRES Science Gateway for Inference of Large Phylogenetic Trees; Institute of Electrical and Electronics Engineers: New Orleans, LA, USA, 2010. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef]
- Jia, X.H.; Wang, W.; Fu, J.F.; Du, Y.M.; Wang, Y.; Zhou, R.J.; Liu, B.L. Biological characteristics and host range of Athelia bombacina causing postharvest fruit rot on pear. Acta Hortic. Sin. 2020, 47, 1253–1263. [Google Scholar]
- Asgari, B.; Zare, R. Two new Coniochaeta species from Iran. Nova Hedwig. 2006, 82, 227–236. [Google Scholar] [CrossRef]
- Davis, E.C.; Franklin, J.B.; Shaw, A.J.; Vilgalys, R. Endophytic Xylaria (Xylariaceae) among liverworts and angiosperms: Phylogenetics, distribution, and symbiosis. Am J Bot. 2003, 90, 1661–1667. [Google Scholar] [CrossRef] [PubMed]
- Rogers, J.D. The Xylariaceae: Systematic, biological and evolutionary aspects. Mycologia 1979, 71, 1–42. [Google Scholar] [CrossRef]
- González, F.E.S.M. A Mycofloristic and Cultural Study of the Xylariaceae of México; Washington State University: Washington, DC, USA, 1992. [Google Scholar]
- Garcia-Aroca, T.; Price, P.P.; Tomaso-Peterson, M.; Allen, T.W.; Wilkerson, T.H.; Spurlock, T.N.; Faske, T.R.; Bluhm, B.; Conner, K.; Sikora, E.; et al. Xylaria necrophora sp. nov. is an emerging root-associated pathogen responsible for taproot decline of soybean in the southern United States. Mycologia 2021, 113, 326–347. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.L.; Jonglaekha, N.; Kshirsagar, A.; Maitland, D.J.; Mekkamol, S.; Nugent, L.K.; Phosri, C.; Rodtong, S.; Ruchichachorn, N.; Sangvichien, E.; et al. The Xylariaceae as phytopathogens. Recent Res. Dev. Plant Sci. 2003, 1, 1–19. [Google Scholar]
- Shen, J.W.; Zhou, S.J.; Yu, H.Y.; Zhang, B.; Zhou, H. Studies on the biological characteristics of Xylaria pedunculata. J. Henan Agric. Sci. 2008, 12, 75–77. [Google Scholar]
- Li, P. Studies on the Physiological Mechanism of Populus bolleana in Response to the Fungal Canker Diseases; Chinese Academy of Forestry Sciences: Beijing, China, 2019. [Google Scholar]
- Jiang, H.; Zhou, F.; Xia, S.C.; Che, J.H.; Wang, G.F. Providencia rettgeri as the causal agent of the brown slime flux of Populus tomentosa. For. Pathol. 2020, 50, e12576. [Google Scholar]
- Alexopoulos, C.J.; Mims, C.W.; Blackwell, M. Introductory Mycology, 4th ed.; Wiley & Sons: New York, NY, USA, 1996. [Google Scholar]
- Balasuriya, A.; Adikaram, N.K.B. Some spatial, temporal and spatio-temporal considerations of wood decay of tea (Camellia sinensis), caused by Nemania diffusa (syn. Hypoxylon vestitum). Crop Prot. 2009, 28, 273–279. [Google Scholar] [CrossRef]
- Pérez-Jiménez, R.M. A review of the biology and pathogenicity of Rosellinia necatrix—The cause of white root rot disease of fruit trees and other plants. J. Phytopathol. 2006, 154, 257–266. [Google Scholar] [CrossRef]
- Eguchi, N.; Kondo, K.I.; Yamagishi, N. Bait twig method for soil detection of Rosellinia necatrix, causal agent of white root rot of Japanese pear and apple, at an early stage of tree infection. J. Gen. Plant Pathol. 2009, 75, 325–330. [Google Scholar] [CrossRef]
- Ushiyam, K. Mass production of conidiophores of Phomopsis spp., Colletotrichum and Pestalotiopsis spp. by agar leaf disk method. Proceeding Kanto-Tosan Plant Prot. Soc. 1995, 42, 101–103. [Google Scholar]
- Liang, D.D.; Zhang, Y.L.; Gu, Z.M. Study on sporulation conditions of Phoma herbarum SYAU-06 Strain. Agrochemicals 2020, 59, 219–222. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species of Coniolariella | Isolate | Host | Country | Sequences | |
|---|---|---|---|---|---|
| ITS | LSU | ||||
| C. ershadii | CBS 119785 | Pistacia vera | Iran | GU553328 | GU553331 |
| C. gamsii | CBS 114379 | Hordeum vulgare | Iran | GU553325 | GU553329 |
| C. gamsii | CFCC 55706 | Populus laurifolia | China | MZ423210 | MZ436978 |
| C. gamsii | XJAU KY-2-1 | Populus laurifolia | China | MZ423211 | MZ436979 |
| C. hispanica | CBS 124506 | Eryngium campestre | Spain | MH863381 | MH874902 |
| C. limoniispora | 108fa | Vitis vinifera | USA | NA | EF489469 |
| C. limoniispora | AH24323 | Nicotiana glauca | Spain | AY908997 | EF489464 |
| C. macrothecia | CBS 125772 | Soil | USA | MH863751 | MH875219 |
| Xylaria badia | BCC 1190 | NA | Thailand | AB376711 | NA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, C.; Lu, H.; Chen, H.; Chen, S.; Wang, C.; Zhou, Z.; Ma, R. Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China. Diversity 2023, 15, 1190. https://doi.org/10.3390/d15121190
Liu C, Lu H, Chen H, Chen S, Wang C, Zhou Z, Ma R. Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China. Diversity. 2023; 15(12):1190. https://doi.org/10.3390/d15121190
Chicago/Turabian StyleLiu, Chuli, Hailong Lu, Hongjin Chen, Shuikang Chen, Caixia Wang, Zhiyong Zhou, and Rong Ma. 2023. "Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China" Diversity 15, no. 12: 1190. https://doi.org/10.3390/d15121190
APA StyleLiu, C., Lu, H., Chen, H., Chen, S., Wang, C., Zhou, Z., & Ma, R. (2023). Coniolariella gamsii Causes Poplar Leaf Spot Disease in Xinjiang, China. Diversity, 15(12), 1190. https://doi.org/10.3390/d15121190