
The Importance of Including Spatial Autocorrelation When Modelling Species Richness in Archipelagos: A Bayesian Approach



 and
and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Statistical Model
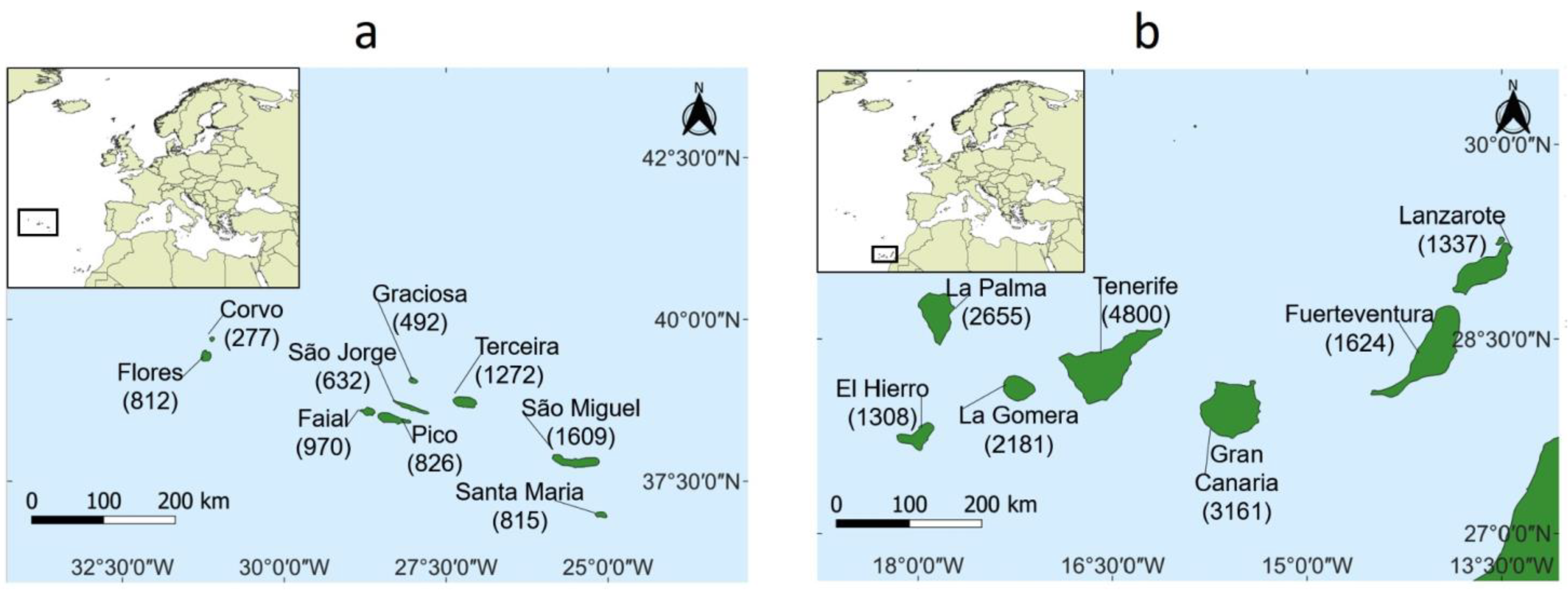
2.2. Case Study—The Azores and Canary Archipelagos
2.2.1. Study Area
2.2.2. Arthropod Data
2.3. Selection of Explanatory Variables
2.4. Choice of Priors
2.5. Software Packages
3. Results
3.1. The Traditional ISAR with and without the GAUSSIAN Process
3.2. Considering All Explanatory Variables
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Matthews, T.J.; Triantis, K.A.; Whittaker, R.J. The Species–Area Relationship: Theory and Application; Cambridge University Press: Cambridge, UK, 2021. [Google Scholar]
- Whittaker, R.J.; Fernández-Palacios, J.M. Island Biogeography: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2007. [Google Scholar]
- Warren, B.H.; Simberloff, D.; Ricklefs, R.E.; Aguilée, R.; Condamine, F.L.; Gravel, D.; Morlon, H.; Mouquet, N.; Rosindell, J.; Casquet, J.; et al. Islands as Model Systems in Ecology and Evolution: Prospects Fifty Years after MacArthur-Wilson. Ecol. Lett. 2015, 18, 200–217. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.M.; Field, R.; Ricklefs, R.E. New Directions in Island Biogeography. Glob. Ecol. Biogeogr. 2016, 25, 751–768. [Google Scholar] [CrossRef]
- Borregaard, M.K.; Amorim, I.R.; Borges, P.A.; Cabral, J.S.; Fernández-Palacios, J.M.; Field, R.; Heaney, L.R.; Kreft, H.; Matthews, T.J.; Olesen, J.M.; et al. Oceanic Island Biogeography through the Lens of the General Dynamic Model: Assessment and Prospect. Biol. Rev. 2016, 92, 830–853. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, R.H.; Wilson, E.O. The Theory of Island Biogeography: Monographs in Population Biology; Princeton University Press: Princeton, NJ, USA, 1967. [Google Scholar]
- MacArthur, R.H.; Wilson, E.O. Equilibrium-Theory of Insular Zoogeography. Evolution 1963, 17, 373. [Google Scholar] [CrossRef]
- Whittaker, R.J.; Triantis, K.A.; Ladle, R.J. A General Dynamic Theory of Oceanic Island Biogeography. J. Biogeogr. 2008, 35, 977–994. [Google Scholar] [CrossRef]
- Witt, C.C.; Maliakal-Witt, S. Why Are Diversity and Endemism Linked on Islands? Ecography 2007, 30, 331–333. [Google Scholar] [CrossRef]
- Emerson, B.C.; Kolm, N. Species Diversity Can Drive Speciation. Nature 2005, 434, 1015–1017. [Google Scholar] [CrossRef] [PubMed]
- Irl, S.D.; Harter, D.E.; Steinbauer, M.J.; Gallego Puyol, D.; Fernández-Palacios, J.M.; Jentsch, A.; Beierkuhnlein, C. Climate vs. Topography–Spatial Patterns of Plant Species Diversity and Endemism on a High-Elevation Island. J. Ecol. 2015, 103, 1621–1633. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Otto, R.; Naranjo-Cigala, A.; Beierkuhnlein, C.; Fernández-Palacios, J.-M. Increase of Island Endemism with Altitude–Speciation Processes on Oceanic Islands. Ecography 2012, 35, 23–32. [Google Scholar] [CrossRef] [Green Version]
- Triantis, K.A.; Hortal, J.; Amorim, I.; Cardoso, P.; Santos, A.M.; Gabriel, R.; Borges, P.A. Resolving the Azorean Knot: A Response to Carine & Schaefer (2010). J. Biogeogr. 2012, 39, 1179–1184. [Google Scholar]
- May, R.M. Patterns of Species Abundance and Diversity. In Ecology and Evolution of Communities; Cody, M.L., Diamond, J.M., Eds.; Harvard University Press: Cambridge, MA, USA, 1975; pp. 81–120. [Google Scholar]
- Diamond, J.M. Distributional Ecology of New Guinea Birds. Science 1973, 179, 759–769. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Cardoso, P.; Rigal, F.; Triantis, K.A.; Borges, P.A. Modeling Directional Spatio-Temporal Processes in Island Biogeography. Ecol. Evol. 2015, 5, 4671–4682. [Google Scholar] [CrossRef] [PubMed]
- Kreft, H.; Jetz, W.; Mutke, J.; Kier, G.; Barthlott, W. Global Diversity of Island Floras from a Macroecological Perspective. Ecol. Lett. 2008, 11, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Selmi, S.; Boulinier, T. Ecological Biogeography of Southern Ocean Islands: The Importance of Considering Spatial Issues. Am. Nat. 2001, 158, 426–437. [Google Scholar] [CrossRef]
- Keitt, T.H.; Bjørnstad, O.N.; Dixon, P.M.; Citron-Pousty, S. Accounting for Spatial Pattern When Modeling Organism-Environment Interactions. Ecography 2002, 25, 616–625. [Google Scholar] [CrossRef] [Green Version]
- Dormann, C.F.; McPherson, J.M.; Araújo, M.B.; Bivand, R.; Bolliger, J.; Carl, G.; Davies, R.G.; Hirzel, A.; Jetz, W.; Daniel Kissling, W.; et al. Methods to Account for Spatial Autocorrelation in the Analysis of Species Distributional Data: A Review. Ecography 2007, 30, 609–628. [Google Scholar] [CrossRef] [Green Version]
- McElreath, R. Statistical Rethinking. In Texts in Statistical Science; CRC Press: Boca Raton, FL, USA, 2020. [Google Scholar]
- Gelman, A.; Carlin, J.B.; Stern, H.S.; Dunson, D.B.; Vehtari, A.; Rubin, D.B. Bayesian Data Analysis, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2014. [Google Scholar]
- Golding, N.; Purse, B.V. Fast and Flexible Bayesian Species Distribution Modelling Using Gaussian Processes. Methods Ecol. Evol. 2016, 7, 598–608. [Google Scholar] [CrossRef] [Green Version]
- Hadjipantelis, P.Z.; Jones, N.S.; Moriarty, J.; Springate, D.A.; Knight, C.G. Function-Valued Traits in Evolution. J. R. Soc. Interface 2013, 10, 20121032. [Google Scholar] [CrossRef] [Green Version]
- Talluto, M.V.; Mokany, K.; Pollock, L.J.; Thuiller, W. Multifaceted Biodiversity Modelling at Macroecological Scales Using Gaussian Processes. Divers. Distrib. 2018, 24, 1492–1502. [Google Scholar] [CrossRef] [Green Version]
- Borges, P.A.V.; Vieira, V.; Amorim, I.R.; Bicudo, N.; Fritzén, N.; Gaspar, C.; Heleno, R.; Hortal, J.; Lissner, J.; Logunov, D.; et al. List of arthropods (Arthropoda). In A List of the Terrestrial and Marine Biota from the Azores; Borges, P.A.V., Costa, A., Cunha, R., Gabriel, R., Gonçalves, V., Martins, A.F., Melo, I., Parente, M., Raposeiro, P., Rodrigues, P., Eds.; Princípia: Cascais, Portugal, 2010; pp. 179–246. [Google Scholar]
- Arechavaleta, M.; Rodríguez, S.; Zurita, N.; García, A. Lista de Especies Silvestres de Canarias. Hongos, Plantas y Animales Terrestres. 2009. Gobierno de Canararias 2010. Available online: https://mc-stan.org (accessed on 10 December 2021).
- Dias, F.S.; Betancourt, M.; Rodríguez-González, P.M.; Borda-de-Água, L. BetaBayes—A Bayesian Approach for Comparing Ecological Communities. Diversity 2022, 15, 858. [Google Scholar] [CrossRef]
- Dias, F.S.; Betancourt, M.; Rodríguez-González, P.M.; Borda-de-Água, L. Analysing the Distance Decay of Community Similarity in River Networks Using Bayesian Methods. Sci. Rep. 2021, 11, 21660. [Google Scholar] [CrossRef] [PubMed]
- Rosenzweig, M.L. Species Diversity in Space and Time, 1st ed.; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar]
- Gelman, A.; Hill, J. Data Analysis Using Regression and Multilevel/Hierarchical Models; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Edge, M.D. Statistical Thinking From Scratch: A Primer For Scientists; Oxford University Press: Oxford, UK, 2019; ISBN 0-19-882762-8. [Google Scholar]
- Rasmussen, C.E.; Williams, C.K.I. Gaussian Processes for Machine Learning; MIT Press: Cambridge, MA, USA, 2006. [Google Scholar]
- Borges, P.A.V.; Lobo, J.M.; de Azevedo, E.B.; Gaspar, C.S.; Melo, C.; Nunes, L.V. Invasibility and Species Richness of Island Endemic Arthropods: A General Model of Endemic vs. Exotic Species. J. Biogeogr. 2006, 33, 169–187. [Google Scholar] [CrossRef] [Green Version]
- Fernandopullé, D. Climatic Characteristics of the Canary Islands. In Biogeography and ecology in the Canary Islands; Springer: Berlin/Heidelberg, Germany, 1976; pp. 185–206. [Google Scholar]
- Ramalho, R.S.; Helffrich, G.; Madeira, J.; Cosca, M.; Thomas, C.; Quartau, R.; Hipólito, A.; Rovere, A.; Hearty, P.J.; Ávila, S.P. Emergence and Evolution of Santa Maria Island (Azores)—The Conundrum of Uplifted Islands Revisited. Bulletin 2017, 129, 372–390. [Google Scholar] [CrossRef] [Green Version]
- Azevedo, J.; Ferreira, M.P. The Volcanotectonic Evolution of Flores Island, Azores (Portugal). J. Volcanol. Geotherm. Res. 2006, 156, 90–102. [Google Scholar] [CrossRef] [Green Version]
- Prægel, N.-O.; Holm, P.M. Lithospheric Contributions to High-MgO Basanites from the Cumbre Vieja Volcano, La Palma, Canary Islands and Evidence for Temporal Variation in Plume Influence. J. Volcanol. Geotherm. Res. 2006, 149, 213–239. [Google Scholar] [CrossRef]
- van den Bogaard, P. The Origin of the Canary Island Seamount Province-New Ages of Old Seamounts. Sci. Rep. 2013, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Ancochea, E.; Huertas, M.J.; Hernán, F.; Brändle, J.L.; Alonso, M. Structure, Composition and Age of the Small Islands of Santa Luzia, Branco and Raso (Cape Verde Archipelago). J. Volcanol. Geotherm. Res. 2015, 302, 257–272. [Google Scholar] [CrossRef]
- Ramalho, R.S.; Brum da Silveira, A.; Fonseca, P.E.; Madeira, J.; Cosca, M.; Cachão, M.; Fonseca, M.M.; Prada, S.N. The Emergence of Volcanic Oceanic Islands on a Slow-Moving Plate: The Example of M Adeira I Sland, NE A Tlantic. Geochem. Geophys. Geosystems 2015, 16, 522–537. [Google Scholar] [CrossRef]
- Ávila, S.; Cachão, M.; Ramalho, R.; Botelho, A.; Madeira, P.; Rebelo, A.; Cordeiro, R.; Melo, C.; Hipólito, A.; Ventura, M.; et al. The Palaeontological Heritage of Santa Maria Island (Azores: NE Atlantic): A Re-Evaluation of Geosites in GeoPark Azores and Their Use in Geotourism. Geoheritage 2016, 8, 155–171. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2015. [Google Scholar]
- Stan Development Team. Stan Modeling Language User’s Guide and Reference Manual; Version 2.17. 0 2017; Stan Development Team: Aberdeen, South Dakota, 2017. [Google Scholar]
- Burnham, K.P.; Anderson, D.R. Model Selection and Multimodel Inference. A Practical Information-Theoretic Approach Second Edition; Springer: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Cowie, R.H.; Holland, B.S. Dispersal Is Fundamental to Biogeography and the Evolution of Biodiversity on Oceanic Islands. J. Biogeogr. 2006, 33, 193–198. [Google Scholar] [CrossRef]
- Williamson, M. Relationship of Species Number to Area, Distance and Other Variables. In Analytical Biogeography. An Integrated Approach to the Study of Animal and Plant Distributions; Myers, A.A., Giller, P.S., Eds.; Chapman and Hall: Boca Raton, FL, USA, 1988; pp. 91–115. [Google Scholar]
- Sanmartín, I.; Van Der Mark, P.; Ronquist, F. Inferring Dispersal: A Bayesian Approach to Phylogeny-Based Island Biogeography, with Special Reference to the Canary Islands. J. Biogeogr. 2008, 35, 428–449. [Google Scholar] [CrossRef]
- Kühn, I. Incorporating Spatial Autocorrelation May Invert Observed Patterns. Divers. Distrib. 2007, 13, 66–69. [Google Scholar] [CrossRef]
- Borregaard, M.K.; Matthews, T.J.; Whittaker, R.J. The General Dynamic Model: Towards a Unified Theory of Island Biogeography? Glob. Ecol. Biogeogr. 2016, 25, 805–816. [Google Scholar] [CrossRef]
- Borges, P.A.V.; Hortal, J. Time, Area and Isolation: Factors Driving the Diversification of Azorean Arthropods. J. Biogeogr. 2009, 36, 178–191. [Google Scholar] [CrossRef] [Green Version]
- Steinbauer, M.J.; Field, R.; Grytnes, J.-A.; Trigas, P.; Ah-Peng, C.; Attorre, F.; Birks, H.J.B.; Borges, P.A.V.; Cardoso, P.; Chou, C.-H.; et al. Topography-Driven Isolation, Speciation and a Global Increase of Endemism with Elevation. Glob. Ecol. Biogeogr. 2016, 25, 1097–1107. [Google Scholar] [CrossRef]
- Cutts, V.; Katal, N.; Löwer, C.; Algar, A.C.; Steinbauer, M.J.; Irl, S.D.; Beierkuhnlein, C.; Field, R. The Effect of Small-Scale Topography on Patterns of Endemism within Islands. Front. Biogeogr. 2019, 11, e43737. [Google Scholar] [CrossRef]
- Pereira, H.M.; Proenca, V.M.; Vicente, L. Does Species Diversity Really Drive Speciation? Ecography 2007, 30, 328–330. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Mean | SD | CV | 2.50% | 97.50% | n_eff | Rhat | ||
|---|---|---|---|---|---|---|---|---|
| Without GP | c | 100.64 | 20.23 | 0.20 | 63.33 | 139.77 | 223 | 1.00 |
| z | 0.41 | 0.05 | 0.12 | 0.32 | 0.52 | 172 | 1.00 | |
| ϕ | 4.53 | 1.99 | 0.44 | 1.66 | 9.36 | 195 | 1.00 | |
| With GP | c | 99.53 | 19.16 | 0.19 | 64.45 | 136.28 | 219 | 1.01 |
| z | 0.42 | 0.06 | 0.14 | 0.31 | 0.56 | 123 | 1.00 | |
| ϕ | 6.47 | 3.17 | 0.49 | 2.05 | 14.54 | 232 | 1.02 | |
| η2 | 0.23 | 0.48 | 2.09 | 0.00 | 1.39 | 363 | 1.01 | |
| ρ2 | 1.56 | 1.83 | 1.17 | 0.02 | 6.22 | 406 | 1.00 |
| Mean | SD | CV | 2.50% | 97.50% | n_eff | Rhat | ||
|---|---|---|---|---|---|---|---|---|
| Without GP | c | 101.91 | 20.31 | 0.20 | 61.69 | 139.74 | 230 | 1.00 |
| z | 0.47 | 0.05 | 0.11 | 0.38 | 0.59 | 172 | 1.01 | |
| ϕ | 2.93 | 1.47 | 0.50 | 0.89 | 6.00 | 224 | 1.00 | |
| With GP | c | 99.31 | 20.4 | 0.20 | 57.9 | 136.31 | 248 | 1.00 |
| z | 0.46 | 0.10 | 0.22 | 0.18 | 0.63 | 54 | 1.00 | |
| ϕ | 5.03 | 2.64 | 0.52 | 1.30 | 11.49 | 292 | 1.00 | |
| η2 | 0.58 | 0.80 | 1.34 | 0.02 | 2.69 | 122 | 1.00 | |
| ρ2 | 1.37 | 1.96 | 1.43 | 0.02 | 7.79 | 204 | 1.00 |
| ISAR Model | WAIC | ΔWAIC | Weight | |
|---|---|---|---|---|
| Azores | With GP | 127.00 | 0.00 | 0.56 |
| Without GP | 127.50 | 0.50 | 0.44 | |
| Canary Islands | With GP | 116.70 | 0.00 | 0.76 |
| Without GP | 119.00 | 2.30 | 0.24 | |
| C | Fl | Fa | P | G | SJ | T | SM | SMa | |
| C | 1.000 | 0.831 | 0.004 | 0.001 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 |
| Fl | 0.831 | 1.000 | 0.005 | 0.001 | 0.001 | 0.000 | 0.000 | 0.000 | 0.000 |
| Fa | 0.004 | 0.005 | 1.000 | 0.810 | 0.499 | 0.654 | 0.185 | 0.000 | 0.000 |
| P | 0.001 | 0.001 | 0.810 | 1.000 | 0.567 | 0.819 | 0.346 | 0.002 | 0.000 |
| G | 0.001 | 0.001 | 0.499 | 0.567 | 1.000 | 0.740 | 0.510 | 0.002 | 0.000 |
| SJ | 0.000 | 0.000 | 0.654 | 0.819 | 0.740 | 1.000 | 0.560 | 0.004 | 0.000 |
| T | 0.000 | 0.000 | 0.185 | 0.346 | 0.510 | 0.560 | 1.000 | 0.040 | 0.001 |
| SM | 0.000 | 0.000 | 0.000 | 0.002 | 0.002 | 0.004 | 0.040 | 1.000 | 0.371 |
| SMa | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.000 | 0.001 | 0.371 | 1.000 |
| F | L | GC | T | LG | EH | LP | |
| F | 1.000 | 0.506 | 0.081 | 0.003 | 0.000 | 0.000 | 0.000 |
| L | 0.506 | 1.000 | 0.008 | 0.000 | 0.000 | 0.000 | 0.000 |
| GC | 0.081 | 0.008 | 1.000 | 0.345 | 0.078 | 0.004 | 0.005 |
| T | 0.003 | 0.000 | 0.345 | 1.000 | 0.579 | 0.097 | 0.166 |
| LG | 0.000 | 0.000 | 0.078 | 0.579 | 1.000 | 0.449 | 0.436 |
| EH | 0.000 | 0.000 | 0.004 | 0.097 | 0.449 | 1.000 | 0.317 |
| LP | 0.000 | 0.000 | 0.005 | 0.166 | 0.436 | 0.317 | 1.000 |
| Archipelago | Model | WAIC | ΔWAIC | Weight |
|---|---|---|---|---|
| Azores | fulle.poisson.gp | 91.6 | 0.0 | 0.23 |
| area.poisson.gp | 91.7 | 0.1 | 0.21 | |
| dist.poisson.gp | 91.9 | 0.3 | 0.20 | |
| diste.poisson.gp | 92.0 | 0.4 | 0.19 | |
| elev.poisson.gp | 92.2 | 0.5 | 0.17 | |
| Canary Islands | area.poisson.gp | 78.3 | 0.0 | 0.20 |
| elev.poisson.gp | 78.4 | 0.1 | 0.19 | |
| dist.poisson.gp | 78.5 | 0.3 | 0.17 | |
| fulle.poisson.gp | 78.7 | 0.4 | 0.16 | |
| full.poisson.gp | 78.9 | 0.7 | 0.14 | |
| diste.poisson.gp | 79.1 | 0.8 | 0.13 |
| Archipelago | Model | WAIC | ΔWAIC | Weight |
|---|---|---|---|---|
| Azores | elev.poisson.gp | 85.7 | 0.0 | 0.18 |
| area.poisson.gp | 85.8 | 0.1 | 0.17 | |
| fulle.poisson.gp | 85.8 | 0.1 | 0.17 | |
| full.poisson.gp | 85.8 | 0.2 | 0.16 | |
| dist.poisson.gp | 85.9 | 0.2 | 0.16 | |
| diste.poisson.gp | 85.9 | 0.2 | 0.16 | |
| Canary Islands | fulle.poisson.gp | 62.1 | 0.0 | 0.20 |
| elev.poisson.gp | 62.2 | 0.1 | 0.19 | |
| area.poisson.gp | 62.4 | 0.3 | 0.17 | |
| full.poisson.gp | 62.4 | 0.3 | 0.17 | |
| dist.poisson.gp | 62.8 | 0.8 | 0.14 | |
| diste.poisson.gp | 62.9 | 0.8 | 0.13 |
| Archipelago | Model | WAIC | ΔWAIC | Weight |
|---|---|---|---|---|
| Azores | area.poisson.gp | 70.7 | 0.0 | 0.38 |
| fulle.poisson.gp | 72.2 | 1.5 | 0.18 | |
| full.poisson.gp | 72.8 | 2.1 | 0.13 | |
| dist.poisson.gp | 73.2 | 2.5 | 0.11 | |
| diste.poisson.gp | 73.3 | 2.6 | 0.10 | |
| Canary Islands | fulle.poisson.gp | 70.8 | 0.0 | 0.20 |
| area.poisson.gp | 70.9 | 0.1 | 0.18 | |
| elev.poisson.gp | 71.0 | 0.2 | 0.17 | |
| full.poisson.gp | 71.2 | 0.4 | 0.16 | |
| dist.poisson.gp | 71.3 | 0.5 | 0.15 | |
| diste.poisson.gp | 71.6 | 0.8 | 0.13 |
| Archipelago | Model | WAIC | ΔWAIC | Weight |
|---|---|---|---|---|
| Azores | fulle.poisson.gp | 57.8 | 0.0 | 0.45 |
| diste.poisson.gp | 59.7 | 2.0 | 0.17 | |
| full.poisson.gp | 60.3 | 2.6 | 0.13 | |
| Canary Islands | dist.poisson.gp | 61.3 | 0.0 | 0.19 |
| elev.poisson.gp | 61.4 | 0.1 | 0.18 | |
| diste.poisson.gp | 61.4 | 0.1 | 0.18 | |
| area.poisson.gp | 61.6 | 0.3 | 0.17 | |
| full.poisson.gp | 61.6 | 0.3 | 0.16 | |
| fulle.poisson.gp | 62.2 | 0.9 | 0.12 |
| Archipelago | Model | WAIC | ΔWAIC | Weight |
|---|---|---|---|---|
| Azores | area.poisson.gp | 69.6 | 0.0 | 0.4.0 |
| fulle.poisson.gp | 70.2 | 0.6 | 0.29 | |
| dist.poisson.gp | 72.1 | 2.6 | 0.11 | |
| elev.poisson.gp | 72.4 | 2.8 | 0.10 | |
| diste.poisson.gp | 72.4 | 2.9 | 0.10 | |
| Canary Islands | elev.poisson.gp | 68.7 | 0.0 | 0.23 |
| dist.poisson.gp | 68.8 | 0.1 | 0.22 | |
| fulle.poisson.gp | 68.9 | 0.2 | 0.20 | |
| area.poisson.gp | 68.9 | 0.3 | 0.20 | |
| diste.poisson.gp | 69.5 | 0.9 | 0.15 |
| C | Fl | Fa | P | G | SJ | T | SM | SMa | |
| C | 1.00 | 1.14 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fl | 1.14 | 1.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 |
| Fa | 0.00 | 0.00 | 1.00 | 1.20 | 2.74 | 1.72 | 14.85 | 0.00 | 0.00 |
| P | 0.00 | 0.00 | 1.20 | 1.00 | 2.20 | 1.17 | 5.13 | 0.00 | 0.00 |
| G | 0.00 | 0.00 | 2.74 | 2.20 | 1.00 | 1.40 | 2.63 | 0.00 | 0.00 |
| SJ | 0.00 | 0.00 | 1.72 | 1.17 | 1.40 | 1.00 | 2.25 | 0.00 | 0.00 |
| T | 0.00 | 0.00 | 14.85 | 5.13 | 2.63 | 2.25 | 1.00 | 204.25 | 0.00 |
| SM | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 204.25 | 1.00 | 4.54 |
| SMa | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | 4.54 | 1.00 |
| F | L | GC | T | LG | EH | LP | |
| F | 1.00 | 1.44 | 5.08 | 47.14 | 0.00 | 0.00 | 0.00 |
| L | 1.44 | 1.00 | 24.88 | 0.00 | 0.00 | 0.00 | 0.00 |
| GC | 5.08 | 24.88 | 1.00 | 1.88 | 5.22 | 37.23 | 35.4 |
| T | 47.14 | 0.00 | 1.88 | 1.00 | 1.31 | 4.51 | 3.11 |
| LG | 0.00 | 0.00 | 5.22 | 1.31 | 1.00 | 1.56 | 1.60 |
| EH | 0.00 | 0.00 | 37.23 | 4.51 | 1.56 | 1.00 | 1.99 |
| LP | 0.00 | 0.00 | 35.4 | 3.11 | 1.6 | 1.99 | 1.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barros, D.D.; Mathias, M.d.L.; Borges, P.A.V.; Borda-de-Água, L. The Importance of Including Spatial Autocorrelation When Modelling Species Richness in Archipelagos: A Bayesian Approach. Diversity 2023, 15, 127. https://doi.org/10.3390/d15020127
Barros DD, Mathias MdL, Borges PAV, Borda-de-Água L. The Importance of Including Spatial Autocorrelation When Modelling Species Richness in Archipelagos: A Bayesian Approach. Diversity. 2023; 15(2):127. https://doi.org/10.3390/d15020127
Chicago/Turabian StyleBarros, Diogo Duarte, Maria da Luz Mathias, Paulo A. V. Borges, and Luís Borda-de-Água. 2023. "The Importance of Including Spatial Autocorrelation When Modelling Species Richness in Archipelagos: A Bayesian Approach" Diversity 15, no. 2: 127. https://doi.org/10.3390/d15020127
APA StyleBarros, D. D., Mathias, M. d. L., Borges, P. A. V., & Borda-de-Água, L. (2023). The Importance of Including Spatial Autocorrelation When Modelling Species Richness in Archipelagos: A Bayesian Approach. Diversity, 15(2), 127. https://doi.org/10.3390/d15020127