
Rotifers of Inter-Forest Springs

Abstract
:1. Introduction
2. Material and Methods
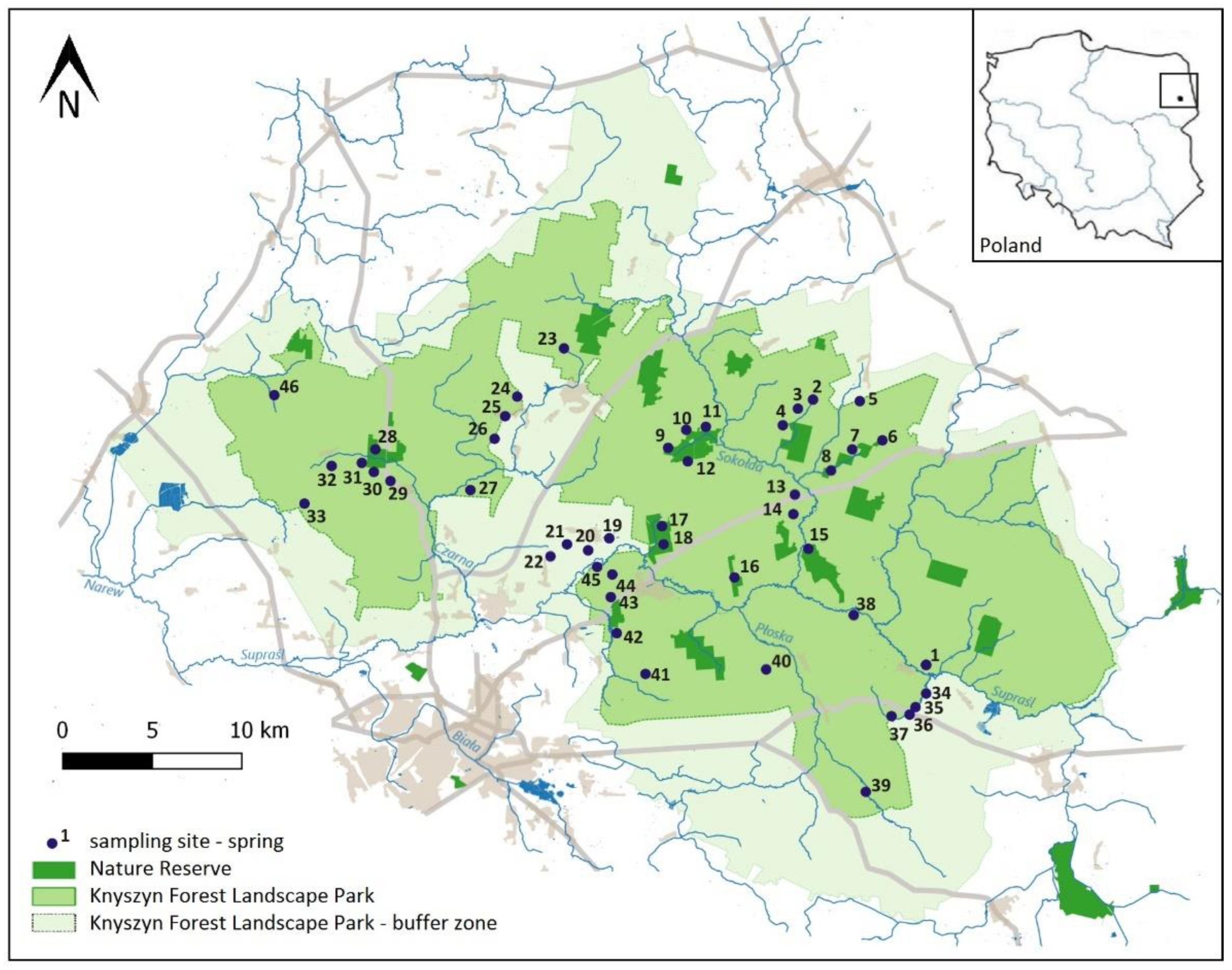
2.1. Study Area
2.2. Sampling Methods
2.3. Analyses
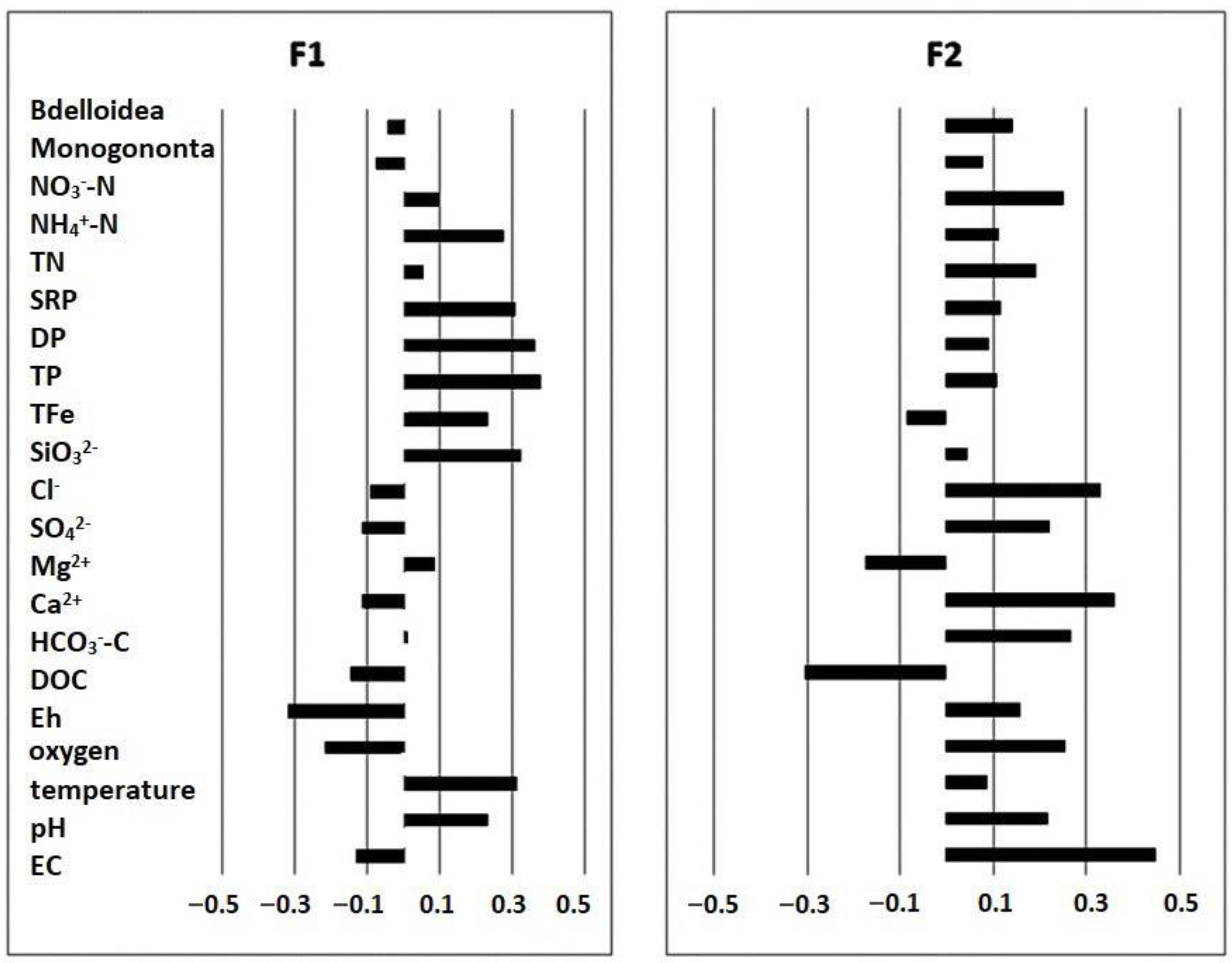
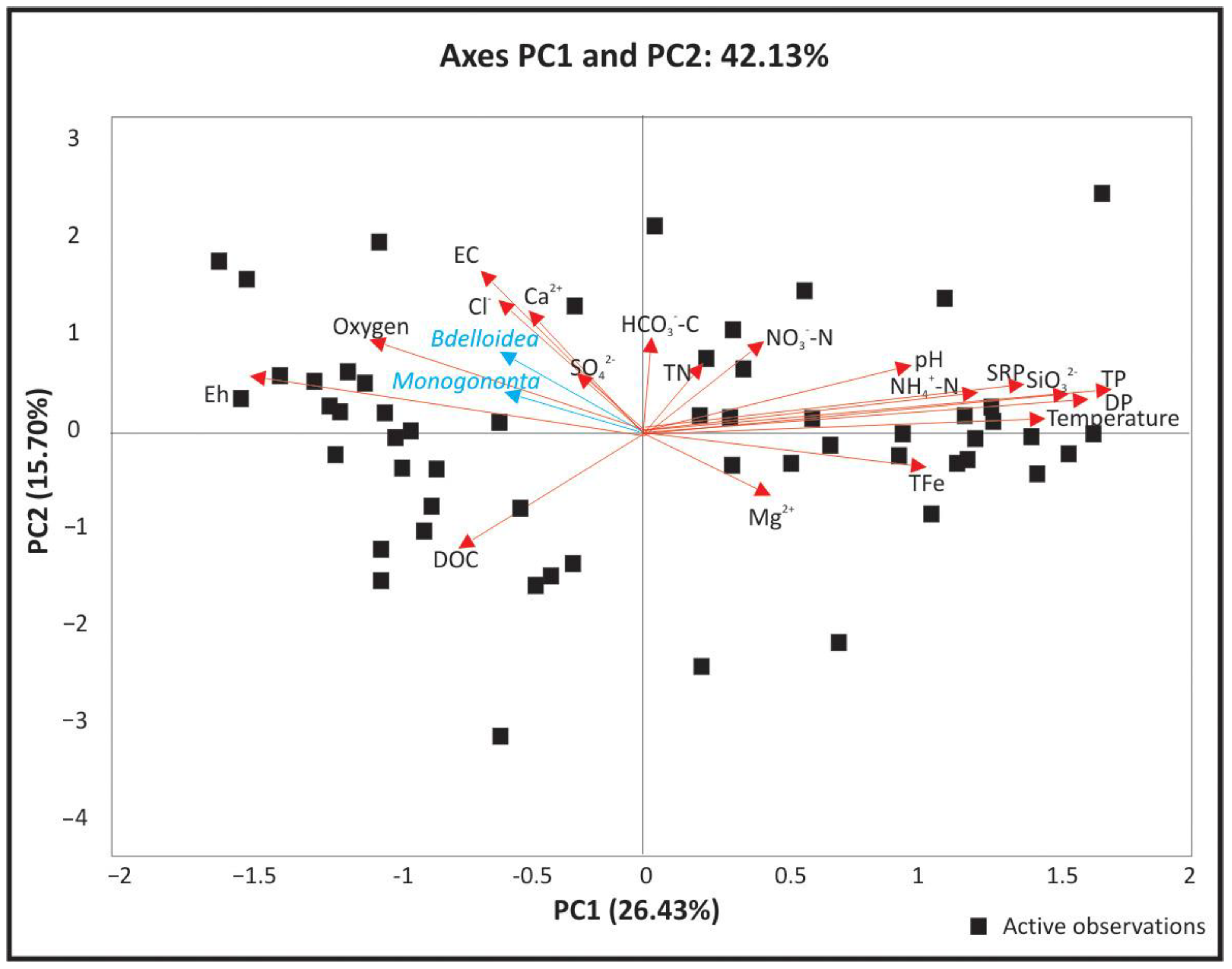
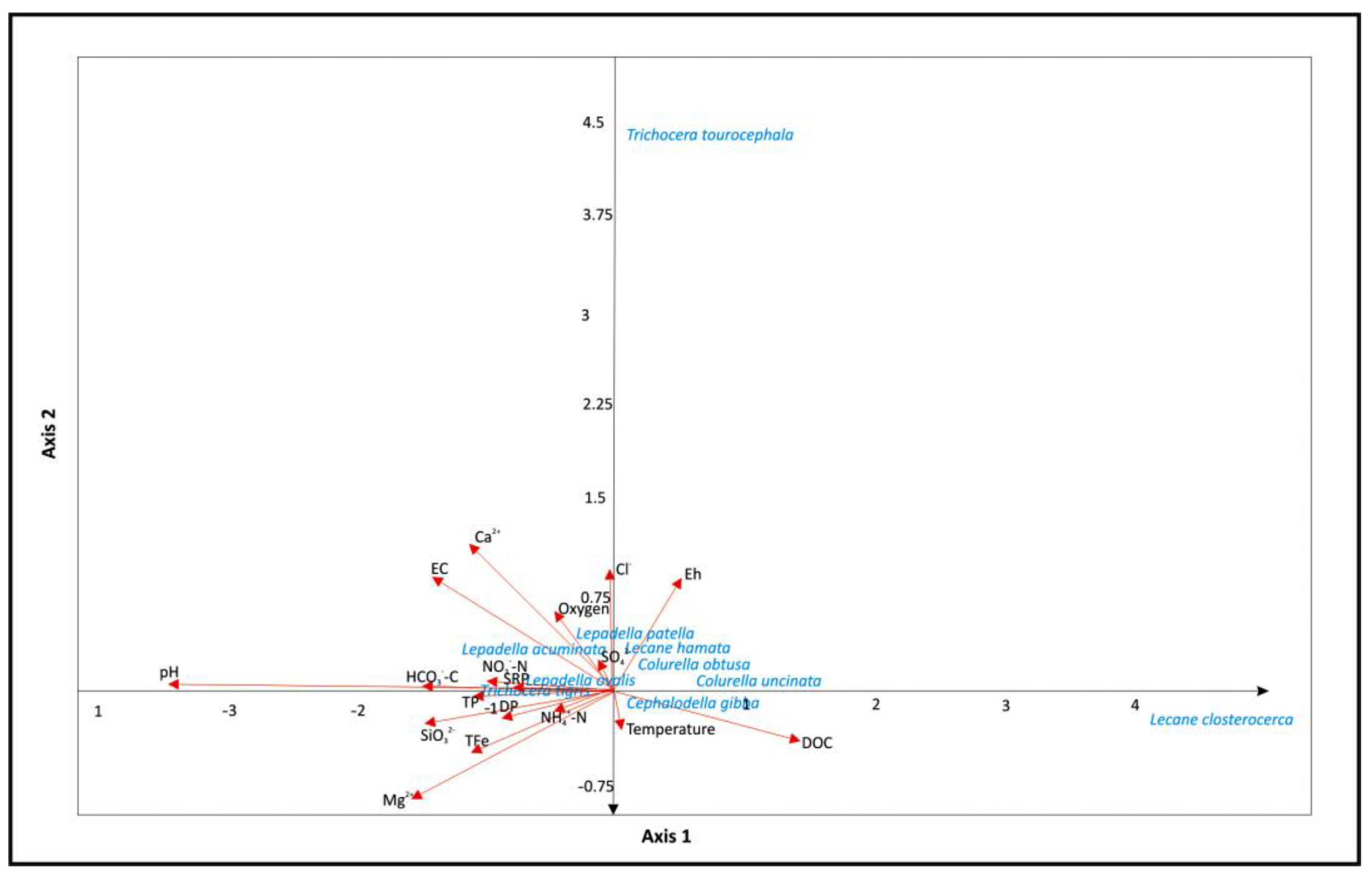
3. Results
3.1. Water Quality in Springs
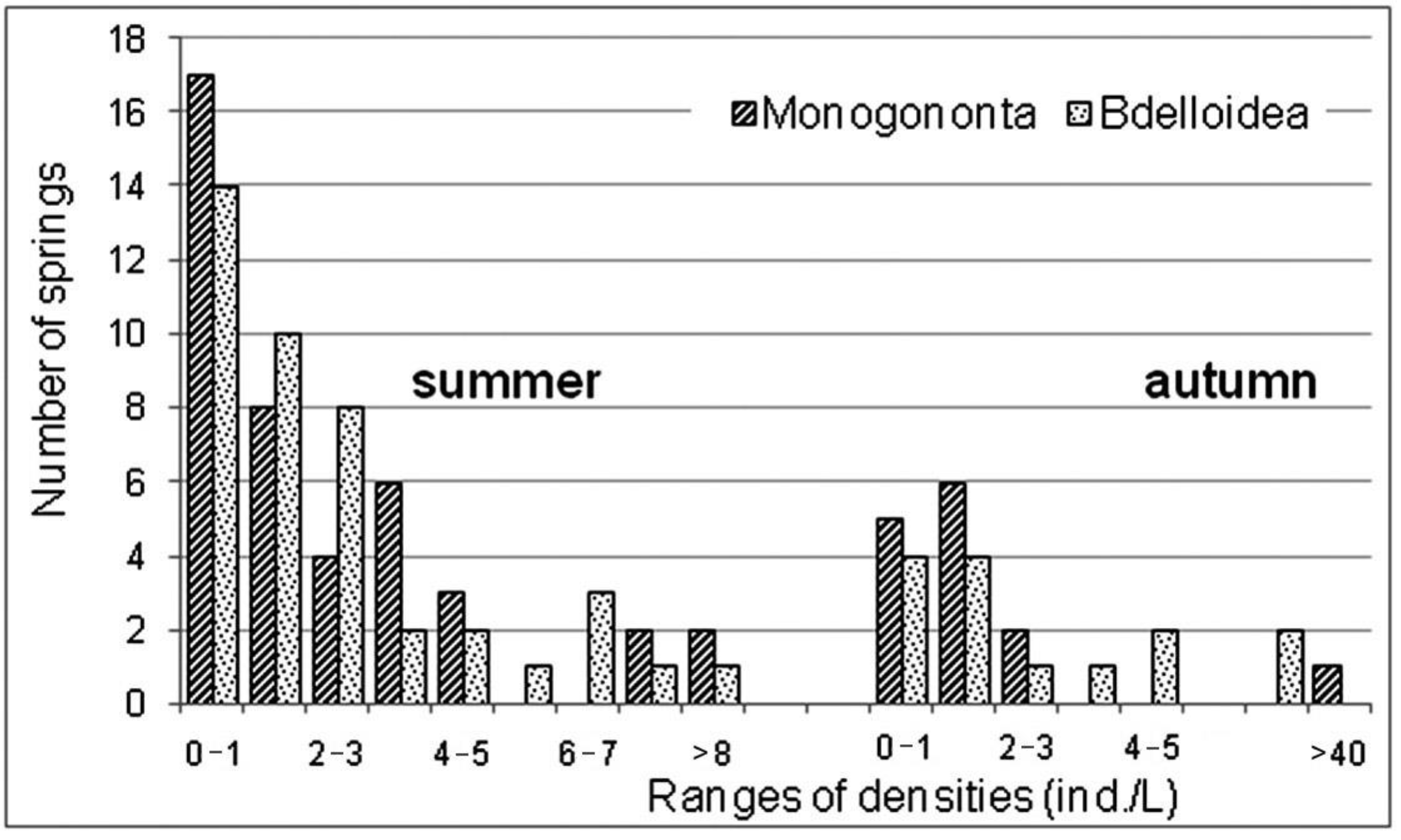
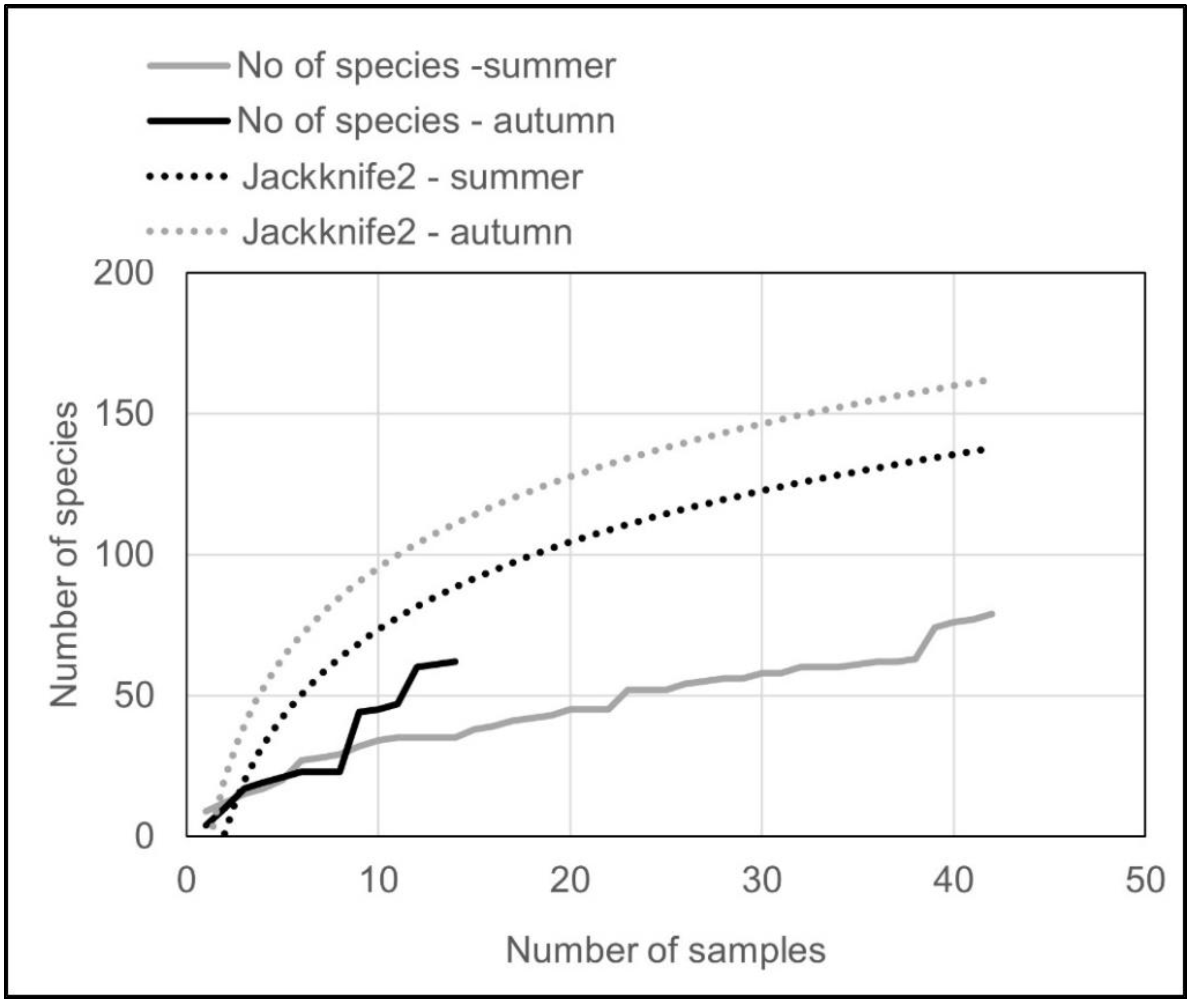
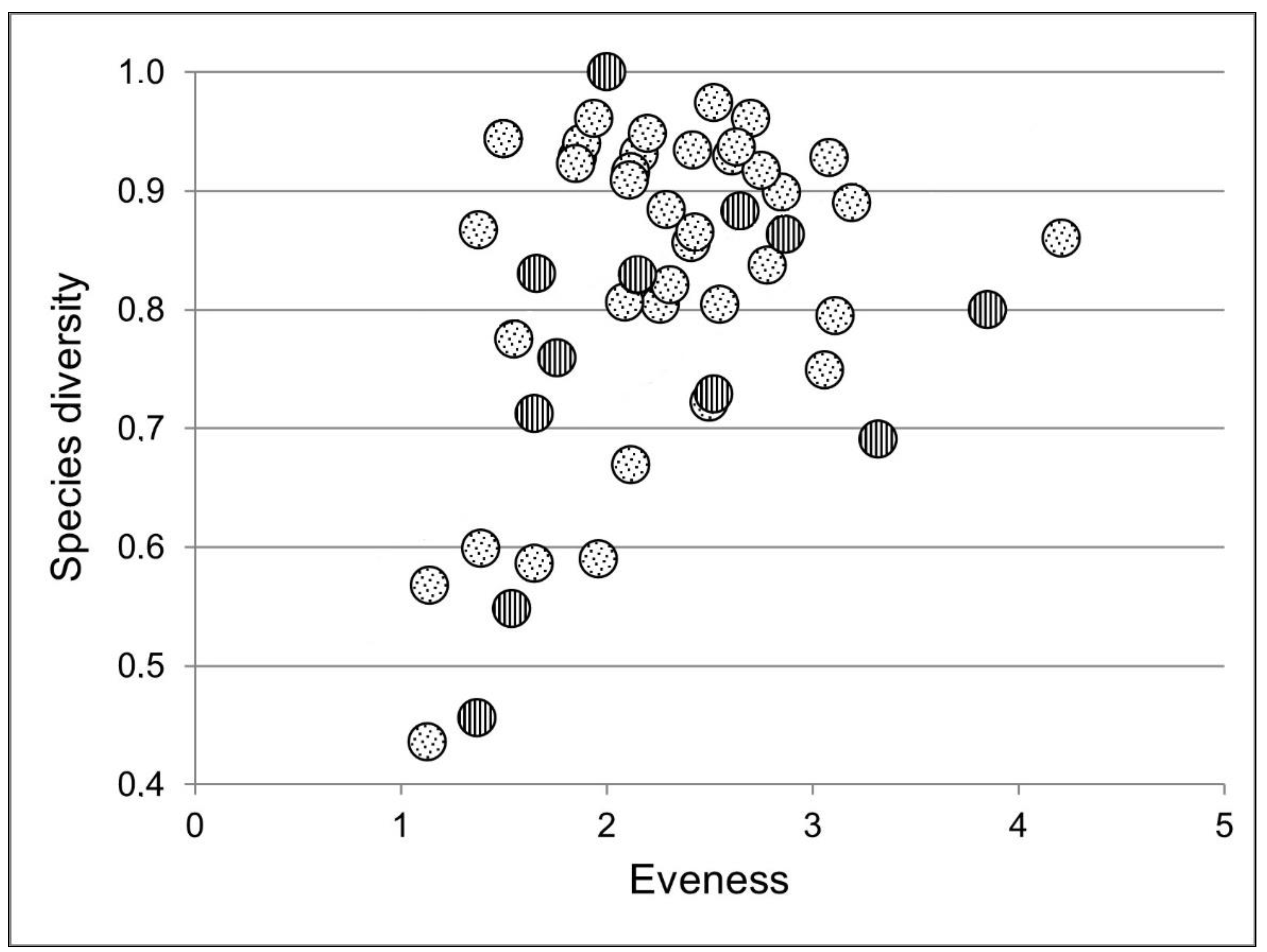
3.2. Structure of Rotifer Communities
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Erman, N.A.; Erman, D.C. Spring permanence. Trichoptera species richness and the role of drought. Kans. Entomol. Soc. Suppl. 1995, 68, 50–64. [Google Scholar]
- Cantonati, M.; Spitale, D.; Calfi, A.S.; Guealla, G. Exploring the contrasting seasonal strategies of two crenic macroalgae. Fottea Olomouc 2016, 16, 133–143. [Google Scholar] [CrossRef] [Green Version]
- Cantonati, M.; Lichtenwöhrer, K.; Leonhardt, G.; Seifert, L.; Mustoni, A.; Hotzy, R.; Schubert, E.; Blattner, L.; Bilous, O.; Lotz, A.; et al. Using Springs as Sentinels of Climate Change inNature Parks North and South of theAlps: A Critical Evaluation ofMethodological Aspects and Recommendations for Long-Term Monitoring. Water 2022, 14, 2843. [Google Scholar] [CrossRef]
- Bottazzi, E.; Bruno, M.C.; Pieri, V.; Di Sabatino, A.; Silveri, L.; Carolli, M.; Rossetti, G. Spatial and seasonal distribution of invertebrates in Northern Apennine rheocrene spring. J. Limnol. 2011, 70 (Suppl. 1), 77–92. [Google Scholar] [CrossRef]
- Meyer, J.L.; Wallace, J.B. Lost Linkages and Lotic Ecology: Rediscovering Small Streams; Blackwell Scientific: Oxford, UK, 2001. [Google Scholar]
- Meyer, J.L.; Strayer, D.L.; Wallace, J.B.; Eggert, S.L.; Helfman, G.S.; Leonard, N.E. The contribution of headwater streams to biodiversity in river networks. J. Am. Water Resour. Assoc. 2007, 43, 86–103. [Google Scholar] [CrossRef] [Green Version]
- Richardson, J.S.; Danehy, R.J. A synthesis of the ecology of headwater streams and their riparian zones in temperate forests. For. Sci. 2007, 53, 131–147. [Google Scholar]
- Leibold, M.A.; Holyoak, M.; Mouquet, N.; Amarasekare, P.; Chase, J.M.; Hoopes, M.F.; Holt, R.D.; Shurin, J.B.; Law, R.; Tilman, D.; et al. The metacommunity concept: A framework for multi-scale community ecology. Ecol. Lett. 2004, 7, 601–613. [Google Scholar] [CrossRef]
- Scarsbrook, M.; Barquin, J.; Gray, D.P. New Zealand Coldwater Springs and Their Biodiversity; Science for Conservation; Department of Conservation: Wellington, New Zealand, 2007; Volume 278. [Google Scholar]
- Botosaneanu, L. Studies in Crenobiology: The Biology of Springs and Springbrooks; Blackhuys Publishers: Leiden, The Netherlands, 1998. [Google Scholar]
- Hoffsten, P.; Malmqvist, B. The macroinvertebrate fauna and hydrogeology of springs in central Sweden. Hydrobiologia 2000, 436, 91–104. [Google Scholar] [CrossRef]
- Zawal, A.; Stryjecki, T.; Buczyńska, E.; Buczyński, P.; Pakulnicka, J.; Bańkowska, A.; Czernicki, T.; Janusz, K.; Szlauer-Łukaszewska, A.; Pešić, V. Water mites (Acari, Hydrachnidia) of riparian springs in a small lowland river valley: What are the key factors for species distribution? PeerJ 2018, 6. [Google Scholar] [CrossRef] [Green Version]
- Staudacher, K.; Fureder, L. Habitat complexity and invertebrates in selected alpine springs (Schütt. Carinthia. Austria). Int. Rev. Hydrobiol. 2007, 92, 465–479. [Google Scholar] [CrossRef]
- Cantonati, M.; Gerecke, R.; Bertuzzi, E. Springs of the Alps—Sensitive ecosystems to environmental change: From biodiversity assessments to long-term studies. Hydrobiologia 2006, 562, 59–96. [Google Scholar] [CrossRef]
- Von Fumetti, S.; Blattner, L. Faunistic assemblages of natural springs in different areas in the Swiss National Park: A small-scale comparison. Hydrobiologia 2017, 793, 175–184. [Google Scholar] [CrossRef]
- Jekatierynczuk-Rudczyk, E.; Puczko, K.; Żukowska, J.; Sawicka, A. Biota communities influence on nutrients circulation in hyporheic zone—A case study in urban spring niches in Bialystok (NE Poland). Aquat. Sci. 2021, 83, 75. [Google Scholar] [CrossRef]
- Reiss, J.; Schmid-Araya, J.M. Existing in plenty: Abundance, biomass and diversity of ciliates and meiofauna in small streams. Freshw. Biol. 2008, 53, 652–668. [Google Scholar] [CrossRef]
- Schmid-Araya, J. Rotifers in interstitial sediments. Hydrobiologia 1998, 387/388, 231–240. [Google Scholar] [CrossRef]
- Ríos-Arana, J.V.; Agüero-Reyes, L.D.C.; Wallace, R.L.; Walsh, E.J. Limnological characteristics and rotifer community composition of Northern Mexico Chihuahuan Desert Springs. J. Arid. Environ. 2019, 160, 32–41. [Google Scholar] [CrossRef]
- Brown, P.D.; Schröder, T.; Ríos-Arana, J.V.; Rico-Martinez, R.; Silva-Briano, M.; Wallace, R.L.; Walsh, E.J. Patterns of Rotifer Diversity in the Chihuahuan Desert. Diversity 2020, 12, 393. [Google Scholar] [CrossRef]
- Jekatierynczuk-Rudczyk, E.; Zieliński, P.; Puczko, K.; Micun, K.; Puczyłowska, E. The Role of the Catchment Area in Shaping Water Quality in the Lowland Springs of the Knyszyn Forest (NE Poland). Water 2022, 14, 3202. [Google Scholar] [CrossRef]
- JekatierynczukRudczyk, E. Effects of drainage basin managment on the chemical composition of waters in lowlands springs. Acta Hydrobiol. 1999, 41, 97–105. [Google Scholar]
- Jekatierynczuk-Rudczyk, E. Transformation in Physicochemical Composition of Groundwaters within Lowland Drainage Zones; Wydawnictwo Uniwersytetu w Białymstoku: Białystok, Poland, 2010. [Google Scholar]
- Puczko, K.; Zieliński, P.; Jusik, S.; Kołakowska, A.; JekatierynczukRudczyk, E. Vascular plant and bryophyte species richness in response to water quality in lowland spring niches with different anthropogenic impacts. Environ. Monit. Assess. 2018, 190, 338. [Google Scholar] [CrossRef] [Green Version]
- Zieliński, P.; Jekatierynczuk-Rudczyk, E.; Puczko, K. Factors affecting the abundance and activity of the bacterioplankton in lowland forest springs in northeastern Poland. Ecohydrol. Hydrobiol. 2020, 20, 675–686. [Google Scholar] [CrossRef]
- Górniak, A. Wody Krajobrazowego Parku Puszczy Knyszyńskiej (Waters of the Landscape Park of te Knyszyn Forest); Abstract; Park Krajobrazowy Puszczy Knyszyńskiej: Supraśl, Poland, 1999; Volume 48. (In Polish) [Google Scholar]
- Jekatierynczuk-Rudczyk, E.; Zieliński, P.; Puczko, K. Is the protection of springs in Knyszyn Forest effective and satisfactory? Prot. Nativ. Nat. 2017, 73, 135–147, (In Polish with English summary). [Google Scholar]
- Łoszewski, H. Springs in the Supraśl river basin. Sci. Pract. 1984, OBN 4, 111–150. (In Polish) [Google Scholar]
- Pawłowska, T.; Miodek, K. Physico-geographic characteristics. In Natural and Cultural Documentation along with the Principles of Spatial Management; Gacka-Grześkiewicz, E., Ed.; Knyszyn Forest Landscape Park: Knyszyn, Poland, 1993; pp. 14–39. (In Polish) [Google Scholar]
- Jersabek, C.D.; Leitner, M.F.; The Rotifer World Catalog. World Wide Web Electronic Publication. 2013. Available online: http://www.rotifera.hausdernatur.at/ (accessed on 2 October 2022).
- APHA. Standard Methods for Examination of Water and Waste Water; APHA, AWWA: Washington, DC, USA, 1992. [Google Scholar]
- Basualdo, C.V. Choosing the best nonparametric richness estimator for benthic macroinvertebrates databases. Rev. Soc. Entomol. Argent. 2011, 70, 27–38. [Google Scholar]
- Margalef, R. Information theory in ecology. Gen. Syst. 1958, 3, 36–71. [Google Scholar]
- Vialle, C.; Sablayrolles, C.; Lovera, M.; Jacob, S.; Huau, M.C.; Montréjaud-Vignoles, M. Monitoring of water quality from roof runoff: Interpretation using multivariate analysis. Water Res. 2011, 45, 3765–3775. [Google Scholar] [CrossRef] [Green Version]
- Helena, B.; Pardo, R.; Vega, M.; Barrado, E.; Fernandez, J.M.; Fernandez, L. Temporal evolution of groundwater composition in an alluvial aquifer (Pisuerga River, Spain) by principal component analysis. Water Res. 2000, 34, 807–816. [Google Scholar] [CrossRef]
- Sliva, L.; Williams, D.D. Buffer zone versus whole catchment approaches to studying land use impact on river water quality. Water Res. 2001, 35, 3462–3472. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Suring, L.H. Lotic Freshwater: Springs. In Encyclopedia of the World’s Biomes; Elsevier: Amsterdam, The Netherlands, 2020; pp. 131–133. [Google Scholar] [CrossRef]
- Barquín, P.; Scarsbrook, M. Management and conservation strategies for coldwater springs. Aquat. Conserv. Mar. Freshw. Ecosyst. 2008, 18, 580–591. [Google Scholar] [CrossRef]
- Pradhan, N. Conserving Springs and its Significance for Water Security in Himalayas. 2005. Available online: https://thesarcist.org/Opinion/103 (accessed on 20 November 2022).
- Hakenkamp, C.C.; Morin, A. The importance of meiofauna to lotic ecosystem functioning. Freshw. Biol. 2000, 44, 165–175. [Google Scholar] [CrossRef]
- Sarkka, J.; Levonen; Makela, J. Meiofauna of spring in Finland in relation to environmental factors. Hydrobiologia 1997, 347, 139–150. [Google Scholar] [CrossRef]
- Robertson, A.L.; Rundle, S.D.; Schmid-Araya, J.M. Putting the meio- into stream ecology: Current findings and future directions for lotic meiofauna research. Freshw. Biol. 2000, 44, 177–183. [Google Scholar] [CrossRef]
- Wallace, R.L.; Walsh, E.J.; Arroyo, M.L.; Starkweather, P.L. Life on the edge: Rotifers from springs and ephemeral waters in the Chihuahuan Desert. Big Bend National Park (Texas. USA). Hydrobiologia 2005, 546, 147–157. [Google Scholar] [CrossRef]
- Schmid-Araya, J. Benthic rotifera inhabiting the bed sediments of a mountain gravel stream. J. Biol. Stn. Lunz 1993, 14, 75–101. [Google Scholar]
- Ejsmont-Karabin, J. Vertical microzonation of psammon rotifers (Rotifera) in the psammolittoral habitat of an eutrophic lake. Pol. J. Ecol. 2008, 56, 351–357. [Google Scholar]
- Ejsmont-Karabin, J.; Radwan, S.; Bielańska-Grajner, I. Monogononta—Atlas gatunków. 32B. In Wrotki (Rotifera). Fauna słodkowodna Polski. 32. Polskie Towarzystwo Hydrobiologiczne; Radwan, S., Ed.; Uniwersytet Łódzki, Oficyna Wydawnicza Tercja: Łódź, Poland, 2004; pp. 147–448. [Google Scholar]
- Gathmann, F.O.; Manne, L.L.; Williams, D.D. Spatial patterns in insect community composition in coldwater springs. Aquat Ecol. 2009, 42, 501–512. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | Geographical Coordinates | Hydrological Location | Hydrobiology Type of Springs | Hydrochemistry Type of Water | Land Use |
|---|---|---|---|---|---|
| 1. | N: 53°09′29.32″; E: 23°34′16.0″ | right-side direct tributary of the Supraśl river → Radulinka catchment | artificial limnocrene | HCO3-Ca-Mg | rural area |
| 2. | N: 53°18′28.05″; E: 23°26′57.28″ | Sokołda catchment → Karnicha catchment | rheocrene | HCO3-Ca-Mg | forest/grassland |
| 3. | N: 53°17′42.44″; E: 23°25′37.06″ | Sokołda catchment → Karnicha catchment | rheocrene | HCO3-Ca-Mg | forest |
| 4. | N: 53°16′41.53″; E: 23°25′04.8″ | Sokołda catchment → Karnicha catchment | rheocrene | HCO3-Ca-Mg | forest |
| 5. | N: 53°17′43.72″; E: 23°30′ 21.1″ | Sokołda catchment → Łanga catchment | rheocrene | HCO3-Ca-Mg | rural area |
| 6. | N: 53°15′23.85″; E: 23°30′03.88″ | Sokołda catchment → Łanga catchment | rheocrene | HCO3-Ca-Mg | rural area |
| 7. | N: 53°16′52.31″; E: 23°30′51.01″ | Sokołda catchment → Łanga catchment | rheocrene | HCO3-Ca | rural area |
| 8. | N: 53°16′05.05″; E: 23°31′01.89″ | Sokołda catchment → Łanga catchment | rheocrene | HCO3-Ca-Mg | rural area |
| 9. | N: 53°16′53.67″; E: 23°22′15.75″ | Sokołda catchment → Migówka catchment | rheocrene | HCO3-Ca | forest |
| 10. | N: 53°16′54.62″; E: 23°22′27.36″ | Sokołda catchment → Migówka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 11. | N: 53°16′49.47″; E: 23°22′44.93″ | Sokołda catchment → Migówka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 12. | N: 53°16′26.07″; E: 23°21′24.73″ | Sokołda catchment → Migówka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 13. | N: 53°14′04.31″; E: 23°29′43.08″ | direct tributary of the Sokołda river → Sokołda catchment | rheocrene | HCO3-Ca | grassland |
| 14. | N: 53°14′35.73″; E: 23°27′38.92″ | direct tributary of the Sokołda river → Sokołda catchment | helocrene | HCO3-Ca | grassland |
| 15. | N: 53°14′53.01″; E: 23°28′59.46″ | direct tributary of the Sokołda river → Sokołda catchment | limnocrene | HCO3-Ca-Mg | forest |
| 16. | N: 53°12′23.77″; E: 23°26′02.44″ | direct tributary of the Sokołda river → Sokołda catchment | helocrene | HCO3-Ca-Mg | rural area |
| 17. | N: 53°14′30.49″; E: 23°20′50.34″ | Sokołda catchment → Jałówka catchment | rheocrene | HCO3-Ca | forest/grassland |
| 18. | N: 53°14′04.58″; E: 23°20′41.57″ | Sokołda catchment → Jałówka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 19. | N: 53°14′28.01″; E: 23°18′07.49″ | right-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca-Mg | forest/grassland |
| 20. | N: 53°13′34.82″; E: 23°15′45.74″ | right-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca | forest/grassland |
| 21. | N: 53°14′31.25″; E: 23°19′12.46″ | right-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca | grassland |
| 22. | N: 53°14′31.38″; E: 23°17′15.09″ | right-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca | grassland |
| 23. | N: 53°19′13.31″; E: 23°15′18.06″ | Czarna catchment → Czapielówka catchment | helocrene | HCO3-Ca | rural area |
| 24. | N: 53°17′24.01″; E: 23°12′25.18″ | Czarna catchment → Czarna Rzeczka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 25. | N: 53°16′50.22″; E: 23°11′31.01″ | direct tributary of the Czarna river → Czarna catchment | rheocrene | HCO3-Ca-Mg | forest |
| 26. | N: 53°16′03.72″; E: 23°12′56.16″ | Czarna catchment → Czarna Rzeczka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 27. | N: 53°15′35.27″; E: 23°10′00.62″ | direct tributary of the Czarna river → Czarna catchment | helocrene | HCO3-Ca | grassland |
| 28. | N: 53°16′55.79″; E: 23°07′05.98″ | Czarna catchment → Krzemianka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 29. | N: 53°16′38.56″; E: 23°03′30.18″ | Czarna catchment → Krzemianka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 30. | N: 53°16′27.53″; E: 23°05′28.25″ | Czarna catchment → Krzemianka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 31. | N: 53°15′05.65″; E: 23°08′40.17″ | Czarna catchment → Krzemianka catchment | rheocrene | HCO3-Ca | forest |
| 32. | N: 53°14′46.25″; E: 23°08′10.17″ | Czarna catchment → Krzemianka catchment | helocrene | HCO3-Ca-Mg | forest |
| 33. | N: 53°13′09.44″; E: 23°01′24.03″ | right-side direct tributary of the Supraśl river → Supraśl catchment | helocrene | HCO3-Ca | rural area |
| 34. | N: 53°08′19.35″; E: 23°33′17.86″ | left-side direct tributary of the Supraśl river → stream from Sofipol catchment | rheocrene/limnocrene | HCO3-Ca | grassland |
| 35. | N: 53°08′17.93″; E: 23°33′12.44″ | left-side direct tributary of the Supraśl river → stream from Sofipol catchment | rheocrene | HCO3-Ca | grassland |
| 36. | N: 53°08′20.26″; E: 23°33′5.42″ | left-side direct tributary of the Supraśl river → stream from Sofipol catchment | rheocrene | HCO3-Ca-Mg | grassland |
| 37. | N: 53°08′21.2″; E: 23° 33′07.51″ | left-side direct tributary of the Supraśl river → stream from Sofipol catchment | rheocrene | HCO3-Ca-Mg | grassland |
| 38. | N: 53°10′35.29″; E: 23°30′03.76″ | left-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca-Mg | forest |
| 39. | N: 53°06′20.46″; E: 23°29′48.26″ | Płoska catchment → Świniobródka catchment | rheocrene | HCO3-Ca | forest |
| 40. | N: 53°09′42.33″; E: 23°27′18.34″ | direct tributary of the Płoska river → Płoska catchment | rheocrene | HCO3-Ca-Mg | forest |
| 41. | N: 53°09′52.69″; E: 23°21′41.36″ | left-side direct tributary of the Supraśl river → Starzynka catchment | rheocrene | HCO3-Ca-Mg | forest |
| 42. | N: 53°09′29.54″; E: 23°17′42.11″ | left-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca | forest |
| 43. | N: 53°09′43.62″; E: 23°17′47.22″ | left-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca-Mg | forest |
| 44. | N: 53°13′17.37″; E: 23°18′01.96″ | left-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca | forest |
| 45. | N: 53°13′13.9″; E: 23°18′5.54″ | left-side direct tributary of the Supraśl river → Supraśl catchment | rheocrene | HCO3-Ca-Mg | forest/grassland |
| 46. | N: 53°18′19.94″; E: 23°1′55.69″ | Narew catchment → Jaskranka catchment | helocrene | HCO3-Ca-Mg | rural area |
| 47. | N: 53°19′12.49″; E: 23°3′23.81″ | Narew catchment → Jaskranka catchment | rheocrene | HCO3-Ca | rural area |
| Parameters | Mean | SD | Minimum | Median | Maximum | |
|---|---|---|---|---|---|---|
| Temperature | °C | 15.18 | 3.18 | 9.5 | 15.20 | 21.2 |
| pH | 7.91 | 0.41 | 6.66 | 8.01 | 9.02 | |
| EC | µS·cm−1 | 403 | 72 | 283 | 385 | 607 |
| Eh | mV | 78.6 | 25.4 | 37.8 | 78.7 | 144.2 |
| Oxygen | mg· L−1 | 7.61 | 1.58 | 1.12 | 7.85 | 9.74 |
| Ca2+ | mg· L−1 | 78.1 | 13.6 | 56.9 | 75.2 | 122.6 |
| Mg2+ | mg· L−1 | 13.9 | 6.3 | 3.6 | 15.0 | 31.9 |
| HCO3−-C | mg· L−1 | 53.3 | 8.3 | 42.2 | 51.9 | 82.3 |
| Cl− | mg· L−1 | 11.5 | 8.1 | 5.5 | 9.1 | 41.5 |
| SO42− | mg· L−1 | 26.5 | 8.9 | 1.6 | 27.2 | 42.3 |
| SiO32− | mg· L−1 | 2.27 | 1.26 | 0.50 | 2.10 | 5.30 |
| TFe | mg· L−1 | 1.32 | 0.50 | 0.31 | 1.46 | 2.21 |
| TN | µg· | 1957 | 2767 | 203 | 1303 | 18160 |
| NO3−-N | µg· L−1 | 868 | 1200 | 52 | 384 | 4957 |
| NH4+-N | µg· L−1 | 278 | 120 | 117 | 244 | 600 |
| TP | µg· L−1 | 217 | 201 | 28 | 216 | 1311 |
| SRP | µg· L−1 | 79 | 110 | 13 | 61 | 717 |
| DP | µg· L−1 | 164 | 183 | 17 | 165 | 1174 |
| DOC | mg· L−1 | 4.15 | 4.59 | 1.09 | 2.34 | 23.38 |
| Species | Summer | Autumn | ||
|---|---|---|---|---|
| Maximum Numbers (ind. L−1) | Frequency (%) | Maximum Numbers (ind. L−1) | Frequency (%) | |
| Anuraeopsis fissa (Gosse, 1851) | 0.10 | 7.1 | 0.05 | * |
| Ascomorpha ovalis (Bergendal, 1892) | - | - | 0.02 | * |
| Asplanchna priodonta Gosse, 1850 | 0.25 | * | - | - |
| Brachionus angularis Gosse, 1851 | 0.10 | * | 0.05 | * |
| Brachionus bidentatus Anderson, 1889 | 0.02 | * | - | - |
| Brachionus calyciflorus s.s. Pallas, 1766 | - | - | 0.02 | 14.3 |
| Cephalodella auriculata (Müller, 1773) | 0.05 | * | - | - |
| Cephalodella catellina (Müller, 1786) | 0.05 | * | - | - |
| Cephalodella exigua (Gosse, 1886) | 0.25 | * | 0.05 | 14.3 |
| Cephalodella forficula (Ehrenberg, 1832) | 0.07 | * | - | - |
| Cephalodella gibba (Ehrenberg, 1832) | 0.20 | 14.3 | 0.05 | 21.4 |
| Cephalodella gracilis (Ehrenberg, 1832) | 0.25 | * | - | - |
| Cephalodella tenuiseta (Burn, 1890) | 0.07 | * | 0.03 | * |
| Cephalodella ventripes (Dixon-Nuttall, 1901) | 0.05 | * | 0.02 | * |
| Colurella adriatica Ehrenberg, 1831 | 1.80 | 73.8 | 1.20 | 57.1 |
| Colurella colurus (Ehrenberg, 1830) | 0.05 | 4.8 | 0.04 | 28.6 |
| Colurella geophila Donner, 1951 | 0.10 | 4.8 | - | - |
| Colurella hindenburgi Steinecke, 1917 | 0.05 | * | 0.05 | 14.3 |
| Colurella obtusa (Gosse, 1886) | 0.40 | 14.3 | 0.10 | 21.4 |
| Colurella uncinata (Müller, 1773) | 1.35 | 28.6 | 0.30 | 35.7 |
| Conochilus hippocrepis (Schrank, 1803) | 0.20 | * | - | - |
| Dicranophorus capucinus Harring & Myers, 1928 | 0.10 | * | - | - |
| Dicranophorus forcipatus (Muller, 1786) | - | - | 0.02 | * |
| Dicranophorus hercules Wiszniewski, 1932 | 0.25 | 4.8 | - | - |
| Dicranophorus luetkeni (Bergendal, 1892) | 0.10 | 7.1 | - | - |
| Dicranophorus rostratus (Dixon-Nuttall & Freeman, 1902) | 0.05 | * | - | - |
| Dicranophorus secretus Donner, 1951 | - | - | 0.02 | * |
| Encentrum diglandula (Zavadovsky, 1926) | - | - | 0.15 | * |
| Encentrum fluviatile Wulfert, 1939 | 0.05 | * | 0.04 | * |
| Encentrum lupus Wulfert, 1936 | 0.15 | 16.7 | 0.03 | * |
| Encentrum saundersiae (Hudson, 1885) | 0.05 | * | 0.02 | * |
| Encentrum sp | 0.10 | 4.8 | 0.25 | 21.4 |
| Encentrum uncinatum (Milne, 1886) | 0.05 | * | - | - |
| Eothinia elongata (Ehrenberg, 1832) | - | - | 0.01 | * |
| Euchlanis deflexa Gosse, 1851 | - | - | 0.02 | * |
| Euchlanis dilatata Ehrenberg, 1832 | 2.00 | 7.1 | - | - |
| Euchlanis incisa Carlin, 1939 | 0.60 | 4.8 | 0.01 | * |
| Keratella cochlearis (Gosse, 1851) | 1.00 | 9.5 | 0.05 | 28.6 |
| Keratella paludosa (Lucks, 1912) | 0.33 | * | - | - |
| Keratella quadrata (Müller, 1786) | - | - | 0.05 | 14.3 |
| Keratella ticinensis (Callerio, 1920) | - | - | 0.02 | * |
| Lecane acus (Harring, 1913) | 1.00 | 4.8 | - | - |
| Lecane closterocerca (Schmarda, 1859) | 3.65 | 52.4 | 0.44 | 35.7 |
| Lecane flexilis (Gosse, 1886) | - | - | 0.01 | * |
| Lecane hamata (Stokes, 1896) | 0.60 | 38.1 | 0.10 | 28.6 |
| Lecane luna (Müller, 1776) | 0.05 | * | - | - |
| Lecane lunaris (Ehrenberg, 1832) | 0.60 | * | 0.02 | * |
| Lecane opias (Harring & Myers, 1926) | 0.05 | * | - | - |
| Lecane perpusilla (Hauer, 1929) | 0.10 | * | - | - |
| Lecane stichaea Harring, 1913 | 0.05 | * | - | - |
| Lepadella acuminata (Ehrenberg, 1834) | 2.75 | 66.7 | 5.00 | 85.7 |
| Lepadella costata Wulfert, 1940 | 0.20 | 4.8 | - | - |
| Lepadella ovalis (Müller, 1786) | 0.25 | 26.2 | 36.5 | 50.0 |
| Lepadella patella (Müller, 1773) | 3.67 | 45.2 | 0.53 | 78.6 |
| Lepadella quadricarinata (Stenroos, 1898) | - | - | 0.05 | 14.3 |
| Lepadella rhomboides (Gosse, 1886) | - | - | 0.05 | * |
| Lepadella rottenburgi (Lucks, 1912) | 0.05 | * | - | - |
| Lepadella triba Myers, 1934 | 0.20 | * | - | - |
| Lepadella triptera (Ehrenberg, 1830) | - | - | 0.01 | * |
| Lindia truncata (Jennings, 1894) | 0.02 | * | - | - |
| Lophocharis oxysternoon (Gosse, 1851) | 0.02 | * | 0.10 | * |
| Lophocharis salpina (Ehrenberg, 1834) | 0.10 | * | 0.03 | * |
| Microcodon clavus Ehrenberg, 1830 | - | - | 0.02 | * |
| Monommata longiseta (Müller, 1786) | - | - | 0.02 | * |
| Monommata phoxa Myers, 1930 | - | - | 0.02 | * |
| Mytilina mucronata (Muller, 1773) | 0.05 | 4.8 | 0.02 | * |
| Mytilina ventralis (Ehrenberg, 1832) | 0.08 | * | 0.01 | * |
| Notholca squamula (Müller, 1786) | 1.00 | * | 0.05 | * |
| Notommata aurita (Müller, 1786) | - | - | 0.02 | * |
| Notommata cyrtopus Gosse, 1886 | 0.05 | * | 0.02 | * |
| Notommata tripus Ehrenberg, 1838 | 0.05 | * | - | - |
| Paradicranophorus aculeatus (Neizvestnova-Zhadina, 1935) | 0.25 | * | 0.15 | * |
| Paradicranophorus hudsoni (Glascott, 1893) | 0.95 | 11.9 | 0.52 | * |
| Pleurotrocha petromyzon Ehrenberg, 1830 | 0.40 | 4.8 | 0.20 | 14.3 |
| Polyarthra major Burckhardt, 1900 | - | - | 0.02 | * |
| Pompholyx sulcata Hudson, 1885 | 1.40 | 4.8 | - | - |
| Proales globulifera (Hauer, 1921) | 0.15 | 4.8 | 0.02 | * |
| Proales micropus (Gosse, 1886) | 0.05 | * | 0.07 | * |
| Proales sigmoidea Skorikov, 1896 | 0.02 | * | - | - |
| Proales theodora (Gosse, 1887) | 0.20 | 9.5 | - | - |
| Ptygura melicerta Ehrenberg, 1832 | 0.05 | 4.8 | - | - |
| Resticula melandocus (Gosse, 1887) | 0.10 | 4.8 | - | - |
| Squatinella lamellaris (Müller, 1786) | 0.95 | * | 0.05 | 21.4 |
| Squatinella rostrum (Schmarda, 1846) | 0.10 | * | - | - |
| Synchaeta longipes Gosse, 1887 | - | - | 0.05 | * |
| Synchaeta oblonga Ehrenberg, 1831 | 0.20 | * | 0.76 | 21.4 |
| Testudinella caeca (Parsons, 1892) | 0.02 | * | - | - |
| Testudinella mucronata (Gosse, 1886) | 0.05 | * | - | - |
| Testudinella patina (Hermann, 1783) | - | - | 0.01 | - |
| Testudinella truncata (Gosse, 1886) | 0.02 | * | 0.01 | - |
| Trichocerca myersi (Hauer, 1931) | 0.10 | 4.8 | - | - |
| Trichocerca pusilla (Jennings, 1903) | 0.12 | * | 0.04 | * |
| Trichocerca similis (Wierzejski, 1893) | 0.20 | * | - | - |
| Trichocerca taurocephala (Hauer, 1931) | 7.90 | 61.9 | 0.30 | 14.3 |
| Trichocerca tenuior (Gosse, 1886) | 0.05 | 7.1 | - | - |
| Trichocerca tigris (Müller, 1786) | 1.55 | 38.1 | 0.10 | 14.3 |
| Trichocerca vernalis (Hauer, 1936) | 0.05 | * | - | - |
| Trichotria pocillum (Müller, 1776) | - | - | 0.03 | 14.3 |
| Trichotria tetractis (Ehrenberg, 1830) | - | - | 0.02 | * |
| Wierzejskiella velox (Wiszniewski, 1932) | 0.05 | * | - | - |
| Principal Component | ||||||
|---|---|---|---|---|---|---|
| F1 | F2 | F3 | F4 | F5 | F6 | |
| Eigenvalue | 5.55 | 3.30 | 1.97 | 1.59 | 1.32 | 1.01 |
| Variability (%) | 26.43 | 15.70 | 9.36 | 7.55 | 6.29 | 4.81 |
| Cumulative % | 26.43 | 42.13 | 51.50 | 59.05 | 65.33 | 70.14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jekatierynczuk-Rudczyk, E.; Ejsmont-Karabin, J. Rotifers of Inter-Forest Springs. Diversity 2023, 15, 153. https://doi.org/10.3390/d15020153
Jekatierynczuk-Rudczyk E, Ejsmont-Karabin J. Rotifers of Inter-Forest Springs. Diversity. 2023; 15(2):153. https://doi.org/10.3390/d15020153
Chicago/Turabian StyleJekatierynczuk-Rudczyk, Elżbieta, and Jolanta Ejsmont-Karabin. 2023. "Rotifers of Inter-Forest Springs" Diversity 15, no. 2: 153. https://doi.org/10.3390/d15020153
APA StyleJekatierynczuk-Rudczyk, E., & Ejsmont-Karabin, J. (2023). Rotifers of Inter-Forest Springs. Diversity, 15(2), 153. https://doi.org/10.3390/d15020153