3.2. An Annotated Checklist of Species of the Genus Paralycus
1. Paralycus chongqingensis Fan, Li et Xuan, 1996
Paralycus chongqingensis Fan et al. 1996: 174, Figures 1 and 2; Smelansky 2003: 181; Xu et al. 2020: 486; Subías 2022: 29.
Type depository. Holotype (female) and 16 paratypes (8 females, 1 larva, 1 protonymph, 6 deutonymph) are in the Department of Plant Protection, College of Plant Protection, Southwest University, Chongqing, China (confirmed by the curator in this institution).
Known instars. Female, larva (not described), protonymph and deutonymph [
14].
Distribution. China: Chongqing (type locality) [
14].
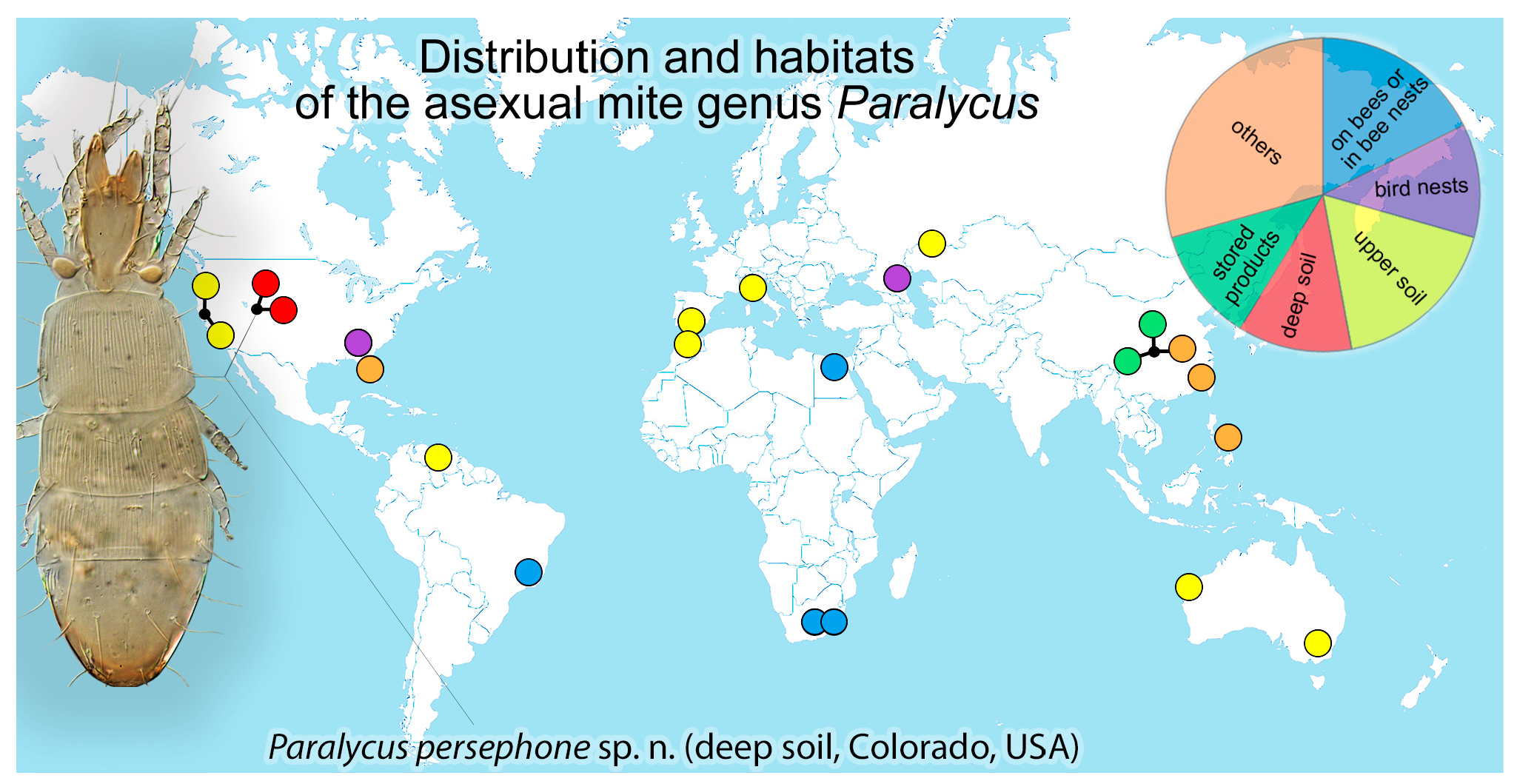
Habitat. Stored products, such as walnut, star anise, chili, garlic, beans, dried kelp (
Figure 1) [
14].
2. Paralycus daeira sp. n. (see below)
3. Paralycus lavoipierrei (Price, 1973)
Pediculochelus lavoipierrei Price 1973: 305, Figures 6–12.
Paralycus lavoipierrei: Norton et al. 1983: 493, Figures 3–11; Marshall et al. 1987: 28; Lebedeva and Poltavskaya 2013: 107; Xu et al. 2020: 486; Subías and Shtanchaeva 2021: 72; Subías 2022: 29.
Paralycus cf. lavoipierrei: Smelansky 2003: 181
Type depository. Holotype (female) and 4 paratypes (females) are in the U. S. National Museum; 15 paratypes (13 females, 1 larva, 1 deutonymph) are in the Entomology Museum, University of California, Berkeley, USA.
Known instars. Female, larva and deutonymph [
4].
Distribution. USA: California [
3,
4,
23] (type locality); Australia: Western Australia, Cape Range [
3]; Russia: Orenburg Oblast [
2], Stavropol’sky Kray [
24]; Venezuela [
25].
Habitat. Grassland soil [
4], carbonate soil [
2], nest of rosy starling (Figure 1) [
24].
4. Paralycus longior Fan, Li et Xuan, 1996
Paralycus longior Fan et al. 1996: 175, Figures 3 and 4; Smelansky 2003: 181; Xu et al. 2020: 486; Subías 2022: 29.
Type depository. Holotype (female) and 7 paratypes (6 females, 1 larva) are in the Department of Plant Protection, College of Plant Protection, Southwest University, Chongqing, China (formerly the Department of Plant Protection, Southwest Agricultural University) (not found by the curator in this institution).
Known instars. Female and larva [
14].
Distribution. China: Chongqing (type locality) [
14].
Habitat. Stored products, such as Auricularia fungi, lily bulbs, and tangerine cake (Figure 1) [
14].
5. Paralycus nortoni Xu, Zhu, Wu et Zhang, 2020
Paralycus nortoni Xu et al. 2020: 482, Figures 1–3; Subías 2022: 29.
Type depository. Holotype (female) and paratype (females) are in the National Zoological Museum of China, Institute of Zoology, Chinese Academy of Sciences, Beijing and Department of Plant Protection, Fujian Agriculture and Forestry University, China.
Known instars. Female [
13].
Distribution. China: Fujian Province (type locality) [
13].
Habitat. Under bark of
Pinus massoniana infested by longhorn beetles
Monochamus alternatus (Figure 1) [
13].
6. Paralycus parasiti Zhang et Li, 2001
Paralycus parasiti Zhang and Li 2001: 317, Figures 1 and 2; Xu et al. 2020: 486; Subías 2022: 29.
Type depository. Holotype (female) is in the Department of Plant Protection, Southwest Agricultural University, China.
Known instars. Female [
12].
Distribution. China: Chongqing (type locality) [
12].
Habitat. From
Coccinella septempunctata (Figure 1) [
12].
7. Paralycus parvulus (Price, 1973)
Pediculochelus parvulus Price 1973: 306, Figures 13–15.
Paralycus parvulus: Norton et al. 1983: 493; Marshall et al. 1987: 28; Smelansky 2003: 181; Xu et al. 2020: 486; Subías 2022: 29.
Type depository. Holotype (female) are in U. S. National Museum; 2 paratype (2 females) are in the Entomology Museum, University of California, Berkeley, USA.
Known instars. Female [
4].
Distribution. USA: California (type locality) [
4,
23].
Habitat. Grassland soil (Figure 1) [
4,
23].
8. Paralycus persephone sp. n. (see below)
9. Paralycus pricei sp. n. (see below)
10. Paralycus pyrigerus (Berlese, 1905) (see below)
11. Paralycus raulti (Lavoipierre, 1946) (see below)
Unidentified specimens
One specimen is from the nest of a pigeon in Atlanta, USA [
4]; several specimens were from Australia [
6] and the USA [
15]. “Paralycus raulti” was reported from Florida (USA) and Samar (Philippines) on rats and chickens [
11]. Baker and Wharton (1952, Figure 254) from [
11] illustrated a single specimen presumably from one of these locations. Based on this figure, their specimen differs from both Price’s specimens identified by him as
P. raulti by the long rostral setae (as in
P. raulti but not
P. pricei) and short cheliceral seta
cha (as in
P. pricei, but
P. raulti). A reexamination is needed to accurately identify Baker and Wharton’s specimens. The one species is a
Paralycus sp. Brazil: Minas Gerais, Sabará, managed nest of
Melipona marginata (Hymenoptera: Apidae) 19°54′50.1″ S 43°49′35.4″ W BMOC 15-0104-030 (this mite species is similar to
P. longior, but cannot be described without studying the types of
P. longior first).
3.3. Descriptions of Species
Paralycus daeira sp. n.
Paralycus sp. Pepato and Klimov, 2015: 8 (included in a molecular phylogeny); Klimov et al., 2017: 109 (included in a molecular phylogeny).
Diagnosis (female). The rostral setae are not reaching half the length of the chelicera; the lamellar setae are situated close to each other. The cheliceral setae cha are shorter than half the length of the chelicera. Gastronotal setae c, d, e, and f are not reaching bases of the next row of setae; h1 reached the bases of p1; p2 is shorter than p1. The epimeral setae 4a is absent. Three pairs of genital setae are present; the distance is g1 − g2 > g2 − g3. The leg trochanteral formula is 0-0-0-0; the genua I had 4 setae (d, l′, l″, v) while the genua II had 2 setae (l′ and l″); the solenidion ω of tarsi I had not expanded in the middle; and the solenidion φ of tibiae III is long (about 1/4 of the length of d).
Description. Female.Measurements. Idiosomal length 219, width 56.
Integument. The body is colorless. The prodorsal shield (except its posterior part) is smooth. The legs, chelicerae, and coxae are smooth. The dorsum (except segment P), ventrum, and ovipositor are striated.
Gnathosoma. The subcapitulum (30 × 22) had 3 pairs of filiform, smooth setae (a 9; m 4; h 4) and 2 pairs of filiform, and smooth adoral setae (4–5). There are 20 palps, and the setal formula is 0-1-0-1-9+ω. Of the setae, 3 setae (sul, acm, ul′) are with expanded tips. The inf is absent, and the postpalpal setae (ep 2) is blunt. The chelicerae is large (24) with 2 filiform and smooth setae (cha 5; chb 4); the cha is shorter than half of the length of the chelicera. The pharyngeal cupola is long, reaching the level of exp.
The prodorsum is covered with a shield-shaped plate in mid-dorsal region. The plate had 2 pairs of setae (ro 7; le 19). The setae ro does not reach half of the length of the chelicera. The bases of setae le are close to each other; the setae in (33) and exa (7) filiform are smooth, the exp is very short (2) and simple, and the bothridial setae (bs 12 × 7) is clavate and smooth.
Gastronotum. Segment C had 4 pairs of setae: c1 (6), c2 (11), c3 (9), and cp (19). Segment DE had 4 pairs of setae: d1 (7), d2 (10), e1 (6), and e2 (10). Segment F had 2 pairs of setae: f1 (20) and f2 (18). Segments H and P fused with 6 pairs of setae: h1 (24), h2 (25), h3 (10), p1 (30), p2 (28), and p3 (5). All gastronotal setae filiform are smooth; the setae f, h, p1, and p2 expanded at the bases; the c1 does not reach the bases of the cp; the d1 does not reach the bases of e; the e1 does not reach the bases of f; the f1 does not reach the bases of h1; the h1 reached the bases of p1; and the p2 is shorter than p1.
Epimeral and podosomal regions. The setal formula of the epimera is 3-2-3-2; the setae 1a (2), 1b (5), 1c (3), 2a (27), 2b (8), 3a (3), 3b (4), 3c (2), 4b (2), and 4c (3) filiform are smooth; the 2a reached the bases of 1a; the bases of 3a are situated close to each other; the 4a is absent. The supracoxal setae (3) is triangular with a rounded tip.
Anogenital region. There are 3 pairs of genital setae: g1-2 (4) and g3 (7). The distance g1 − g2 > g2 − g3. All genital setae are situated medially; the eugenital setae are minute (2). There are 3 pairs of adanal setae: ad1 (14), ad2 (30), and ad3 (7) and 2 pairs of anal setae: an1 (6) and an2 (3). All anogenital setae filiform are smooth. The genital tracheae are reduced and represented by short cavities.
Legs. The leg chaeto- and solenidiotaxy is reported: I 0-2-4-2-9 (0-1-1), II 0-2-2-3-6 (0-0-1), III 0-2-0-2-5 (0-1-0), and IV 0-2-0-2-5 (0-0-0). The famulus of tarsi I baculiform is thin and expanded at end; other setae filiform are smooth. The solenidion ω of tarsi I 6 × 1 is not expanded in the middle; the ω of tarsi II 3 × 1 is smaller and not expanded in middle; the φ of tibiae I elongate is attenuate; and the φ of tibiae III 5 is not shorter than half the length of tibia III and baculiform. The length of the tibial seta is
d 19 (
Figure 2,
Figure 3 and
Figure 4 and
Table 1).
Male. Unknown.
Juvenile instars.Measurements. The body length of the larva is 144 (excluding gnathosoma), protonymph is 160, deutonymph is 200, tritonymph is 214; body width of larva is 45, protonymph is 53, deutonymph is 55, and tritonymph is 56.
Integument is similar to the female but with segment P transversely striated dorsally.
Gnathosoma is similar to the female except the measurements. Subcapitulum is as follows: L 24 × 18, PN 23 × 19, DN 27 × 21, TN 29 × 23. The length of setae h is: L 3, PN and DN 4, TN 5; a L and PN 6, DN and TN 7; m L and PN 2, DN and TN 3. The adoral setae is: L and PN 3, DN and TN 4–5. The palps length is: L and PN 15, DN 17, TN 20. The chelicerae length is: L 18, PN 20, DN 22, TN 23. The cheliceral setae cha is: L and PN 2, DN 3, TN 4; chb L and PN 2, DN and TN 3.
Prodorsum is similar to the female except the bases of le–le are well separated (L, PN) or close (DN, TN). The length of the setae are: ro L 4, PN 5, DN 6, TN 7; le L 14, PN 14, DN and TN 16; in L 23, PN 24, DN and TN 30; exa L 5, PN and DN 6, TN 7; exp L 1, PN 2, DN and TN 3; bs L 13 × 6, PN 14 × 6, DN and TN 14 × 7.
Gastronotum is similar to the female, but transverse furrows are weakly developed in L, PN, and DN. The tf3 is absent in L; the gastronotal setae p is short and situated around the anal opening in L and the posterior anal opening (except p3) in other stages. The length of the setae are: c1 5; c2 L and PN 7, DN and TN 9; c3 L 6, PN and DN 9, TN 12; cp L and PN 13, DN 18, TN 19; d1 L 3, PN, DN and TN 4; d2 L 7, PN and DN 8, TN 9; e1 L 6, PN, DN and TN 7; e2 L 8, PN and DN 9, TN 11; f1 18–20; f2 L and PN 18, DN and TN 20; h1 L 15, PN 17, DN and TN 20; h2 L 18, PN 19, DN and TN 24; h3 L 3, PN 6, DN 7, TN 12; p1 L 2, PN 9, DN 16, TN 25; p2 L 2, PN 17, DN 25, TN 27; and p3 L 2, PN and DN 5, TN 6.
For the epimeral and podosomal regions, the claparède’s organs are absent in L and other instars. The chaetotaxy of the epimera is: L 3-1-2-0; PN 3-2-3-1; DN 3-2-3-2; TN 3-2-3-2. All setae filiform are smooth. The length of the setae is: 1a 2; 1b L 3, PN, DN, and TN 5; 1c L 2, PN, DN, and TN 3; 2a L, PN, and DN 20, TN 25; 2b PN 4, DN, and TN 6; 3a L and PN 2, DN and TN 3; 3b 4; 3c PN, DN, and TN 2; 4b 2; 4c 3. The supracoxal setae L is 1, PN, DN, and TN 2.
The anogenital region showed larva without genital, anal, and adanal setae. The protonymph had 1 pair of genital (3) and 3 pairs of adanal (3) setae; the anal setae are absent. The deutonymph had 2 pairs of genital (4) and 3 pairs of adanal (ad1 7, ad2 23, ad3 5) setae, and the anal setae an2 (1) are simple while the an1 are represented by alveoli. The tritonymph had 3 pairs of genital (3–5), 3 pairs of adanal (ad1 11, ad2 28, ad3 7), and 2 pairs of anal (6) setae; the eugenital setae are absent. All setae in all juvenile instars filiform (except an1 is alveoli) are smooth. The eugenital setae and other genital structures are absent.
The
legs are similar to the female, but the legs IV are absent in L. The leg setae and solenidia are as follows: L I 0-2-3-2-9 (0-1-1), II 0-2-2-3-6 (0-0-1), III 0-2-0-2-5 (0-1-0); PN I 0-2-3-2-9 (0-1-1), II 0-2-2-3-6 (0-0-1), III 0-2-0-2-5 (0-1-0), IV 0-0-0-0-5 (0-0-0); DN and TN I 0-2-4-2-9 (0-1-1), II 0-2-2-3-6 (0-0-1), III 0-2-0-2-5 (0-1-0), IV 0-2-0-2-5 (0-0-0). (
Figure 5,
Figure 6,
Figure 7,
Figure 8,
Figure 9,
Figure 10,
Figure 11 and
Figure 12 and
Table 1).
Type material. The holotype (female) and paratypes (1 larva, 1 protonymph, 2 deutonymphs, 1 tritonymph, 1 female) came from the USA: Colorado, Weld Co., 2 mi NE Masters, 40°20′02.4″ N 104°13′49.13″ W, 1371m, sandy soil, 90 cm depth, kerosene flotation, 12 Oct 2008, B.M. OConnor & J. Wilke, BMOC 08-1012-006. They are deposited at UMMZ.
DNA voucher. AD1370 BMOC 08-1012-006 is used with the same data with the following GenBank sequence accession ids: KY922457 (Cytochrome oxidase subunit I (COXI) gene); KY922705 (Heat shock protein cognate 5 (Hsc70-5) gene); KY922585 (Signal recognition particle protein 54k (Srp54k) gene); KY922330 (Elongation factor 1-alpha (Ef1alpha100E) gene); KP325061 (Small subunit ribosomal RNA (18S) gene); KY922209 (Small subunit ribosomal RNA (18S) gene); KP325023 (large subunit ribosomal RNA (28S) gene); KY922080 (large subunit ribosomal RNA (28S) gene).
Type deposition. UMMZ.
Additional material. We used numerous specimens with the same data, preserved for DNA work as the frozen tissue samples at UMMZ (accession BMOC 08-1012-006).
Etymology. Daeira is an underworld nymph and companion of the goddess Persephone (Greek mythology).
Remark 2. Females of Paralycus daeira are similar to those of P. lavoipierrei by the following character states: the rostral and cheliceral setae do not reach half of the length of chelicera; the bases of setae le are adjacent; epimeral setae 2a are long; gastronotal setae c, d, e, and f are not protruding beyond the bases of setae in the subsequent rows; setae f, h, p1, and p2 are slightly widened at their bases; there are 3 pairs of genital setae; solenidion ω of tarsus I is not expanded. However, P. daeira differs from P. lavoipierrei by the absence of epimeral setae 4a and seta v′ on leg trochanters III (present in P. lavoipierrei); solenidion φ on leg tibiae III is longer than half the length of tibia III (distinctly shorter in P. lavoipierrei); the absence of palpal seta inf and presence of setae cm, lt″ and vt″ (inf present; cm, lt″ and vt″ are absent in P. lavoipierrei); the striated posterior part of the propodosomal shield (vs. with only one medial line is present in P. lavoipierrei). Furthermore, P. daeira is similar to P. chongqingensis by the cheliceral setae cha not reaching half the length of the chelicera, gastronotal setae c1, d1 and e not protruding beyond the bases of setae in the subsequent rows, and by the presence of three pairs of genital setae; but differs from P. chongqingensis by the absence of epimeral setae 4a, setae v′ on leg trochanters III and setae v on leg genua II (all these setae are present in P. chongqingensis), and by the adjacent bases of the lamellar setae le (slightly distant in P. chongqingensis).
The larvae and deutonymphs of P. daeira and P. laviopierrei are very similar. They differ by solenidion φ of tibiae III longer than half the length of tibia III in P. daeira (distinctly shorter in P. laviopierrei); in addition, deutonymphs of these two species can be distinguished by the absence of leg seta v′ on trochanters III in P. daeira (present in P. laviopierrei).
Paralycus persephone sp. n.
Diagnosis (female). The rostral setae are long, reaching half the length of chelicerae; the bases of the lamellar setae are separate. The cheliceral setae cha are shorter than half the length of chelicerae. The gastronotal setae are not expanded at the bases; setae c1, d1, and e1 not reaching bases of next row of setae; setae f1 reached the bases of h1; setae h1 reached bases of p1; p2 are shorter than other dorsal gastronotal setae. The epimeral setae 4a are absent. There are 3 pairs of genital setae present, distance g1 − g2 > g2 − g3. The leg trochanteral formula is 0-0-0-0; genua I had 3 setae (d and l), and genua II had 2 setae (l). The solenidion ω of tarsi I are large and expanded in the middle; solenidion φ of tibiae III are short.
Description. Female.Measurements. The body length is 191 (excluding gnathosoma); the width is 64.
Integument. The body is colorless. The shield of prodorsum had one short medial strip in the posterior part and legs, and the coxae I and II are smooth. The dorsum (except segment P and part of segment H), ventrum, chelicerae (basal part), and ovipositor are striated.
Gnathosoma had a subcapitulum 29 × 22 with 3 pairs of filiform, smooth setae (a 10; m 4; h 5), and 2 pairs of filiform and smooth adoral setae (3–5). It had 24 palps; the setal formula is 0-1-0-1-9+ω; 3 setae had expanded tips (sul, acm, ul′); inf is absent; and the postpalpal setae (ep 2) is blunt. The chelicerae are 21 with 2 filiform and smooth setae (cha 6; chb 5). The cha is shorter than half of the length of the chelicera. The pharyngeal cupola is long, reaching the level of the exa.
Prodorsum is covered with a shield-shaped plate in mid-dorsal region, bearing 2 pairs of filiform and smooth setae (ro 12; le 16); ro reached half of the cheliceral length. The bases of le are distant from each other; in (25) and exa (10) filiform is smooth; exp is very short (3) and simple; and bothridial setae (12 × 9) clavate is smooth.
Gastronotum. Segment C had 4 pairs of setae: c1 (10), c2 (13), c3 (12), and cp (26). Segment DE had 4 pairs of setae: d1 (8), d2 (13), e1 (20), and e2 (25); segment F had 2 pairs of setae: f1 (30) and f2 (26). Segments H and P fused with 6 pairs of setae: h1 (30), h2 (28), h3 (20), p1 (23), p2 (25), and p3 (10). All gastronotal setae filiform are smooth and not expanded at the base; c1 does not reach the base of cp; d1 does not reach the base of e; e1 does not reach the base of f; f1 reached the base of h1; h1 reached the base of p1, and p2 is shorter than other dorsal gastronotal setae.
For the epimeral and podosomal regions, the setal formula of epimera is 3-2-3-2; the setae 1a (4), 1b (6), 1c (5), 2a (8), 2b (6), 3a (4), 3b (6), 3c (3), 4b (3), and 4c (4) filiform are smooth; 2a does not reach the bases of 1a, the bases of 3a are situated close to each other; and 4a is absent. The supracoxal setae (2) are triangular with a rounded tip.
In the anogenital region, there are 3 pairs of genital setae (g1 4, g2 5, g3 6). The distance g1 − g2 > g2 − g3; g1 and g3 are medial, and g2 is slightly more lateral than g1 and g3. Eugenital setae are minute (2). There are 3 pairs of adanal setae: ad1 (14), ad2 (23), and ad3 (5), and 2 pairs of anal setae: an1 (6) and an2 (3). All anogenital setae filiform are smooth. Genital tracheae are reduced and represented by short cavities.
For the
legs, the chaeto- and solenidotaxy are: I 0-2-3-2-9 (0-1-1), II 0-2-2-3-6 (0-0-1), III 0-2-0-2-5 (0-1-0), and IV 0-2-0-2-5 (0-0-0). Famulus of tarsi I baculiform are thin and expanded at the end; other setae filiform are smooth. Solenidion ω of tarsi I are large (6 × 3) and distinctly widened in the middle; ω of tarsi II (3 × 1) are shorter and had a widened middle; φ of tibiae I are elongated and attenuated; φ of tibiae III 2 are short and expanded at the end. The length of tibial seta
d is 13 (
Figure 13,
Figure 14 and
Figure 15 and
Table 2).
Male. Unknown.
Type material. Holotype (female), 1 paratype female, 2 paratype deutonymphs, 1 paratype protonymph: USA: Colorado, Weld Co., 2 mi NE Masters, 40°20′02.4″ N 104°13′49.13″ W, 1371m, sandy soil, 90 cm depth, kerosene flotation, 12 Oct 2008, B.M. OConnor & J. Wilke, BMOC 08-1012-006. Deposited at UMMZ.
Type deposition. UMMZ.
Additional material. Numerous specimens, same data, preserved for DNA work as frozen tissue samples at UMMZ.
Etymology. Persephone is the goddess of the underworld (Greek mythology).
Remark 3.Paralycus persephone is similar to P. parvulus in having long gastronotal setae cp, f and h distinctly extending beyond the bases of setae in the subsequent rows, the rostral setae reaching half the length of the chelicera; the position of setae le (bases are distinctly separated from each other), the absence of epimeral seta 4a, the presence of 3 pairs of genital setae, and the absence of setae v′ on leg trochanters III. However, P. persephone differs from P. parvulus as follows: gastronotal setae c1, d, and e shorter, not reaching the bases of the following pair of setae (extending beyond the bases of the next row setae in P. parvulus); seta v on leg genua I are absent (present in P. parvulus); and the greatly widened solenidion ω on leg tarsi I (weakly widened in P. parvulus). P. pyrigerus also has long gastronotal setae f, h, and p, but P. persephone has shorter setae c2, d, and e1, not reaching the bases of the next pair of setae (extending beyond these bases in P. pyrigerus) and the long, distinctly widened solenidion ω on tarsi I, longer than half of the length of tarsi I (not widened and shorter than half of the length of tarsi I in P. pyrigerus).
Paralycus pricei sp. n.
Pediculochelus raulti Price 1973 (specimen no. 1): 302, Figures 3–5 (misidentification).
Diagnosis (females). The rostral setae does not reach half the length of the chelicera; the bases of lamellar setae are distinctly separated from each other. The subcapitulum has four pairs of setae (h, a, m1, m2). The cheliceral setae cha are shorter than half the length of chelicera. The gastronotal setae d, e, f, and h1 does not reach the bases of setae in the next row; p2 is the longest. The epimeral setae 4a are present. There are 4 pairs of genital setae (g1–4). The leg trochanteral formula is 0-0-1-0. The genu I has 3 setae (d and l); the genu II has 2 setae (l); and the solenidion ω of tarsus I is simple and not widened.
Description. Female. Idiosoma 180 x 58.
Integument. The body is colorless. The shield of prodorsum, legs, and coxae are smooth; the dorsal and ventral sides of the body are striated.
Gnathosoma has subcapitulum (28 × 22) with 4 pairs of filiform and smooth setae (a 11; m1 and m2 4; h 10) and 2 pairs of filiform, smooth adoral setae (4). The palps length is 24. The chelicerae are large (22) with 2 filiform and smooth setae (cha 5; chb 4); the cha is shorter than half the length of the chelicera.
The prodorsum has a shield-shaped plate in mid-dorsal region, bearing 2 pairs of setae (ro 6; le 28); ro does not reach half the length of the chelicera. The bases of le are adjacent; in (38), exa (16), and exp (2) filiform are smooth; bothridial setae (13 × 6) clavate are smooth.
For the Gastronotum, Segment C has 4 pairs of setae: c1 (13), c2 (13), c3 (25), and cp (25). Segment DE has 4 pairs of setae: d1 (10), d2 (12), e1 (12), and e2 (16). Segment F has 2 pairs of setae: f1 (20) and f2 (18). Segments H and P fused and bear 6 pairs of setae: h1 (14), h2 (16), h3 (7), p1 (14), p2 (30), and p3 (8). All setae filiform are smooth and not widened at the bases; c1 nearly reached the base of cp, d1 does not reach the bases of e1, d2 does not reach the bases of e2; e1 does not reach the bases of f; f1 does not reach the bases of h1; h1 does not reach the bases of p1; and p2 is longer than other dorsal gastronotal setae.
In the epimeral and podosomal regions, chaetotaxy of epimera are 3-2-3-3; setae 1a (7), 1b (8), 1c (6), 2a (11), 2b (11), 3a (5), 3b (12), 3c (8), 4a (6), 4b (5), and 4c (4) are filiform and smooth; 2a does not reach the bases of 1a, and the bases of 3a are adjacent.
In the anogenital region, there are 4 pairs of genital setae (g1 4, g2 4, g3 5, g4 9); the distance is g1 − g2 > g2 − g3; g1 and g2 are medial, and g3 and g4 are slightly more lateral than g1 and g2. There are 3 pairs of adanal setae: ad1 (12), ad2 (26), and ad3 (8); 2 pairs of anal setae: an1 (6) and an2 (5); and all anogenital setae are filiform and smooth.
For the legs, the genua I has 3, and the genua II has 2 setae; the trochanteral formula is 0-0-1-0. All setae are filiform and smooth; solenidion ω of tarsus I 5 × 1 is slightly expanded at the tip and situated in the basal part of tarsus.
Male. Unknown.
Type material. The holotype (female) (=paratype of
P. raulti designated by Price (1973) [
4] is specimen no. 1 from the personal collection of Dr. M. Lavoipierre) has the following data: South Africa, Durban, 1940 from bees
Amegilla fallax and
Apis mellifera adansonii (as
Anthophora fallax and
Apis mellifera adansonii).
Type deposition. It is probably deposited at the University of California, Berkeley [
26].
Etymology. The species is named after Dr. Douglas W. Price who suggested that specimen no. 1 from M. Lavoipierre’s collection could represent an undescribed species.
Remark 4. Price (1973) [
4] did not give information about the structure of the ovipositor, palp, and legs as the specimen was old, and these structures could not be studied in detail. The differences between
P. pricei (specimen no. 1) and
P. raulti (specimen no. 2) were given previously [
4]; see also the key to species below.
Paralycus pricei is similar to P. parasiti by the well separated bases of the lamellar setae; the dorsal gastronotal setae (c1, d, e, f, h) not reaching the bases of the setae in the next row; and the presence of epimeral setae 4a and 4 pairs of genital setae g1–g4. However, P. pricei differs from P. parasiti by the rostral setae ro not reaching half the length of the chelicerae (reaching in P. parasiti). The leg genua I has 3 setae: l, d (4 setae: v, l, d in P. parasiti); the leg genua II has 2 l setae, and setae v are absent (3 setae: v, l in P. parasiti).
Paralycus pyrigerus (Berlese, 1905)
Alycus pyrigerus Berlese 1905: 232; Berlese 1910: 13, Figure 18 from [
27].
Paralycus pyrigerus: Womersley 1944: 134; Price 1973: 302; Norton et al. 1983: 493; Ruiz et al. 1991: 59; Gil-Martín et al. 1992: 55; Smelansky 2003: 181; Xu et al. 2020: 486; Subías 2022: 29.
Diagnostic description (female). The rostral setae reaches half the length of chelicera. Gastronotal setae c1 do not reach the bases of cp setae, c2 reaches the bases of cp; setae cp, d, e, f, and h are distinctly protruding beyond the bases of setae in the subsequent rows. Tarsus I is more than 2 times longer than wide. Solenidion ω of tarsus I is slightly expanded in the middle, distinctly shorter than half length of tarsus I.
Male. Unknown.
Type material. Holotype (female): Italy: Monte Pisano, chestnut forest, soil/decaying wood.
Type deposition. Holotype: CREA Research Centre for Plant Protection and Certification, Florence, Italy.
Known instars. Female (Berlese 1905).
Distribution. Italy: near Pisa (type locality) [
5]; Spain: Andalusia [
8]; Morocco [
7].
Habitat. Soil (Figure 1) [
5,
7,
8].
Remark 5. This species is inadequately described [
5], making it difficult to identify
P. pyrigerus based on the original description. In 1965, Donald E. Johnston examined the single holotype specimen preserved in the Berlese collection at the Agricultural Station near Florence, Italy, and found that it is congeneric with Pediculochelus [
3], but he did not re-describe it. We examined this specimen as images provided by Dr. Sauro Simoni and Silvia Guidoni, and we report that it is in poor condition, thus preventing a detailed study (
Figure 16).
However, here, we report some diagnostic characters that could be observed.
Paralycus pyrigerus and
P. parvulus are similar species, both having long rostral setae reaching half the length of the chelicera and the dorsal notogastral setae distinctly protruding beyond the bases of setae in the subsequent rows. Based on the original figure of Berlese (1910) [
27] and our study of the photos of the type,
P. pyrigerus has setae
c1 not protruding beyond the bases of
cp, and
c1 are subequal to
ro (vs. setae
c1 reaching the level of bases of setae
cp and are longer than
ro in
P. parvulus). We were not able to study the entire ventral side of the type specimen (except setae
g1 and
g2, which were clearly seen; however, additional genital setae are probably present as well). Because the original description of
P. pyrigerus lacks important diagnostic detail, we suggest that all subsequent reports of this species in the literature need confirmation.
Paralycus raulti (Lavoipierre, 1946)
Pediculochelus raulti Lavoipierre 1946: 130, Figure 1 (part.); Price 1973 (specimen no. 2): 302, Figures 1–5; Abou Senna 1997: 667; Baker and Wharton 1952: 325, Figure 254 from [
11] (specimens need to be examined to confirm identification).
Paralycus raulti: Krantz 1970: 145; Norton et al. 1983: 493; Smelansky 2003: 181; Xu et al. 2020: 486; Subías 2022: 29
Diagnostic description (female). The rostral setae reaches half the length of chelicera; the bases of lamellar setae are separated from each other. The cheliceral setae
cha is longer than half the length of the chelicera. The gastronotal setae
d1,
e, and
f does not reach the bases of the setae in the next row. Setae
h1 reaches the bases of
p1;
p2 is the longest among the dorsal gastronotal setae. The epimeral setae
4a is present. There are 5 pairs of genital setae. The leg trochanteral formula is 0-0-1-0; the genual formula is 4-2-0-0 [
3]. Solenidion ω of tarsus I slightly expanded in the middle (after [
4,
9]).
Known instars. Female [
4,
9].
Distribution. South Africa: Durban (type locality) [
9]; Egypt [
10].
Habitat. It lives on adult bees Amegilla fallax and
Apis mellifera adansonii [
9] and in tentorial pits on the head of the honeybee
Apis mellifera (Figure 1) [
10].
Type material. Holotype female and 3 female paratype (including specimen no. 2 designated by [
4]) are from: South Africa, Durban, 1940 from bees
Amegilla fallax and
Apis mellifera adansonii (as
Anthophora fallax,
Apis adansonii).
Type depository. Holotype (female) and 2 paratypes (females) are in the National Museum of Natural History, Paris, France; 1 paratype (female, specimen 2) and 1 paratype female specimen 1 (identified here as
P. pricei) are probably at the University of California, Berkeley [
26].
Remark 6. Price (1973) [
4] studied two paratypes of
P. raulti from Lavoipierre’s personal collection and found that both paratypes differ in a number of character states. He suggested that two separate species were involved but did not formally propose a new species for one of them. Here, we propose a new species,
P. pricei, based on Price’s original description of specimen no. 1 [
4]. Unfortunately, Price (1973) [
4] did not study the type of specimens deposited in the National Museum of Natural History, France, which include the holotype. We contacted this institution but received no response. Here, we identify
P. raulti based on the specimen figured by Lavoipierre (1946) [
9] as it is likely to be the holotype.
Our study of
P. raulti was based on the original description and figures of [
3,
4,
9]. Price (1973) [
4] reported that the paratype studied by him was in poor condition, so the gnathosoma and legs could not be observed in detail. The original description [
9] shows that
P. raulti has long
cha, which is consistent with specimen no. 2 in [
4]. Price figured leg genua I with 3 setae, but Norton et al. (1983) [
3], who studied the same specimens reported 4 setae.
Paralycus raulti is very similar to
P. longior but cheliceral setae
cha are longer than half the length of the chelicera in
P. raulti (vs. shorter in
P. longior).






















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}