
Checklist of the Fishes of the Kundelungu National Park (Upper Congo Basin, DR Congo): Species Diversity and Endemicity of a Poorly Known Ichthyofauna

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
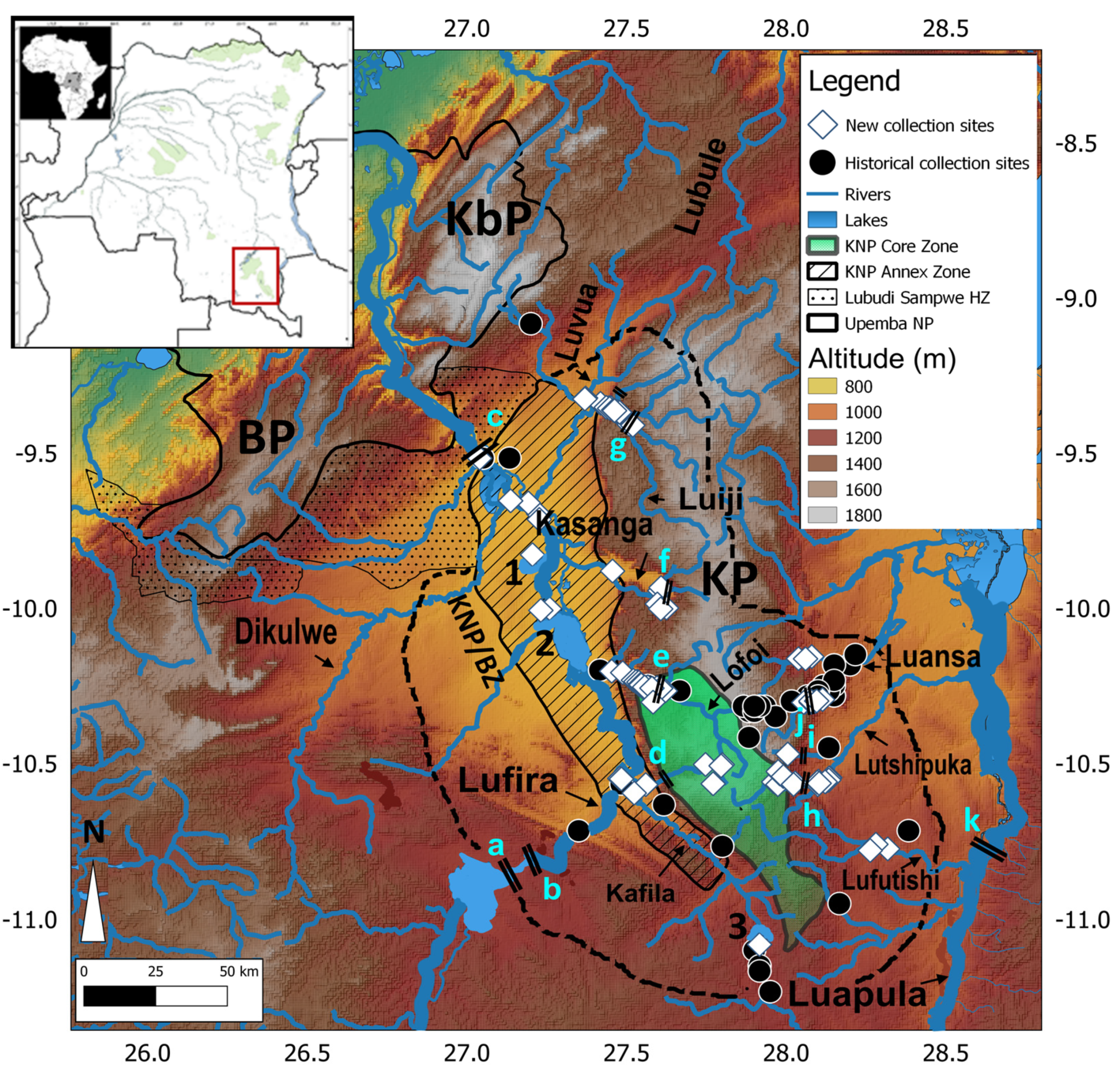
2.1. Hydrography of the Study Area
2.2. Data Collection and Analyses
3. Results
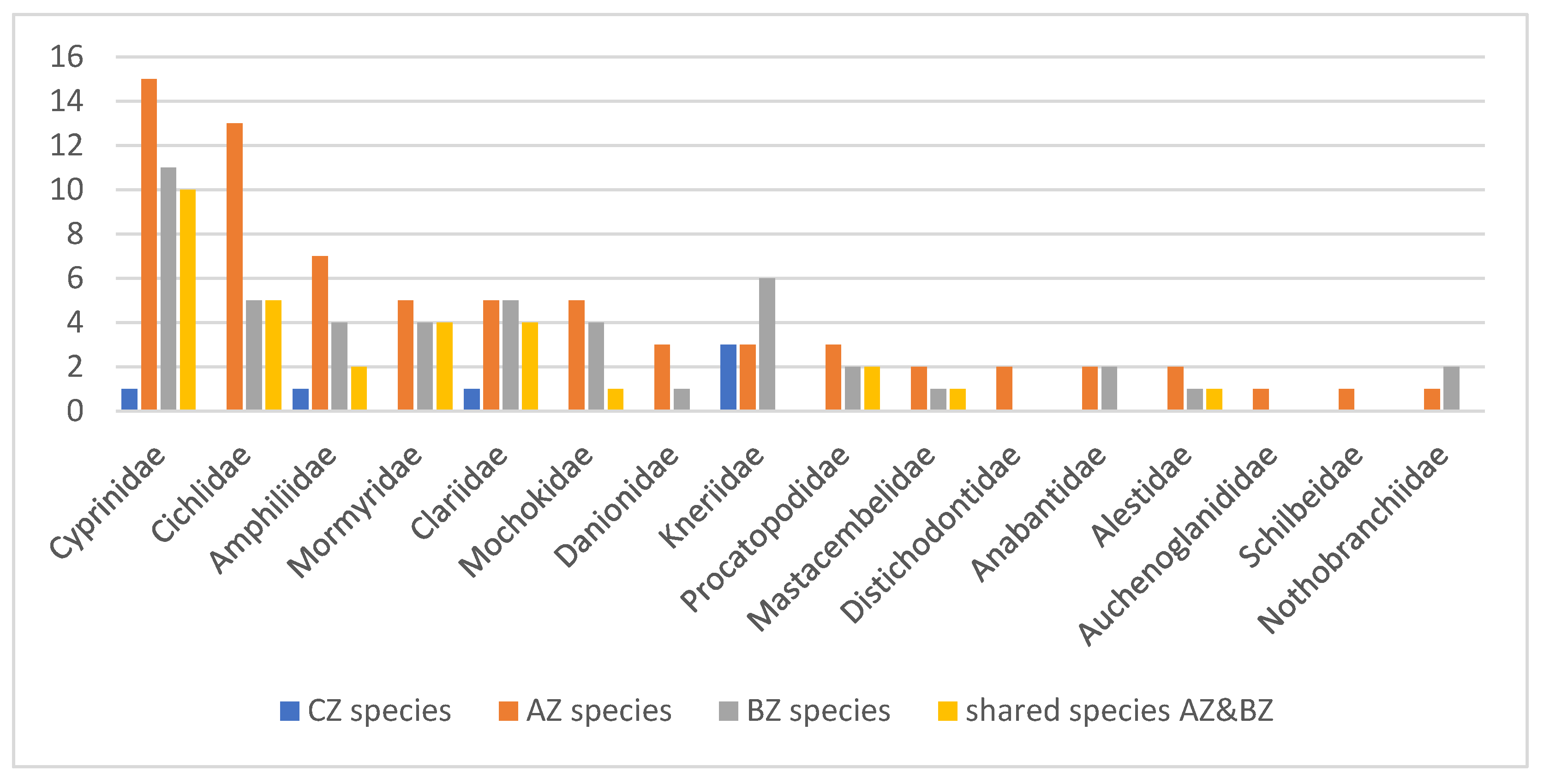
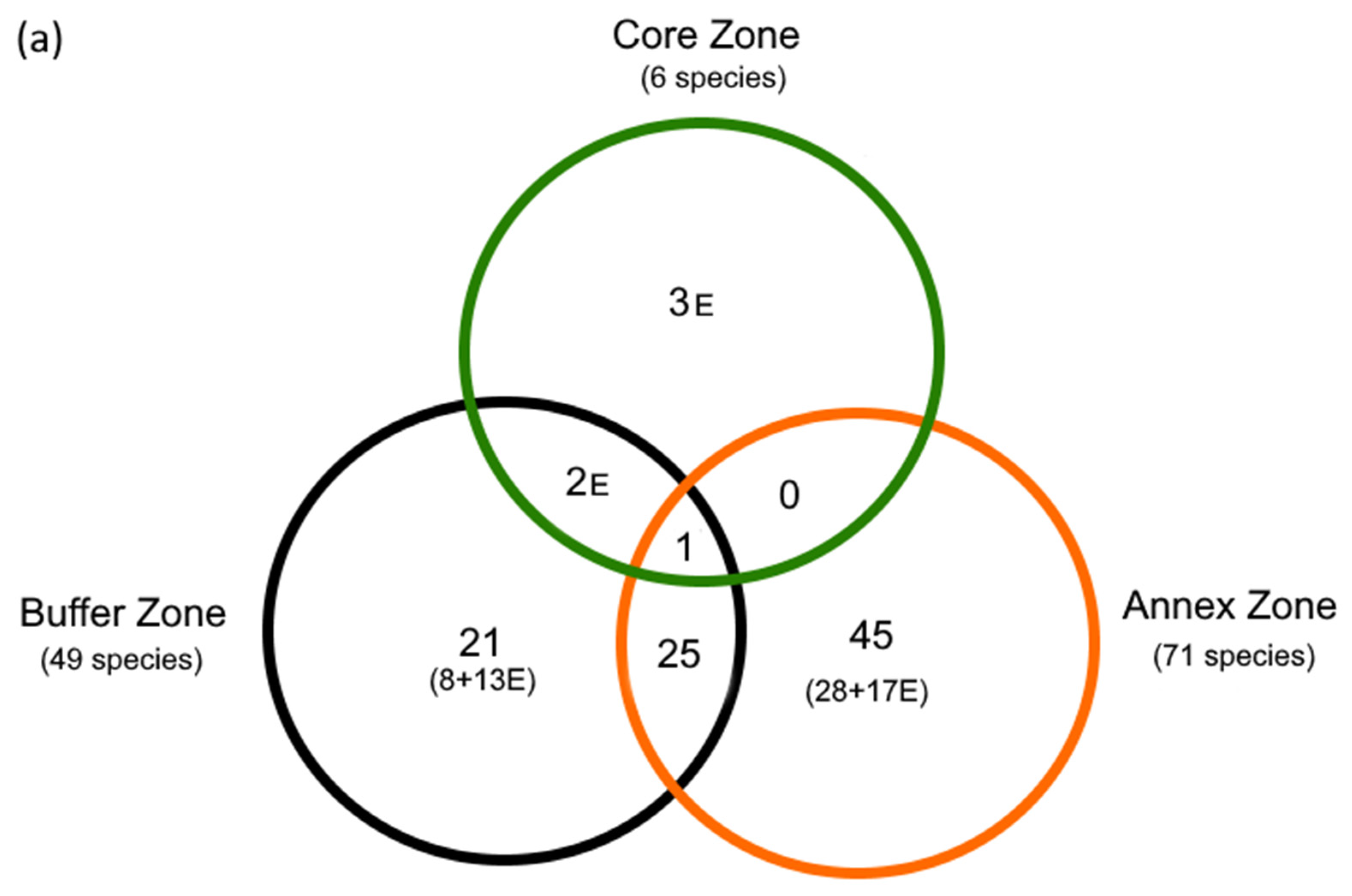
3.1. The Fish Diversity in the Kundelungu National Park (KNP) s.l.
3.2. Taxonomic Problems: Evidence for Hidden Species Diversity
3.2.1. Mormyridae
3.2.2. Kneriidae
3.2.3. Cyprinidae
3.2.4. Danionidae
3.2.5. Amphiliidae
3.2.6. Mochokidae
3.2.7. Auchenoglanididae
3.2.8. Procatopodidae
3.2.9. Cichlidae
4. Discussion
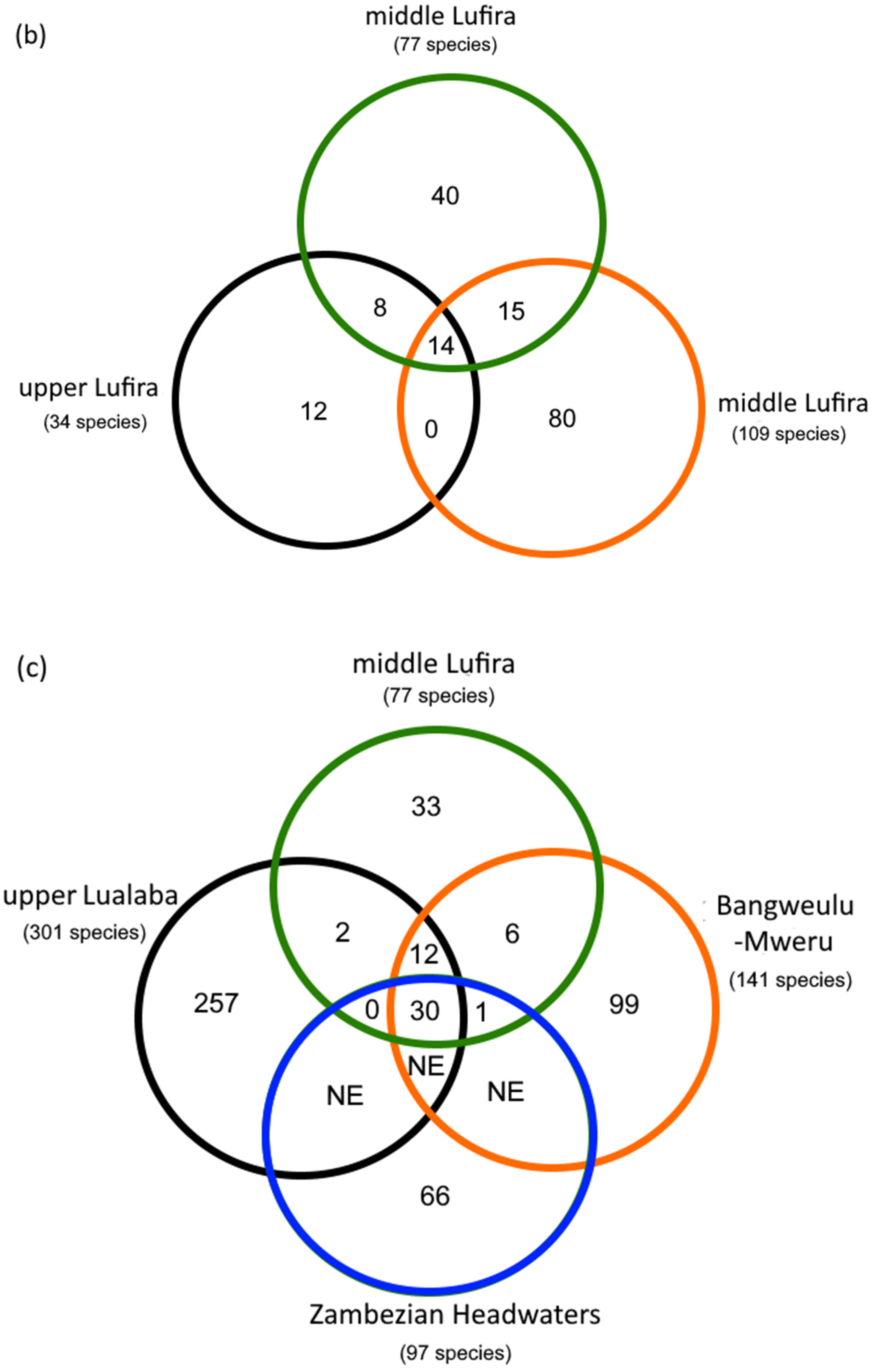
4.1. Contribution to Our Current Knowledge of the Fish Fauna Diversity of the Upper Lualaba and Bangweulu-Mweru Ecoregions
4.2. Ichthyobiogeography of the Rivers of the KNP s.l.
4.3. Protection/Conservation Issues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A









Appendix B

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Major Basins | Conservation Zones | IUCN | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species and Vernacular Names | Lufira Basin | BM | ZH n | Core Zone | Annex Zone | Buffer Zone | |||||||||
| lLf n | mLf | uLf n | Lfus | Ltus | mLfb | Kfl | Lfds | Ksds | Lv | mLf | BM | ||||
| Osteoglossiformes | |||||||||||||||
| Mormyridae (6) | |||||||||||||||
| Cyphomyrus discorhynchus (Peters, 1852) | + * | + * | LC | ||||||||||||
| Cyphomyrus lufirae Mulelenu et al. (2020), Lububu (S) | XE + * | E + | E + | E + | |||||||||||
| Marcusenius macrolepidotus (Peters, 1852), Lububu (S) | X + | + | X | X + | X | + | + | + | X + | LC | |||||
| Paramormyrops tavernei (Poll, 1972), Sengi (S) | XE | XE + | LC | ||||||||||||
| Petrocephalus catostoma (Günther, 1866), Bushimba drotea (B), Sengi (S) | + | + | + | + | + | LC | |||||||||
| Pollimyrus stappersii (Boulenger, 1915) | X | + | X + | + * | X + | LC | |||||||||
| Gonorynchiformes | |||||||||||||||
| Kneriidae (13) | |||||||||||||||
| Kneria stappersii Boulenger, 1915 | XE | + | LC | ||||||||||||
| Kneria sp. ‘katwei-ab’, Mujingiri, Muringiri, Mulingili (S), Mwinjo (K) | E + | E + p | E + p | ||||||||||||
| Kneria sp. ‘kasangaensis-ab’, Mujingiri, Muringiri (S) | E + | E + p | |||||||||||||
| Kneria sp. ‘luijiensis-ab’, Mujingiri, Muringiri, Mulingili (S), Mwinjo (K) | E + | E + p | |||||||||||||
| Kneria sp. ‘masansa-ab-uniform’, Munjinjingiri (B) | E + | E + p | |||||||||||||
| Kneria sp. ‘musipasi-ab’, Munjinjingiri (B) | E + | E + p | |||||||||||||
| Kneria sp. ‘luansa-ab’, Munjinjingiri (B) | E + | E + * p | |||||||||||||
| Kneria sp. ‘luansa-bl’, Munjinjingiri (B) | E + * | E + * | |||||||||||||
| Kneria sp. ‘seegersi-lofoi-mwena-bl’, Mujingiri, Muringiri (S), Mwinjo (K) | E + * | E + * | |||||||||||||
| Kneria sp. ‘restrictus-luiji-bl’, Mujingiri, Muringiri, Mulingili (S), Mwinjo (K) | E + | E + | |||||||||||||
| Kneria sp. ‘lutshipuka-bl’, Munjinjingiri (B) | E + | E + | |||||||||||||
| Parakneria malaissei Poll, 1969 complex, Kipala matete (B) | XE + | XE + | DD | ||||||||||||
| Parakneria thysi Poll, 1965 complex, Mujingiri, Muringiri, Kalumbu (S) | XE + | XE + | + | + | + | DD | |||||||||
| Cypriniformes | |||||||||||||||
| Cyprinidae (17) | |||||||||||||||
| Enteromius afrovernayi (Nichols & Boulton, 1927) | + * | X | X | + | + * | LC | |||||||||
| Enteromius eutaenia (Boulenger, 1904), Matule (B, S), Kipulumushi (S) | X | + * | X + | X | + | + | + | + | + | X + | LC | ||||
| Enteromius haasianus (David, 1936) | X + | X | X | X | X | LC | |||||||||
| Enteromius kerstenii (Peters, 1868) | + | X | X | + | + | + | + | LC | |||||||
| Enteromius lineomaculatus (Boulenger, 1903), Kasepa (S) | X | + * | X + | X + | X | + | + | + * | + * | + * | + * | LC | |||
| Enteromius multilineatus (Worthington, 1933) | X + | + * | X | X | + | + * | LC | ||||||||
| Enteromius neefi (Greenwood, 1962), Kasepa (B) | X + | + | + | + | + * | LC | |||||||||
| Enteromius paludinosus (Peters, 1852), Mulumbwe (S), Lubwe (B) Kanfisha (B) | X + | + * | X + | X + | X | + * | + | + * | + | + * | X + | LC | |||
| Enteromius radiatus (Peters, 1853) | + * | X + | X | + | + * | + | + | LC | |||||||
| Enteromius trimaculatus (Peters, 1852), Kyaka (B,S) Malandala (B), Sese, Shamatonga (S) | X + | + * | X + | + * | + | + * | + * | X + | LC | ||||||
| Enteromius unitaeniatus (Günther, 1867), Pulumuji, Lwebe (S) | + * | X | X | + | + | + | + * | LC | |||||||
| Enteromius sp. ‘kundelungu’, Kalumbulwa (B), Mulumbwe, Kipulumushi (S) | XE + | XE + | E + p | E + p | XE + | ||||||||||
| Enteromius sp. ‘luiji’, Mulumbwe, Kipulumushi (S) | E + | E + | + p & p’ | ||||||||||||
| Enteromius sp. ‘lutshipuka-bl’, Kalumbulwa (B) | E + | E + | |||||||||||||
| Labeo cylindricus Peters, 1852, Kalongwe (B, S) | X + | + * | X | X + | X | + * | + | + | + | X + | LC | ||||
| Labeo cf. rosae Steindachner, 1894 | X * | X * | * | LC | |||||||||||
| Labeobarbus trachypterus (Boulenger, 1915) complex, Nkeni (K,S), Kaleya (B), Kachila (S) | X | X + | X + | + | X + | + | + | X + | LC | ||||||
| Danionidae (4) | |||||||||||||||
| Chelaethiops congicus (Nichols & Griscom, 1917), Mulela hulu (S) | X + | X + | X + | + | LC | ||||||||||
| Engraulicypris brevianalis (Boulenger, 1908), Kisense (S) | + | X | + | + | LC | ||||||||||
| Neobola moeruensis (Boulenger, 1915) | XE + | + | LC | ||||||||||||
| Opsaridium zambezense (Peeters, 1852), Mukelekele (S) | X + | X | X | + | + | + | LC | ||||||||
| Characiformes | |||||||||||||||
| Distichodontidae (2) | |||||||||||||||
| Distichodus maculatus Boulenger, 1898, Kikwanya (S) | X | X + | X | X | + | LC | |||||||||
| Nannocharax dageti Jerep, Vari & Vreven, 2014 | + | X | X | + | NE | ||||||||||
| Alestidae (3) | |||||||||||||||
| Brycinus lateralis (Boulenger, 1900), Mbelebembe (K) Pambia (S) | X + | X + | X + | X | X | X + * | + | + | LC | ||||||
| Brycinus peringueyi (Boulenger, 1923), Lutuntu (B) | XE + | + * | LC | ||||||||||||
| Micralestes humilis Boulenger, 1899, Mukelekele (B), Mbelebembe, Lupambia (S) | X | + | + | + | + | + | + | + | LC | ||||||
| Siluriformes | |||||||||||||||
| Amphiliidae (9) | |||||||||||||||
| Amphilius frieli Thomson & Page, 2015, Kilamba (S,K), Kilamba mabwe (B) | X + * | X + | X | X + * | + | + | X + | NE | |||||||
| Amphilius sp. ‘murielae-mansansa-ab/bl’, Kilamba-mabwe (B) | E + | E + p & p’ | E + p & p’ | ||||||||||||
| Amphilius sp. ‘luansaensis-ab’, Kilamba mabwe (B) | XE + | XE + p | |||||||||||||
| Amphilius sp. ‘vandewallei-kyubo-ab’, Kilamba (S,K) | E + | E + | |||||||||||||
| Amphilius sp. ‘luijiensis-ab/bl-2′, Kilamba (S) | E + | E + | E + p & p’ | ||||||||||||
| Amphilius sp. ‘elachymystax-luiji-bl-1′, Kilamba (S) | E + | E + | |||||||||||||
| Congoglanis sagitta Ferraris, Vari & Skelton, 2011, Kamimbi (K, S) | + | X | + | + | + | NE | |||||||||
| Zaireichthys brevis (Boulenger, 1915), Ngoyi wa Lulenda (K, S) Kanyensha (B) | X | + | X * | X + | + | + | + | X + | LC | ||||||
| Zaireichthys sp. ‘luiji’, Ngoyi wa Lulenda (K, S) Kanyensha (B) | E + | + | |||||||||||||
| Mochokidae (8) | |||||||||||||||
| Chiloglanis cf. elisabethianus (Boulenger, 1915), Kaminye (S), Kinimba (B) | + | + n | X + | VU | |||||||||||
| Chiloglanis micropogon Poll, 1952 | + | + | NE | ||||||||||||
| Chiloglanis microps Matthes, 1965 | XE | XE + | XE + | LC | |||||||||||
| Chiloglanis sp. ‘luansa-luisé’ | E + | XE + | |||||||||||||
| Chiloglanis sp. ‘lufira’ | E + | + | + | ||||||||||||
| Synodontis lufirae Poll, 1971, Bongwe (S), Nkonkonko (K), Kakonkola (K, S) | XE + | X | XE + | X + | + | + | + | VU | |||||||
| Synodontis nigromaculatus Boulenger, 1905, Bongwe (B) | X + | X | + | LC | |||||||||||
| Synodontis denticulatus Ilunga et al., 2020, Bongwe (S), Nkonkonko (K), Kakonkola (K, S) | E + | E + * | + | + | NE | ||||||||||
| Auchenoglanididae (1) | |||||||||||||||
| Parauchenoglanis cf. punctatus (Boulenger, 1902), Sokosoko (S) | + | X | + | + | + | + | LC | ||||||||
| Clariidae (6) | |||||||||||||||
| Clarias buthupogon Sauvage, 1879, Mulonge (B) | X + | X + | LC | ||||||||||||
| Clarias dumerilii Steindachner, 1866, Kibobe (S, B) | X | + | + | + p & p’ | + | + | + | + | + | LC | |||||
| Clarias gariepinus (Burchell, 1822), Hondehonde (B), Mulombe (S), Kabambale (Sw) | X | + | X | X + | X | + | + | + | X + | LC | |||||
| Clarias ngamensis Castelnau, 1861, Mulombe, Mulombe kilundwe (S) | X | + | X | X | X | + | + | + | LC | ||||||
| Clarias stappersii Boulenger, 1915, Kineke (S, B) | X | + | X + | X | + | + | + | X + | NE | ||||||
| Clarias theodorae Weber, 1897, Mulemba, Musonfi (S) Mulonge (B) | X | + | X | X + | X | + | + | + | + | X + | LC | ||||
| Schilbeidae (1) | |||||||||||||||
| Schilbe intermedius Rüppell, 1832, Kandja (S) | X | X + | X | X | X | + | + | + | + | + | LC | ||||
| Cyprinodontiformes | |||||||||||||||
| Nothobranchiidae (3) | |||||||||||||||
| Nothobranchius hassoni Valdesalici & Wildekamp, 2004 | XE | XE | DD | ||||||||||||
| Nothobranchius malaissei Wildekamp, 1978 | XE + | XE | DD | ||||||||||||
| Nothobranchius chochamandai Nagy, 2014 | XE + | XE + | NE | ||||||||||||
| Procatopodidae (5) | |||||||||||||||
| Lacustricola moeruensis (Boulenger, 1914) | + | X + | + | + * | LC | ||||||||||
| Lacustricola cf. hutereaui (Boulenger, 1913) | + | X | X | + | + | LC | |||||||||
| Lacustricola cf. johnstoni (Günther, 1894), Tulumenso (K) Kisense, Kafunta (S) | + | + | X | + | + | LC | |||||||||
| Lacustricola sp. ‘lofoi’ | E + | E + | + | ||||||||||||
| Lacustricola sp. ‘luiji’ | E + | E + | |||||||||||||
| Poeciliidae (1) | |||||||||||||||
| Poecilia reticulata Peters, 1859 | I | I | |||||||||||||
| Synbranchiformes | |||||||||||||||
| Mastacembelidae (2) | |||||||||||||||
| Mastacembelus frenatus Boulenger, 1901, Muntongwa (B), Mwanzaloka (S) | + * | X | X + * | + | + | X + * | LC | ||||||||
| Mastacembelus. sp. ‘lufiraensis’ | X | X | X | ||||||||||||
| Cichliformes | |||||||||||||||
| Cichlidae (13) | |||||||||||||||
| Coptodon rendalli (Boulenger, 1897), Kituku (S, B), Kakenge (S) | X | + * | X | X + | X | + | + | + | + | + | LC | ||||
| Coptodon/Tilapia sp. ‘pungwe’ | E + | E + | |||||||||||||
| Oreochromis mweruensis Trewavas, 1983, Pale (S) | X + | X | X + | X + | + | + | + | LC | |||||||
| Oreochromis salinicola (Poll, 1948) | XE | XE | VU | ||||||||||||
| Orthochromis torrenticola (Thys van den Audenaerde, 1963), Lufindu (K, S) | X | XE + | XE + | + | + | + | LC | ||||||||
| Pseudocrenilabrus philander (Weber, 1897), Bututulu, Kandongorio (S), Lufindu (K, S) | X | + * | X | X + | X | + | + * | LC | |||||||
| Pseudocrenilabrus sp. ‘lufira’, Bututulu, Kandongorio (S), Lufindu (K, S) | E + | E + | + | + | + | + | |||||||||
| Sargochromis sp. ‘lufira’, Marichongo, Musase, Likobo (S) | E + | E + | + | + | + | ||||||||||
| Serranochromis robustus (Günther 1864), Makobo (S, B) | X + | X | X | + | + | CR | |||||||||
| Serranochromis macrocephalus (Boulenger, 1899), Makobo (S, B) | + * | X | X + | + | + | + | + | + * | LC | ||||||
| Serranochromis thumbergi (Castelnau, 1861), Makobo (S, B) | + * | X | X | X | + | + | LC | ||||||||
| Tilapia ruweti (Poll & Thys van den Audenaerde, 1965) | + | X | X + | X | + | + | LC | ||||||||
| Tilapia sparrmanii Smith, 1840, Kifinsa (S,B), Samba (B) | X | + * | X | X + | X | + | + | + | + | + | + * | LC | |||
| Anabantiformes | |||||||||||||||
| Anabantidae (2) | |||||||||||||||
| Ctenopoma multispine Peters, 1844, Nkomo (S) | X | + * | X | X | X | + | * | LC | |||||||
| Microctenopoma intermedium (Pellegrin, 1920) | + | X | X | + | * | LC | |||||||||
| Sub-total (KNP s.l. species) | 29 | 77 | 22 | 64 | 31 | 3 | 5 | 43 | 34 | 34 | 23 | 45 | 6 | 45 | |
| Total (all species of the sub-basin/zone) | 109 | 77 | 34 | 141 | 97 | 6 | 71 | 49 | |||||||
| (a) | mLf | uLf | lLf | uC | BM | ZH |
| N | 77 | 34 | 109 | 301 | 141 | 97 |
| Shared | 22 | 29 | 44 | 49 | 31 | |
| Shared (%) | 28.6 | 37.7 | 57.1 | 63.6 | 40.3 | |
| J | 0.25 | 0.18 | 0.13 | 0.29 | 0.22 | |
| (b) | CZ | AZ | BZ | |||
| N | 6 | 71 | 49 | |||
| Shared | 1 | 3 | ||||
| Shared (%) | 16.7 | 50.0 | ||||
| J | 0.013 | 0.058 | ||||
| (c) | AZ | BZ | CZ | |||
| N | 71 | 49 | 6 | |||
| Shared | 26 | 1 | ||||
| Shared (%) | 36.6 | 1.4 | ||||
| J | 0.277 | 0.013 |
References
- Vanleeuwe, H.; Henschel, P.; Pelissier, C.; Moyer, D.; Gotanegre, A. Recensement des Grands Mammifères et Impacts Humains. Parcs Nationaux de L’upemba et de Kundelungu, République Démocratique du Congo; Wildlife Conservation Society: Kinshasa, Democratic Republic of the Congo, 2008; p. 30. [Google Scholar]
- UICN/PACO. Parcs et réserves de la République Démocratique du Congo: Évaluation de L’efficacité de Gestion des Aires Protégées; BF: UICN/PACO:: Gland, Switzerland, 2010; p. 140. [Google Scholar]
- Cotterill, F.P.D. The Upemba lechwe, Kobus anselli: An antelope new to science emphasizes the conservation importance of Katanga, Democratic Republic of Congo. J. Zool. Soc. Lond 2005, 265, 113–132. [Google Scholar] [CrossRef]
- Malaisse, F. Ecologie de la rivière Luanza, Cercle Hydrobiologique de Bruxelles; Séries Exploration Hydrobiologique du lac Bangweolu et du Luapula: Brussels, Belgique, 1976; p. 151. [Google Scholar]
- Lisowski, S.; Malaisse, F. Groupement Végétaux des Mares et des Anses Calmes des Rivières du Plateau de Kundelungu; Cercle hydrobiologique de Bruxelles: Brussels, Belgique, 1989; p. 41. [Google Scholar]
- Scott, L. Bangweulu-Mweru. In Freshwater Ecoregions of Africa and Madagascar: A conservation assessment; Thieme, M.L., Abell, R., Stiassny, M.L.J., Skelton, P., Lehner, B., Teugels, G.G., Dinerstein, E., Kamdem-Toham, A., Burgess, N., Olson, D., Eds.; Island Press: Washington DC, USA, 2005; pp. 185–186. [Google Scholar]
- Brown, A.; Abell, R. Upper Lualaba. In Freshwater ecoregions of Africa and Madagascar: A conservation assessment; Thieme, M.L., Abell, R., Stiassny, M.L., Skelton, P., Lehner, B., Teugels, G.G., Dinerstein, E., Kamdem-Toham, A., Burgess, N., Olson, D., Eds.; Island Press: Washington DC, USA, 2005; pp. 203–205. [Google Scholar]
- Misser, F. Les aires protégées en République Démocratique du Congo: Menaces et défis, l’action de l’Union européenne. Rev. Trimest. De Conserv. De La Nat. Et De Gest. Durable D’ardenne Et Gaume 2013, 68, 1–51. [Google Scholar]
- Poll, M. Exploration du Parc National de l’Upemba, Poissons; Fondation pour favoriser les Recherches Scientifiques en Afrique, Bruxelles: Brussels, Belgique, 1976; p. 127. [Google Scholar]
- Banister, K.E.; Bailey, R.G. Fishes collected by the Zaïre River Expedition, 1974–75. Zool. J. Lin. Soc 1979, 66, 205–249. [Google Scholar] [CrossRef]
- Banister, K.E. Fish of the Zaïre River. In The Ecology of River Systems; Davies, B.R., Walker, K.F., Eds.; Springer: New York, NY, USA, 1986; pp. 215–224. [Google Scholar]
- Magis, N. La pêche dans le lacs de retenue de Koni et de N’zilo. (Haut Katanga); Université de Liège, F.U.L.R.E.A.C: Liège, Belgium, 1961; p. 52. [Google Scholar]
- Magis, N. Nouvelle contribution à l’étude hydrobiologique des lacs de Mwadingusha, Koni et N’zilo; Université de Liège F.U.L.R.E.A.C: Liège, Belgium, 1961; p. 71. [Google Scholar]
- Goorts, P.; Wilmet, J.; Magis, N. Les Aspects Biologiques, Humains et Économiques de la Pêche dans le lac de Barrage de la Lufira; Université de Liège, F.U.L.R.E.A.C: Liège, Belgium, 1961; p. 127. [Google Scholar]
- Poll, M. Poissons recueillis au Katanga par H. J. Bredo. Bul. Mus. R. Hist. Nat. Bel. 1948, 24, 1–24. [Google Scholar]
- Thys van den Audenaerde, D.F.E. Description d’une espèce nouvelle d’Haplochromis (Pisces, Cichlidae) avec observations sur les Haplochromis rhéophilus du Congo oriental. Rev. Zool. Bot. Afr 1963, 68, 140–151. [Google Scholar]
- Poll, M. Contribution à la faune ichthyologique du Katanga. Annal. Mus. Congo Bel. Zool. 1933, 3, 101–152. [Google Scholar]
- David, L.; Poll, M. Contribution à la faune ichthyologique du Congo Belge: Collections du Dr. H. Schouteden (1924–1926) et d’autres récolteurs. Ann. Mus. Congo Bel. Zool. 1937, 3, 189–294. [Google Scholar]
- De Kimpe, P. Contribution à l’étude hydrobiologique du Luapula-Moero. Ann. Mus. R. Afr. Cent. Sci. Zool. 1964, 8A, 128–238. [Google Scholar]
- Van Steenberge, M.; Vreven, E.; Snoeks, J. The fishes of the Upper Luapula area (Congo basin): A fauna of mixed origin. Ichth. Expl. Fresh 2014, 24, 329–345. [Google Scholar]
- Balon, E.K.; Stewart, D.J. Fish assemblages in a river with unusual gradient (Luongo, Africa-Zaire system), reflections on river zonation, and description of another new species. Env. Biol. Fish 1983, 9, 225–252. [Google Scholar] [CrossRef]
- Poll, M. Révision des Synodontis africains (famille Mochokidae). Ann. Mus. R. Afr. Cent. Sci. Zool 1971, 191, 1–497. [Google Scholar]
- Howes, G.J. A review of the anatomy, taxonomy, phylogeny and biogeography of the African neoboline cyprinid fishes. Bull. Brit. Mus. (Nat. Hist) Zool. 1984, 47, 151–185. [Google Scholar]
- Teugels, G.G. A systematic revision of the African species of the genus Clarias (Pisces; Clariidae). Ann. Mus. R. Afr. Cent. Sci. Zool 1986, 247, 1–199. [Google Scholar]
- Paugy, D. Révision Systématique des Alestes et Brycinus africains (Pisces, Characidae); Collection Études et Thèses; Éditions de l’O.R.S.T.O.M.: Paris, France, 1986; p. 295. [Google Scholar]
- De Vos, L. A systematic revision of the African Schilbeidae (Teleostei, Siluriformes), with an annotated bibliography. Ann. Mus. R. Afr. Centr. Sci Zool 1995, 271, 1–450. [Google Scholar]
- Tshibwabwa, S.M. Systématique des Espèces Africaines du Genre Labeo (Teleostei, Cyprinidae) Dans Les Régions Ichtyo Géographiques de Basse-Guinée et du Congo. II. Ph.D. Thesis, Université de Namur, Namur, Belgium, 1997. [Google Scholar]
- Vreven, E. A Systematic Revision of the African Spiny-Eels (Mastacembelidae; Synbranchiformes). Volume I–VII. Ph.D. Thesis, Katholieke Universiteit Leuven, Leuven, Belgium, 2001. [Google Scholar]
- Geerinckx, T.; Adriaens, D.; Teugels, G.G.; Verraes, W. A systematic revision of the African catfish genus Parauchenoglanis (Siluriformes: Claroteidae). J. Nat. Hist. 2004, 38, 775–803. [Google Scholar] [CrossRef]
- Thomson, A.W.; Page, L.M. Taxonomic revision of the Amphilius uranoscopus group (Teleostei: Siluriformes) in Kenya, with the description of a new species form the Athi River. Bull. Flor. Mus. Nat. Hist 2010, 49, 45–66. [Google Scholar]
- Moelants, T.; Mbadu Zebe, V.; Snoeks, J.; Vreven, E. A review of the Distichodus antonii assemblage (Characiformes: Distichodontidae) from the Congo basin. J. Nat. Hist 2014, 48, 1707–1735. [Google Scholar] [CrossRef]
- Thomson, A.W.; Page, L.M.; Hilber, S.A. Revision of the Amphilius jacksoni complex (Siluriformes: Amphiliidae), with the description of five new species. Zootaxa 2015, 3986, 061–087. [Google Scholar] [CrossRef] [PubMed]
- Daget, J.; Gosse, J.-P.; Thys van den Audenaerde, D.F.E. Check-List of the Freshwater Fishes of Africa (CLOFFA); Volume 1, RMCA: Tervuren, Belgium; Orstom: Paris, France, 1984; p. 429. [Google Scholar]
- Daget, J.; Gosse, J.-P.; Thys van den Audenaerde, D.F.E. Check-List of the Freshwater Fishes of Africa (CLOFFA); Volume 2, RMCA: Tervuren, Belgium; ISNB: Bruxelles, Belgium; Orstom: Paris, France, 1986; p. 535. [Google Scholar]
- Daget, J.; Gosse, J.-P.; Thys van den Audenaerde, D.F.E. Check-List of the Freshwater Fishes of Africa (CLOFFA); Volume 4, RMCA: Tervuren, Belgium; ISNB: Bruxelles, Belgium; Orstom: Paris, France, 1991; p. 740. [Google Scholar]
- Froese, R.; Pauly, D. FishBase. Available online: http://www.fishbase.org (accessed on 1 July 2022).
- Skelton, P.H. A Complete Guide to the Fresh Water Fishes of Southern Africa; Struik Publishers: Cape Town, South Africa, 2001; p. 395. [Google Scholar]
- Sigovini, M.; Keppel, E.; Tagliapietra, D. Open nomenclature in the biodiversity era. Meth. Ecol. Evol. 2016, 7, 1217–1225. [Google Scholar] [CrossRef]
- Fricke, R.; Eschmeyer, W.N.; Van der Laan, R. Catalog of Fishes: Genera, Species, References. Available online: http://researcharchive.calacademy.org/research/ichthyology/catalog/fishcatmain.asp (accessed on 5 July 2022).
- Nelson, J.S.; Grande, T.C.; Wilson, M.V.H. Fishes of the World, 5th ed.; John Wiley: Hoboken, NJ, USA, 2016; p. 746. [Google Scholar]
- Poll, M. Contribution à l’étude des Kneriidae et description d’un nouveau genre, le genre Parakneria (Pisces, Kneriidae). Mém. Acad. Roy. Belg. 1965, 36, 1–28. [Google Scholar]
- Poll, M. Contribution à la connaissance des Parakneria. Rev. Zool. Bot. Afr. 1969, 80, 359–368. [Google Scholar]
- Kiwele, P.; Abwe, E.; Schedel, F.; Chocha Manda, A.; Schliewen, U.; Vreven, E. The Parakneria Poll, 1965 (Gonorhynchiformes: Kneriidae) “Miringiri” in the Upper Lualaba and Luapula-Mweru basins (Upper Congo, DRC). In Proceedings of the Sixth International Conference of the Panafrican fish and fisheries (PAFFA), Mangochi, Malawi, 24–28 September 2018; African Fish and Fisheries: Diversity, Conservation and Sustainable Management, a Book of Abstract of a Panel of Experts Meeting; Mangochi, Malawi, 24–28 September 2018, pp. 35–36.
- Malaisse, F. Se Nourrir en Forêt Clair Africaine: Approche Écologique et Nutritionnelle; Les presses Agronomiques de Gembloux: Gembloux, Belgium, 1997; p. 383. [Google Scholar]
- da Costa, L.M. Systematic Studies on the Chubbyhead Barbs Species Complex (Osteichthyes, Cyprinidae) from Southern Africa. Ph.D. Thesis, University of Lisbon, Lisbon, Portugal, 2013. Available online: http://hdl.handle.net/10451/7854 (accessed on 22 December 2019).
- Lévêque, C.; Daget, J. Cyprinidae. In Check-list of the Freshwater Fishes of Africa (CLOFFA); Daget, J., Gosse, J.P., Thys van den Audenaerde, D.F.E., Eds.; ORSTOM: Paris, France; RMCA: Tervuren, Belgium, 1984; Volume 1, pp. 217–342. [Google Scholar]
- Hayes, M.M.; Armbruster, J.W. The taxonomy and relationships of the African small barbs (Cypriniformes: Cyprinidae). Copeia 2017, 105, 348–362. [Google Scholar] [CrossRef]
- Chocha Manda, A.; Abwe, E.; Bills, R.; da Costa Vreven, E. On some chubbyhead minnows of the Upper Lualaba (Upper Congo basin: DR Congo): The case of E. motebensis (Cypriniformes: Cyprinidae) and the populations of the Kundelungu highland plateau. In Proceedings of the Sixth International Conference of the Panafrican Fish and Fisheries (PAFFA), African Fish and Fisheries: Diversity, Conservation and Sustainable Management, a Book of Abstract of a Panel of Experts Meeting. Mangochi, Malawi, 24–28 September 2018; p. 55. [Google Scholar]
- Banister, K.E. A revision of the large Barbus (Pisces, Cyprinidae) of east and central Africa. Studies on African Cyprinidae Part II. Bull. Brit. Mus. (Nat. Hist) Zool. 1973, 26, 1–148. [Google Scholar]
- Vreven, E.J.W.M.N.; Musschoot, T.; Snoeks, J.; Schliewen, U.K. The African hexaploid Torini (Cypriniformes: Cyprinidae): Review of a tumultuous history. Zool. J. Linn. Soc. 2016, 177, 231–305. [Google Scholar]
- Vreven, E.J.; Musschoot, T.; Decru, E.; Lunkayilakio, S.W.; Obiero, K.; Cerwenka, A.F.; Schliewen, U.K. The complex origins of mouth polymorphism in the Labeobarbus (Cypriniformes: Cyprinidae) of the Inkisi River basin (Lower Congo, DRC, Africa): Insights from an integrative approach. Zool. J. Linn. Soc. 2018, 186, 414–482. [Google Scholar] [CrossRef]
- Reid, G.M. A revision of the African species of Labeo (Pisces: Cyprinidae) and a re-definition of the genus. Theses Zool. 1985, 6, 1–322. [Google Scholar]
- Van Steenberge, M.; Gajdzik, L.; Chilala, A.; Snoeks, J.; Vreven, E. Labeo rosae (Cypriniformes: Cyprinidae) in the Congo basin: A relict distribution or a historical introduction? J. Fish Biol. 2014, 85, 1733–1738. [Google Scholar] [CrossRef] [PubMed]
- Nichols, J.T.; Griscom, L. Fresh-water fishes of the Congo basin obtained by the American Museum Congo Expedition, 1909–1915. Bull. Amer. Mus. Nat. Hist. 1917, 37, 653–756. [Google Scholar]
- Banyankimbona, G.; Vreven, E.; Ntakimazi, G.; Snoeks, J. The riverine fishes of Burundi (East Central Africa): An annotated checklist. Ichthyol. Explor. Freshwaters 2012, 23, 273–288. [Google Scholar]
- Abwe, E. The Fish Fauna of the Kundelungu National Park (KNP/DR Congo): Diversity and Conservation. Ph.D. Thesis, Ku Leuven University, Leuven, Belgium, 2022. [Google Scholar]
- Thomson, W.A. Systematics of the African Catfish Family Amphiliidae (Teleostei: Siluriformes). Ph.D. Thesis, University of Florida, Gainesville, FL, USA, 2013. [Google Scholar]
- Roberts, T.R. Systematics and osteology of Leptoglaninae, a new subfamily of the African catfish family Amphiliidae, with descriptions of three new genera and six new species. Proc. Calif. Acad. Sci. 2003, 54, 81–132. [Google Scholar]
- Seegers, L. The Catfishes of Africa: A Handbook for Identification and Maintenance; Aqualog Verlag A.C.S. GmbH: Rodgau, Germany, 2008; p. 604. [Google Scholar]
- Gosse, J.-P. Mochokidae. In Check-List of the Freshwater Fishes of Africa (CLOFFA).; Daget, J., Gosse, J.P., Thys van den Audenaerde, D.F.E., Eds.; RMCA: Tervuren, Belgium, 1986; Volume 2, pp. 105–152. [Google Scholar]
- Boulenger, G.A. Poissons recueillis au Congo belge par l’expédition du Dr. C. Christy. Ann. Mus. Congo Bel. Sci. Zool. 1920, 2, 1–39. [Google Scholar]
- Poll, M. Recherches sur la faune ichthyologique de la région du Stanley-Pool. Ann. Mus. Congo Bel. Sci. Zool. 1959, 71, 75–174. [Google Scholar]
- Boulenger, G.A. Mission Stappers au Tanganika-Moero. Diagnoses de poissons nouveaux. II. Mormyrides, Kneriides, Characinides, Cyprinides, Silurides. Rev. Zool. Afr. 1915, 4, 162–171. [Google Scholar]
- Geerinckx, T.; Adriaens, D.; Teugels, G.G. Auchenoglanidinae. In The Fresh and Brackish Water Fishes of Lower Guinea, West-Central Africa; Stiassny, M.L.J., Teugels, G.G., Hopkins, C.D., Eds.; Collection Faune et Flore tropicales, 42; Institut de Recherche pour le Développement: Paris, France; Muséum National d’Histoire Naturelle: Paris, France; Musée Royal de l’Afrique Centrale: Tervuren, Belgium, 2007; Volume I, pp. 587–607. [Google Scholar]
- Boulenger, G.A. Additions à la faune ichthyologique de bassin du Congo. Matériaux pour la faune du Congo. Ann. Mus. Congo Bel. Sci. Zool. 1902, 2, 19–57. [Google Scholar]
- Bragança, P.H.N.; Costa, W.J.E.M. Multigene fossil-calibrated analysis of the African lampeyes (Cyprinodontoidei: Procatopodidae) reveals an early Oligocene origin and Neogene diversification driven by palaeogeographic and palaeoclimatic events. Org. Div. Evol. 2019, 19, 303–320. [Google Scholar] [CrossRef]
- Bragança, P.H.N.; van Zeeventer, R.; Bills, R.; Tweddle, D.; Chakona, A. Diversity of the southern Africa Lacustricola Myers, 1924 and redescription of Lacustricola johnstoni (Boulenger, 1906) and Lacustricola myaposae (Boulenger, 1908) (Cyprinodontiformes: Procatopodidae). Zookeys 2020, 923, 91–113. [Google Scholar] [CrossRef]
- Bragança, P.H.N.D.; Skelton, P.H.; Bills, R.; Tweddle, D.; Chakona, A. Revalidation and redescription of “Lacustricola” chobensis (Fowler, 1935) and description of a new miniature species of “Lacustricola” from Southern Africa (Cyprinodontiformes: Procatopodidae). Ichthyol. Herp. 2021, 109, 123–137. [Google Scholar]
- Huber, J.H. Updates to the phylogeny and systematics of the African lampeye schooling cyprinodonts (Cyprinodontiformes: Aplocheilichthyinae). Cybium 1999, 23, 53–77. [Google Scholar]
- Bell-Cross, G. A revision of certain Haplochromis species (Pisces: Cichlidae) of Central Africa. Occas. Pap. Nat. Mus. Rhod. Ser. B 1975, 5, 405–464. [Google Scholar]
- Poll, M. Contribution à la Faune Ichthyologique de l’Angola; çōes Culturais de la Companhia dos Diamentes de Angola (DIAMANG): Lisbon, Portugal, 1967; p. 381. [Google Scholar]
- Crispo, E.; Chapman, L.J. Geographic variation in phenotypic plasticity in response to dissolved oxygen in an African cichlid fish. J. Evol. biol. 2010, 23, 2091–2103. [Google Scholar] [CrossRef]
- Muschick, M.; Barluenga, M.; Salzburger, W.; Meyer, A. Adaptive phenotypic plasticity in the Midas cichlid fish pharyngeal jaw and its relevance in adaptive radiation. BMC Evol. Biol. 2011, 11, 1–12. [Google Scholar] [CrossRef]
- Theis, A.; Ronco, F.; Indermaur, A.; Salzburger, W.; Egger, B. Adaptive divergence between lake and stream populations of an East African cichlid fish. Mol. Ecol. 2014, 23, 5304–5322. [Google Scholar] [CrossRef] [PubMed]
- Neat, F.C.; Lengkeek, W.; Westerbeek, E.P.; Laarhoven, B.; Videler, J.J. Behavioural and morphological differences between lake and river populations of Salaria fluviatilis. J. Fish Biol. 2003, 63, 374–387. [Google Scholar] [CrossRef]
- Çakmak, E.; Alp, A. Morphological differences among the mesopotamian spiny eel, Mastacembelus mastacembelus (Banks & Solander 1794), populations. Turk. J. Fish. Aqua. Sci. 2010, 10, 87–92. [Google Scholar]
- Kamal, S.; Bakhtiyari, M.; Abdoli, A.; Eagderi, S.; Karami, M. Life-history variations of killifish (Aphanius sophiae) populations in two environmentally different habitats in central Iran. J. Appl. Ichthyo. 2009, 25, 474–478. [Google Scholar] [CrossRef]
- Katongo, C.; Seehausen, O.; Snoeks, J. A new species of Pseudocrenilabrus (Perciformes: Cichlidae) from Lake Mweru in the Upper Congo River System. Zootaxa 2017, 4237, 181–190. [Google Scholar] [CrossRef]
- Loiselle, P.V. Pseudocrenillabrus, the Dwarf African Mouthbrooders, Part Two: The Pseudocrenillabrus ventralis and Pseudocrenillabrus philander Complex. Freshw. Mar. Aquar. Mag. 1982, 5, 66–71. [Google Scholar]
- Katongo, C.; Koblmuller, S.; Duftner, N.; Makasa, L.; Sturmbauer, C. Phylogeography and speciation in the Pseudocrenilabrus philander species complex in Zambian Rivers. Hydrobiologia 2005, 542, 221–233. [Google Scholar] [CrossRef]
- Lamboj, A. The Cichlid Fishes of Western Africa; Birgit Schmettkamp Verlag: Bornheim, Germany, 2004; p. 255. [Google Scholar]
- Dunz, A.R.; Schliewen, U.K. Molecular phylogeny and revised classification of the haplotilapiine cichlid fishes formerly referred to as “Tilapia”. Mol. Phyl. Evol. 2013, 68, 64–80. [Google Scholar] [CrossRef] [PubMed]
- Schliewen, U.K.; SNSB-Bavarian State Collection of Zoology, Department of Ichthyology, München, Germany. Personal communication, 2017.
- Trewavas, E. Tilapiine fishes of the genera Sarotherodon, Oreochromis and Danakilia; British Mus. Nat. Hist.: London, UK, 1983; p. 583. [Google Scholar]
- Kasongo Ilunga, K.M.; Abwe, E.; Decru, E.; Manda, A.C.; Vreven, E. Description of a new small-sized Synodontis species (Siluriformes: Mochokidae) that is important for local subsistence fisheries in the Middle Lufira (upper Congo River, DR Congo). J. Fish Biol. 2020, 96, 1142–1159. [Google Scholar] [CrossRef]
- Mukweze Mulelenu, C.; Katemo Manda, B.; Decru, E.; Chocha Manda, A.; Vreven, E. The Cyphomyrus Myers 1960 (Osteoglossiformes: Mormyridae) of the Lufira Basin (Upper Lualaba: DR Congo): A generic reassignment and the description of a new species. J. Fish Biol. 2020, 96, 1123–1141. [Google Scholar] [CrossRef] [PubMed]
- Gosse, J.-P. Protopteriidae & Polypteridae. In Check-List of the Freshwater Fishes of Africa (CLOFFA); Daget, J., Gosse, J.P., Thys van den Audenaerde, D.F.E., Eds.; ORSTOM: Paris, France; MRAC: Tervuren, Belgium, 1984; Volume I, pp. 8–29. [Google Scholar]
- Katemo Manda, B. Ichtyofaune du Parc National de l’Upemba (PNU/RD Congo): Diversité, Écologie, Conservation et Gestion Durable. Ph.D. Thesis, Ku Leuven University, Leuven, Belgium, 2020. [Google Scholar]
- Skelton, P.H. Name changes and additions to the southern African freshwater fish fauna. Afr. J. Aqua. Sci 2016, 41, 345–351. [Google Scholar] [CrossRef]
- Chakona, A.; Kadye, W.T.; Bere, T.; Mazungula, D.N.; Vreven, E. Evidence of hidden diversity and taxonomic conflicts in five stream fishes from the eastern Zimbabwe highlands freshwater ecoregion. ZooKeys 2018, 768, 69–95. [Google Scholar] [CrossRef]
- Hugueny, B. Richesse des peuplements de poissons dans le Niandan Haut Niger, Afrique en fonction de la taille de la rivière et de la diversité du milieu. Rev. Hydro. Trop. 1990, 23, 351–364. [Google Scholar]
- Hugueny, B.; Lévêque, C. Freshwater fish zoogeography in west Africa: Faunal similarities between river basins. Env. Biol. Fishes 1994, 39, 365–380. [Google Scholar] [CrossRef]
- Oberdorff, T.; Tedesco, P.A.; Hugueny, B.; Leprieur, F.; Beauchard, O.; Brosse, S.; Durr, H.H. Global and regional patterns in riverine fish species richness, A review. Inter. J. Ecol. 2011, 2011, 967631. [Google Scholar] [CrossRef]
- Broadley, D.G.; Cotterill, F.P.D. The reptiles of southeast Katanga, an overlooked ‘hot spot’. Afr. J. Herp. 2004, 53, 35–61. [Google Scholar] [CrossRef]
- Thys van den Audenaerde, D.F.E. Révision systématique des espèces congolaises du genre Tilapia (Pisces, Cichlidae). Ann. Mus. R. Afr. Centr. Sci. Zool. 1964, 124, 1–155. [Google Scholar]
- Roberts, T.R. Geographical distribution of African freshwater fishes. Zool. J. Linn. Soc. 1975, 57, 249–319. [Google Scholar] [CrossRef]
- Moore, A.E.; Cotterill, F.P.D.; Eckardt, F.D. The evolution and ages of Makgadikgadi palaeo-lakes: Consilient evidence from Kalahari drainage evolution. S. Afr. J. Geol. 2012, 115, 385–413. [Google Scholar] [CrossRef]
- Bell-Cross, G. Movement of fish across the Congo-Zambezi watershed in the Mwinilunga district of Northern Rhodesia. In Proceedings of the Central African Scientific and Medical Congress, Lusaka, Zambia, 26–30 August 1963; Snowball, G.J., Ed.; Pergamon Press: Oxford, UK, 1965; pp. 415–424. [Google Scholar]
- Snoeks, J.; Harrison, I.J.; Stiassny, M.L.J. The status and distribution of freshwater fishes. In The Diversity of Life in African Freshwaters: Underwater, under Threat. An Analysis of the Status and Distribution of Freshwater Species throughout Mainland Africa; Darwall, W.R.T., Smith, K., Allen, D., Holland, R., Harrison, I., Brooks, E., Eds.; IUCN: Gland, Switzerland; Cambridge, UK, 2011; pp. 42–91. [Google Scholar]
- Stiassny, M.L.J.; Brummett, R.E.; Harrison, I.J.; Monsembula, R.; Mamonekene, V. The status and distribution of freshwater fishes of Central Africa. In The Status and Distribution of Freshwater Biodiversity in Central Africa; Brooks, E.G.E., Allen, D.J., Darwall, W.R.T., Eds.; IUCN: Gland, Switzerland; Cambridge, UK, 2011; pp. 27–47. [Google Scholar]
- IUCN. IUCN Red List Categories and Criteria: Version 3.1, 2nd ed.; IUCN: Gland, Switzerland; Cambridge, UK, 2012; p. 36. [Google Scholar]
- Van Ginneken, M.; Decru, E.; Verheyen, E.; Snoeks, J. Morphometry and DNA barcoding reveal cryptic diversity in the genus Enteromius (Cypriniformes: Cyprinidae) from the Congo basin, Africa. Euro. J. Taxo. 2017, 310, 1–32. [Google Scholar]
- Mace, G.M.; Collar, N.J.; Gaston, K.J.; Hilton-Taylor, C.; Akçakaya, H.R.; Leader-Williams, N.; Milner-Gulland, E.J.; Stuart, S.N. Quantification of extinction risk: IUCN’s system for classifying threatened species. Cons. Biol. 2008, 22, 1424–1442. [Google Scholar] [CrossRef] [PubMed]
- Abwe, E.; Chocha Manda, C.; BEZHU-Unité de recherche en Biodiversité et Exploitation durable des Zones Humides, Lubumbashi, RD, Congo; Vreven, E.; MRAC-Royal Museum for Central Africa, Tervuren, Belgium. Personal communication, 2014.
- Feller, M.C. Human impacts on streams and rivers: Deforestation and nutrient loading to fresh waters. In River Ecosystem Ecology: A Global Perspective; Likens, G.E., Ed.; Elsevier: Oxford, UK, 2009; pp. 221–236. [Google Scholar]
- Soto, D.X.; Decru, E.; Snoeks, J.; Verheyen, E.; Van de Walle, L.; Bamps, J.; Mambo, T.; Bouillon, S. Terrestrial contributions to Afrotropical aquatic food webs: The Congo River case. Ecol. Evol. 2019, 9, 10746–10757. [Google Scholar] [CrossRef]
- Katemo Manda, B.; Colinet, G.; André, L.; Chocha Manda, A.; Marquet, J.P.; Micha, J.-C. Evaluation de la contamination de la chaîne trophique par les éléments traces (Cu, Co, Zn, Pb, Cd, U, V et As) dans le bassin de la Lufira supérieure (Katanga/RD Congo). Tropicultura 2010, 28, 246–252. [Google Scholar]
- Mees., F.; Masalehdani, M.N.N.; De Putter, T.; D’Hollander, C.; Van Biezen, E.; Mujinya, B.B.; Potdevin, J.L.; Van Ranst, E. Concentrations and forms of heavy metals around two ore processing sites in Katanga, Democratic Republic of Congo. J. Afr. Earth Sci. 2012, 77, 22–30. [Google Scholar] [CrossRef]
- Thieme, M.L.; Shapiro, A.; Colom, A.; Schliewen, U.; Sindorf, N.; Toham, A.K. Inventaire Rapide des Zones Humides Représentatives en République Démocratique du Congo. 2008. Available online: http://www.ramsar.org/pdf/wurc/wurc_dr-congo_inventaire2008.pdf (accessed on 5 July 2022).
- Foster, M.N. Synthesis for all taxa: Section 8.3.4. Conservation action. In The Diversity of Life in African Freshwaters: Underwater, under Threat. An Analysis of the Status and Distribution of Freshwater Species throughout Mainland Africa; Darwall, W., Smith, K., Allen, D., Holland, R., Harrison, I., Brooks, E., Eds.; IUCN: Gland, Switzerland; Cambridge, UK, 2011; pp. 247–249. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Abwe, E.; Snoeks, J.; Manda, B.K.; Mutambala, P.K.; Kalumba, L.N.; Bragança, P.H.N.; Kipanga, K.; Mulelenu, C.M.; Kayaba, M.K.I.; Manda, A.C.; et al. Checklist of the Fishes of the Kundelungu National Park (Upper Congo Basin, DR Congo): Species Diversity and Endemicity of a Poorly Known Ichthyofauna. Diversity 2023, 15, 259. https://doi.org/10.3390/d15020259
Abwe E, Snoeks J, Manda BK, Mutambala PK, Kalumba LN, Bragança PHN, Kipanga K, Mulelenu CM, Kayaba MKI, Manda AC, et al. Checklist of the Fishes of the Kundelungu National Park (Upper Congo Basin, DR Congo): Species Diversity and Endemicity of a Poorly Known Ichthyofauna. Diversity. 2023; 15(2):259. https://doi.org/10.3390/d15020259
Chicago/Turabian StyleAbwe, Emmanuel, Jos Snoeks, Bauchet Katemo Manda, Pacifique Kiwele Mutambala, Lewis Ngoy Kalumba, Pedro H. N. Bragança, Kamwanya Kipanga, Christian Mukweze Mulelenu, Micheline Kasongo Ilunga Kayaba, Auguste Chocha Manda, and et al. 2023. "Checklist of the Fishes of the Kundelungu National Park (Upper Congo Basin, DR Congo): Species Diversity and Endemicity of a Poorly Known Ichthyofauna" Diversity 15, no. 2: 259. https://doi.org/10.3390/d15020259
APA StyleAbwe, E., Snoeks, J., Manda, B. K., Mutambala, P. K., Kalumba, L. N., Bragança, P. H. N., Kipanga, K., Mulelenu, C. M., Kayaba, M. K. I., Manda, A. C., & Vreven, E. J. W. M. N. (2023). Checklist of the Fishes of the Kundelungu National Park (Upper Congo Basin, DR Congo): Species Diversity and Endemicity of a Poorly Known Ichthyofauna. Diversity, 15(2), 259. https://doi.org/10.3390/d15020259