


Do Suburban Populations of Lizards Behave Differently from Forest Ones? An Analysis of Perch Height, Time Budget, and Display Rate in the Cuban Endemic Anolis homolechis

Abstract
:1. Introduction
2. Materials and Methods
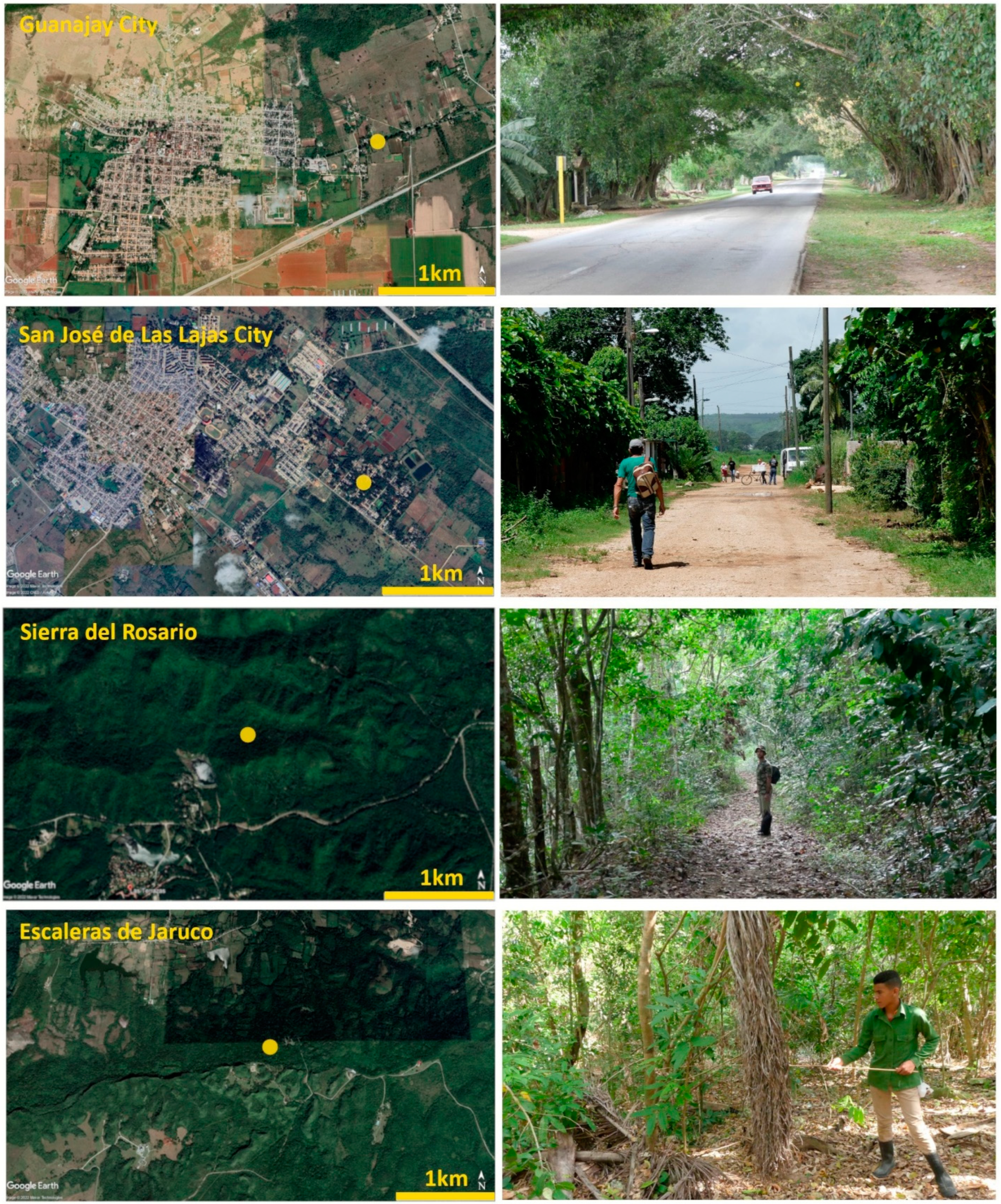
2.1. Species and Study Sites
2.2. Data Collection
2.3. Statistical Analysis
2.3.1. Perch Height
2.3.2. Time Budget and Display Rate
3. Results
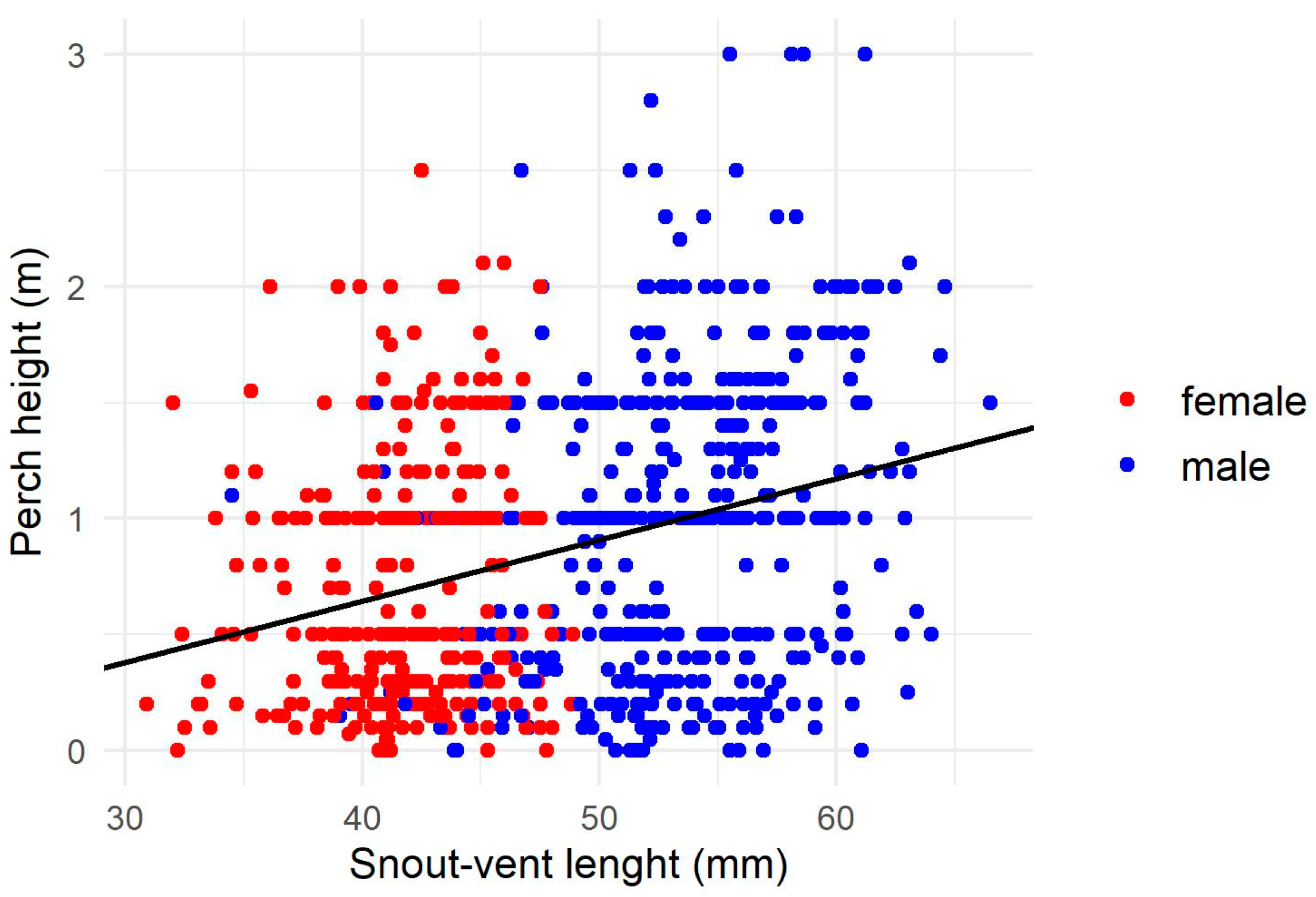
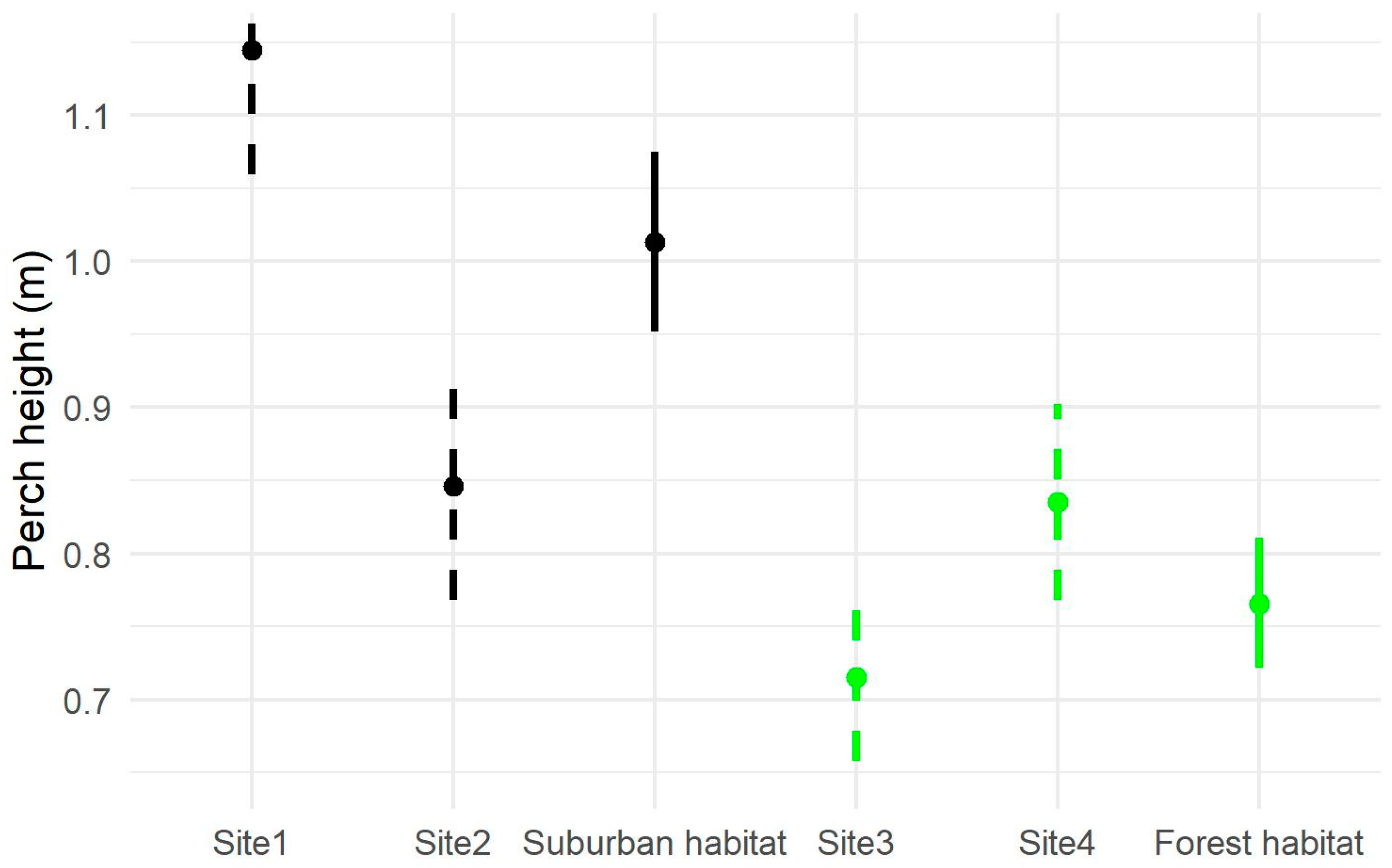
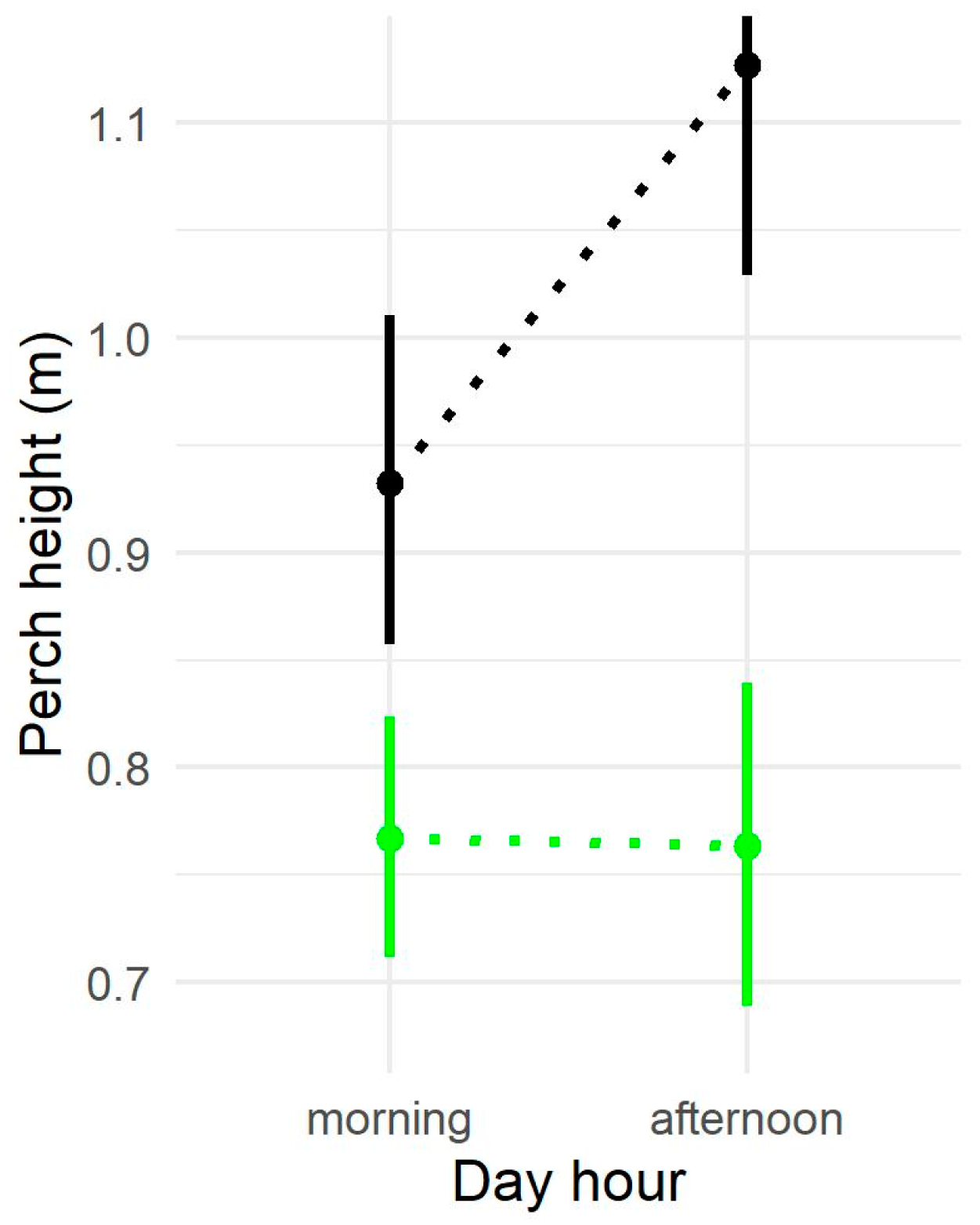
3.1. Perch Height
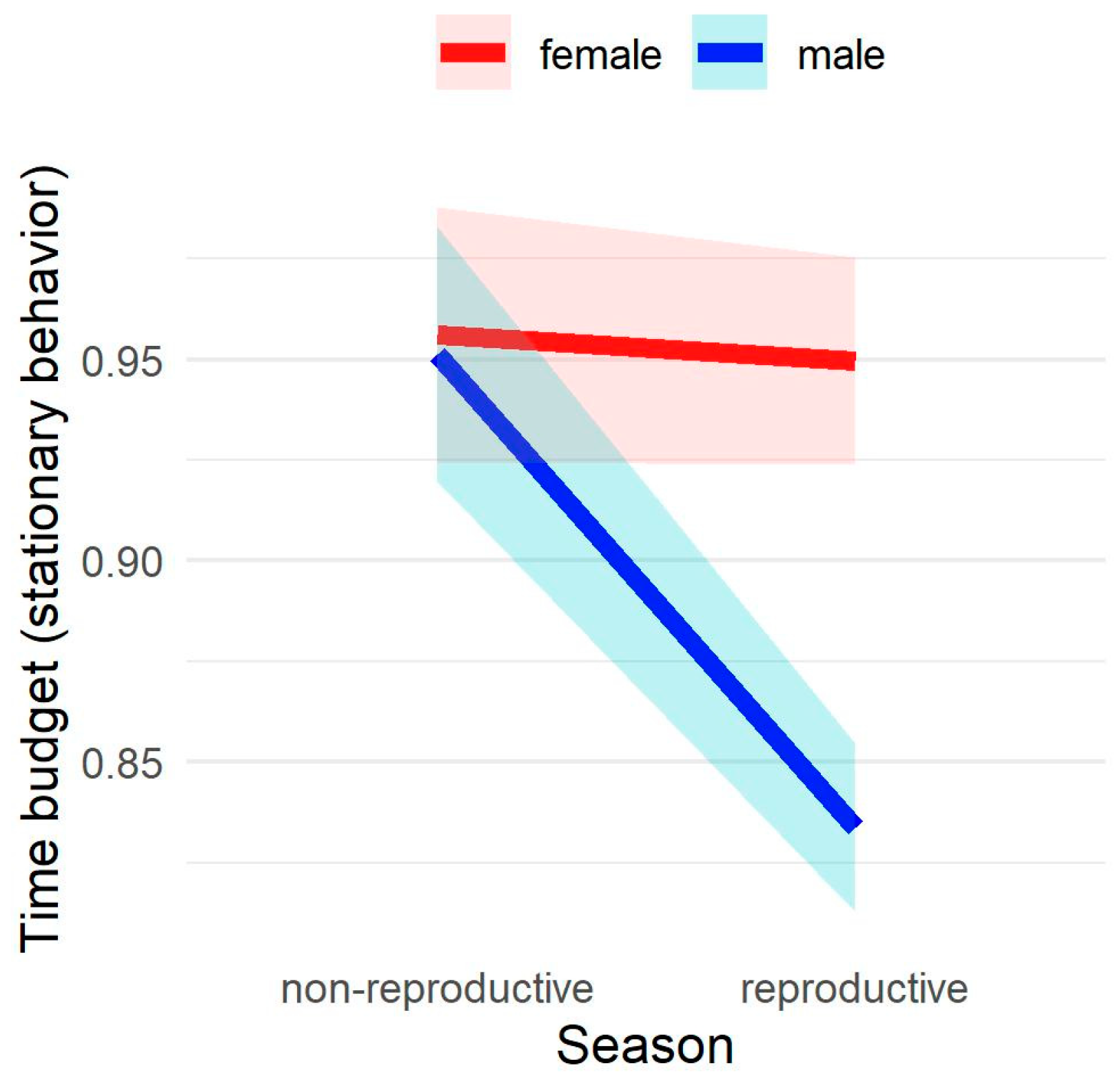
3.2. Time Budget
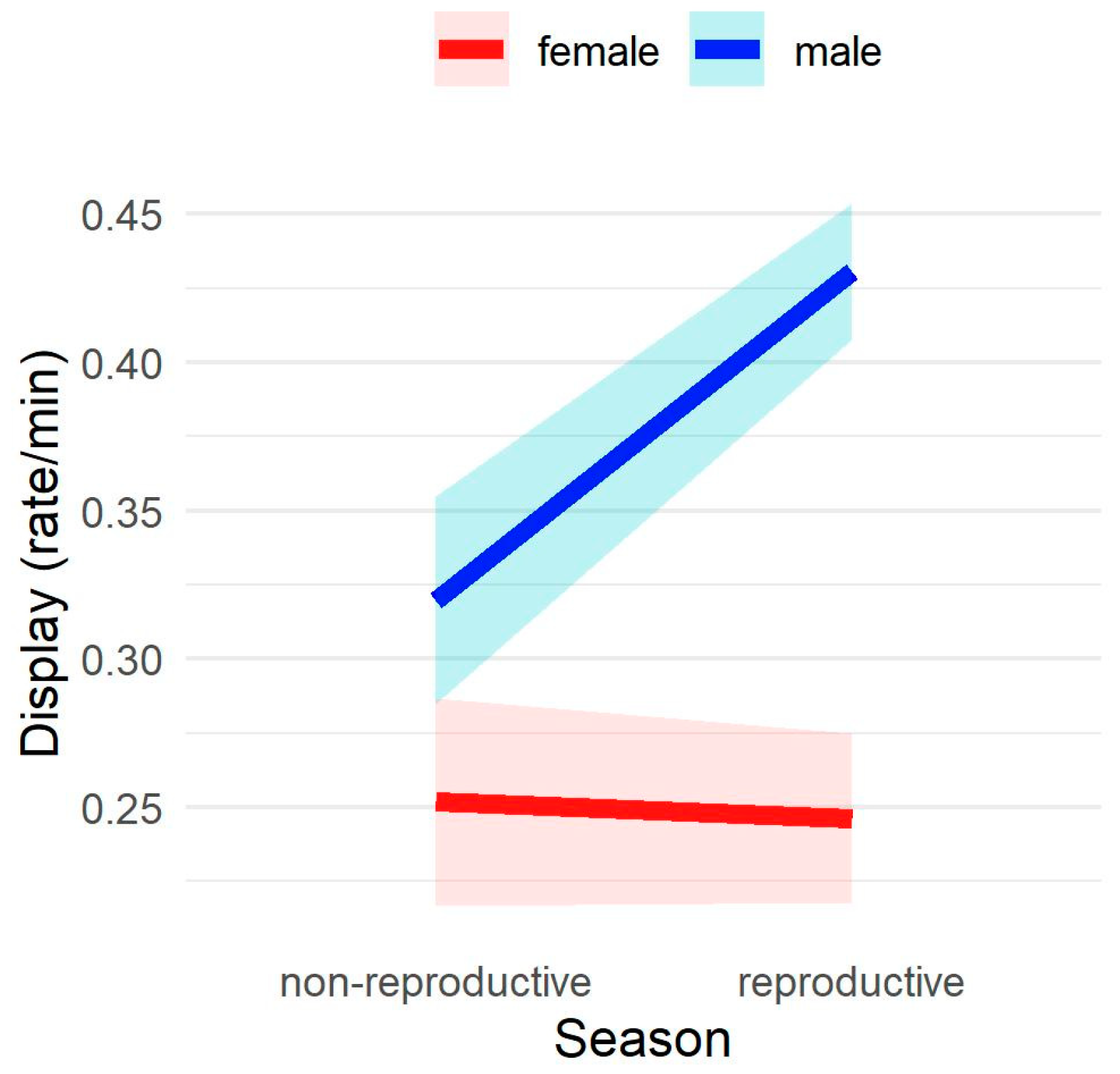
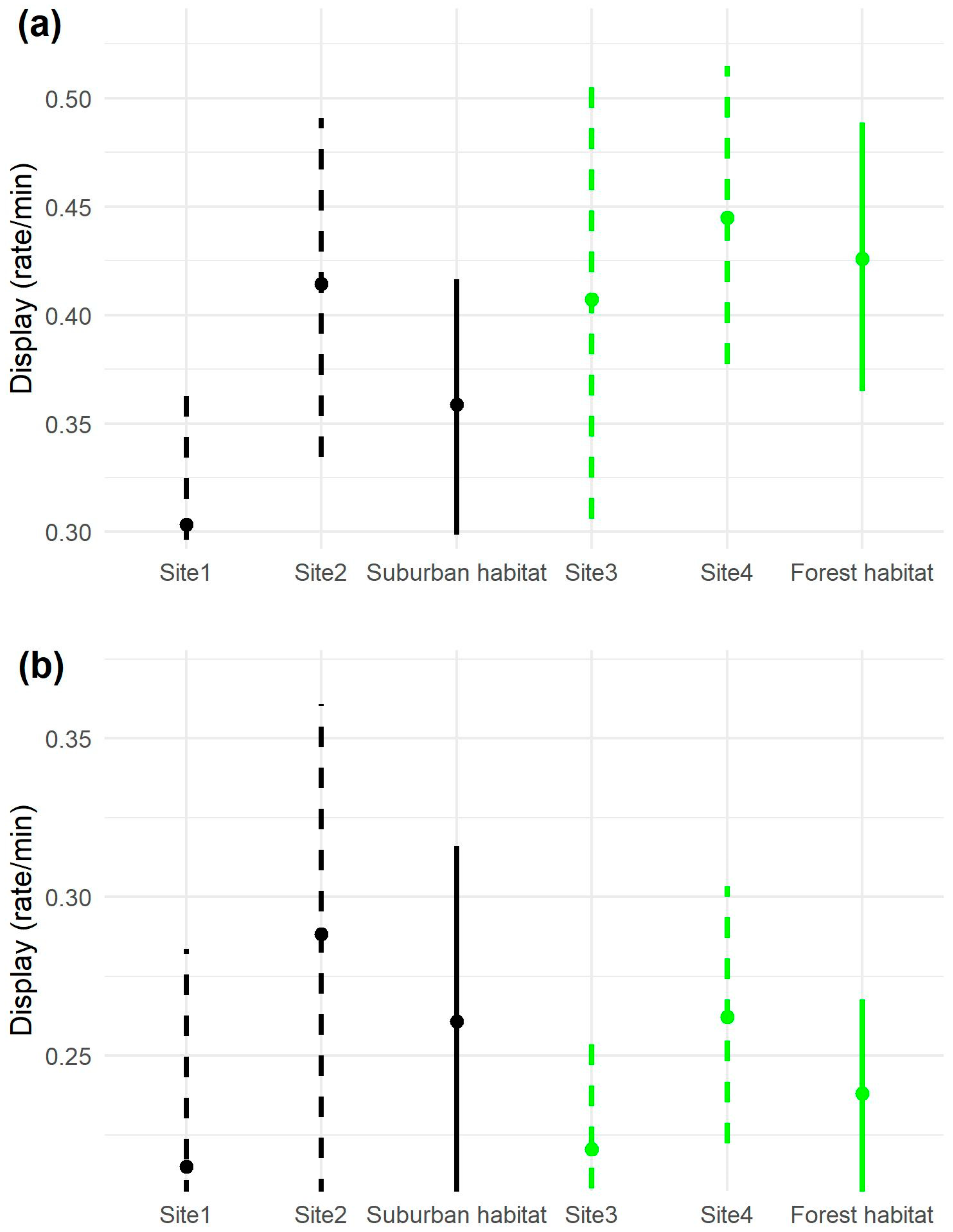
3.3. Display Rate
4. Discussion
4.1. Variation in Perch Height
4.2. Time Budget and Display Rate
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Morning | Afternoon | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Males | Females | Total | Males | Females | Total | Males | Females | Total | ||
| Suburban sites | ||||||||||
| Guanajay | 66 | 30 | 96 | 52 | 32 | 84 | 118 | 62 | 180 | |
| San José de las Lajas | 73 | 45 | 116 | 39 | 21 | 60 | 112 | 66 | 178 | |
| Forest sites | ||||||||||
| Sierra del Rosario | 72 | 72 | 144 | 53 | 42 | 95 | 125 | 114 | 239 | |
| Escaleras de Jaruco | 57 | 55 | 112 | 40 | 30 | 70 | 95 | 85 | 182 | |
Appendix B
| Reproductive Season | Non-Reproductive Season | Total | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Males | Females | Total | Males | Females | Total | Males | Females | Total | ||
| Suburban sites | ||||||||||
| Guanajay | 9 | 6 | 15 | 1 | 0 | 1 | 10 | 6 | 16 | |
| San José de las Lajas | 6 | 5 | 11 | 4 | 5 | 9 | 10 | 10 | 20 | |
| Forest sites | ||||||||||
| Sierra del Rosario | 9 | 3 | 12 | 4 | 8 | 12 | 13 | 11 | 24 | |
| Escaleras de Jaruco | 8 | 7 | 15 | 5 | 1 | 6 | 13 | 8 | 21 | |
References
- Angel, S.; Parent, J.; Civco, D.L.; Blei, A.; Potere, D. The dimensions of global urban expansion: Estimates and projections for all countries, 2000–2050. Prog. Plann. 2011, 75, 53–107. [Google Scholar] [CrossRef]
- Seto, K.C.; Fragkias, M.; Güneralp, B.; Reilly, M.K. A meta-analysis of global urban land expansion. PLoS ONE 2012, 6, e23777. [Google Scholar] [CrossRef] [PubMed]
- Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; Redman, C.L.; Wu, J.; Bai, X.; Briggs, J.M.; Grimm, N.B.; Faeth, S.H.; Golubiewski, N.E.; et al. Global change and the ecology of cities. Science 2008, 319, 756–760. [Google Scholar] [CrossRef]
- McKinney, M.L. Urbanization, biodiversity, and conservation. Bioscience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Gaston, K.J.; Duffy, J.P.; Gaston, S.; Bennie, J.; Davies, T.W. Human alteration of natural light cycles: Causes and ecological consequences. Oecologia 2014, 176, 917–931. [Google Scholar] [CrossRef]
- Arnfield, A.J. Two decades of urban climate research: A review of turbulence, exchanges of energy and water, and the urban heat island. Int. J. Climatol. 2003, 23, 1–26. [Google Scholar] [CrossRef]
- Kowarik, I. Novel urban ecosystems, biodiversity, and conservation. Environ. Pollut. 2011, 159, 1974–1983. [Google Scholar] [CrossRef] [PubMed]
- Aronson, M.F.J.; La Sorte, F.A.; Nilon, C.H.; Katti, M.; Goddard, M.A.; Lepczyk, C.A.; Warren, P.S.; Williams, N.S.G.; Cilliers, S.; Clarkson, B.; et al. A global analysis of the impacts of urbanization on bird and plant diversity reveals key anthropogenic drivers. Proc. R. Soc. B 2014, 281, 20133330. [Google Scholar] [CrossRef]
- Hahs, A.K.; McDonnell, M.J.; McCarthy, M.A.; Vesk, P.A.; Corlett, R.T.; Norton, B.A.; Clemants, S.E.; Duncan, R.P.; Thompson, K.; Schwartz, M.W.; et al. A global synthesis of plant extinction rates in urban areas. Ecol. Lett. 2009, 12, 1165–1173. [Google Scholar] [CrossRef]
- Angold, P.G.; Sadler, J.P.; Hill, M.O.; Pullin, A.; Rushton, S.; Austin, K.; Small, E.; Wood, B.; Wadsworth, R.; Sanderson, R.; et al. Biodiversity in urban habitat patches. Sci. Total Environ. 2006, 360, 196–204. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Luniak, M. Synurbization: Adaptation of animal wildlife to urban development. In Proceedings of the 4th International Urban Wildlife Symposium, Tucson, AZ, USA, 1–5 May 1999; University of Arizona: Tucson, AZ, USA, 2004; pp. 50–55. [Google Scholar]
- Alberti, M.; Correa, C.; Marzluff, J.M.; Hendry, A.P.; Palkovacs, E.P.; Gotanda, K.M.; Hunt, V.M.; Apgar, T.M.; Zhou, Y. Global urban signatures of phenotypic change in animal and plant populations. Proc. Natl. Acad. Sci. USA 2017, 114, 8951–8956. [Google Scholar] [CrossRef]
- Liker, A. Biologia futura: Adaptive changes in urban populations. Biol. Futur. 2020, 71, 1–8. [Google Scholar] [CrossRef]
- Ouyang, J.Q.; Isaksson, C.; Schmidt, C.; Hutton, P.; Bonier, F.; Dominoni, D. A new framework for urban ecology: An integration of proximate and ultimate responses to anthropogenic change. Integr. Comp. Biol. 2018, 58, 915–928. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Jenkins, J.C.; Law, N.L.; Nilon, C.H.; Pouyat, R.V.; Szlavecz, K.; Warren, P.S.; Wilson, M.A.; Cadenasso, M.L.; Grove, J.M.; et al. Beyond urban legends: An emerging framework of urban ecology, as illustrated by the Baltimore ecosystem study. Bioscience 2008, 58, 139–150. [Google Scholar] [CrossRef]
- Phearson, T.M.; Pickett, S.T.A.A.; Grimm, N.B.; Niemelä, J.; Alberti, M.; Elmqvist, T.; Weber, C.; Haase, D.; Breuste, J.; Qureshi, S.; et al. Advancing urban ecology toward a science of cities. Bioscience 2016, 66, 198–212. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.S.; Saalfeld, S.T.S.; Gibson, C.J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Kralj-Fišer, S.; Hebets, E.A.; Kuntner, M. Different patterns of behavioral variation across and within species of spiders with differing degrees of urbanization. Behav. Ecol. Sociobiol. 2017, 71, 125. [Google Scholar] [CrossRef]
- Łopucki, R.; Klich, D.; Kiersztyn, A. Changes in the social behavior of urban animals: More aggression or tolerance? Mamm. Biol. 2021, 101, 1–10. [Google Scholar] [CrossRef]
- Sarkar, R.; Bhadra, A. How do animals navigate the urban jungle? A review of cognition in urban-adapted animals. Curr. Opin. Behav. Sci. 2022, 46, 101177. [Google Scholar] [CrossRef]
- Sih, A.; Stamps, J.; Yang, L.H.; McElreath, R.; Ramenofsky, M. Behavior as a key component of integrative biology in a human-altered world. Integr. Comp. Biol. 2010, 50, 934–944. [Google Scholar] [CrossRef] [PubMed]
- Shochat, E.; Warren, P.S.; Faeth, S.H.; McIntyre, N.E.; Hope, D. From patterns to emerging processes in mechanistic urban ecology. Trends Ecol. Evol. 2006, 21, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Sol, D.; Lapiedra, O.; González-Lagos, C. Behavioural adjustments for a life in the city. Anim. Behav. 2013, 85, 1101–1112. [Google Scholar] [CrossRef]
- Carrete, M.; Tella, J.L. Behavioral correlations associated with fear of humans differ between rural and urban burrowing owls. Front. Ecol. Evol. 2017, 5, 1–9. [Google Scholar] [CrossRef]
- Vincze, E.; Papp, S.; Preiszner, B.; Seress, G.; Bókony, V.; Liker, A. Habituation to human disturbance is faster in urban than rural house sparrows. Behav. Ecol. 2016, 27, 1304–1313. [Google Scholar] [CrossRef]
- Uchida, K.; Suzuki, K.K.; Shimamoto, T.; Yanagawa, H.; Koizumi, I. Decreased vigilance or habituation to humans? Mechanisms on increased boldness in urban animals. Behav. Ecol. 2019, 30, 1583–1590. [Google Scholar] [CrossRef]
- Lapiedra, O.; Chejanovski, Z.; Kolbe, J.J. Urbanization and biological invasion shape animal personalities. Glob. Chang. Biol. 2017, 23, 592–603. [Google Scholar] [CrossRef]
- Avilés-Rodríguez, K.J.; Kolbe, J.J. Escape in the city: Urbanization alters the escape behavior of Anolis lizards. Urban Ecosyst. 2019, 22, 733–742. [Google Scholar] [CrossRef]
- Batabyal, A.; Balakrishna, S.; Thaker, M. A multivariate approach to understanding shifts in escape strategies of urban lizards. Behav. Ecol. Sociobiol. 2017, 71, 83. [Google Scholar] [CrossRef]
- Bloch, N.; Irschick, D.J. An analysis of inter-population divergence in visual display behavior of the green anole lizard (Anolis carolinensis). Ethology 2006, 112, 370–378. [Google Scholar] [CrossRef]
- Baxter-Gilbert, J.; Riley, J.L.; Whiting, M.J. Bold New World: Urbanization promotes an innate behavioral trait in a lizard. Behav. Ecol. Sociobiol. 2019, 73, 105. [Google Scholar] [CrossRef]
- Atwell, J.W.; Cardoso, G.C.; Whittaker, D.J.; Campbell-Nelson, S.; Robertson, K.W.; Ketterson, E.D. Boldness behavior and stress physiology in a novel urban environment suggest rapid correlated evolutionary adaptation. Behav. Ecol. 2012, 23, 960–969. [Google Scholar] [CrossRef]
- Miranda, A.C.; Schielzeth, H.; Sonntag, T.; Partecke, J. Urbanisation and its effects on personality traits: A result of microevolution or phenotypic plasticity? Glob. Chang. Biol. 2013, 19, 2634–2644. [Google Scholar] [CrossRef]
- Cronin, A.D.; Smit, J.A.H.; Muñoz, M.I.; Poirier, A.; Moran, P.A.; Jerem, P.; Halfwerk, W. A comprehensive overview of the effects of urbanisation on sexual selection and sexual traits. Biol. Rev. 2022, 97, 1325–1345. [Google Scholar] [CrossRef]
- Ríos-Chelén, A.A. Bird song: The interplay between urban noise and sexual selection. Oecologia Bras. 2009, 13, 153–164. [Google Scholar] [CrossRef]
- Stroud, J.T.; Colom, M.; Ferrer, P.; Palermo, N.; Vargas, V.; Cavallini, M.; Lopez, J.; Jones, I. Behavioral shifts with urbanization may facilitate biological invasion of a widespread lizard. Urban Ecosyst. 2019, 22, 425–434. [Google Scholar] [CrossRef]
- Halfwerk, W.; Blaas, M.; Kramer, L.; Hijner, N.; Trillo, P.A.; Bernal, X.E.; Page, R.A.; Goutte, S.; Ryan, M.J.; Ellers, J. Adaptive changes in sexual signalling in response to urbanization. Nat. Ecol. Evol. 2019, 3, 374–380. [Google Scholar] [CrossRef]
- Lintott, P.R.; Bunnefeld, N.; Fuentes-Montemayor, E.; Minderman, J.; Mayhew, R.J.; Olley, L.; Park, K.J. City life makes females fussy: Sex differences in habitat use of temperate bats in urban areas. R. Soc. Open Sci. 2014, 1, 140200. [Google Scholar] [CrossRef]
- Magle, S.B.; Hunt, V.M.; Vernon, M.; Crooks, K.R. Urban wildlife research: Past, present, and future. Biol. Conserv. 2012, 155, 23–32. [Google Scholar] [CrossRef]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [Green Version]
- Uetz, P.; Freed, P.; Aguilar, R.; Hošek, J. The Reptile Database. Available online: http://www.reptile-database.org (accessed on 10 September 2022).
- Lapiedra, O. Urban behavioral ecology: Lessons from Anolis lizards. Integr. Comp. Biol. 2018, 58, 939–947. [Google Scholar] [CrossRef] [PubMed]
- Losos, J.B. Integrative approaches to evolutionary ecology: Anolis lizards as model systems. Annu. Rev. Ecol. Syst. 1994, 25, 467–493. [Google Scholar] [CrossRef]
- Losos, J.B. Lizards in an Evolutionary Tree: Ecology and Adaptive Radiation of Anoles; University of California Press: Berkeley, CA, USA, 2009; p. 507. [Google Scholar]
- Schwartz, A.; Henderson, R.W. Amphibians and Reptiles of the West Indies: Descriptions, Distributions, and Natural History; University of Florida Press: Gainesville, FL, USA, 1991; p. 720. [Google Scholar]
- Andrews, R.M. Structural habitat and time budget of a tropical Anolis lizard. Ecol. Soc. Am. 1971, 52, 262–270. [Google Scholar] [CrossRef]
- Staats, C.M.; Bartolotia, T.; Schall, J.J. Body size and perching behavior of Anolis sabanus, the solitary anole of Saba, Netherland Antilles. Herpetol. Nat. Hist. 1997, 5, 157–164. [Google Scholar]
- Gilman, C.A.; Irschick, D.J. Foils of flexion: The effects of perch compliance on lizard locomotion and perch choice in the wild. Funct. Ecol. 2013, 27, 374–381. [Google Scholar] [CrossRef]
- Jenssen, T.A. The ethoecology of Anolis nebulosus (Sauria, Iguanidae). J. Herpetol. 1970, 4, 1–38. [Google Scholar] [CrossRef]
- Rodríguez Schettino, L. The Iguanid Lizards of Cuba; University of Florida Press: Gainesville, FL, USA, 1999; p. 428. [Google Scholar]
- Ramírez-Bautista, A.; Benabib, M. Perch height of the arboreal lizard Anolis nebulosus (Sauria: Polychrotidae) from a tropical dry forest of México: Effect of the reproductive season. Copeia 2001, 2001, 187–193. [Google Scholar] [CrossRef]
- Paterson, A. Effects of prey availability on perch height of female bark anoles, Anolis distichus. Herpetologica 1999, 42, 412–449. [Google Scholar]
- Barquero, M.D.; Arguedas, V.P. Structural habitat use by the many-scaled anole, Anolis polylepis (Squamata: Polychrotidae). Acta Herpetol. 2009, 4, 135–142. [Google Scholar] [CrossRef]
- Cooper, W.E. Risk factors affecting escape behavior by the Jamaican lizard Anolis lineatopus (Polychrotidae, Squamata). Caribb. J. Sci. 2010, 46, 216–227. [Google Scholar] [CrossRef]
- Steffen, J.E. Perch height differences among female Anolis polylepis exhibiting dorsal pattern polymorphism. Amphib. Reptil. Conserv. 2010, 17, 172–174. [Google Scholar]
- Hall, J.M.; Warner, D.A. Body size and reproduction of a non-native lizard are enhanced in an urban environment. Biol. J. Linn. Soc. 2017, 122, 860–871. [Google Scholar] [CrossRef]
- Prado-Irwin, S.; Revell, L.J.; Winchell, K.M. Variation in tail morphology across urban and forest populations of the crested anole (Anolis cristatellus). Biol. J. Linn. Soc. 2019, 128, 632–644. [Google Scholar] [CrossRef]
- Thawley, C.J.; Moniz, H.A.; Merritt, A.J.; Battles, A.C.; Michaelides, S.N.; Kolbe, J.J. Urbanization affects body size and parasitism but not thermal preferences in Anolis lizards. J. Urban Ecol. 2019, 5, 1–9. [Google Scholar] [CrossRef]
- Vidal, A.; Iturriaga, M.; Mancina, C.A.; Cézilly, F. Differences in sex ratio, tail autotomy, body size and body condition between suburban and forest populations of the Cuban endemic lizard Anolis homolechis. Urban Ecosyst. 2022, 25, 1711–1723. [Google Scholar] [CrossRef]
- Winchell, K.M.; Reynolds, R.G.; Prado-Irwin, S.R.; Puente-Rolón, A.R.; Revell, L.J. Phenotypic shifts in urban areas in the tropical lizard Anolis cristatellus. Evolution 2016, 70, 1009–1022. [Google Scholar] [CrossRef]
- Lailvaux, S.P. It’s not easy being green: Behavior, morphology, and population structure in urban and natural populations of green anole (Anolis carolinensis) lizards. Front. Ecol. Evol. 2020, 8, 570810. [Google Scholar] [CrossRef]
- Fokidis, H.B.; Brock, T. Hurricane Irma induces divergent behavioral and hormonal impacts on an urban and forest population of invasive Anolis lizards: Evidence for an urban resilience hypothesis. J. Urban Ecol. 2020, 6, 1–15. [Google Scholar] [CrossRef]
- McMillan, D.M.; Irschick, D.J. Experimental test of predation and competition pressures on the green anole (Anolis carolinensis) in varying structural habitats. J. Herpetol. 2010, 44, 272–278. [Google Scholar] [CrossRef]
- Winchell, K.M.; Carlen, E.J.; Puente-Rolón, A.R.; Revell, L.J. Divergent habitat use of two urban lizard species. Ecol. Evol. 2018, 8, 25–35. [Google Scholar] [CrossRef] [PubMed]
- Chejanovski, Z.A.; Avilés-Rodríguez, K.J.; Lapiedra, O.; Preisser, E.L.; Kolbe, J.J. An experimental evaluation of foraging decisions in urban and natural forest populations of Anolis lizards. Urban Ecosyst. 2017, 20, 1011–1018. [Google Scholar] [CrossRef]
- Battles, A.C.; Moniz, M.; Kolbe, J.J. Living in the big city: Preference for broad substrates results in niche expansion for urban Anolis lizards. Urban Ecosyst. 2018, 21, 1087–1095. [Google Scholar] [CrossRef]
- Irschick, D.J.; Carlisle, E.; Elstrott, J.; Ramos, M.; Buckley, C.; Vanhooydonck, B.; Meyers, J.; Herrel, A. A comparison of habitat use, morphology, clinging performance and escape behaviour among two divergent green anole lizard (Anolis carolinensis) populations. Biol. J. Linn. Soc. 2005, 85, 223–234. [Google Scholar] [CrossRef] [Green Version]
- Borden, J.B.; Bohlman, S.; Scheffers, B.R. Niche lability mitigates the impact of invasion but not urbanization. Oecologia 2022, 198, 1–10. [Google Scholar] [CrossRef]
- Leal, M.; Powell, B.J. Behavioural flexibility and problem-solving in a tropical lizard. Biol. Lett. 2012, 8, 28–30. [Google Scholar] [CrossRef]
- Jenssen, T.A.; Congdon, J.D.; Fischer, R.U.; Estes, R.; Kling, D.; Edmands, S.; Berna, H. Behavioural, thermal, and metabolic characteristics of a wintering lizard (Anolis carolinensis) from South Carolina. Funct. Ecol. 1996, 10, 201–209. [Google Scholar] [CrossRef]
- Driessens, T.; Vanhooydonck, B.; Van Damme, R. Deterring predators, daunting opponents or drawing partners? Signaling rates across diverse contexts in the lizard Anolis sagrei. Behav. Ecol. Sociobiol. 2014, 68, 173–184. [Google Scholar] [CrossRef]
- Fischer, E.M.; Oleson, K.W.; Lawrence, D.M. Contrasting urban and rural heat stress responses to climate change. Geophys. Res. Lett. 2012, 39, 1–8. [Google Scholar] [CrossRef]
- Williams, E.E.; Rand, A.S. Species recognition, dewlap function, and faunal size. Am. Zool. 1977, 17, 261–270. [Google Scholar] [CrossRef]
- Ruiz García, F.N. Observaciones etológicas sobre Anolis homolechis (Lacertilia: Iguanidae) en Cuba. Miscelánea Zoológica 1975, 1, 4. [Google Scholar]
- González Carralero, A.; Rodríguez Schettino, L. Reproducción en cautiverio de Anolis homolechis Cope, 1864 (Squamata: Iguanidae) en Cuba. Cubazoo 2009, 1, 3–9. [Google Scholar]
- Silva, A.; Estrada, A.R. Ciclos reproductivos de dos lagartos del género Anolis: A. homolechis y A. allogus en la estación Sierra del Rosario, Cuba. Ciencias Biológicas 1984, 12, 81–89. [Google Scholar]
- Calsbeek, R.; Irschick, D.J. The quick and the dead: Correlational selection on morphology, performance, and habitat use in island lizards. Evolution 2007, 61, 2493–2503. [Google Scholar] [CrossRef]
- Vidal, A.; Cézilly, F.; Pradel, R. Contemporary survival selection fails to explain observed patterns of phenotypic divergence between suburban and forest populations of the cuban endemic lizard, Anolis homolechis. in preparation.
- Johnson, M.A. A new method of temporarily marking lizards. Herpetol. Rev. 2005, 36, 277–279. [Google Scholar]
- Altmann, J. Observational study of behavior: Sampling methods. Behaviour 1974, 49, 227–267. [Google Scholar] [CrossRef]
- Burnham, K.P.; Anderson, D.R. Model selection and multimodel inference: A practical information-theoretic approach. Ecol. Modell. 2002, 172, 488. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: http://www.R-project.org (accessed on 30 May 2022).
- Talbot, J.J. Time budget, niche overlap, inter-and intraspecific aggression in Anolis humilis and A. limifrons from Costa Rica. Copeia 1979, 472–481. [Google Scholar] [CrossRef]
- Butler, M.A.; Losos, J.B. Multivariate sexual dimorphism, selection, and adaptation in greater antillean Anolis lizards. Ecol. Monogr. 2002, 72, 541–559. [Google Scholar] [CrossRef]
- Perry, G. The evolution of sexual dimorphism in the lizard Anolis polylepis (Iguania): Evidence from intraspecific variation in foraging behaviour and diet. Can. J. Zool. 1996, 74, 1238–1425. [Google Scholar] [CrossRef]
- Rodríguez-Schettino, L. Diet of Anolis homolechis (Cope, 1864) at the National Botanical Garden, Cuba. Rev. Colomb. Cienc. Anim. 2010, 2, 147–152. [Google Scholar] [CrossRef]
- Sampedro-Marín, A.; Berovides-Álvarez, V.; Rodríguez-Schettino, L. Algunos aspectos ecológicos sobre dos especies cubanas del género Anolis (Sauria: Iguanidae). Ciencias Biológicas 1982, 7, 87–103. [Google Scholar]
- Crandell, K.E.; Herrel, A.; Sasa, M.; Losos, J.B.; Autumn, K. Stick or grip? Co-evolution of adhesive toepads and claws in Anolis lizards. Zoology 2014, 117, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Kolbe, J.J.; Battles, A.C.; Avilés-Rodríguez, K.J. City slickers: Poor performance does not deter Anolis lizards from using artificial substrates in human-modified habitats. Funct. Ecol. 2016, 30, 1418–1429. [Google Scholar] [CrossRef]
- Putman, B.J.; Tippie, Z.A. Big city living: A global meta-analysis reveals positive impact of urbanization on body size in lizards. Front. Ecol. Evol. 2020, 8, 580745. [Google Scholar] [CrossRef]
- Chejanovski, Z.A.; Kolbe, J.J. Close encounters of the urban kind: Predators influence prey body size variation in an urban landscape. Evol. Ecol. 2019, 33, 791–809. [Google Scholar] [CrossRef]
- Battles, A.C.; Kolbe, J.J. Miami heat: Urban heat islands influence the thermal suitability of habitats for ectotherms. Glob. Chang. Biol. 2019, 25, 562–576. [Google Scholar] [CrossRef]
- Lapiedra, O.; Schoener, T.W.; Leal, M.; Losos, J.B.; Kolbe, J.J. Predator-driven natural selection on risk-taking behavior in anole lizards. Science 2018, 360, 1017–1020. [Google Scholar] [CrossRef]
- Magura, T.; Mizser, S.; Horváth, R.; Nagy, D.D.; Tóth, M.; Csicsek, R.; Lövei, G.L. Are there personality differences between rural vs. urban-living individuals of a specialist ground beetle, Carabus convexus? Insects 2021, 12, 646. [Google Scholar] [CrossRef]
- Garland, T.; Albuquerque, R.L. Locomotion, energetics, performance, and behavior: A mammalian perspective on lizards, and vice versa. Integr. Comp. Biol. 2017, 57, 252–266. [Google Scholar] [CrossRef] [Green Version]
- Pough, F.H. The advantages of ectothermy for tetrapods. Am. Nat. 1980, 115, 92–112. [Google Scholar] [CrossRef]
- Hertz, P.E.; Huey, R.B.; Garland, T. Time budgets, thermoregulation, and maximal locomotor performance: Are reptiles olympians or boy scouts? Integr. Comp. Biol. 1988, 28, 927–938. [Google Scholar] [CrossRef]
- Lister, B.C.; García Aguayo, A. Seasonality, predation, and the behaviour of a tropical mainland anole. J. Anim. Ecol. 1992, 61, 717–733. [Google Scholar] [CrossRef]
- Fleming, T.H.; Hooker, R.S. Anolis cupreus: The response of a lizard to tropical seasonality. Ecology 1975, 56, 1243–1261. [Google Scholar] [CrossRef]
- García, A.; Valtierra-Azotla, M.; Lister, B.C. Behavioral responses to seasonality by two Sceloporine lizard species from a tropical dry forest. Anim. Biol. 2010, 60, 97–113. [Google Scholar] [CrossRef]
- Simon, V.B. Not all signals are equal: Male brown anole lizards (Anolis sagrei) selectively decrease pushup frequency following a simulated predatory attack. Ethology 2007, 113, 793–801. [Google Scholar] [CrossRef]
- Thorpe, R.S.; Reardon, J.T.; Malhotra, A. Common garden and natural selection experiments support ecotypic differentiation in the Dominican anole (Anolis oculatus). Am. Nat. 2005, 165, 495–504. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vidal, A.; Pradel, R.; Cézilly, F. Do Suburban Populations of Lizards Behave Differently from Forest Ones? An Analysis of Perch Height, Time Budget, and Display Rate in the Cuban Endemic Anolis homolechis. Diversity 2023, 15, 261. https://doi.org/10.3390/d15020261
Vidal A, Pradel R, Cézilly F. Do Suburban Populations of Lizards Behave Differently from Forest Ones? An Analysis of Perch Height, Time Budget, and Display Rate in the Cuban Endemic Anolis homolechis. Diversity. 2023; 15(2):261. https://doi.org/10.3390/d15020261
Chicago/Turabian StyleVidal, Annabelle, Roger Pradel, and Frank Cézilly. 2023. "Do Suburban Populations of Lizards Behave Differently from Forest Ones? An Analysis of Perch Height, Time Budget, and Display Rate in the Cuban Endemic Anolis homolechis" Diversity 15, no. 2: 261. https://doi.org/10.3390/d15020261
APA StyleVidal, A., Pradel, R., & Cézilly, F. (2023). Do Suburban Populations of Lizards Behave Differently from Forest Ones? An Analysis of Perch Height, Time Budget, and Display Rate in the Cuban Endemic Anolis homolechis. Diversity, 15(2), 261. https://doi.org/10.3390/d15020261