
Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland


Abstract
:1. Introduction
2. Materials and Methods
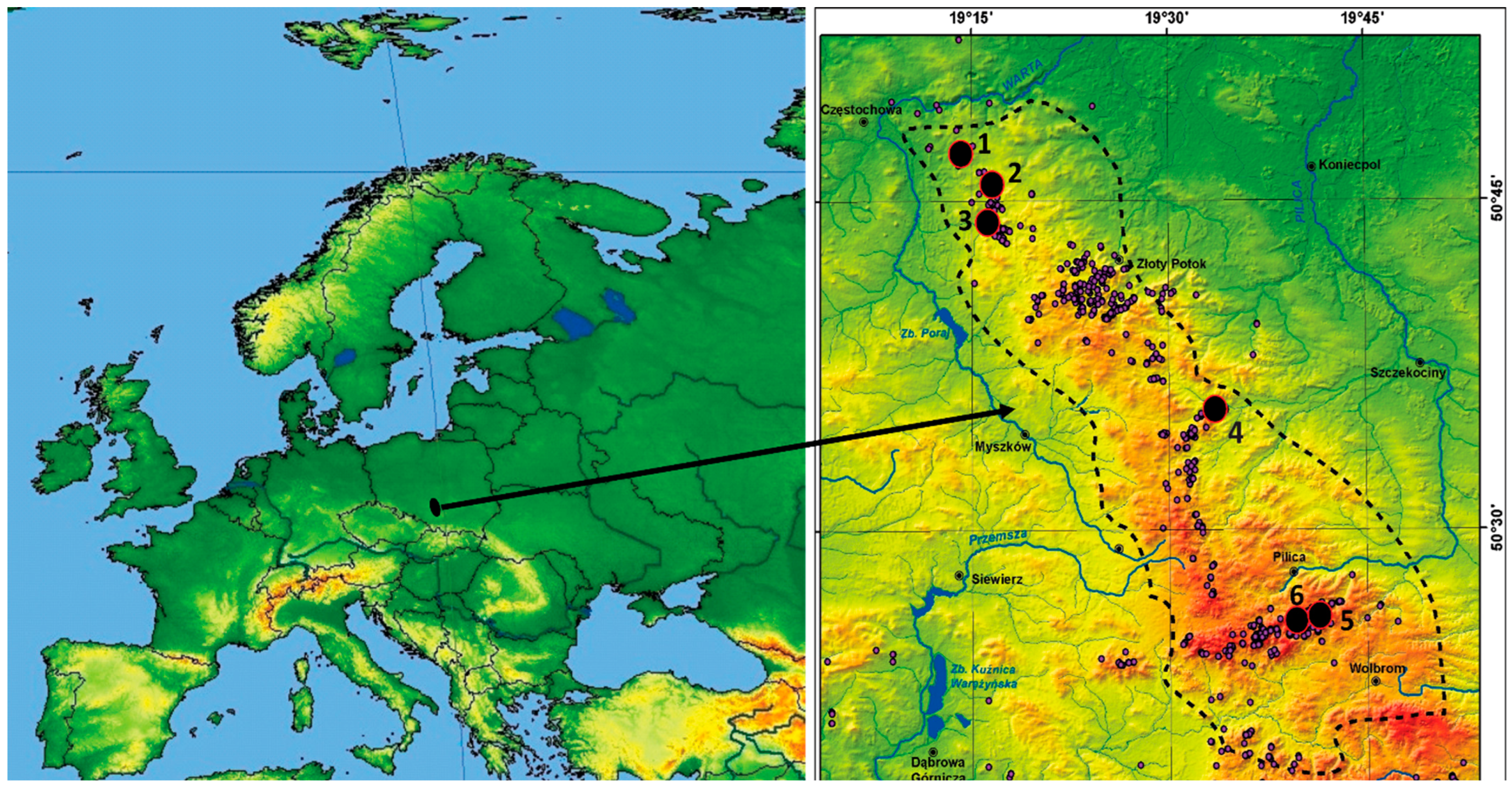
2.1. Study Area
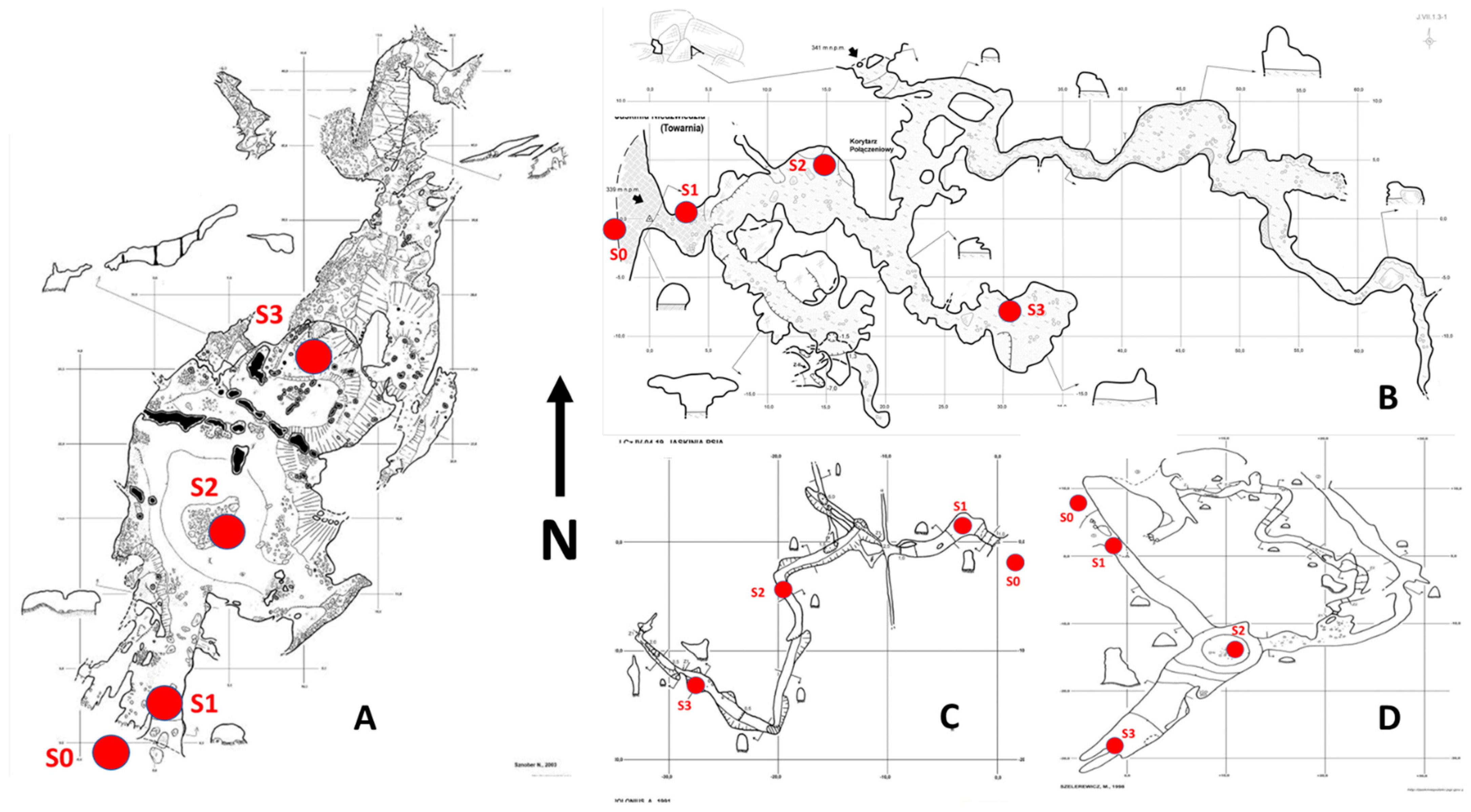
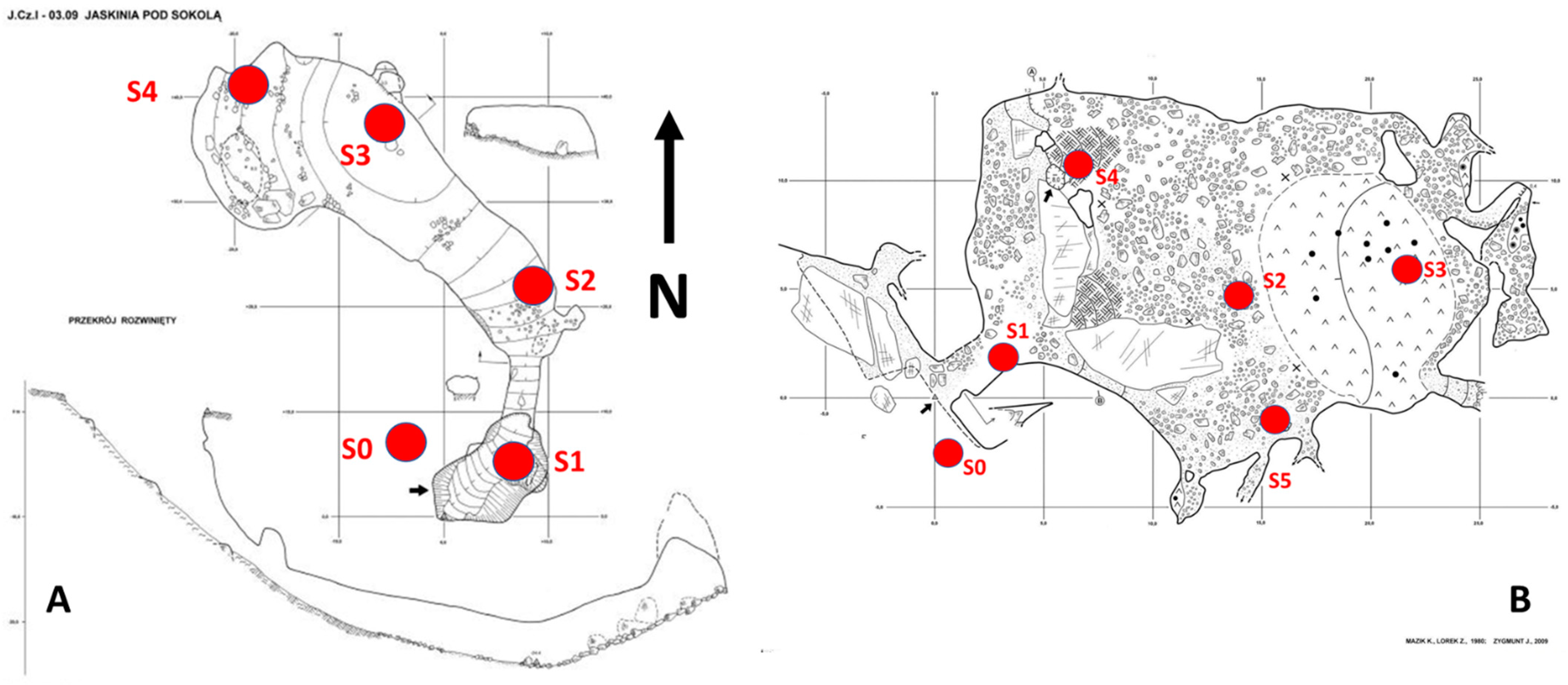
2.2. Sampling Design
2.3. Microclimate Measurements
2.4. Data Analysis
3. Results
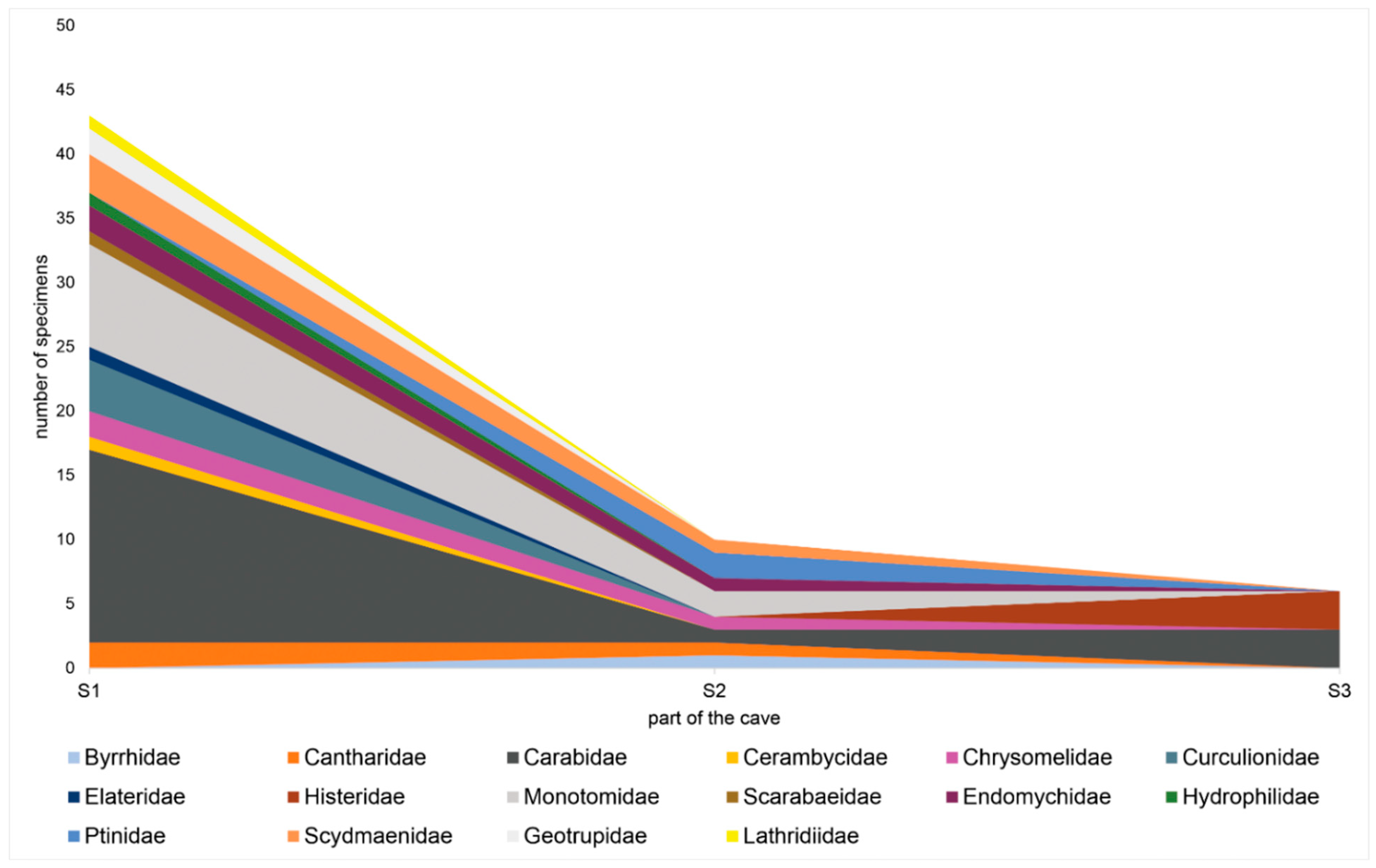
3.1. Species Dispersal
3.2. Species Richness
3.3. Species Dominance
3.4. Habitat and Food Preferences
3.5. Ecological Classification
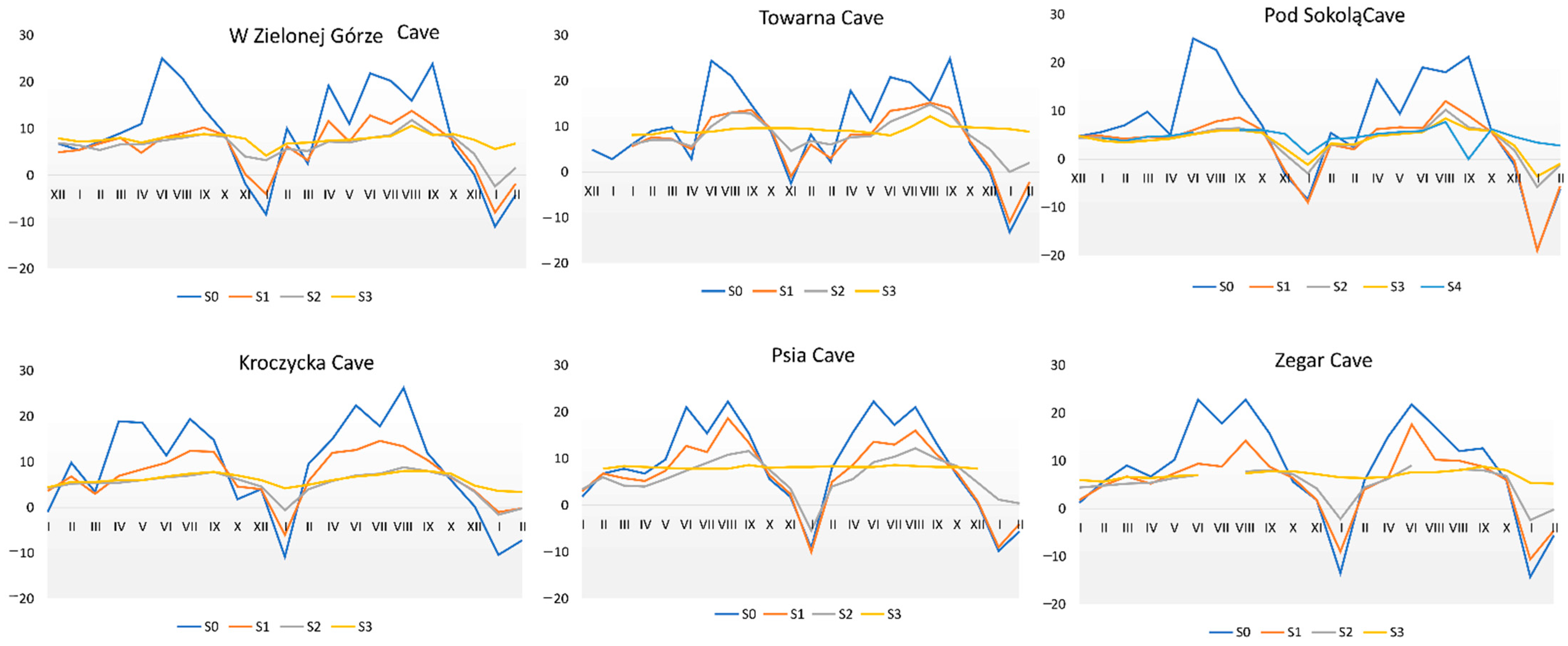
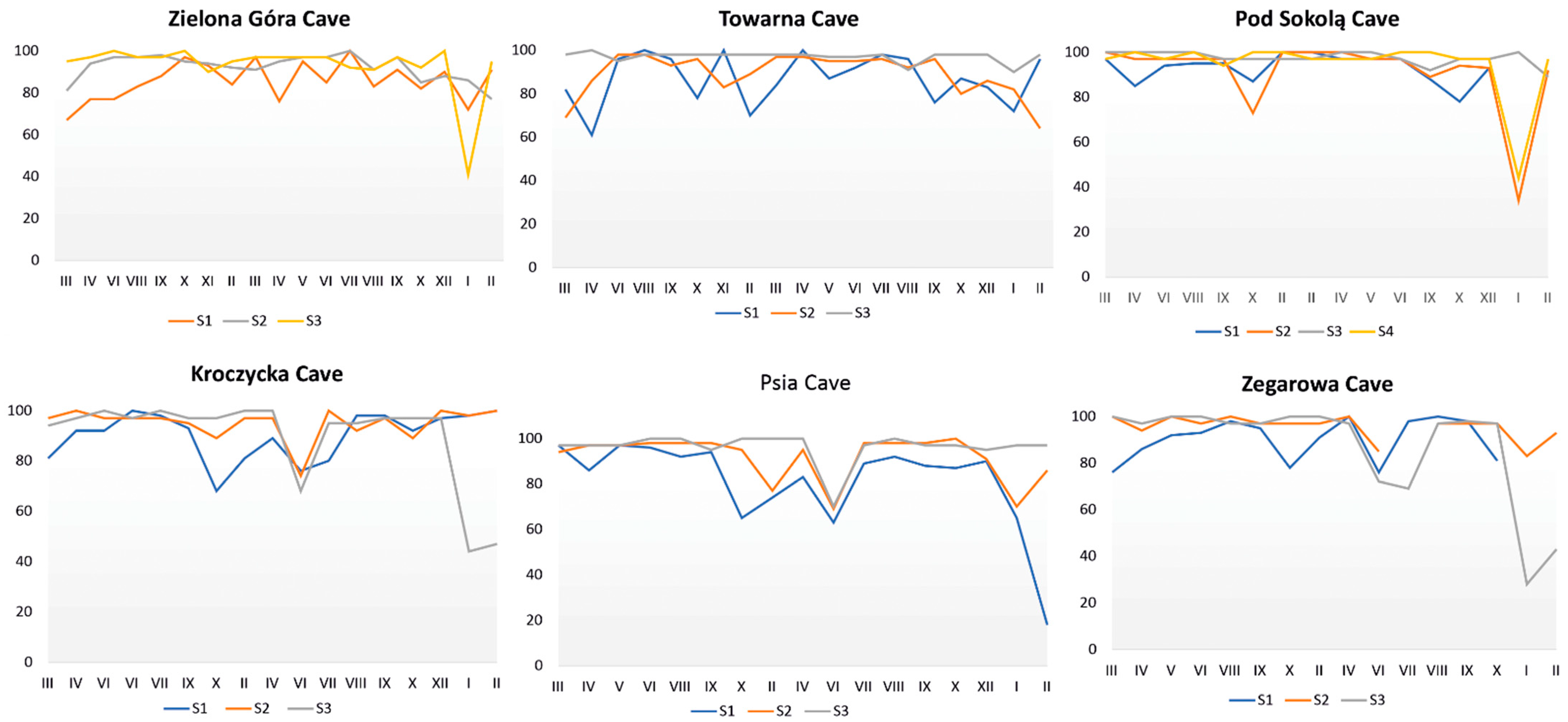
3.6. Microclimatic Analyses
4. Discussion
4.1. Species Richness
4.2. Species Dominance
4.3. Habitat and Food Preferences
4.4. Ecological Classification
5. Conclusions
- Many beetle species choose to dwell in caves, despite lacking specific adaptations for living in such environments.
- The entrance zone is the most attractive place for surface species to inhabit because the climatic conditions there are more stable than outside the cave (due to the presence of some light and high availability of organic matter).
- In the deeper parts of the studied caves, the number of occurring species rapidly decreased, though this zone sometimes had a stable environmental condition (namely, humidity).
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- 1D—Shannon diversity;
- H′—the Shannon index;
- pi —the proportion (ni/N) of individuals of ith species found (ni) divided by the total number of individuals found (N).
- Σ pi2—the Simpson index;
- pi—number of specimens of species ith in the community.
- Di—dominance index;
- ni—number of individuals in ith species;
- N—total number of individuals for all species.
References
- Stack, C. (Ed.) Beetles: Biodiversity, Ecology and Role in the Environment; Nova Science Publishers Inc.: New York, NY, USA, 2015; p. 262. [Google Scholar]
- Marshall, S.A. Beetles: The Natural History and Diversity of Coleoptera; Firefly Books Ltd.: Richmond Hill, ON, Canada, 2018; p. 784. [Google Scholar]
- Racovitza, E.G. Essai sur les problèmes biospéologiques. Arch. Zool. Exp. Gén. 1907, 4, 371–488. [Google Scholar]
- Culver, D.C. Cave Life. Evolution and Ecology; Harvard University Press: Cambridge, MA, USA, 1982; p. 189. [Google Scholar]
- Juberthie, C.; Delay, B.; Bouillon, M. Extension du milieu souterrain en zone non-calcaire: Description d’un nouveau milieu et de son peuplement par les Coléoptéres troglobies. Mém. Biospéol. 1980, 7, 19–52. [Google Scholar]
- Juberthie, C. Le milieu souterrain: Étendue et Composition. Mém. Biospéol. 1983, 10, 17–65. [Google Scholar]
- Pipan, T.; Culver, D.C. Shallow Subterranean Habitats. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: London, UK, 2012; pp. 886–895. [Google Scholar]
- Rendoš, M.; Mock, A.; Jászay, T. Spatial and temporal dynamics of invertebrates dwelling karstic mesovoid shallow substratum of Sivec National Nature Reserve (Slovakia), with emphasis on Coleoptera. Biologia 2012, 67, 1143–1151. [Google Scholar] [CrossRef] [Green Version]
- Nae, I.; Băncilă, R.I. Mesovoid shallow substratum as a biodiversity hotspot for conservation priorities: Analysis of oribatid mite (Acari: Oribatida) fauna. Acarologia 2017, 57, 855–868. [Google Scholar] [CrossRef]
- Ledesma, E.; Jiménez-Valverde, A.; Baquero, E.; Jordana, R.; de Castro, A.; Ortuño, V.M. Arthropod biodiversity patterns point to the Mesovoid Shallow Substratum (MSS) as a climate refugium. Zoology 2020, 141, 125771. [Google Scholar] [CrossRef] [PubMed]
- Skalski, A.W. Origin and composition of underground fauna in Poland. In Proceedings of the 8th International Congress of Speleology, Volumes I & II, Department of Geography and Geology, Western Kentucky University, Bowling Green, KY, USA, 18–24 July 1981. [Google Scholar]
- Kocot-Zalewska, J.; Domagała, P. Terrestrial invertebrate fauna of Polish caves—A summary of 100 years of research. Subterr. Biol. 2020, 33, 45–69. [Google Scholar] [CrossRef] [Green Version]
- Gradziński, M.; Kicińska, D. Caves in Poland. In Polish Caving 2013–2017, Proceedings of the 17th International Congress of Speleology, Sydney, Australia, 23–29 July 2017; Kicińska, D., Ed.; Pracownia Kreatywna Bezliku; Komisja Taternictwa Jaskiniowego Polskiego Związku Alpinizmu: Warszawa, Poland, 2017; pp. 4–6. [Google Scholar]
- Kocot-Zalewska, J.; Woźnica, A.J. Cave-dwelling heleomyzid flies (Diptera: Heleomyzidae) from the Polish caves. Historical overview and new data. Int. J. Speleol. 2021, 50, 203–211. [Google Scholar] [CrossRef]
- Jaskinie Polski. Available online: http://geoportal.pgi.gov.pl/jaskinie_polski (accessed on 27 April 2020).
- Szelerewicz, M.; Górny, A. Jaskinie Wyżyny Krakowsko-Wieluńskiej; Wydawnictwo PTTK Kraj: Warszawa-Kraków, Poland, 1986; p. 200. [Google Scholar]
- Zygmunt, J. Jaskinie Okolic Olsztyna; ZH-U Kontur: Częstochowa, Poland, 2013; p. 316. [Google Scholar]
- Jost, L. Entropy and diversity. Oikos 2006, 113, 363–375. [Google Scholar] [CrossRef]
- Chao, A.; Gotelli, N.J.; Hsieh, T.C.; Sander, E.L.; Ma, K.H.; Colwell, R.K.; Ellison, A.M. Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 2014, 84, 45–67. [Google Scholar] [CrossRef] [Green Version]
- Chao, A.; Kobuta, Y.; Zelený, D.; Chiu, C.H.; Li, C.F.; Kosumuto, B.; Yasuhara, M.; Thorn, S.; Wei, C.L.; Costello, M.J.; et al. Quantifying sample completeness and comparing diversities among assemblages. Ecol. Res. 2020, 35, 292–314. [Google Scholar] [CrossRef]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Kasprzak, K.; Niedbała, W. Wskaźniki biocenotyczne stosowane przy porządkowaniu i analizie danych w badaniach ilościowych. In Metody Stosowane w Zoologii Gleby; Górny, M., Grüm, L., Eds.; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1981; pp. 397–416. [Google Scholar]
- Biesiadka, E.; Kowalik, W. Water miters (Hydracarina) of the Western Bieszczady Mountains. I. Stagnant waters. Acta Hydrobiol. 1980, 22, 279–298. [Google Scholar]
- Szymczakowski, W. Klucze Do Oznaczania Owadów Polski. Część XIX: Chrząszcze—Coleoptera. Zeszyt 13: Catopidae; Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1961; p. 69. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Biegaczowate—Carabidae, Część 1. Katalog Fauny Polski, XXIII (2); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1973; p. 430. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Biegaczowate—Carabidae, Część 2. Katalog Fauny Polski, XXIII (3); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1974; p. 430. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Adephaga prócz Carabidae, Myxophaga, Polyphaga: Hydrophiloidea. Katalog Fauny Polski, XXIII (4); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1976; p. 307. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Histeroidea i Staphylinoidea prócz Staphylinidae. Katalog Fauny Polski, XXIII (5); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1978; p. 356. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 1. Katalog Fauny Polski, XXIII (6); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1979; p. 309. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 2. Katalog Fauny Polski, XXIII (7); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1980; p. 271. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Kusakowate—Staphylinidae, Część 3: Aleocharinae. Katalog Fauny Polski, XXIII (8); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1981; p. 329. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Scarabaeoidea, Dascilloidea, Byrrhoidea i Parnoidea. Katalog Fauny Polski, XXIII (9); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1983; p. 394. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Buprestoidea, Elateroidea i Cantharoidea. Katalog Fauny Polski, XXIII (10); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1985; p. 400. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 1. Katalog Fauny Polski, XXIII (12); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1986; p. 265. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 2. Katalog Fauny Polski, XXIII (13); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1986; p. 277. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cucujoidea, Część 3. Katalog Fauny Polski, XXIII (14); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1987; p. 308. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Cerambycidae i Bruchidae. Katalog Fauny Polski, XXIII (15); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1990; p. 311. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowcowate prócz ryjkowców—Curculionioidea prócz Curculionidae. Katalog Fauny Polski, XXIII (18); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1992; p. 323. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 1. Katalog Fauny Polski, XXIII (19); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1993; p. 304. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 2. Katalog Fauny Polski, XXIII (20); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1995; p. 310. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Ryjkowce—Curculionidae, Część 3. Katalog Fauny Polski, XXIII (21); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 1997; p. 307. [Google Scholar]
- Burakowski, B.; Mroczkowski, M.; Stefańska, J. Chrząszcze—Coleoptera. Uzupełnienia Tomów 2–21. Katalog Fauny Polski, XXIII (22); Państwowe Wydawnictwo Naukowe: Warszawa, Poland, 2000; p. 252. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 1; Goecke & Evers: Krefeld, Germany, 1989; p. 440. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 2; Goecke & Evers: Krefeld, Germany, 1989; p. 382. [Google Scholar]
- Koch, K. Die Käfer Mitteleuropas. Ökologie. Band 3; Goecke & Evers: Krefeld, Germany, 1992; p. 389. [Google Scholar]
- Crouau-Roy, B.; Crouau-Roy, Y.; Ferre, C. Dynamic and temporal structure of the troglobitic beetle Speonomus hydrophilus (Coleoptera: Bathysciinae). Ecography 1992, 15, 12–18. [Google Scholar] [CrossRef]
- Smoleński, M. Kusakowate (Coleoptera: Staphylinidae) borów bażynowych (Empetro-nigri Pinetum) Mierzei Łebskiej w Słowińskim Parku Narodowym. Wiad. Entomol. 2000, 18, 207–222. [Google Scholar]
- Päivinen, J.; Ahlroth, P.; Kaitala, V. Ant-associated beetles of Fennoscandia and Denmark. Entomol. Fenn. 2002, 13, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Ruta, R.; Jałoszyński, P.; Konwerski, S. Nowe stanowiska gnilików (Coleoptera: Histeridae) w Polsce. Część 1. Onthophilinae —Dendrophilinae. Wiad. Entomol. 2004, 23, 13–20. [Google Scholar]
- Tréfás, H.; Van Lenteren, J. Egg-laying-site preferences of Pterostichus melanarius in mono- and intercrops. Bull. Insectology 2008, 61, 225–231. [Google Scholar]
- Shockley, F.W.; Tomaszewska, K.W.; McHugh, J.V. Review of the Natural History of the Handsome Fungus Beetles (Coleoptera: Cucujoidea: Endomychidae). Insecta Mundi 2009, 72, 1–24. [Google Scholar]
- Huruk, S.; Huruk, A. Zgrupowania biegaczowatych (Coleoptera: Carabidae) borów sosnowych oraz jodłowych w Górach Świętokrzyskich. Wiad. Entomol. 2010, 29, 247–258. [Google Scholar]
- Huruk, S.; Barševskis, A. Charakterystyka Biegaczowatych (Coleoptera, Carabidae) Gór Świętokrzyskich; Wydawnictwo Emporium: Kielce, Poland, 2014; p. 219. [Google Scholar]
- Szujecki, A. Kusakowate (Staphylinidae) Lasów Polski. Aspekt Różnorodności i Monitoringu Zooindykacyjnego; Lasy Państwowe: Warszawa, Poland, 2017; p. 150. [Google Scholar]
- Sket, B. Can we agree on an ecological classification of subterranean animals? J. Nat. Hist. 2008, 42, 1549–1563. [Google Scholar] [CrossRef]
- Kowalski, K. Fauna jaskiń Tatr Polskich. Ochr. Przyr. 1955, 23, 283–333. [Google Scholar]
- Skalski, A.W. Materiały do znajomości bezkręgowców jaskiń Wyżyny Krakowsko-Częstochowskiej. Roczn. Muz. Częstochowskiego 1973, 3, 161–200. [Google Scholar]
- Skalski, A.W. Charakterystyka fauny podziemnej Wyżyny Krakowsko-Częstochowskiej. Roczn. Muz. Okręg. Częstochowie 1981, 5, 51–60. [Google Scholar]
- Kováč, Ľ.; Mock, A.; Ľuptáčik, P.; Košel, V.; Fenďa, P.; Svatoň, J.; Mašán, P. Terrestrial Arthropods of the Domica Cave system and the Ardovská Cave (Slovak Karst)—Principal microhabitats and diversity. In Contributions to Soil Zoology in Central Europe, Proceedings of the 7th Central European Workshop on Soil Zoology, České Budějovice, Czech Republic, 14–16 April 2003; Tajovský, K., Schlaghamerský, J., Pižl, V., Eds.; Institute of Soil Biology, Academy of Sciences of the Czech Republic: České Budějovice, Czech Republic, 2005; pp. 61–70. [Google Scholar]
- Weber, D. Die Höhlenfauna Luxemburgs. Ferrantia 69; Musée National d’Histoire Naturelle: Luxembourg, 2013; p. 408. [Google Scholar]
- Zaenker, S.; Weber, D.; Weigand, A. Liste der cavernicolen Tierarten Deutschlands mit Einschluss der Grundwasserfauna (Version 1.9). Available online: https://www.hoehlentier.de/taxa.pdf (accessed on 27 April 2020).
- Arndt, W. Beitrag zur Kenntnis der Höhlenfauna. Ergebnis einer faunistischen Untersuchung der Höhlen Schlesiens. Zool. Anz. 1921, 52, 310–315. [Google Scholar]
- Arndt, W. Speläobiologische Untersuchungen in Schlesien. Speläologisch. Jahrb. 1923, 4, 95–114. [Google Scholar]
- Demel, K. Fauna jaskiń Ojcowskich. Sprawozd. Posiedz. Tow. Nauk. Warsz. Wydz. Nauk Mat.-Przyr. 1918, 11, 623–659. [Google Scholar]
- Pax, F.; Maschke, K. Höhlenfauna des Glatzer Schneeberges. Die rezente Metazoenfauna. Beitr. Biol. Glatzer Schneeberges 1935, 1, 4–72. [Google Scholar]
- Moldovan, E. Beetles. In Encyclopedia of Caves; White, W.B., Culver, D.C., Eds.; Academic Press: London, UK, 2012; pp. 96–107. [Google Scholar]
- Buse, J.; Alexander, K.N.A.; Ranius, T.; Assmann, T. Saproxylic Beetles—Their Role and Diversity in European Woodland and Tree Habitats. In Proceedings of the 5th Symposium and Workshop on the Conservation of Saproxylic Beetles, Lüneberg, Germany, 14–16 June 2008; Pensoft Publishers: Sofia, Bulgaria, 2009; p. 236. [Google Scholar]
- Plachter, H.; Plachter, J. Ökologische Studien zur terrestrischen Höhlenfauna Südduetschlands. Zoologica 1988, 139, 1–67. [Google Scholar]
- Szymczakowski, W. Catopidae (Coleoptera) des grottes dans les Sokole Góry près de Częstochowa. Acta Zool. Cracov. 1957, 1, 65–115. [Google Scholar]
- Nitzu, E. The Cholevinae of Romania (exclusive of Leptodirini) (Coleoptera, Leiodidae) with special reference to the hypogeal records. Zootaxa 2013, 3620, 351–378. [Google Scholar] [CrossRef] [PubMed]
- Mazur, A. Omalium validum KRAATZ, 1858 (Coleoptera, Staphylinidae) im Bielengebirge (poln. Góry Bialskie) und Glatzer Schneegebirge (poln. Masyw Śnieżnika). Przyr. Sudet. 2007, 10, 97–100. [Google Scholar]
- Mlejnek, R.; Hamet, A.; Růžička, J. Brouci (Coleoptera) v jeskyních a propastech České republiky: Beetles (Coleoptera) in caves and chasms of the Czech Republic. Acta Speleol. 2015, 6, 1–111. [Google Scholar]
- Nitzu, E. Microhabitats—Integrative environmental factors for species communities of Coleoptera in the karst landscape. Biologia 2021, 76, 1775–1783. [Google Scholar] [CrossRef]
- Tobin, B.W.; Hutchins, B.T.; Schwartz, B.F. Spatial and temporal changes in invertebrate assemblage structure from the entrance to deep-cave zone of a temperate marble cave. Int. J. Speleol. 2013, 42, 203–214. [Google Scholar] [CrossRef] [Green Version]
- Lunghi, E.; Manenti, R.; Ficetola, G.F. Do cave features affect underground habitat exploitation by non-troglobite species? Acta Oecol. 2014, 55, 29–35. [Google Scholar] [CrossRef]
- Killing-Heinze, M.; Pflitsch, A.; Furian, W.; Allison, S. The importance of air temperature as a key parameter to identify climatic processes inside Carlsbad Cavern, New Mexico, USA. J. Caves Karst Stud. 2017, 79, 153–167. [Google Scholar] [CrossRef]
- Mayoral, J.; Barranco, P.; Fernandez-Cortes, A. First biological data, associated fauna, and microclimate preferences of the enigmatic cave-dwelling beetle Dalyat mirabilis Mateu, 2002 (Coleoptera, Carabidae). Subterr. Biol. 2022, 44, 103–121. [Google Scholar] [CrossRef]
- Skalski, A.W. Experimental acclimatization of Speonomus hydrophilus (Jeannel 1907) (Coleoptera, Catopidae, Bathysciinae) in Poland. Mem. Biospeol. 1994, 21, 127–131. [Google Scholar]
- Kocot-Zalewska, J. Speonomus hydrophilus (Jeannel 1907) w Jaskini Towarnej. In Materiały 50 Sympozjum Speleologicznego, Proceedings of the 50th Speleological Symposium, Kielce, Poland, 20–23 October 2016; Urban, J., Ed.; Sekcja Speleologiczna Polskiego Towarzystwa Przyrodników im. M. Kopernika: Kraków, Poland, 2016; p. 124. [Google Scholar]
- Kocot-Zalewska, J.; Domagała, P.J.; Lis, B. Living in isolation for almost 40 years: Molecular divergence of the 28S rDNA and COI sequences between French and Polish populations of the cave beetle Speonomus normandi hydrophilus (Jeannel, 1907). Subterr. Biol. 2021, 37, 75–88. [Google Scholar] [CrossRef]
- Howarth, F.G. Ecology of cave arthropods. Ann. Rev. Entomol. 1983, 28, 365–389. [Google Scholar] [CrossRef]
- Poulson, T.L. Food sources. In Encyclopedia of Caves; Culver, D.C., White, W.B., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2005; pp. 255–264. [Google Scholar]
- Culver, D.C.; Pipan, T. Shallow Subterranean Habitats: Ecology, Evolution, and Conservation; Oxford University Press: Oxford, UK, 2014; p. 258. [Google Scholar]
- Prous, X.; Ferreira, R.L.; Jacobi, C.M. The entrance as a complex ecotone in a Neotropical cave. Int. J. Speleol. 2015, 44, 177–189. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cave | S0 | S1 | S2 | S3 | S4 | S5 |
|---|---|---|---|---|---|---|
| W Zielonej Górze | 22 | 34 | 11 | 7 | ---- | ---- |
| Towarna | 53 | 24 | 9 | 8 | ---- | ---- |
| Pod Sokolą | 13 | 35 | 11 | 5 | 3 | ---- |
| Kroczycka | 17 | 10 | ---- | 5 | 11 | 7 |
| Psia | 20 | 10 | 8 | 1 | ---- | ---- |
| Zegar | 23 | 33 | 8 | 11 | ---- | ---- |
| Cave | Number of Species | Shannon Diversity Index | Simpson Diversity Index |
|---|---|---|---|
| W Zielonej Górze | 38 | 7.614086359 | 1.375137514 |
| Towarna | 27 | 1.447734615 | 7.692307692 |
| Pod Sokolą | 42 | 5.053090317 | 1.497005988 |
| Psia | 17 | 8.758284041 | 2.949852507 |
| Zegar | 37 | 15.95863401 | 1.051524711 |
| Kroczycka | 21 | 8.846306259 | 1.112718371 |
| Cave | Eudominant | Dominant |
|---|---|---|
| W Zielonej Górze | Cryptophagus distinguendus J. Sturm, 1845 (25.76%) Cryptophagus punctipennis Ch.N.F. Brisout de Barneville, 1863 (28.46%) Quedius mesomelinus mesomelinus Marsham, 1802 (12.30%) | |
| Towarna | Speonomus normandi hydrophilus (Jeannel, 1907) (92.86%) | |
| Pod Sokolą | Choleva lederiana gracilenta Szymczakowski, 1957 (55.6%) Catops picipes (Fabricius, 1787) (10.97%) | Catops tristis infernus Szymczakowski, 1957 (9%). |
| Kroczycka | Quedius mesomelinus mesomelinus Marsham, 1802 (22.2%) Othius subuliformis Stephens, 1833 (16.65%) Cryptophagus distinguendus J. Sturm, 1845 (12.5%). | Choleva paskoviensis E. Reitter, 1913 (8.3%) Proteinus brachypterus (Fabricius, 1792) (6.9%). |
| Psia | Omalium caesum Gravenhorst, 1806 (23.8%) Choleva cisteloides (Frölich, 1799) (14.28%). | |
| Zegar | Catops picipes (Fabricius, 1787) (13.56%) Omalium validum Kraatz, 1857 (10.17%) | Omalium caesum Gravenhorst, 1806 (8.47%), Choleva glauca Britten, 1918 (5.1%) Lesteva longoelytrata (Goeze, 1777) (5.1%) Quedius mesomelinus mesolmelinus Marsham, 1802 (5.1%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kocot-Zalewska, J.; Lis, B. Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland. Diversity 2023, 15, 345. https://doi.org/10.3390/d15030345
Kocot-Zalewska J, Lis B. Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland. Diversity. 2023; 15(3):345. https://doi.org/10.3390/d15030345
Chicago/Turabian StyleKocot-Zalewska, Joanna, and Barbara Lis. 2023. "Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland" Diversity 15, no. 3: 345. https://doi.org/10.3390/d15030345
APA StyleKocot-Zalewska, J., & Lis, B. (2023). Species Composition and Structure of Beetle Associations in Caves of the Częstochowa Upland, Poland. Diversity, 15(3), 345. https://doi.org/10.3390/d15030345