
On New Species of Three Genera of Zosimeidae Seifried, 2003 (Copepoda: Harpacticoida) from the Deep Sea of the Gulf of California and Gulf of Mexico, with Notes on the Phylogeny of the Family and on the Species Groups of Zosime Boeck, 1873 †


Abstract
:1. Introduction
2. Materials and Methods
2.1. Field and Laboratory Work
2.2. Taxonomic and Descriptive Work
2.3. Phylogenetics
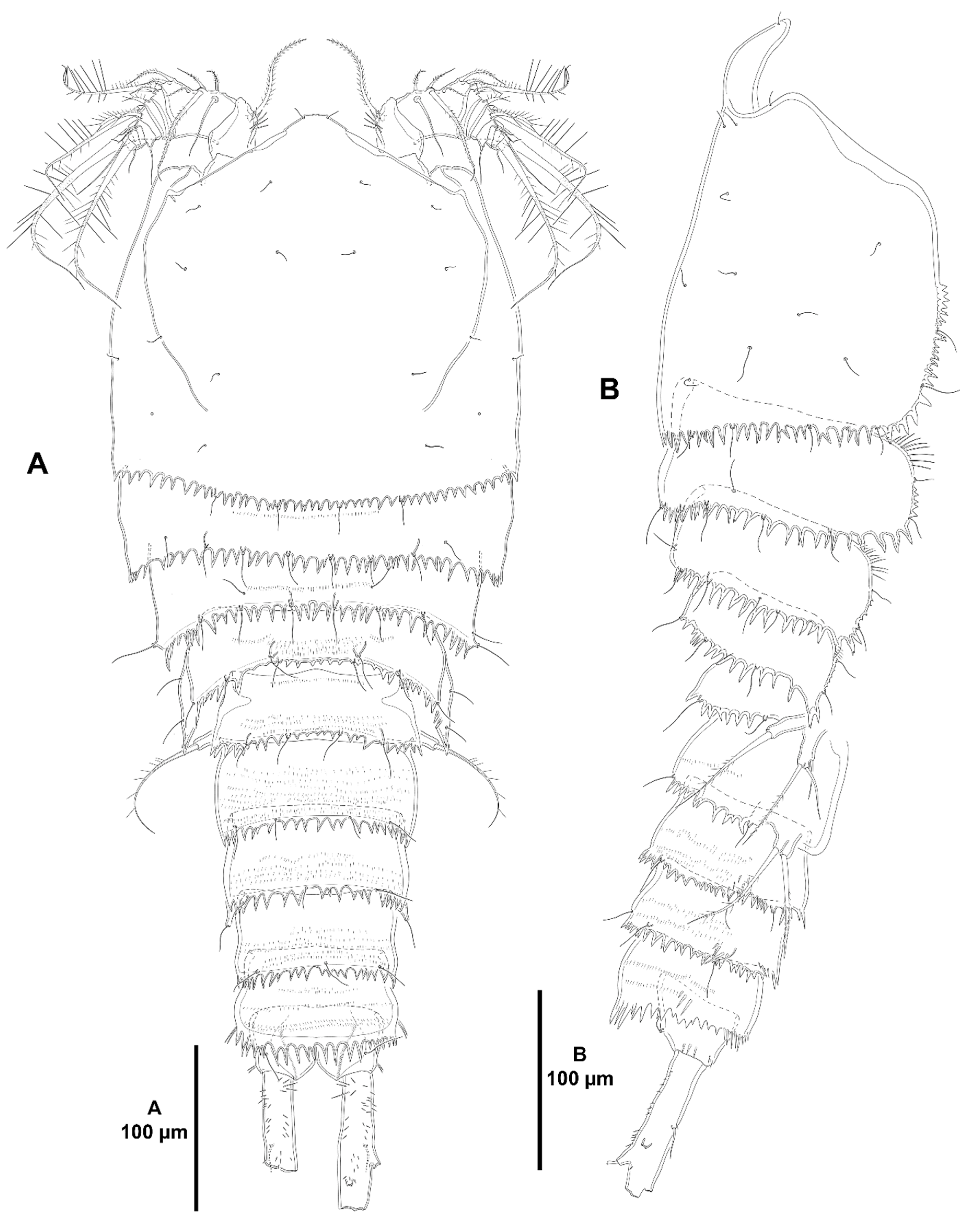
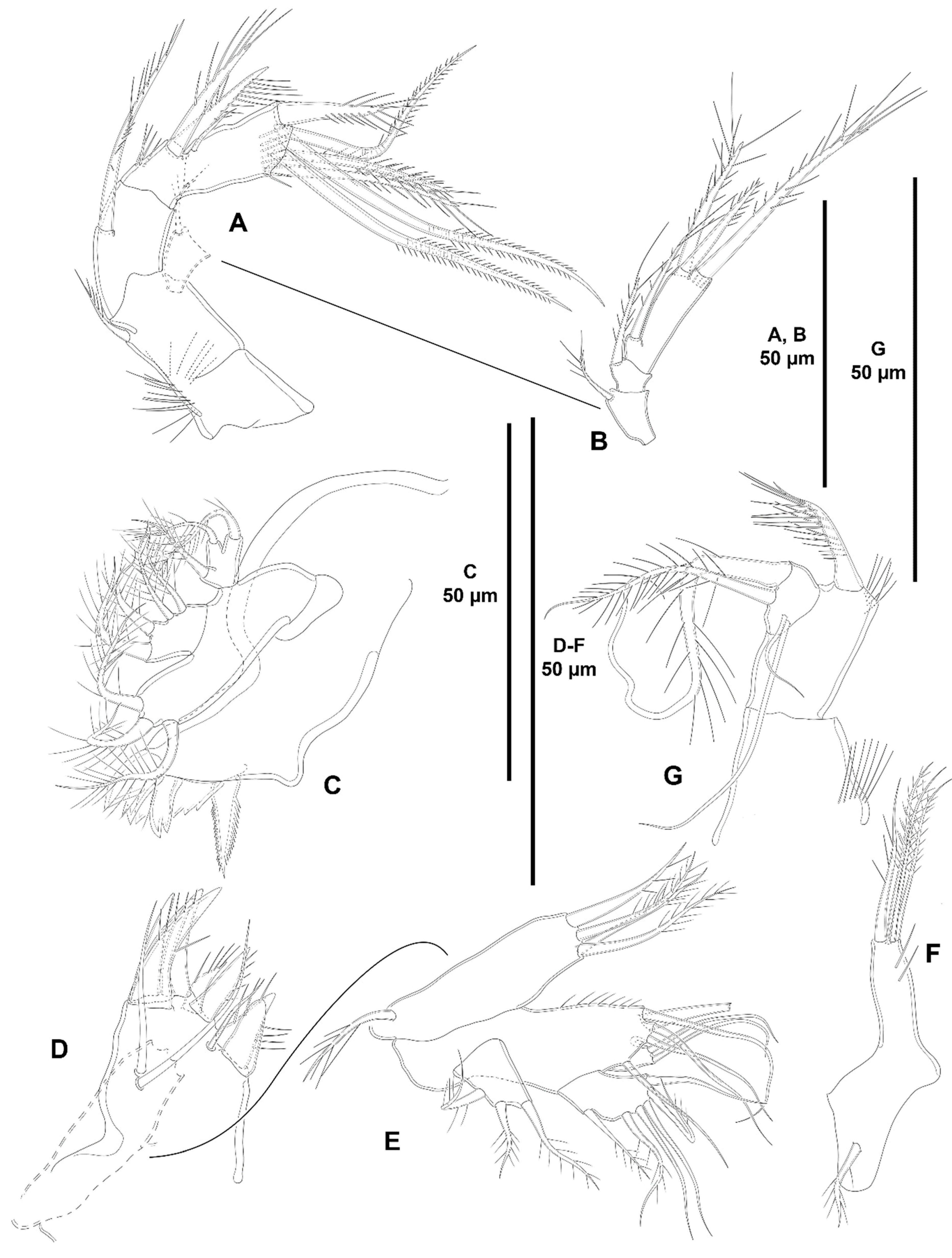
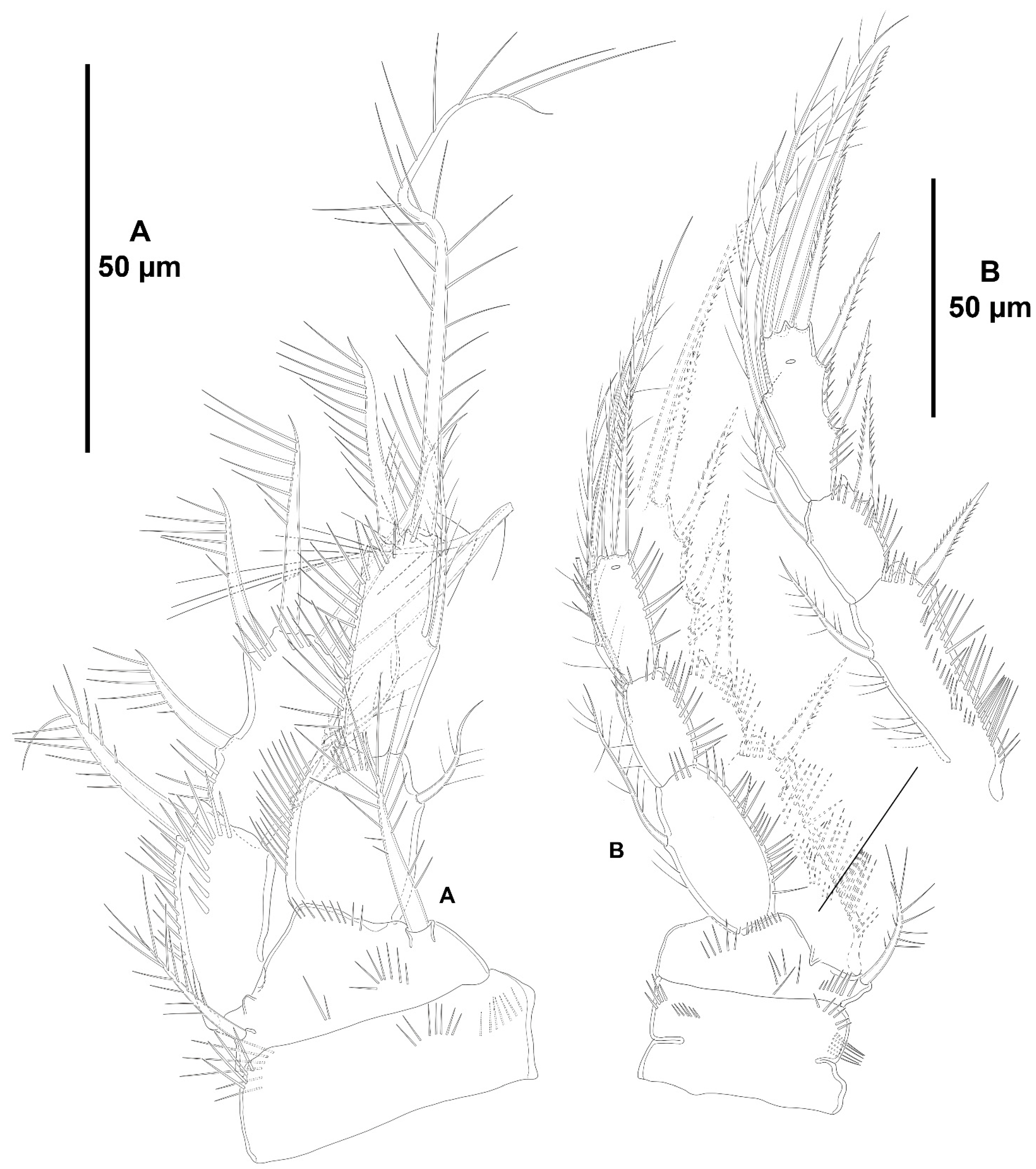
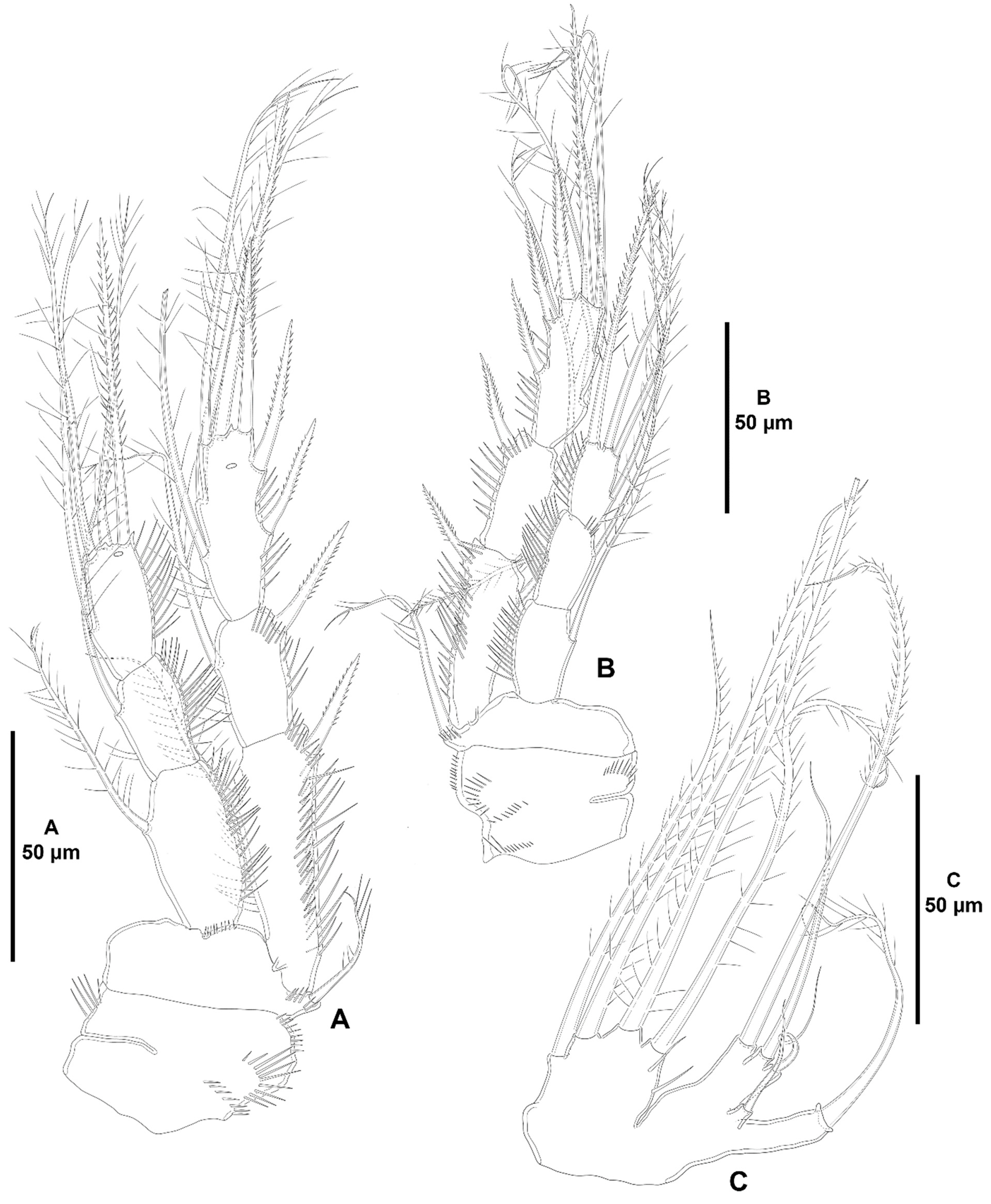
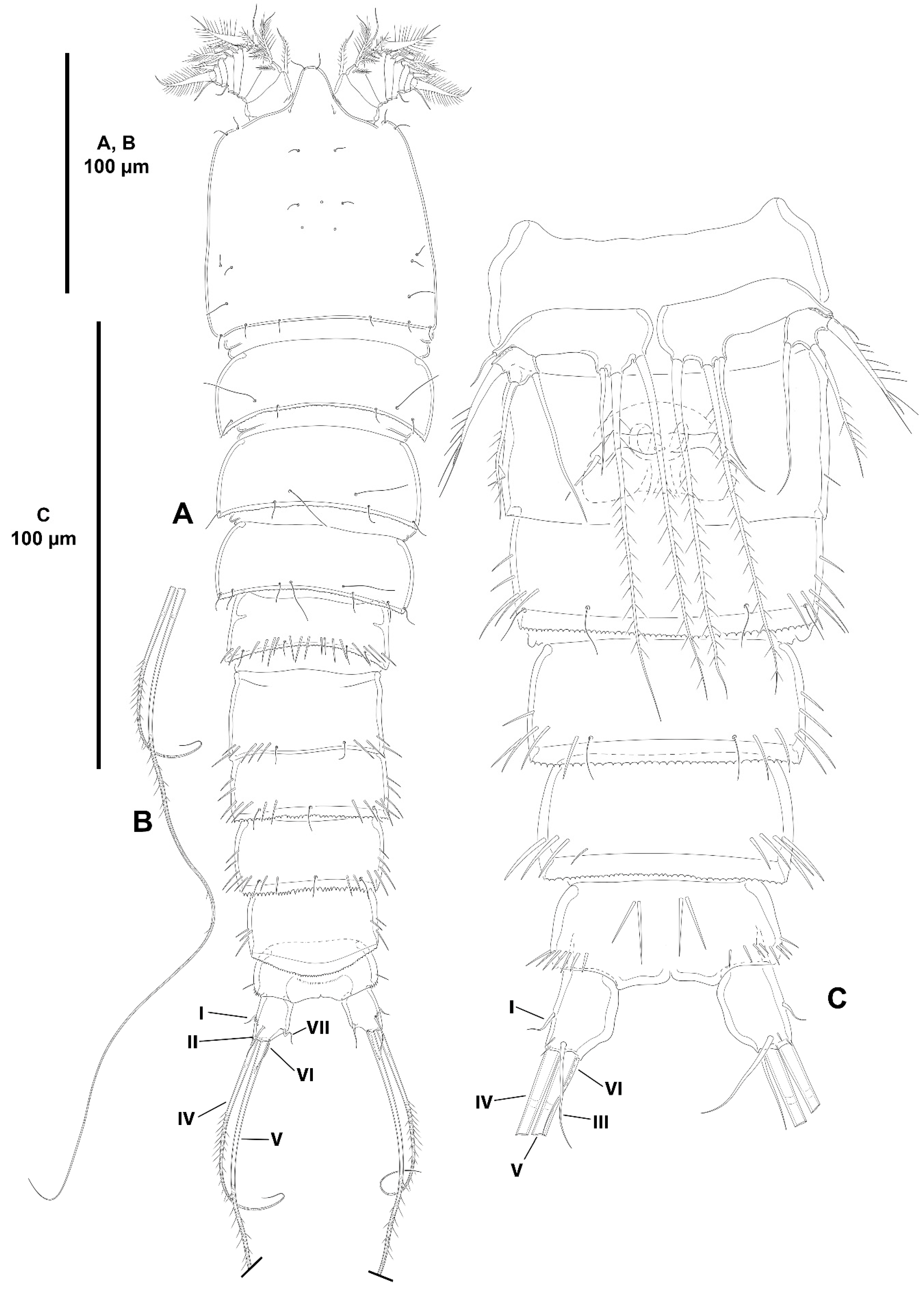
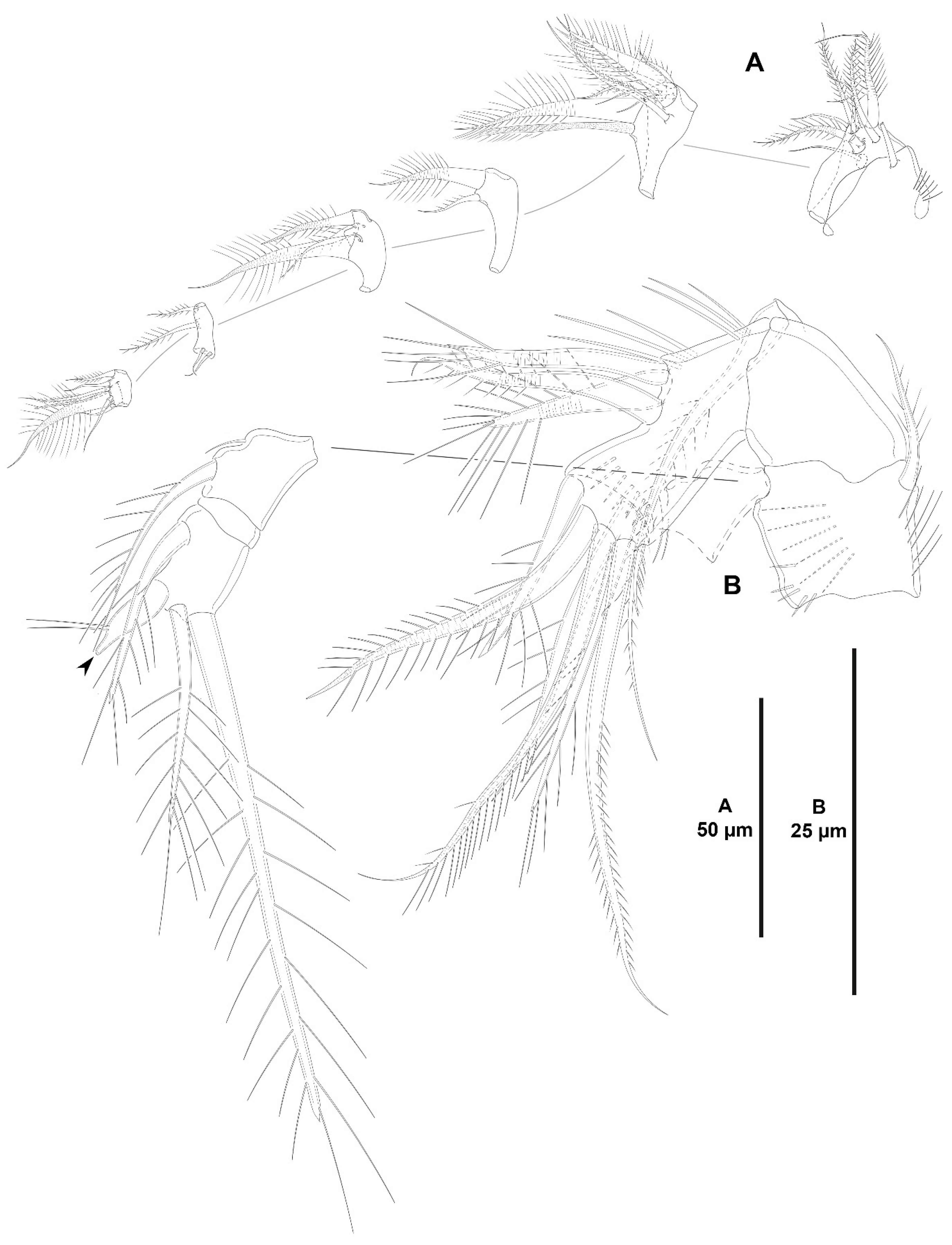
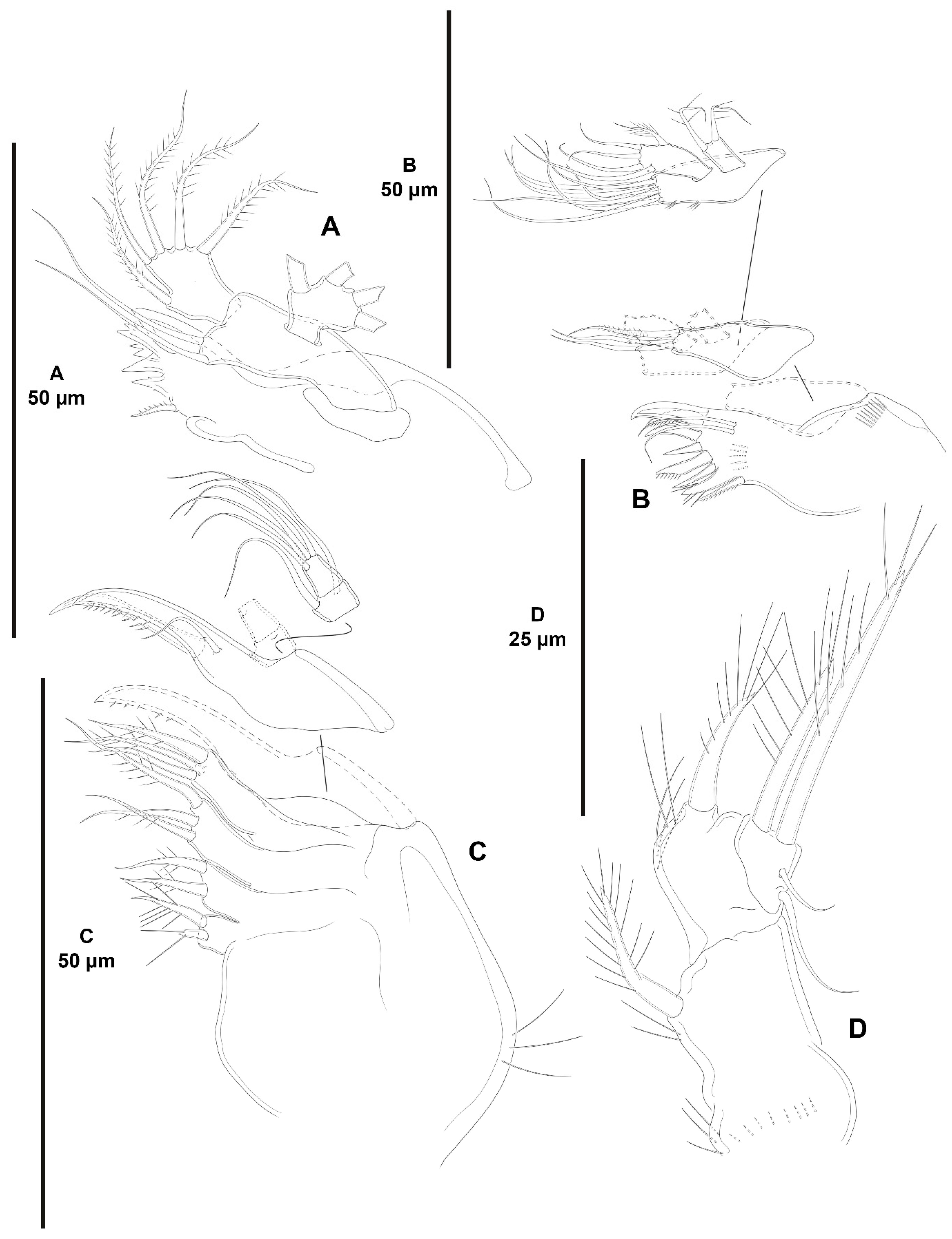
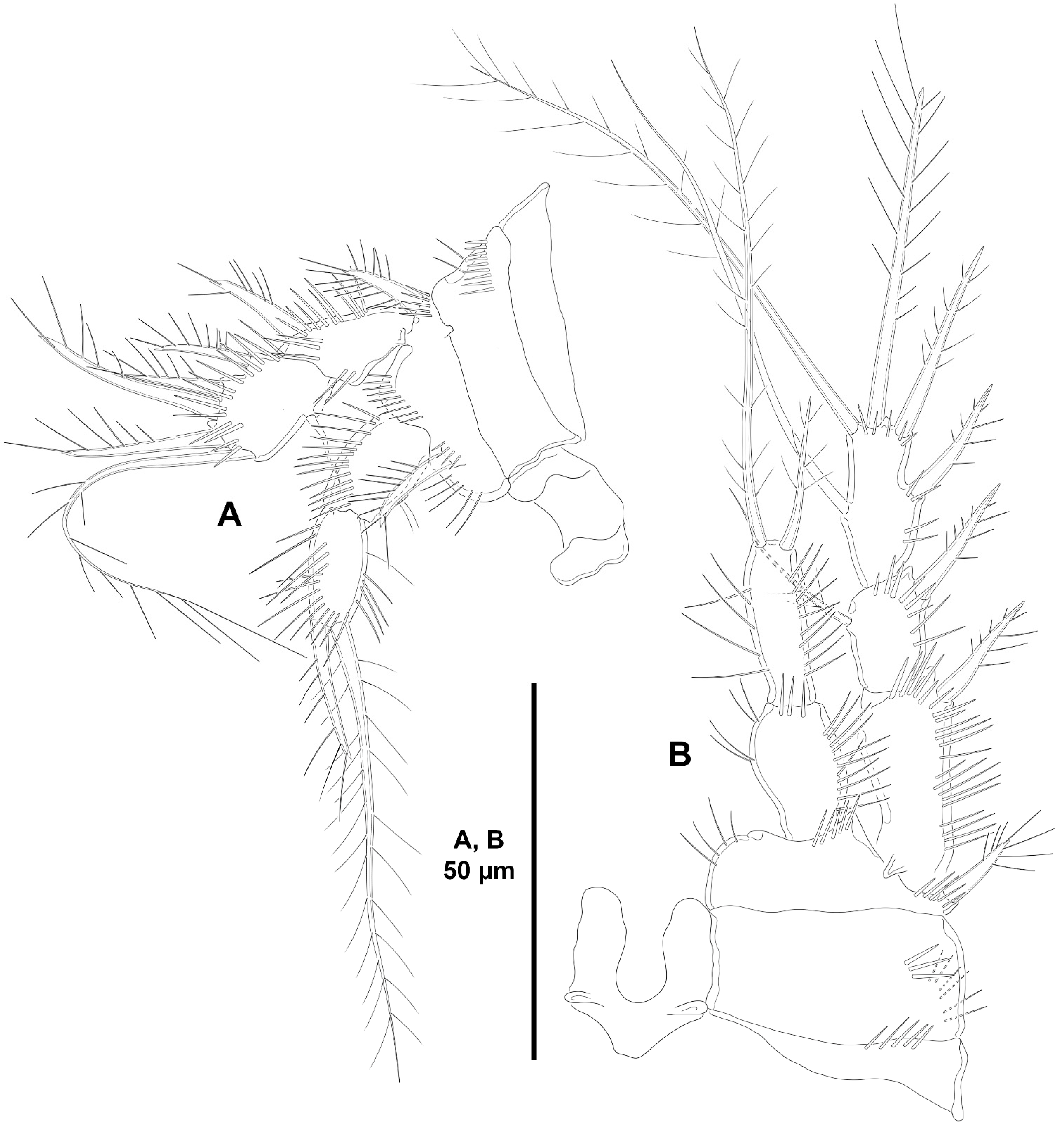
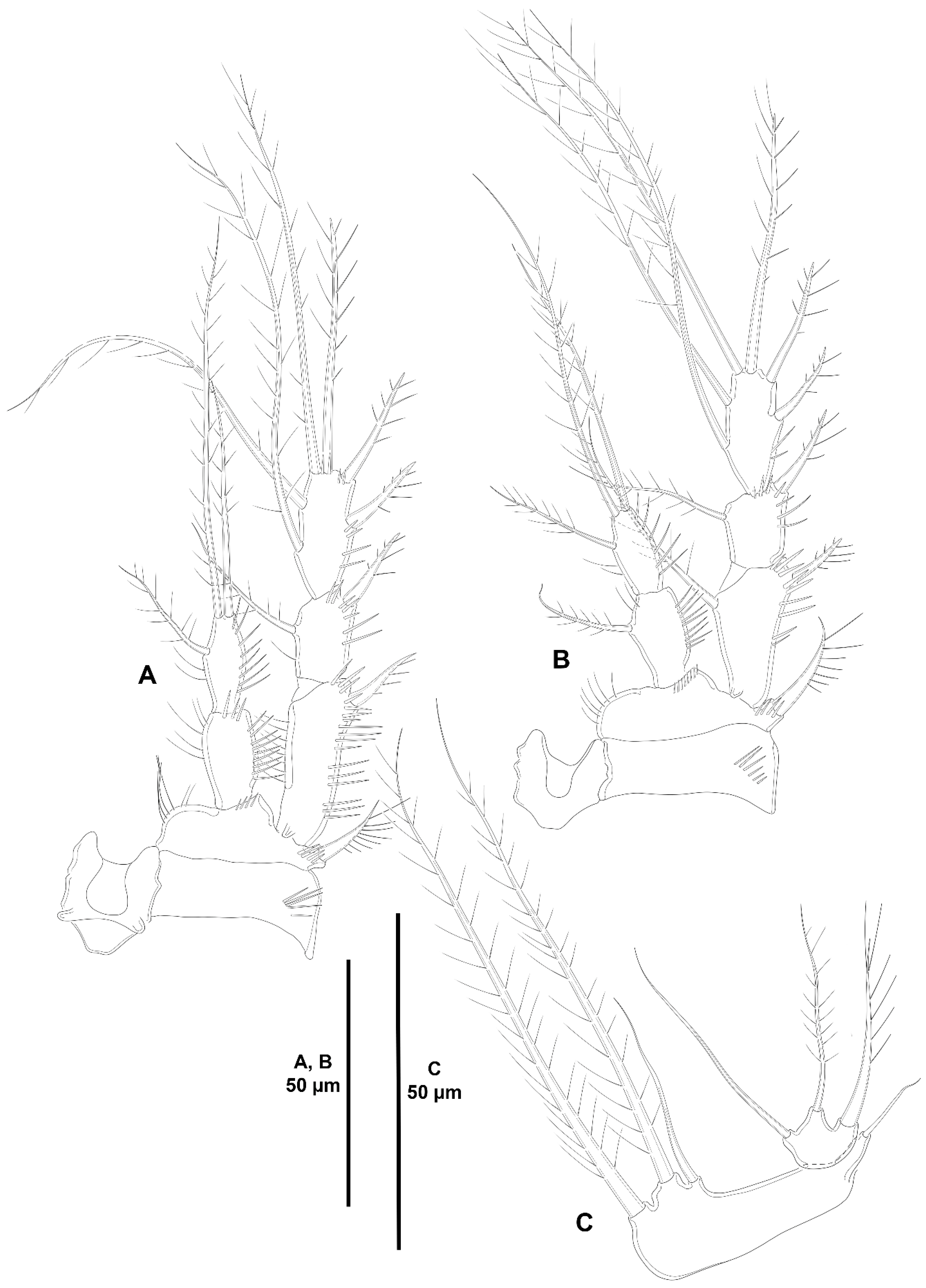
3. Results
3.1. Systematics
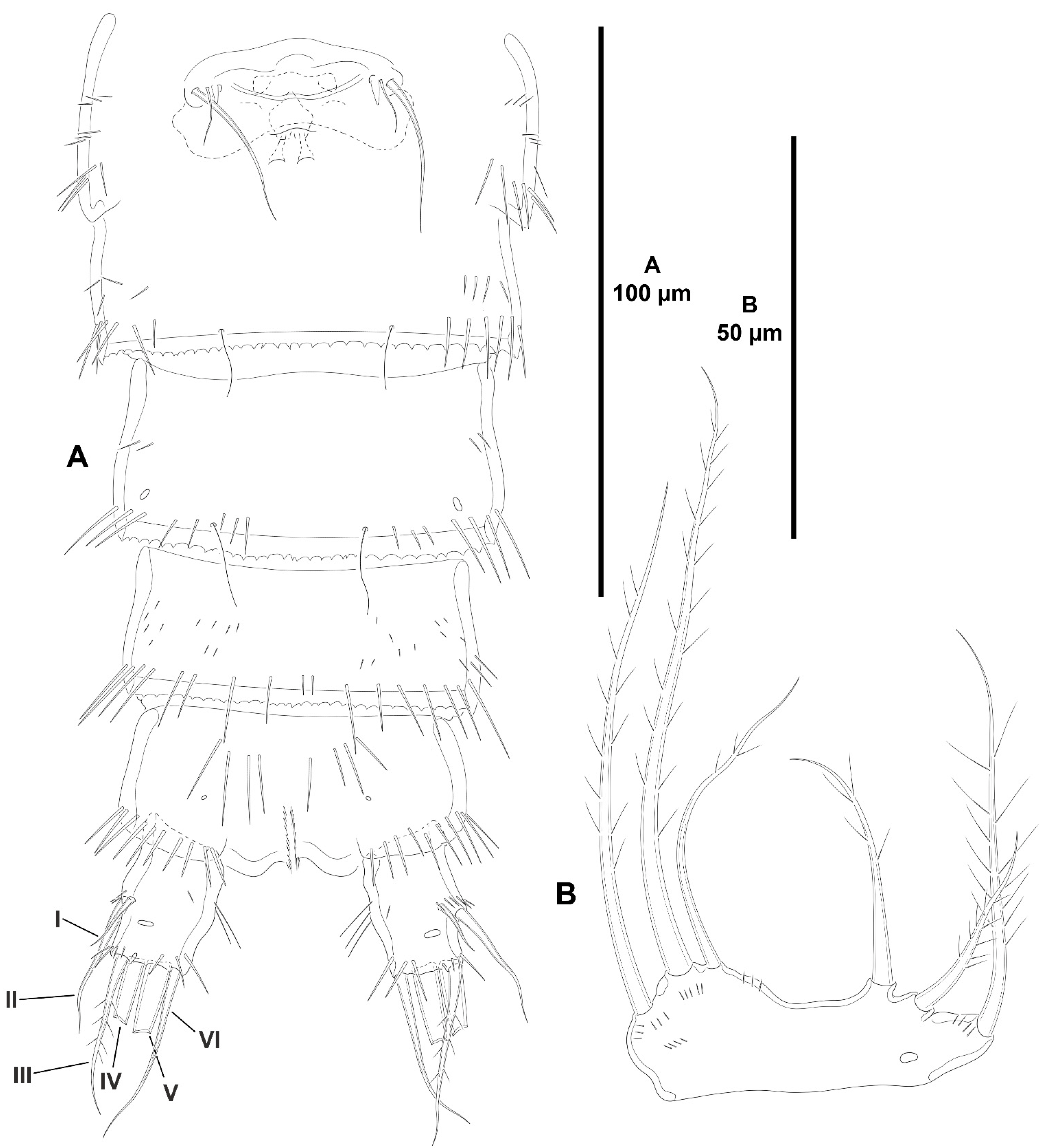
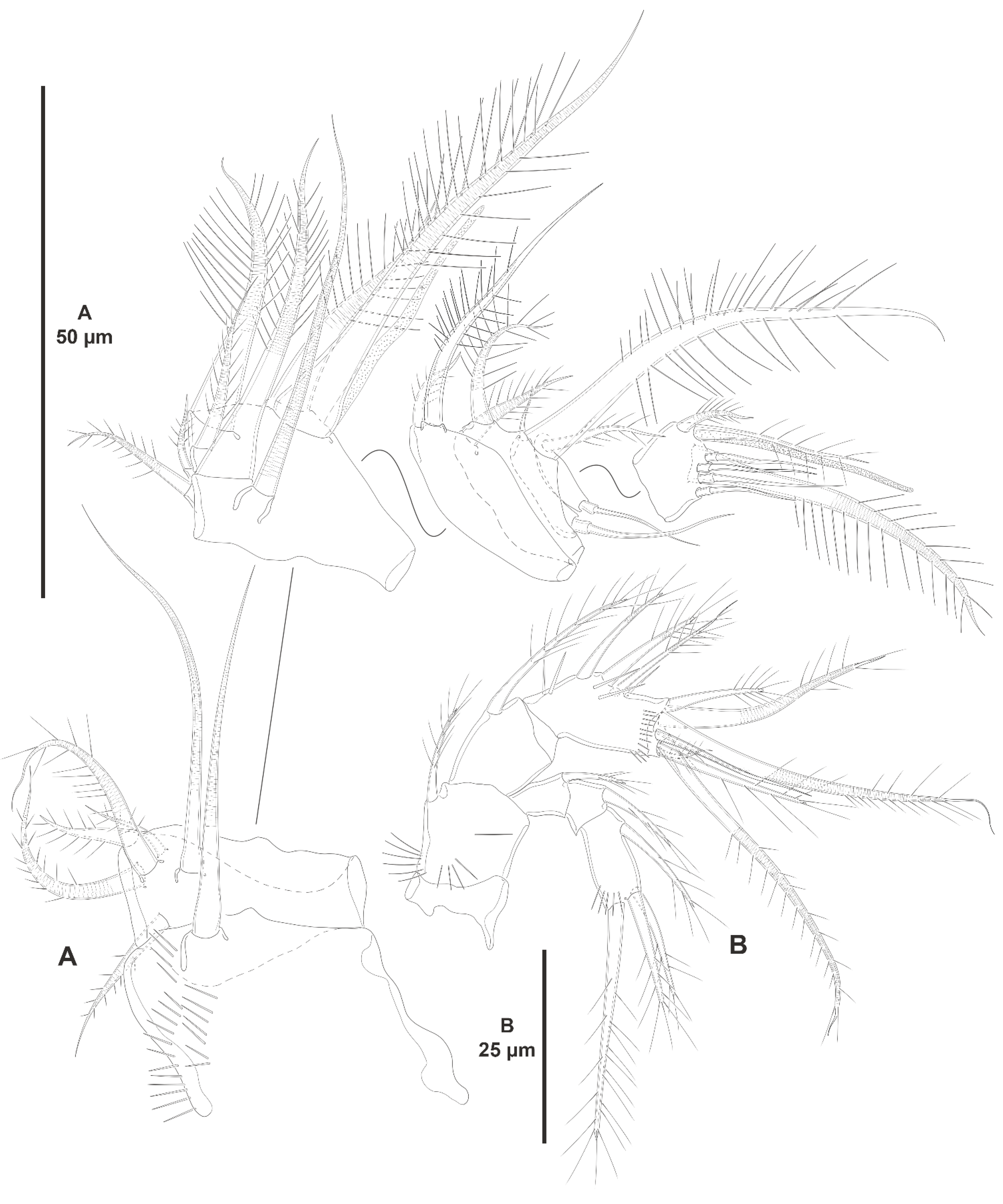
3.1.1. Genus Zosime Boeck, 1873
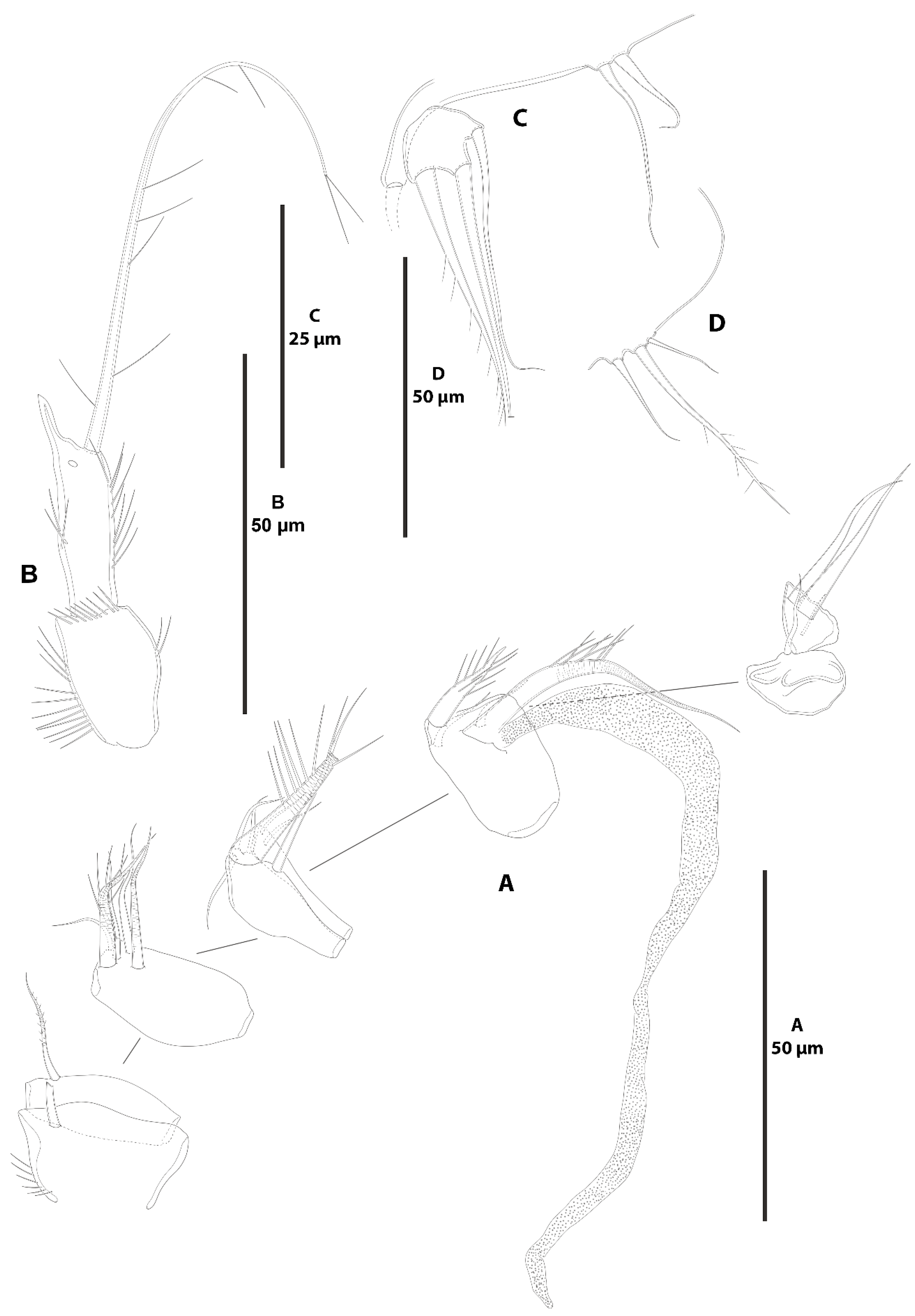
- Zosime xiximi sp. nov.
- Zosime mexicana sp. nov.
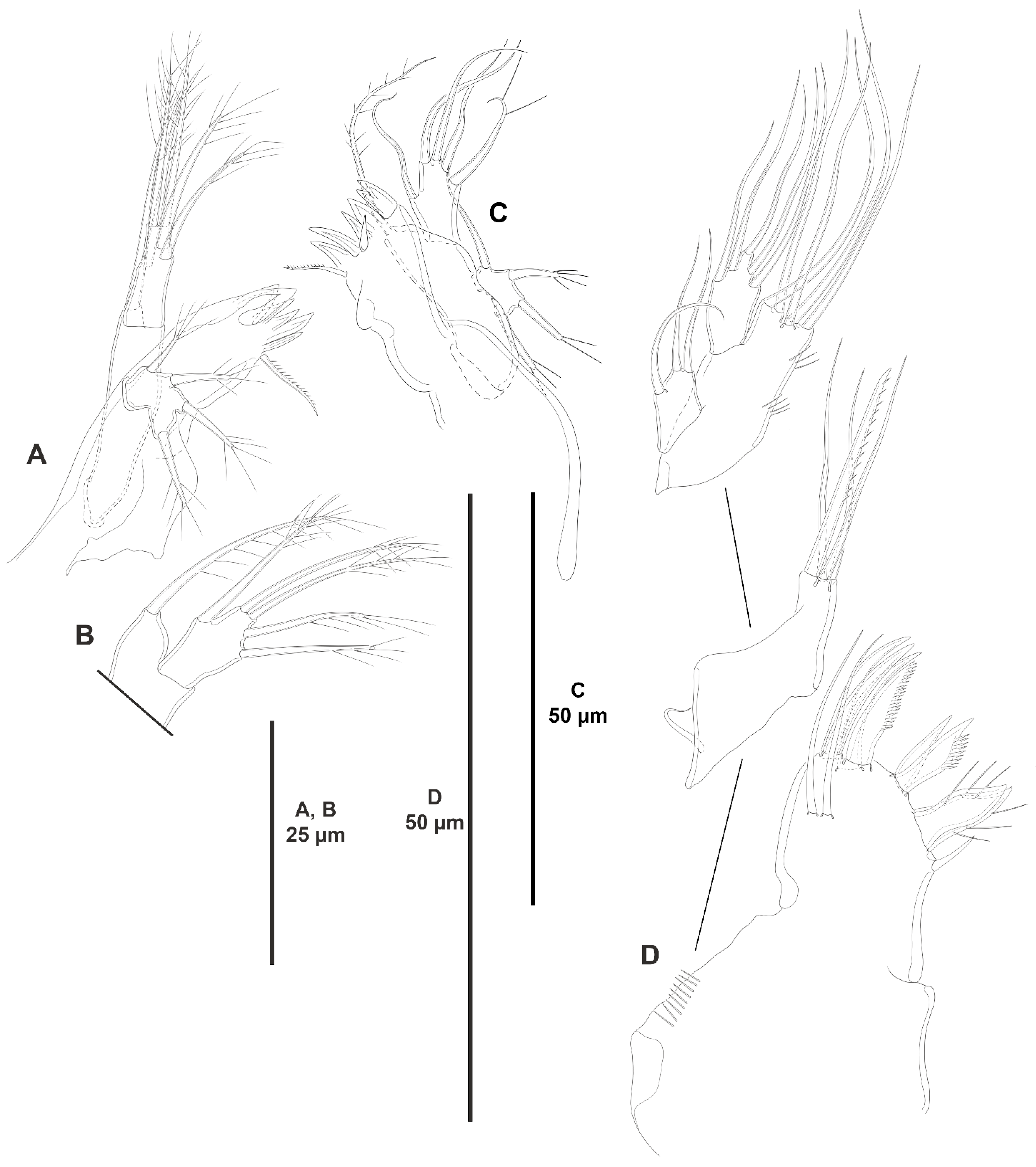
3.1.2. Genus Peresime Dinet, 1974
- Peresime pryocta sp. nov.
3.1.3. Genus Heterozosime Kim and Lee, 2021
- lateral armature o A2 ENP2 composed of two spines and one reduced short seta
- middle apical seta of P4 ENP3 well developed or reduced to a small slender seta
- armature formula of P1–P5 as follows:
| P1 | P2 | P3 | P4 | P5 | |
| EXP | I-0;III,II,2 or I-0;III,2,2 | I-1;I-1;II,I1,2 or I-1;I-1;III;I1,2 | I-1;I-1;II,I1,2 or I-1;I-1;III, I1,2 | I-1;I-1;II,I1,2 or I-1;I-1;III;I1,2 | 3+1 ss |
| ENP | 0-1;I,I,2 or 0-1;1,1,1 | 0-1;0-1;I,2,1♀ 0-1;0,Apo1,1♂ | 0-1;0,1;I,2,1 | 0-1;0-1;I,2,1 | 2 or 4♀ 2♂ |
| Apo, apophysis; ss, surface seta | |||||
- Heterozosime klemie sp. nov.
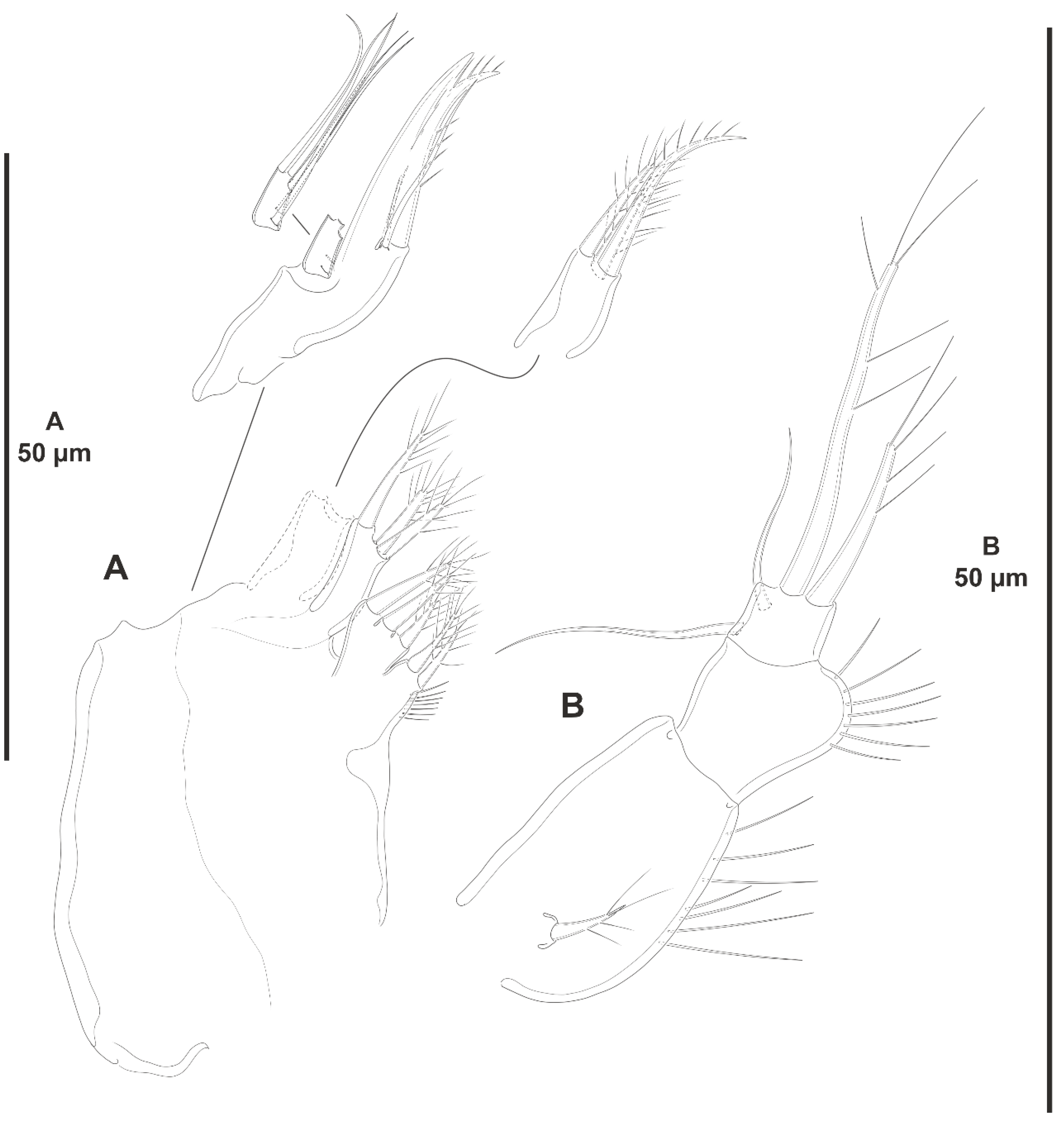
3.1.4. Genus Acritozosime Kim and Lee, 2021
- (i).
- lateral armature o A2 ENP2 composed of two spines and one well-developed seta
- (ii).
- syncoxa of maxilliped with one or two setae
- (iii).
- maxilliped geniculate or non-geniculate
- (iv).
- armature formula of P1–P5 as follows:
| P1 | P2 | P3 | P4 | P5 | |
| EXP | I-0;III,I1,1 | I-1;I-1;II,I1,1 or I-0;I-1;II;I1,1 | I-1;I-1;II,I1,2 or I-0;I-1;II, I1,2 | I-1;I-1;II,I1,2 | 3 |
| ENP | 0-0;0,I1,0 | 0-0;0,I1,0♀ 0-0;0,Apo1,0♂ | 0-0;0,2,1 | 0-1;0,2,1 | 3♀ 2♂ |
| Apo, apophysis | |||||
- Acritozosime sigsbeeiensis sp. nov.
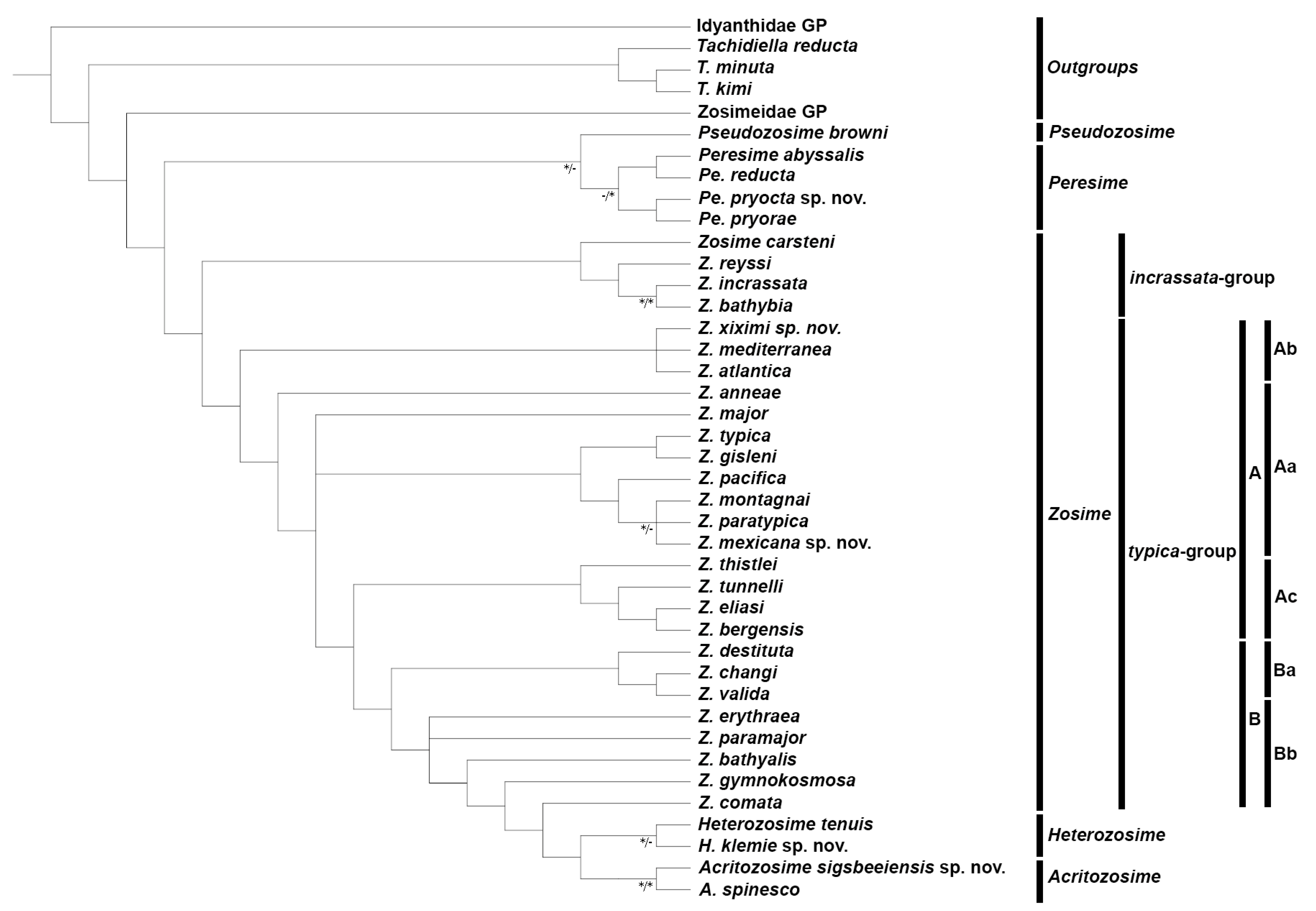
3.2. Phylogenetics
4. Discussion
4.1. Phylogenetic Analysis
- −
- Characters 2–5 (fusion of segments of the female antennule; see also Table S2). Seifried (2003) proposed the ground pattern of the female antennule of the Idyanthidimorpha and Idyanthidae with nine segments and armature formulae I-[1], II-[11], III-[10], IV-[4+ae], V-[3], VI-[4], VII-[2], VIII-[2], IX-[7+ae], and I-[1], II-[11], III-[10], IV-[4+ae], V-[3], VI-[3], VII-[2], VIII-[2], IX-[7+ae], respectively, and of Zosimeidae with eight segments and armature formula I-[1], II-[7], III-[9], IV-[3+ae], V-[1], VI-[4], VII-[4], VIII-[7+ae], being the tetrasetose segment VII of the ground pattern of Zosimeidae, the result of fusion of the idyanthid bisetose primitive segments VII and VIII.
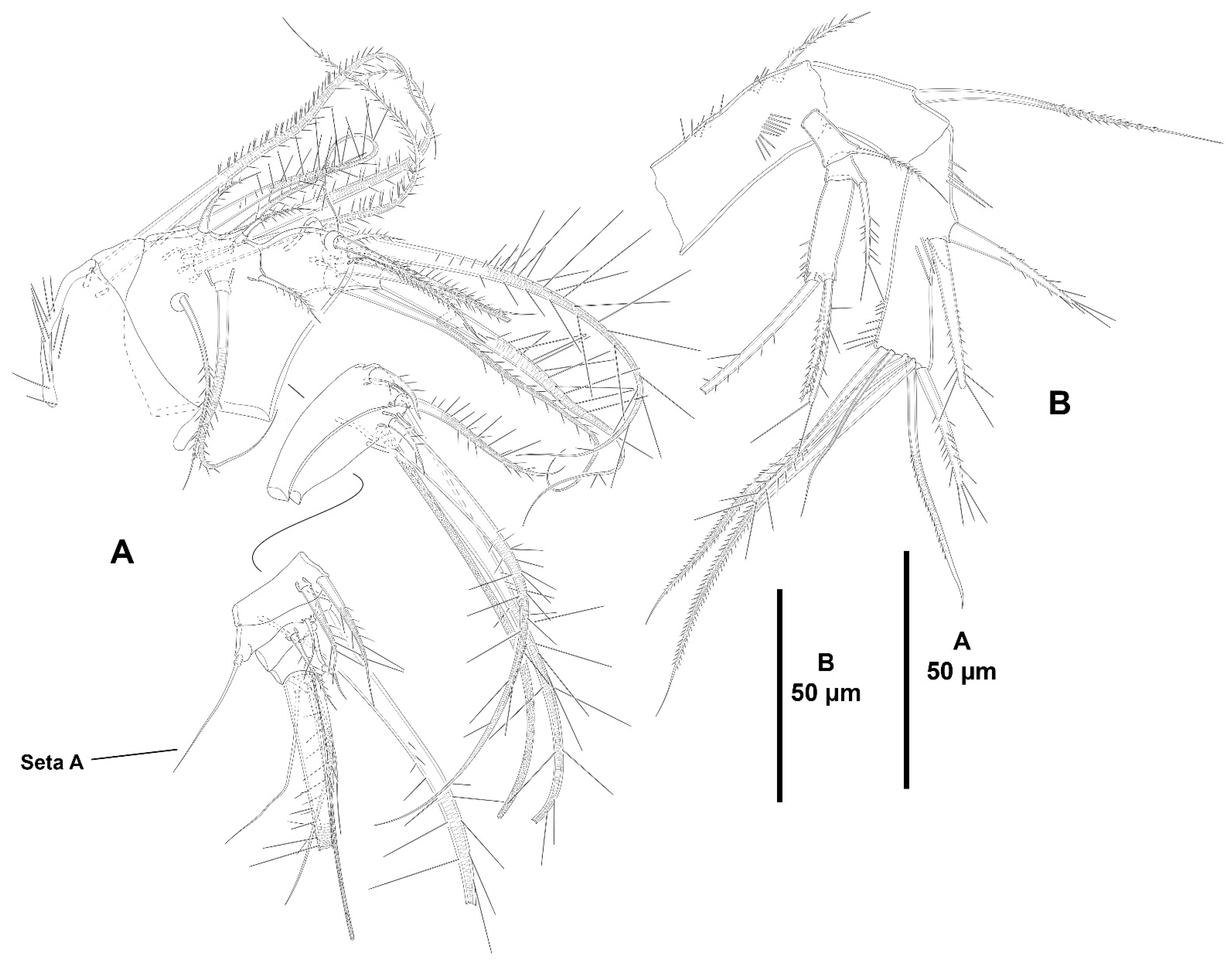
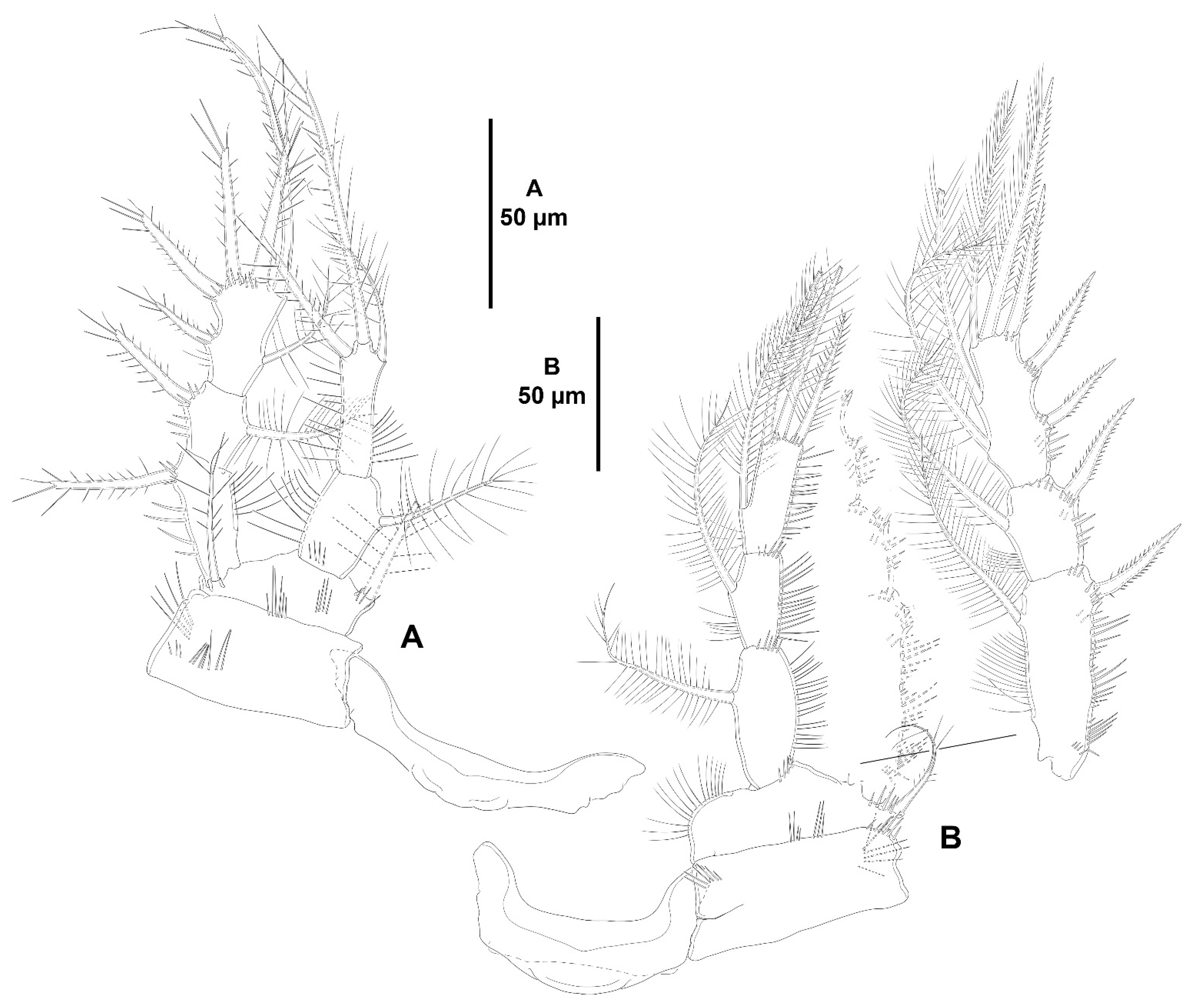
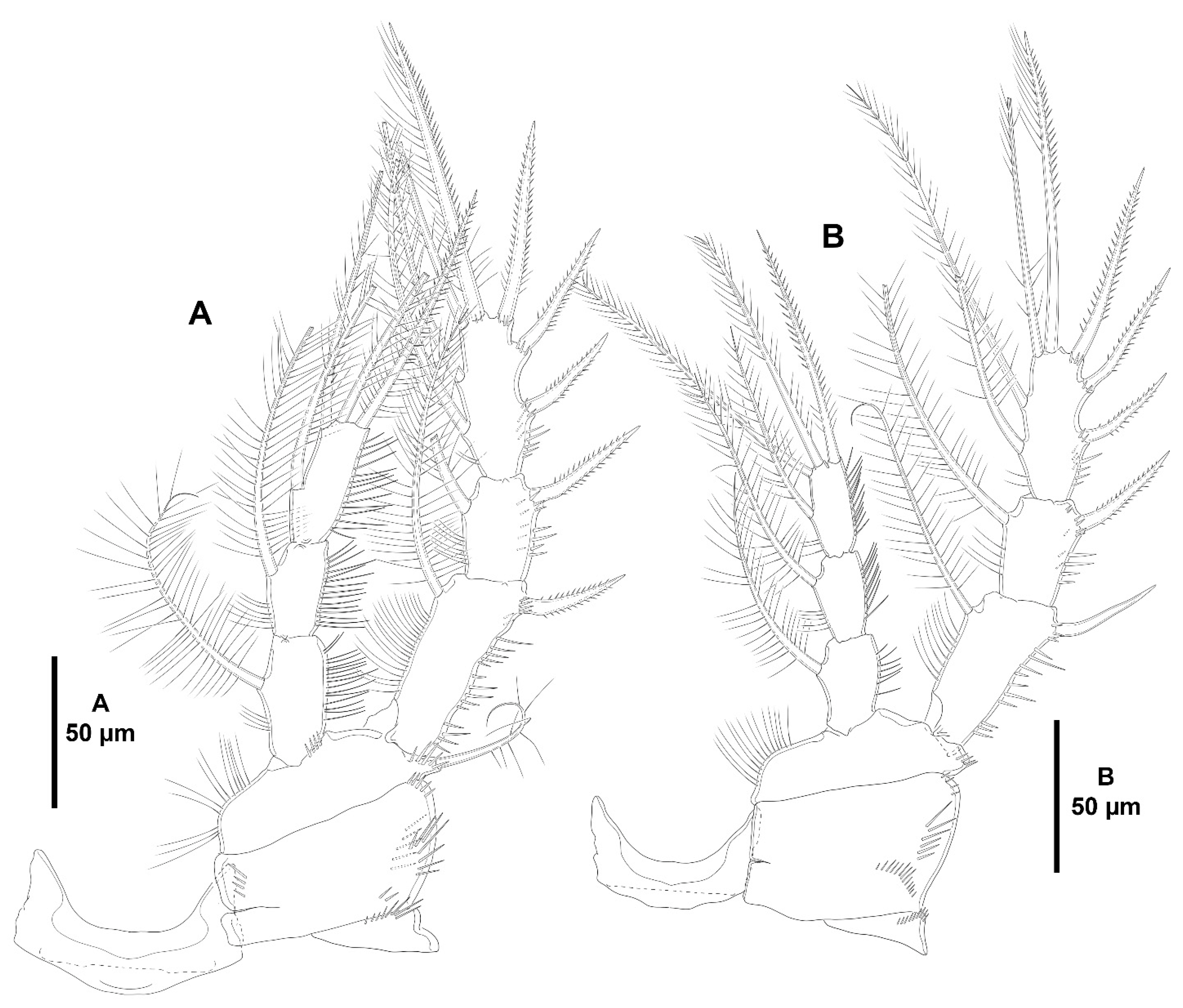
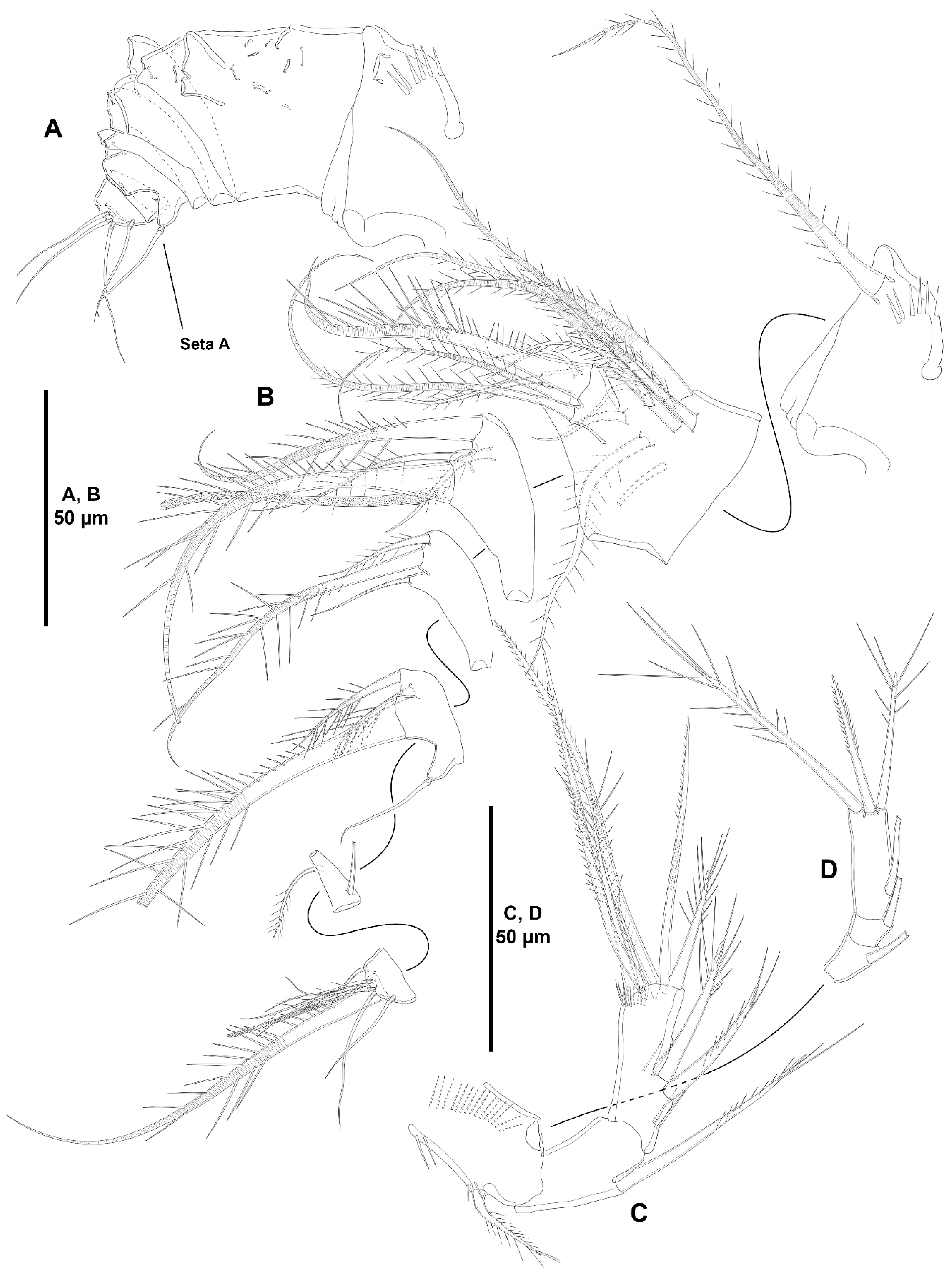
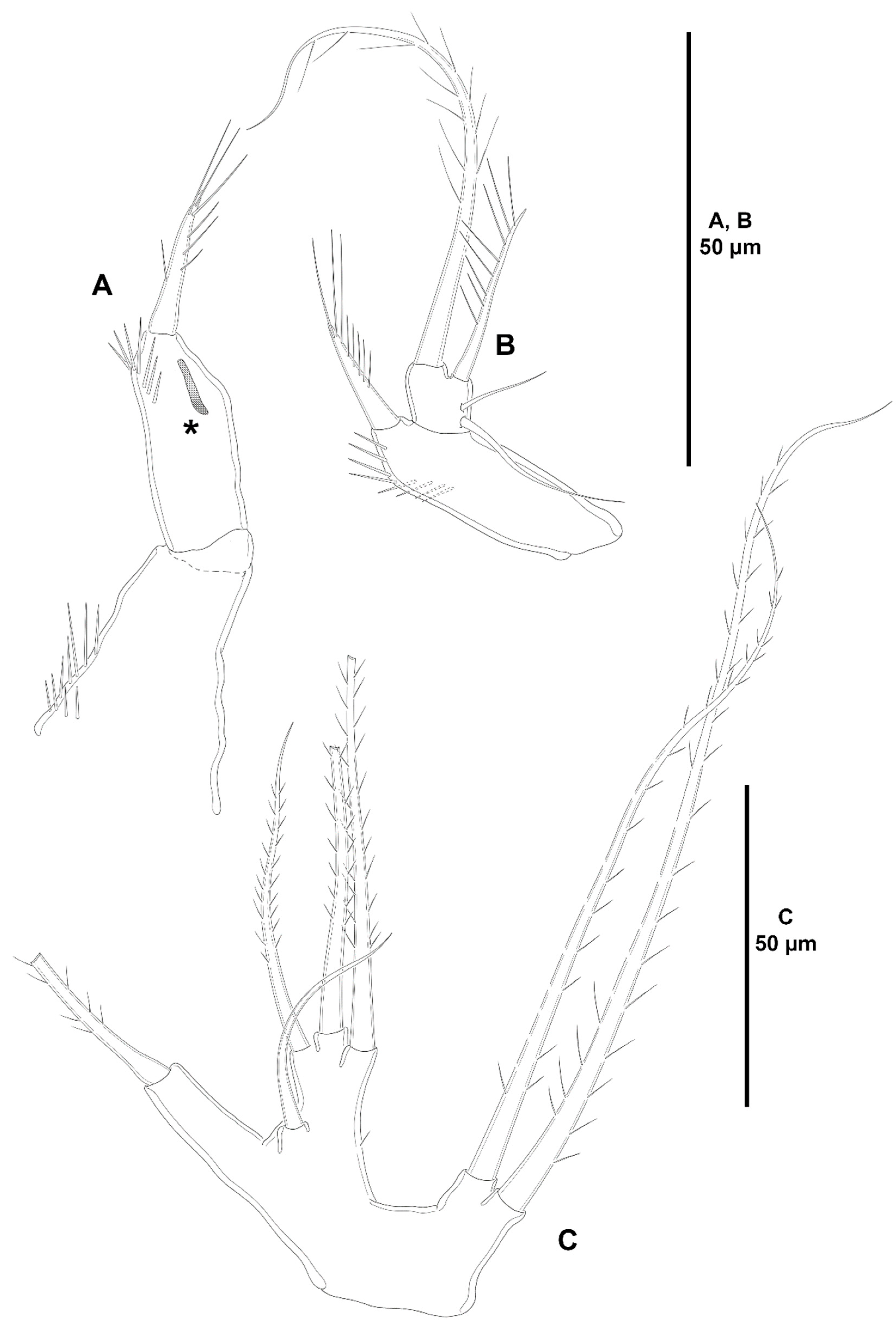
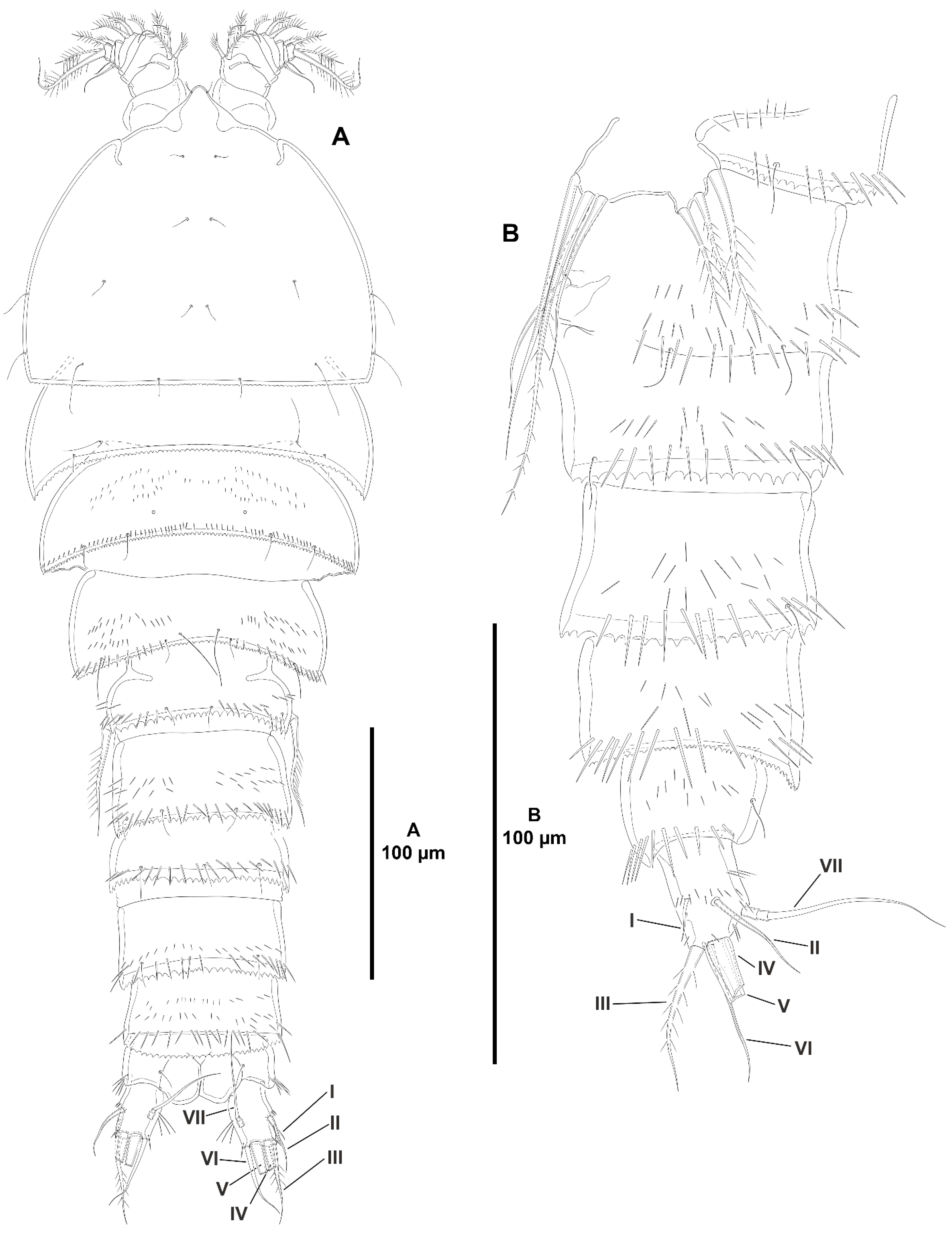
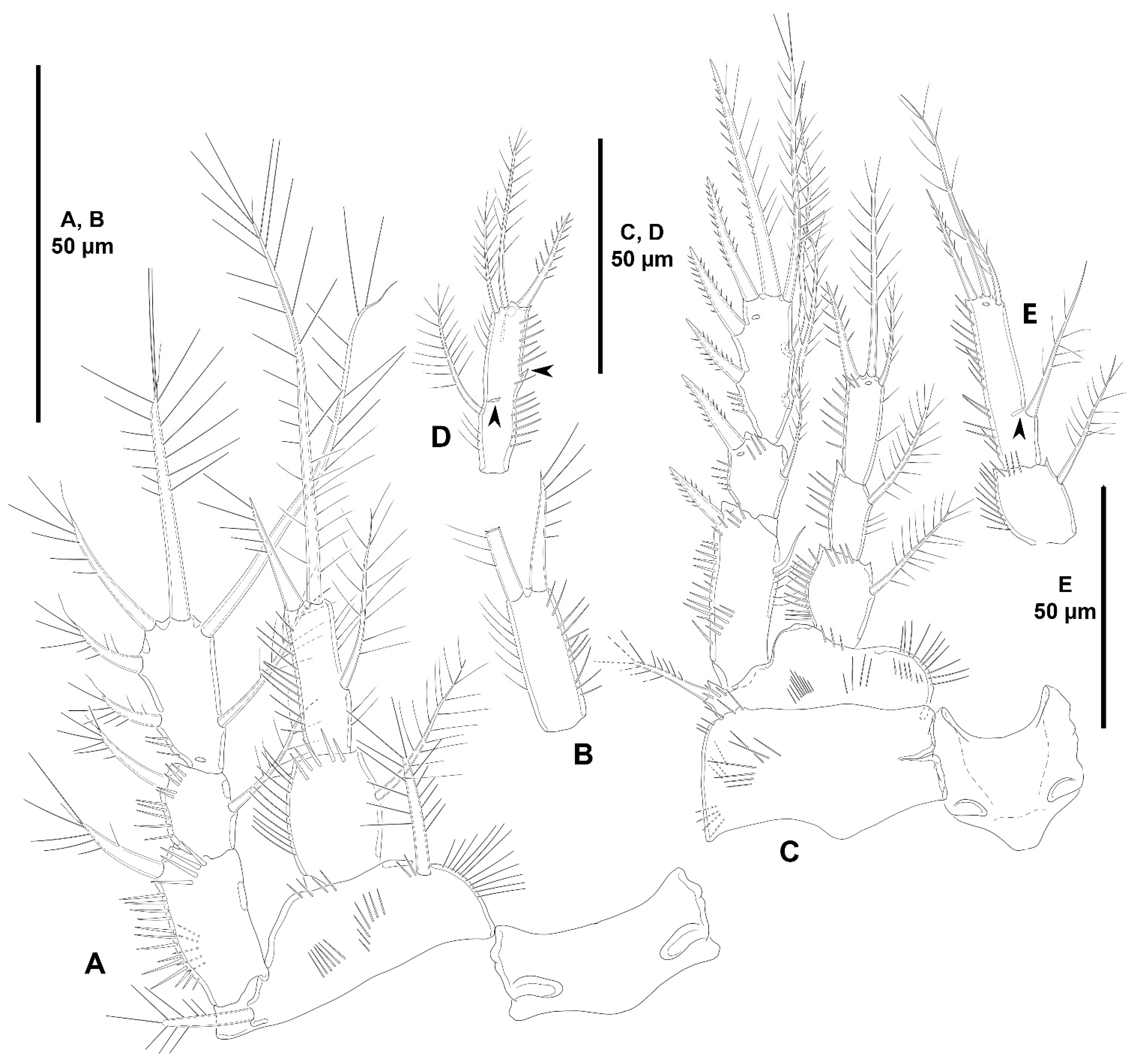
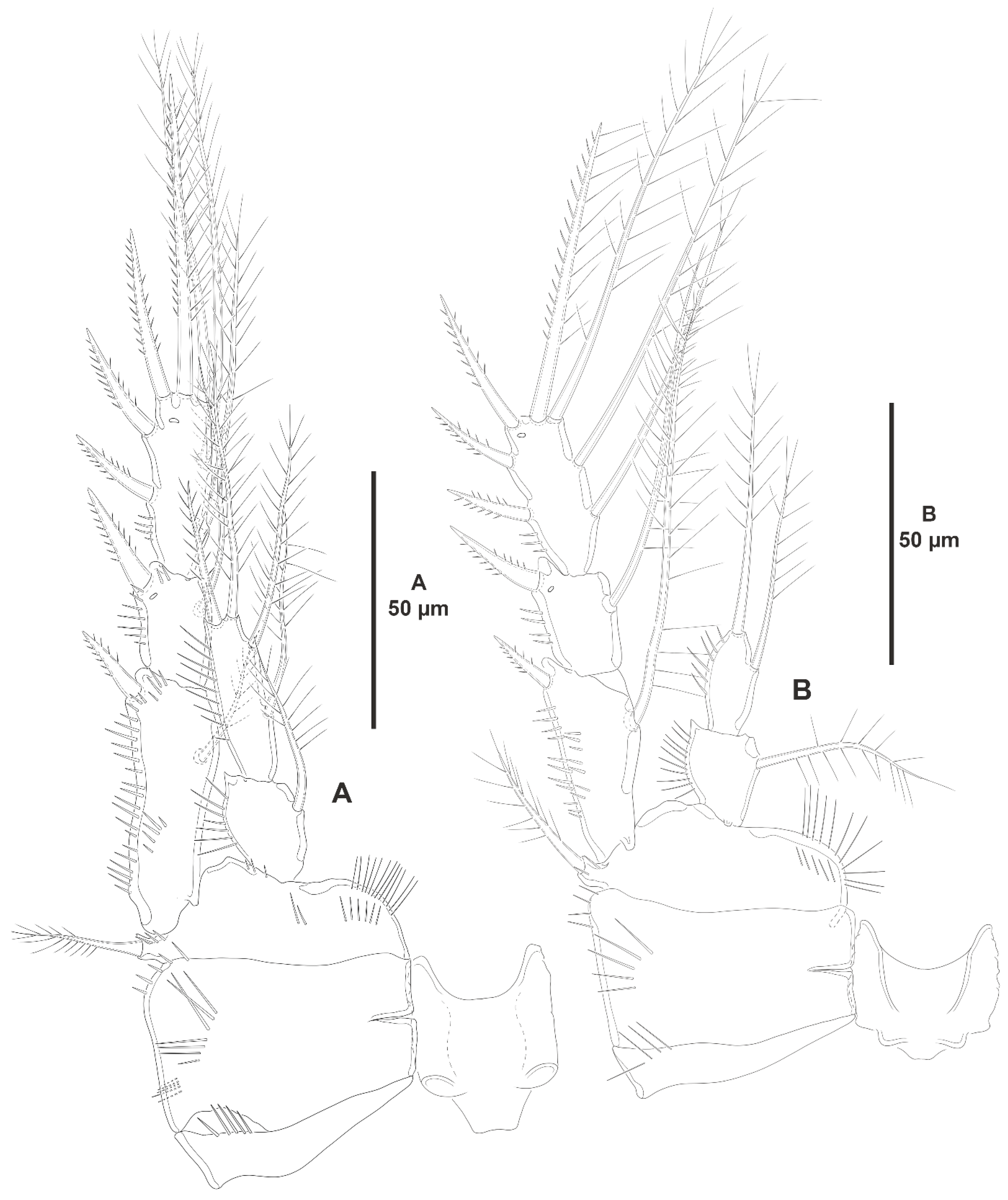
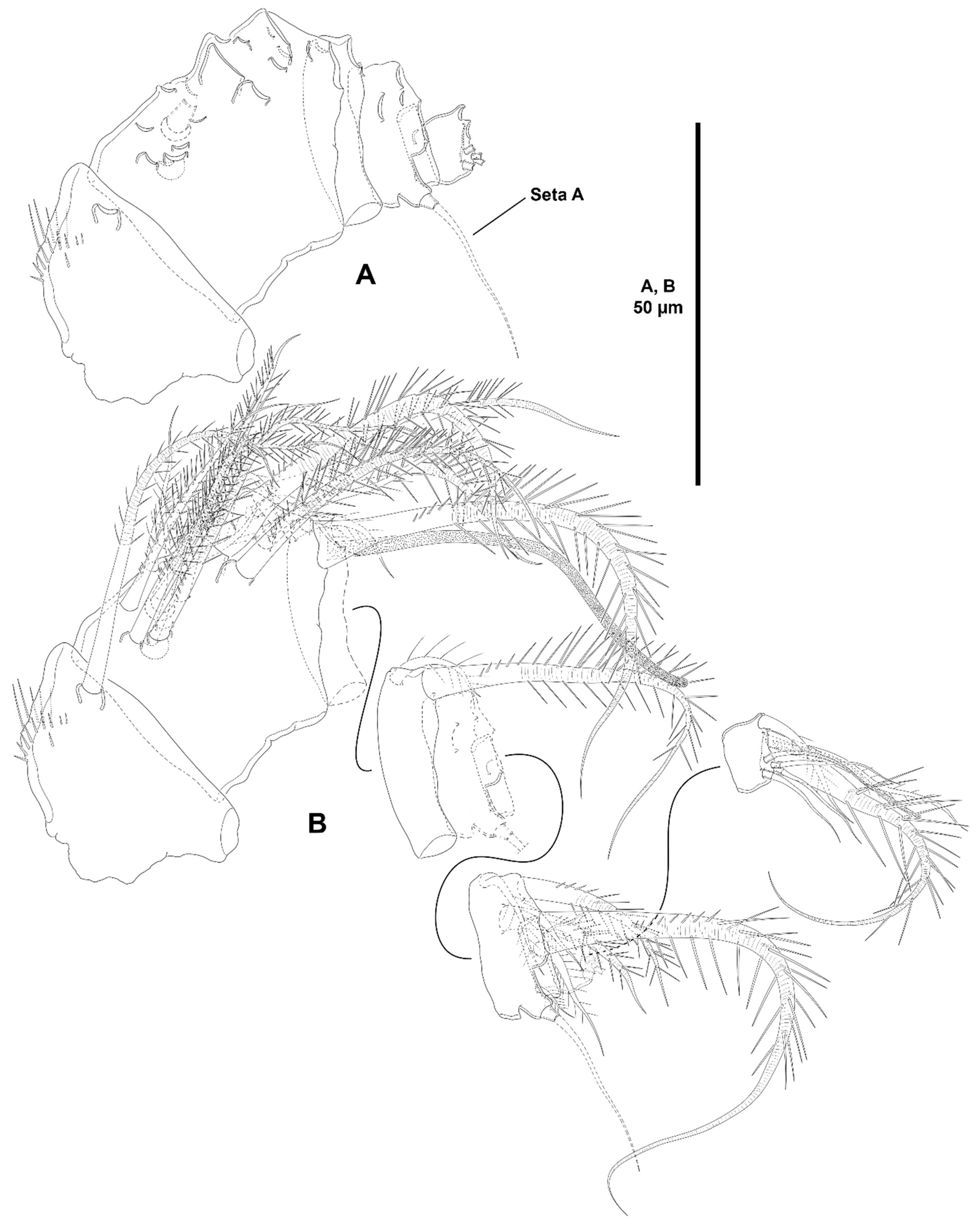
- The female antennules in Zosimeidae are very compact and short, the boundaries between segments are sometimes difficult to see, some segments are ill-separated, and the setae are piled making the counting and homologation of the armature elements very difficult. However, there are two elements that are easy to identify, the proximal aesthetasc and an outer biarticulate seta arising from a strong outer extension of segment 4, 5, or 6 (hereafter referred to as seta A; see Figure 3A, Figure 9A and Figure 22A).The proximal aesthetasc is primitively located on the fourth segment in Idyanthidae and Zosimeidae [11], and is commonly fused basally to a single seta. This armature complex is located either on the second, third, or fourth actual segments in Zosimeidae (Table S2). Its position in the fourth segment indicates that there has not been any fusion of the proximal primitive segments I–IV like in Z. valida [26], Z. pacifica, and Z. destituta, and Pe. pryorae; its position on the third segment indicates fusion of primitive segments II and III like in Z. anneae, Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, Z. carsteni, Z. mexicana sp. nov., Z. montagnai, Z. thistlei, Z. atlantica, Z. changi, and Z. tunnelli, or fusion of primitive segments III and IV like in Pe. pryocta sp. nov. and A. sigsbeeiensis sp. nov.; its position on the second segment indicates that primitive segments II–IV became fused like in Z. comata, Heterozosime, and A. spinesco. However, the fact that (i) the aesthetasc lies on the third segment of the eight-segmented female antennule of Z. anneae, Z. xiximi. sp. nov., Z. eliasi, and Z. gymnokosmosa (these species underwent secondary fusion of primitive segments II and III forming the actual second segment), or in the second segment of the seven-segmented female antennule of A. spinesco (the aesthetasc-bearing second segment is the result of the fusion of primitive segments II–IV), and (ii) that the primitive segments II and III are separated in the seven-segmented female antennule of Z. valida, in the six-segmented female antennules of Z. pacifica, and Z. destituta, and in the eight-segmented female antennule of Pe. pryorae, indicates that the ground pattern of Zosimeidae includes a nine-segmented female antennule, instead of an eight-segmented appendage as in Seifried [11]. Moreover, some species display two setae on the antennulary segment that is most probably homologous to the primitive segment V (fourth segment in Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, Z. incrassata, Z. changi, Pe. pryocta sp. nov., and A. sigsbeeiensis sp. nov.; third segment in Z. comata, Heterozosime, and A. spinesco). Therefore, the armature formula of the ground pattern of Zosimeidae is assumed to be 1-[1], 2-[7], 3-[9], 4-[3+ae], 5-[2], 6-[4], 7-[2], 8-[2], 9-[7+ae]. Some other amendments in the armature complement of the antennulary segments are probable, but this is beyond the scope of the present discussion.Seta A is present in most members of the family, and it can be located either on the fourth, fifth, or sixth antennulary segment (Table S2). This seta can be traced back to the primitive segment VII of Idyanthidae. The outer seta on the actual seventh and eighth segments is still present in the nine-segmented antennule of Idyanthe australis Pallares, 1970. The fusion of the idyanthid primitive segments VII and VIII resulted in the actual seventh segment of the eight-segmented female antennule of Idyella kunzi Bodin, 1968, I. nilmaensis Kornev and Chertoprud, 2008, Tachidiella kimi Lee and Huys, 1999, and T. sarsi Kornev and Chertoprud, 2008, with two outer setae, amongst other inner elements. Zosime anneae and P. pryocta sp. nov. kept the—primitive—idyanthid bisetose sixth and seventh segments (homologous to the primitive segments VII and VIII) in their eight-segmented female antennules, and they became fused in A. sigsbeeiensis sp. nov. forming its tetrasetose sixth segment (see Figure 27A). The outer biarticulate seta of the sixth segment in Z. carsteni, Z. eliasi, and Z. gymnokosmosa, and in the fifth segment of Z. thistlei (homologous to the primitive segment VII) arises from poorly developed extension of the segment. This extension is well-developed in the sixth segment of Z. atlantica, Z. xiximi sp. nov., in the fifth segment of Z. changi, Z. tunnelli, Z. montagnai, Z. destituta, Z. pacifica, Z. mexicana, and in the fourth segment of Z. comata and H. klemie sp. nov. and is regarded here as apomorphic for these species.The position of the proximal aesthetasc and the position of seta A on the female antennule are used here as landmarks to better understand the segmentation patterns of the female antennule of Zosimeidae.The number of segments of the female antennule of Zosime varies from six to eight (see Table S2) and seems to be distributed randomly amongst the species of the typica-group and its subordinate subgroups; the number of segments of the female antennule varies from seven to eight in the incrassata-group, but the armature formula is known only for Z. carsteni. The female antennule is composed of six or eight segments in Peresime, but the armature formula is known only for Pe. pryorae and Pe. pryocta sp. nov. with eight-segmented antennules. The monotypic genus Pseudozosime has five-segmented female antennules, but the armature formula remains unknown. It is six-segmented in Heterozosime and seven-segmented in Acritozosime.The fusion pattern of the primitive antennulary segments of Zosimeidae is not clear. Briefly, fusion took place in primitive segments II and III, II–IV, IV and V, V and VI, V–VII, V–VIII, VI and VII, VII and VIII in Zosime; in V and VI in Pe. pryorae; in III and IV in Pe. pryocta sp. nov.; in II–IV, and VI and VII in H. tenuis and H. klemie sp. nov.; in II–IV in A. spinesco; and in III and IV, and VII and VIII in A. sigsbeeiensis sp. nov.The eight-segmented antennule of Z. anneae, Z. xiximi sp. nov., Z. eliasi, Z. gymnokosmosa, and Z. carsteni is the result of fusion of primitive segments II and III, resulting in the third and sixth segment with the proximal aesthetasc and seta A, respectively, except for Z. anneae in which the strong extension of the supporting segment from which seta A arises seems to have been suppressed.The seven-segmented female antennules in Zosime are the result of different processes. Zosime mexicana sp. nov., Z. montagnai, and Z. thistlei underwent fusion of primitive segments II and III, and IV and V, resulting in the third and fifth segment with the proximal aesthetasc and seta A, respectively. The seven-segmented female antennule of Z. atlantica, Z. changi, and Z. valida is the result of fusion of primitive segments II and III, and VII and VIII in Z. atlantica with the proximal aesthetasc and seta A on the third and sixth segment, respectively; II and III, and VI and VII in Z. changi with the proximal aesthetasc and seta A on the third and fifth segment, respectively; and V–VII in Z. valida with the proximal aesthetasc on the fourth segment (the position of seta A is inconclusive, but it is, in all probability, on the sixth segment).The six-segmented female antennules of Z. tunnelli, Z. comata, and Z. pacifica and Z. destituta is the result of fusion of primitive segments II and III, V and VI, and VII and VIII resulting in the third and fifth segments with the proximal aesthetasc and seta A, respectively, in Z. tunnelli; II–IV, and VI and VII in Z. comata with the proximal aesthetasc on the second segment and seta A on the fourth segment; and V–VIII in the latter two species with the proximal aesthetasc on the fourth segment and seta A on the fifth segment.The fusion pattern is also different in the eight-segmented female antennules of the two species of Peresime for which the armature formulae are known. It is the result of fusion of primitive segments V and VI in Pe. pryorae with the proximal aesthetasc and seta A on the fourth and sixth segment, respectively, but III and IV in Pe. pryocta sp. nov. with the proximal aesthetasc on the third segment (the development of the strong extension of the supporting segment from which seta A arises seems to have been suppressed).Both species of Heterozosime underwent fusion of primitive segments II–IV, and VI and VII, resulting in six-segmented antennules with the proximal aesthetasc on the second segment, and seta A on the fourth segment.The fusion pattern in both species of Acritozosime is different. The seven-segmented female antennule of A. spinesco seems to be the result of fusion of primitive segments II–IV, resulting in the second and fifth segment with the proximal aesthetasc and seta A, respectively; but in A. sigsbeeiensis sp. nov. the fusion took place in primitive segments III and IV, and VII and VIII, resulting in the third segment with the proximal aesthetasc (the outer extension of the supporting segment from which seta A arises seems to have been suppressed).The phylogenetic value of the fusion pattern between the species of each genus is not clear, but some species share a particular set of fusion events, e.g., (i) Z. anneae, Z. xiximi sp. nov., Z. eliasi, and Z. gymnokosmosa (the armature complement of the third segment of Z. carsteni—12 setae+ae—is significantly different); (ii) Z. mexicana sp. nov., Z. montagnai, and Z. thistlei; (iii) Z. pacifica and Z. destituta; (iv) H tenuis and H. klemie sp. nov. (Table S2).
- −
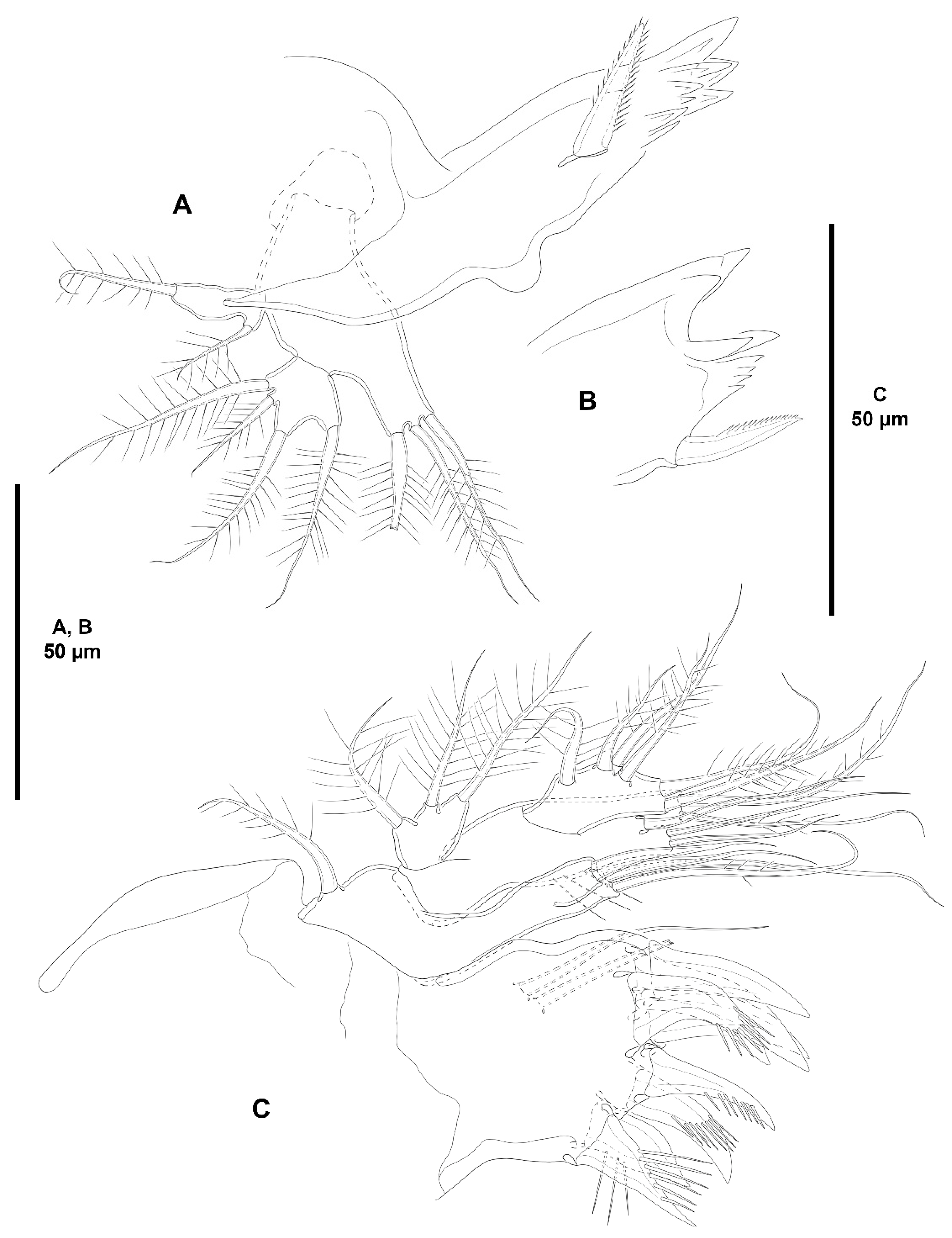
- Characters 6–10 (armature of A2 EXP). The A2 EXP1 is armed with one seta in all species of Zosime for which this ramus has been described (it is unknown in Z. gisleni, Z. major, Z. mediterranea, Z. erythraea, Z. bathyalis, and Z. reyssi), and in Peresime, Pseudozosime, and Heterozosime. Acritozosime is the only genus of the family with the plesiomorphic bisetose A2 EXP1 (see Kim and Lee [1] for an in-depth discussion of this character).
- The second exopodal segment bears one seta in all species of the typica-group of Zosime, in Peresime, Pseudozosime, and Heterozosime. This seta is absent in the incrassata-group except for Z. carsteni with one seta; it is absent also in Acritozosime. The loss of the seta of A2 EXP2 seems to have occurred independently several times within the family and is regarded here as synapomorphic for Z. incrassata, Z. bathybia and Z. reyssi, and apomorphic for Acritozosime (Table S1; Figure 32).The armature of EXP3 is variable. One inner lateral seta is present in all species of subgroup A of Zosime except for Z. atlantica and Z. xiximi sp. nov. (typica-group, subordinate subgroup Ab; the condition in Z. mediterranea remains unverifiable), and for the incrassata-group (except for Z. carsteni with one lateral inner seta). Zosime atlantica, Z. xiximi sp. nov., Z incrassata, Z. bathybia, and Z. reyssi underwent secondary—apomorphic—loss of this seta, and it is the most derived condition in the family. The inner lateral seta of A2 EXP3 is also present in all the other genera of the family.The distal armature complement of the A2 EXP3 varies from three to two. Three setae are present in some species of Zosime (Z. anneae, Z. paratypica, Z. mexicana sp. nov., and Z. pacifica (typica-group of Zosime, subordinate subgroup Aa), Z. atlantica and Z. xiximi sp. nov. (typica-group of Zosime, subordinate subgroup Ab), Z. thistlei (typica-group of Zosime, subordinate subgroup Ac), and Z. bathybia (incrassata-group)), in Pe. pryorae, Pe. reducta, and Pe. pryocta sp. nov., and in Heterozosime, and Acritozosime. Two setae are present in some species of Zosime (Z. typica and Z. montagnai (typica-group, subordinate subgroup Aa), and Z. eliasi, Z. bergensis, and Z. tunnelli (typica-group, subordinate subgroup Ac); the condition of Z. gisleni and Z. major (typica-group, subordinate subgroup Aa), and Z. mediterranea (typica-group, subordinate subgroup Ab) remain unverifiable)), in Z. carsteni, Z. incrassata, and Z. reyssi (incrassata-group of Zosime), and in Pe. abyssalis.
- −
- Character 11 (presence/absence of the inner seta of A2 ENP1). Most species of Zosimeidae possess one inner seta on A2 ENP1. This seta is present also in Idyanthidae. The inner seta of A2 ENP1 is missing in Z. paratypica, Z. thistlei, and A. spinesco. The loss of this element in two species of Zosime and in one species of Acritozosime seems to have occurred independently and is considered apomorphic for A. spinesco. It is also an apomorphy (potential synapomorphy?) for Z. paratypica and Z. thistlei in their respective subordinate subgroups.
- −
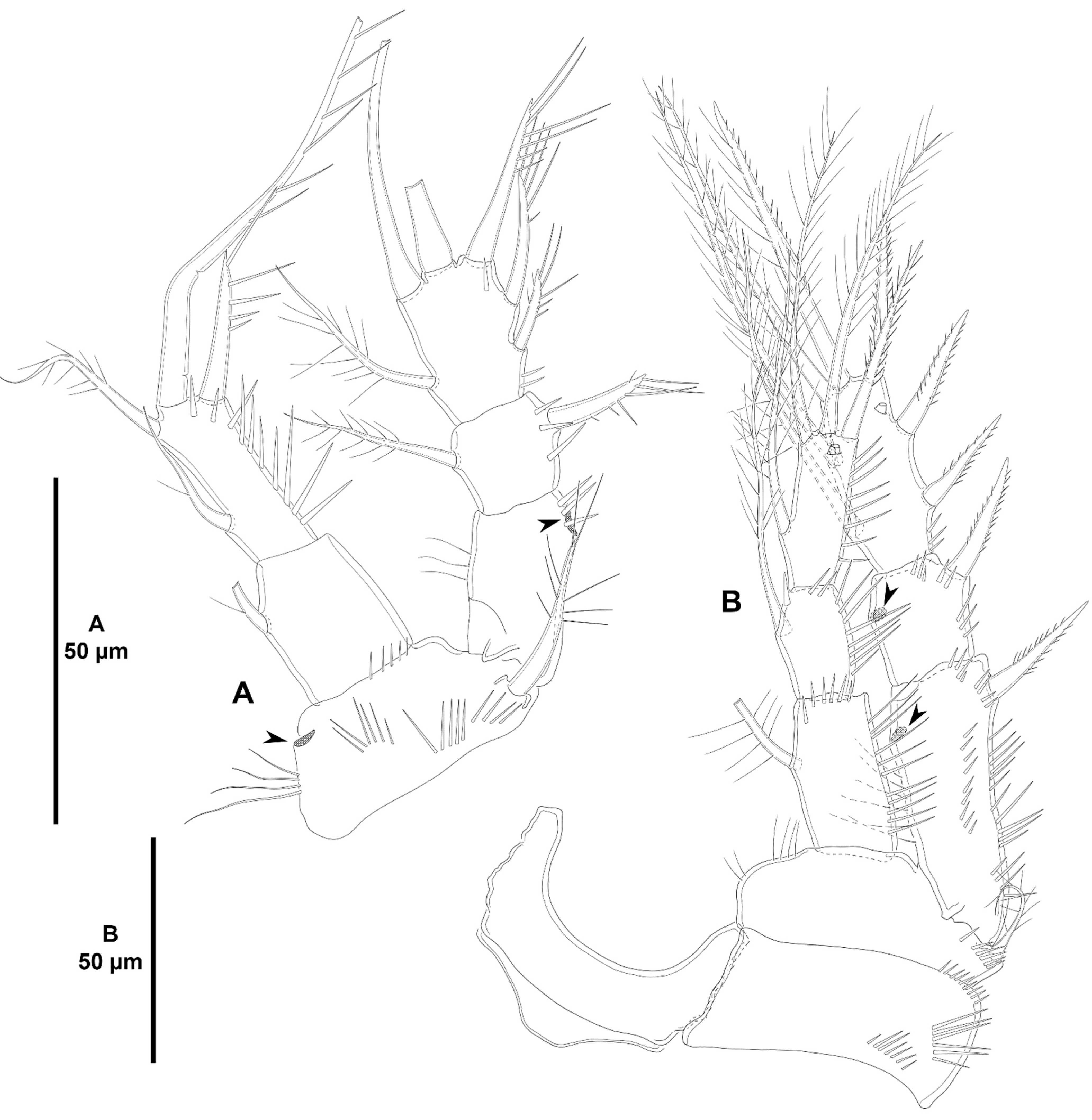
- Characters 12–13 (A2 ENP2, lateral armature). Most species of Zosimeidae display the armature of the second antennary endopod as described by Seifried [11] in her diagnosis and ground pattern of the family, i.e., with one proximal short (element I in Seifried [11]), and one subdistal spine (element III in Seifried [11]) set close to a reduced slender seta (element 4 in Seifried [11]). This setal arrangement occurs in Zosime except for some species with only two lateral spines (Seifried’s [11] elements I and III) on the second endopodal segment of the antenna, Z. anneae, Z. montagnai, Z. eliasi, Z. tunnelli, Z. valida, and Z. incrassata. It is unclear whether the absence of the small seta 4 is a true secondary loss or if this small seta has been overlooked in these species since it is often masked by element III. On the other hand, Peresime and Acritozosime deviates from the general ground pattern suggested by Seifried [11] for the family. The lateral armature of A2 ENP2 of Peresime and Acritozosime consists of three well-developed elements, i.e., spiniform elements I and III, and—primitive—well-developed setiform element 4. Element 4 is primitively well-developed and geniculate in Idyanthidae [11]. Elements I, III, and 4 are geniculate in Pe. pryocta sp. nov. and in A. sigsbeeiensis sp. nov., and their conditions in the other species of Peresime, and in A. spinesco remain uncertain. As noted above, the well-developed element 4 in Peresime and Acritozosime seems to be a plesiomorphy, and the reduction in seta 4 seems to have occurred independently in Zosime, Pseudozosime, and Heterozosime.
- −
- Character 14 (number of distal elements on A2 ENP2). The ground pattern of Idyanthidimorpha and Idyanthidae include seven distal elements on A2 ENP2 [11]. Most members of the family Zosimeidae underwent the synapomorphic loss of one element resulting on six distal setae/spines on A2 ENP2. Zosime atlantica, Z. incrassata, and Pe. reducta were described with five distal elements on A2 ENP2, but this is probably erroneous and requires confirmation. Acritozosime kept the plesiomorphic idyanthid armature complement of seven distal elements on A2 ENP2. Kim and Lee [1] believed that the presence of seven distal elements on A2 ENP2 is a plesiomorphy, and, therefore, ruled out the possibility of the extra seta being a novelty. This view is followed here.
- −
- Characters 15–19 (mandibular palp). The primitive mandibular basis of Idyanthidimorpha and Idyanthidae possesses four setae [11]. The family Zosimeidae underwent loss of one seta resulting in a trisetose basis. The primitive zosimeid trisetose basis is present in the typica-group of Zosime, and in Heterozosime. The mandibular basis of Acritozosime bears two setae and is regarded here as a potential autapomorphy for the genus. One seta is present in Peresime, and in the incrassata-group of Zosime, except for Z. carsteni. The latter kept the primitive trisetose condition. The reduced unisetose mandibular palp of Z. reyssi is unique in the family and is regarded here as autapomorphic for the species. The reduction from three to one seta seems to have occurred independently twice: the unisetose basis is a synapomorphy for Z. incrassata and Z. bathybia within Zosime and is an apomorphy for Peresime. The mandibular palp of Pseudozosime remains undescribed.
- The mandibular exopod is two-segmented in the ground pattern of Idyanthidimorpha and Idyanthidae, the proximal segment bearing four lateral—inner—setae, the distal segment with two apical setae [11]. The same architecture of that ramus is autapomorphic for Idyanthidae [11]. Both segments of the primitive idyanthidimorph mandibular exopod became fused and underwent loss of some setae, resulting in the—autapomorphic—one-segmented ramus in Zosimeidae, with three lateral—inner—setae and one distal element [11]. The one-segmented mandibular exopod with the primitive setal arrangement (three lateral and one distal element) is still present in Z. atlantica. The tetrasetose mandibular exopod is also present in P. reducta, Pe. pryocta sp. nov., A. tenuis, and A. sigsbeeiensis sp. nov. but the four exopodal setae of these species seem to have undergone secondary arrangement consisting of the displacement of the two distalmost lateral setae to the apical margin, giving the impression of one lateral—inner—and three distal setae. Almost all the species of the family underwent secondary loss of one seta, resulting in a trisetose ramus. The trisetose ramus is present in almost all species of Zosime, in Pe. abyssalis and Pe. pryorae, and Heterozosime. The bisetose mandibular exopod of Z. xiximi sp. nov. is autapomorphic for the species and is the most derived condition within the typica-group of Zosime. The knob-like, unisetose mandibular exopod in the incrassata-group was regarded as autapomorphic for that group by Pointner [13]. The one-segmented mandibular exopod with one seta is autapomorphic for Z. carsteni (incrassata-group of Zosime), the mandibular exopod absorbed into the basis and represented by one seta is a synapomorphy for Z. incrassata and Z. bathybia (incrassata-group of Zosime), and the reduced one-segmented mandibular palp of Z. reyssi with one seta only is autapomorphic for the species.The primitive idyanthidimorph and idyanthid mandibular endopod is one-segmented, with three lateral—inner—setae and three distal sets of three, two, and two setae fused basally [11]. Seifried [11] suggested that the tetrasetose mandibular endopod of Zosimeidae, with one lateral—inner—and three distal setae, is an autapomorphy for the family. However, the mandibular endopod of Peresime and Acritozosime bears one lateral and four distal elements, and the pentasetose mandibular endopod seems to be the autapomorphic condition for Zosimeidae. The—apomorphic—secondary reduction from four to three lateral setae is widespread in the family and seems to have occurred independently in Zosime (this condition is present in all species of the typica-group, and in Z. carsteni of the incrassata-group), and in Heterozosime. Two species of the incrassata-group of Zosime, Z. incrassata and Z. bathybia, underwent synapomorphic loss of two apical setae, resulting in a bisetose ramus, with one lateral and one distal seta. As noted above, the one-segmented unisetose mandibular palp is autapomorphic for Z. reyssi.
- −
- Character 20 (number of epipodal setae of the maxillulary coxa). The primitive idyanthidimorph maxillulary coxa is armed with two epipodal setae, and it is unisetose in Idyanthidae [11]. The primitive idyanthidimorph bisetose coxal epipodite is present in the ground pattern of Zosimeidae [11] and is still present in some species of Zosime (Z. anneae, Z. pacifica, Z. atlantica, and Z. valida, all of the typica-group of the genus). The secondary loss of one epipodal seta occurred independently in Zosime (this condition is present in the other species of the genus for which this appendage has been described; Z. reyssi was described without epipodal seta, but this requires confirmation), in two species of Peresime (Pe. abyssalis and Pe. pryocta sp. nov.), and in Heterozosime. The compete loss of the epipodal setae occurred independently in some species of Peresime (Pe. pryorae and Pe. reducta) and in Acritozosime. The bisetose condition of Z. anneae, Z. pacifica, Z. atlantica, and Z. valida is a plesiomorphy, and the unisetose condition of the other species of Zosime is regarded here as apomorphic within the genus. The unisetose condition of Pe. abyssalis and Pe. pryocta sp. nov. is primitive within that genus, and the unarmed coxal epipodite is a potential synapomorphy for Pe. pryorae and Pe. reducta. The complete loss of the epipodal armature in Acritozosime is regarded here as apomorphic for that genus.
- −
- Character 21 (number of segments of the maxillary endopod). Following Seifried [11], the ground pattern of the maxilla comprises a three-segmented endopod in Idyanthidimorpha and Idyanthidae, and a one-segmented endopod in Zosimeidae. However, some species of Zosime (Z. mexicana sp. nov., Z. xiximi sp. nov., Z. eliasi, Z. paramajor, Z. carsteni, and Z. bathybia; the condition of Z. incrassata is uncertain), H. tenuis, and both species of Acritozosime, display a two-segmented maxillary endopod. The secondary—apomorphic—fusion of both segments of the maxillary endopod seems to have occurred convergently in Zosime, Peresime, and Pseudozosime.
- −
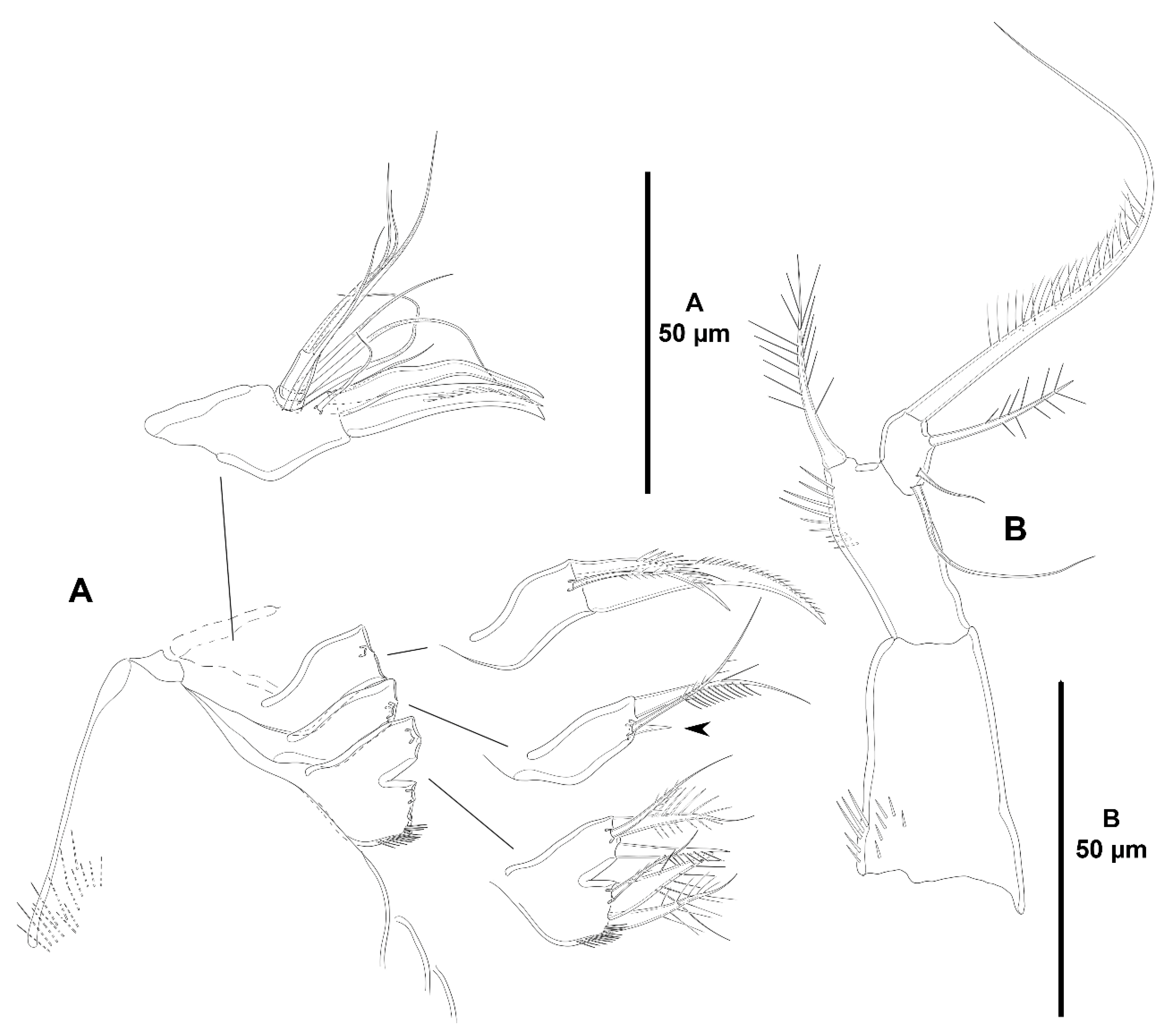
- Characters 22–27 (maxilliped). Following Seifried [11], the ground pattern of the maxilliped in Idyanthidimorpha and Idyanthidae includes (i) the geniculation between syncoxa and basis and between basis and endopod; (ii) the syncoxa with two subdistal setae; (iii) the basis with one inner seta; and (iv) the two-segmented endopod with one seta and one thin and one large claw on proximal segment, and two outer small and two distal geniculate long setae on distal segment. The ground pattern of the maxilliped of Zosimeidae includes (i) the autapomorphic loss of geniculation between syncoxa and basis and between basis and endopod; (ii) the syncoxa with two subdistal setae; (iii) the basis with one inner seta; and (iv) the autapomorphic loss of the first endopodal segment and its armature, resulting in the one-segmented ramus with the primitive setal complement of two outer small elements and two geniculate long distal setae [11].
- The syncoxa is mostly unarmed in the family. Zosime eliasi is the only species of Zosime with one seta. One seta is also present in Pe. pryorae, Pe. pryocta sp. nov., and in A. sigsbeeiensis sp. nov. Acritozosime spinesco kept the full primitive armature complement of two setae. Kim and Lee [1] believed that the maxilliped of A. spinesco was the most plesiomorphic maxilliped in Zosimeidae and that it was similar to the idyanthid maxilliped in the geniculation between the syncoxa and basis and in the bisetose syncoxa and ruled out the possibility that the geniculation between the syncoxa and the basis and the two setae on the syncoxa were acquired secondarily. This view is followed here.The basis of the maxilliped bears one seta in most species of Zosime, except for Z. bathybia (incrassata-group) and Pe. pryocta sp. nov. with unarmed bases.The primitive tetrasetose maxillipedal endopod is present in most species of Zosime, in Pe. pryorae and Pe. pryocta sp. nov., in Heterozosime, and Acritozosime. Secondary reduction to three setae occurred independently in Pseudozosime, and in Zosime (Z. typica (typica-group, subordinate subgroup Aa), Z. eliasi and Z. bergensis (typica-group, subordinate subgroup Ac), Z. comata (typica-group, subordinate subgroup Bb), and in the two species of the incrassata-group of Zosime for which this appendage has been described, Z. carsteni, and Z. bathybia). The armature underwent reduction to two setae independently in Z. paratypica (typica-group of Zosime, subordinate subgroup Aa) and in Pe. reducta.
- −
- Character 28 (length:width ratio of caudal rami). Pointner [13] regarded the longer-than-wide caudal rami as (syn)apomorphic for Z. anneae and the typica-group. The longer-than-wide caudal rami is also present in Heterozosime and Acritozosime, and the plesiomorphic subquadrate caudal rami is present in the incrassata-group of Zosime, Peresime and Pseudozosime. The caudal rami seem to have undergone secondary elongation independently at least in the typica-group of Zosime, in Heterozosime, and in Acritozosime.
- −
- Characters 66–68 (ornamentation of outer spines of P1–P4 EXP). In her ground pattern of Zosimeidae, Seifried [11] noted the subdistal very long spinules on the outer and distal exopodal spines of P1. Pointner [13] noticed differences in the spinular ornamentation of the outer exopodal spines of P1 EXP of Zosime. Following Pointner [13], the typica-group of Zosime display the plesiomorphic outer exopodal spines of P1 EXP with distal and subdistal elongate spinules, and the incrassata-group possesses the derived outer spines of P1 EXP homogeneously pinnate (either with thin and pointed, or with thick and blunt spinules). The outer exopodal spines of P2–P4 show the derived condition in all species of Zosime, except for Z. gymnokosmosa in which the subdistal and distal spinules of the outer spines of P2–P4 EXP are somewhat longer than the proximal spinules (see Kim and Lee [27] (p. 229, Figure 13D; p. 231, Figure 15A–C). The outer spines of P1 EXP of Z. tunnelli display a mix of both conditions: the outer distal spine shows the plesiomorphic spinules, but the other spines of that ramus show the derived condition (see Yeom et al. [2] (p. 15, Figure 10A)). The plesiomorphic condition of P1 EXP is also present in Peresime, Pseudozosime, Heterozosime, and Acritozosime. The outer exopodal spines of P1 and P2 of Pe. reducta display the plesiomorphic condition, but the outer spines of P3 and P4 show the derived condition. Heterozosime shows the derived condition in P2–P4; Acritozosime displays the plesiomorphic condition.
- −
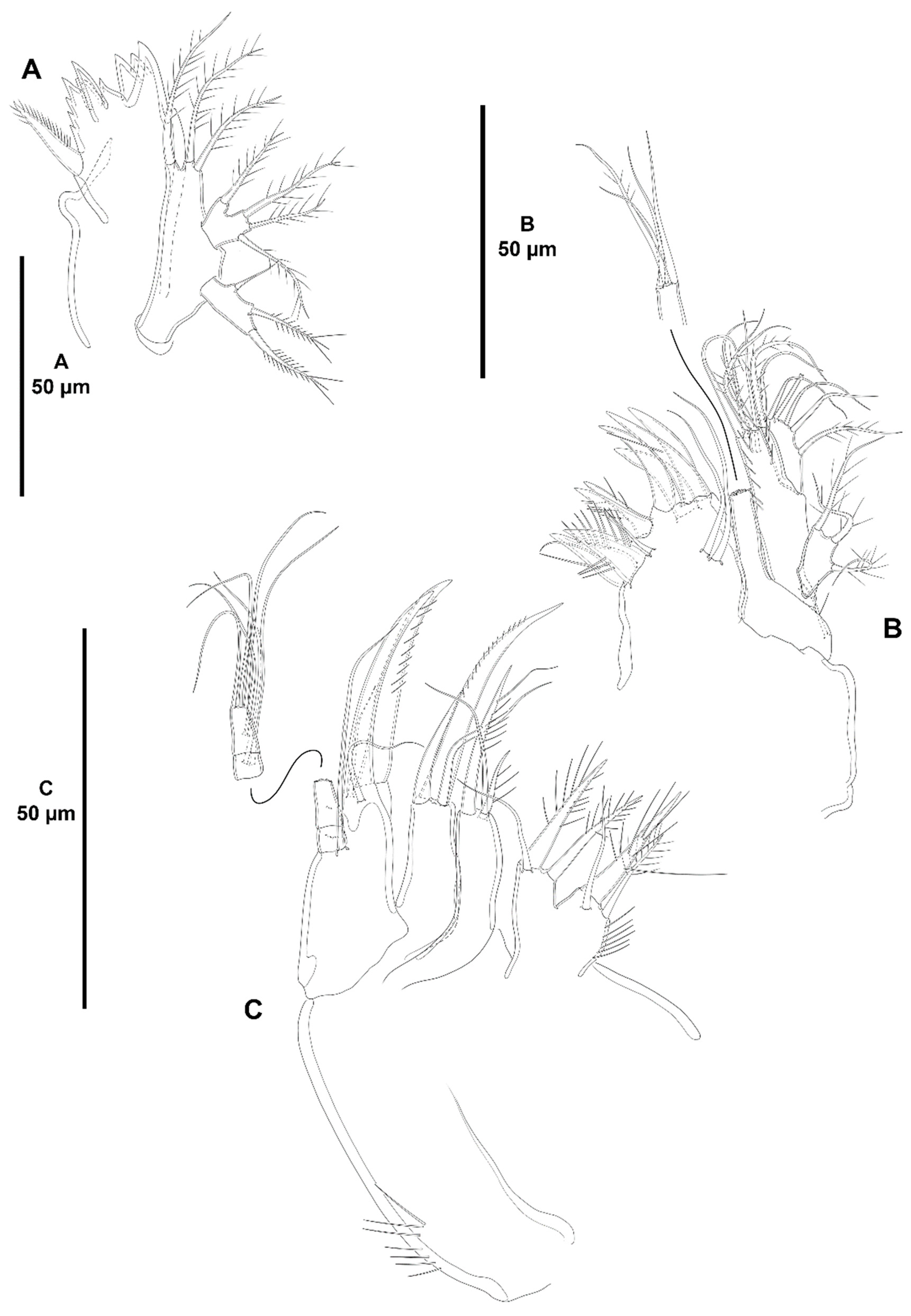
- Character 69 (number and length of the distal armature complements of the last segment of P4 ENP). Three distal setae on the last endopodal segment of P4 are present in all species of Zosime, except for Z. tunnelli, Z. gymnokosmosa, and Z. comata. Three distal setae are also present in H. tenuis and H. klemie sp. nov. The middle distal seta of P4 ENP3 is reduced to a very short seta in Z. mexicana sp. nov. (see Figure 13B, this study), Z. montagni (described as a tube-pore in Yeom et al. [2] (p. 8, Figure 5D)), and Z. pacifica (see Fiers [28] (p. 18–19, Figure 3d)). A similar reduction is present in H. tenuis and seems to have evolved independently. The reduction in the middle seta of P4 ENP3 is regarded here as synapomorphic for Z. mexicana sp. nov., Z. montagni, and Z. pacifica within Zosime, and apomorphic for H. tenuis against the primitive well-developed seta of H. klemie sp. nov. Zosime tunnelli and Z. gymnokosmosa share the—synapomorphic—loss of one distal seta of P4 ENP3; Z. comata underwent further loss of one more distal seta, for which one distal seta on P4 ENP3 is regarded here as autapomorphic within Zosime. The—apomorphic—loss of one distal seta on the last endopodal segment occurred also in the two-segmented endopod of Pseudozosime and Acritozosime. The most derived state amongst the genera of Zosimeidae is that of Peresime, in which the distal segment of the two-segmented endopod of P4 lost two setae, resulting in an unisetose distal margin of P4 ENP2; the latter is regarded here as autapomorphic for Peresime.
- −
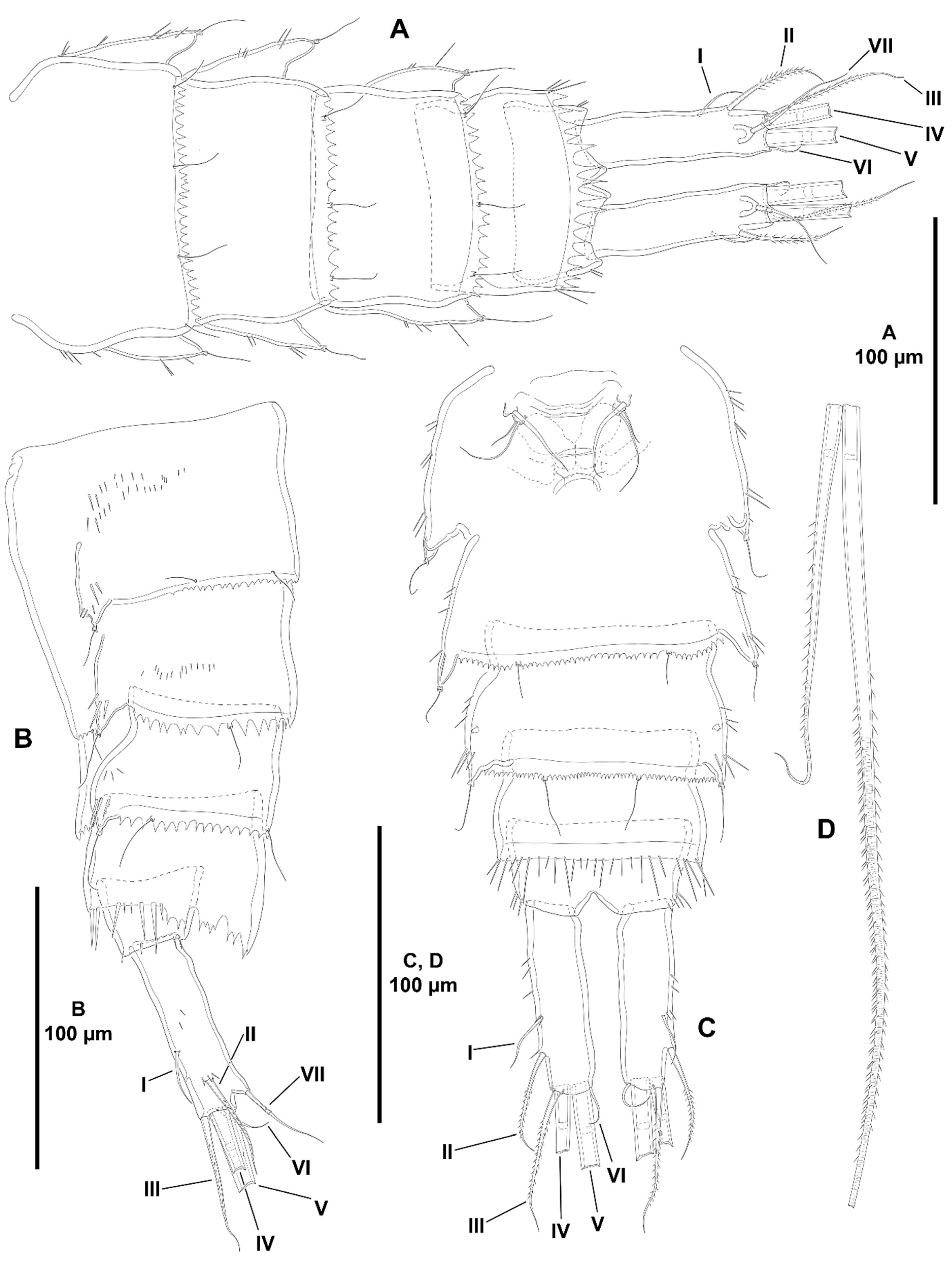
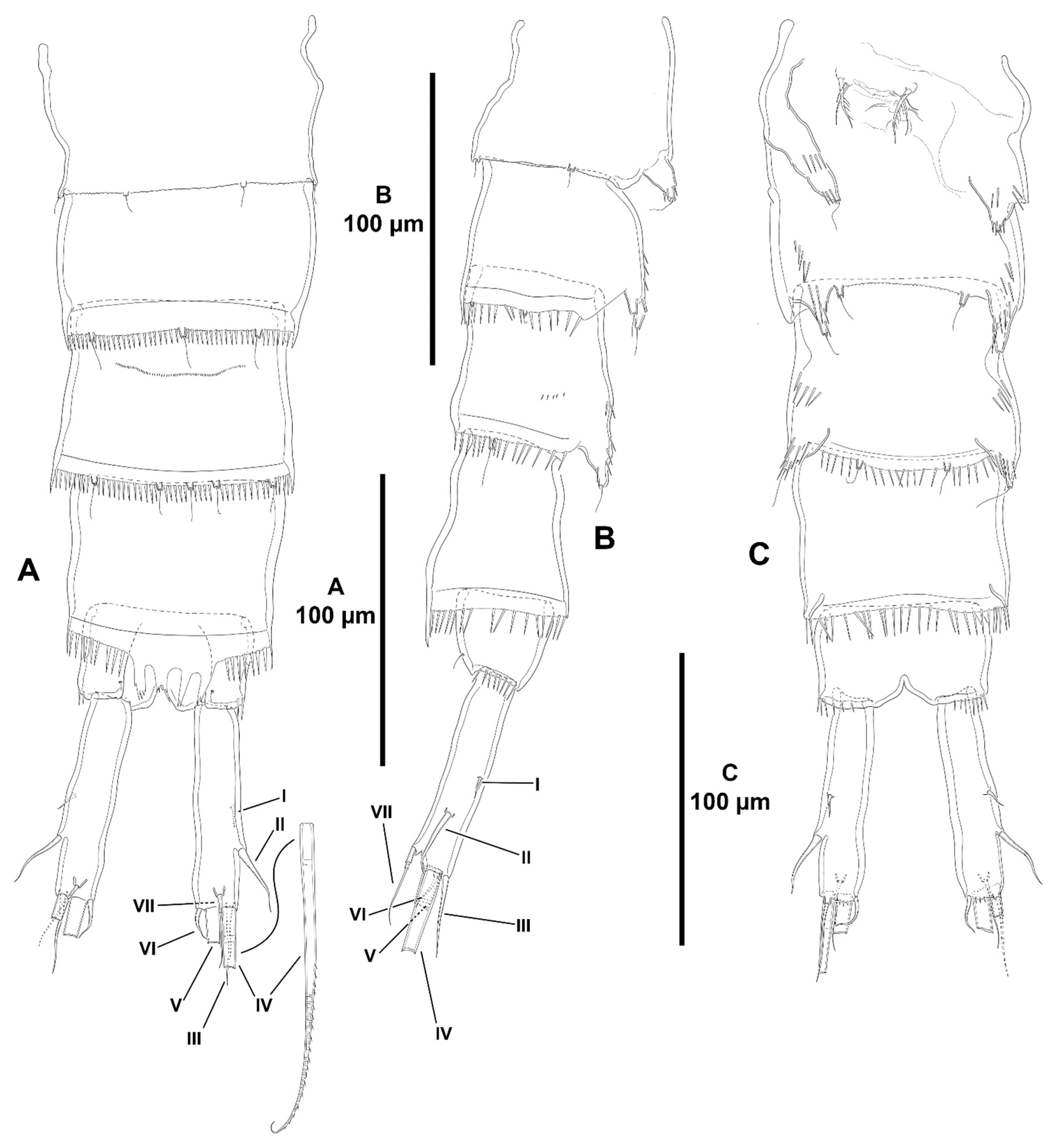
- Characters 70–74 (P5). Seifried [11] showed that the ground pattern of P5 of Zosimeidae includes (i) separated baseoendopods in the females, but fused medially in the males; (ii) P5 EXP separated from the baseoendopod in both sexes; (iii) four setae on the female and male P5 EXP; and (iv) four setae on the endopodal lobe of females, but two setae in males.
- Both baseoendopods of the female P5 are separated throughout the family but became fused medially in Z. xiximi sp. nov. and Z. thistlei. The baseoendopods of both legs are fused medially in the males of most species of the family; both baseoendopods have been described as separate in the males of Z. pacifica, and Z. typica.Acritozosime sigsbeeiensis sp. nov. is the only species known to date in which both the female and male P5 EXP and BENP are separated; P5 EXP and BENP are also separated in the female of A. spinesco, but the male of the species has not been discovered yet. The female P5 EXP and BENP of two species of the incrassata-group of Zosime, Z. incrassata and Z. bathybia, are separated (the males of these species remain unknown). The exopod and baseoendopod of the female P5 of another species of the incrassata-group, Z. carsteni, are imperfectly separated and an anterior suture is still visible; P5 EXP and BENP are completely fused in the male. The exopod and baseoendopod of P5 are separated in the male of Z. pacifica but completely fused in the female. In this regard, Fiers [28] noted that the separation of both rami is most probably suppressed from early copepodid stages onwards in the females of most zosimeids. He [28] observed that the P5 of Z. pacifica appears at the copepodid III as a bud with three setae. The final armature complement of P5 in both the female and male (exopod with three marginal and one surface seta; endopod with two setae) appears at the copepodid IV and both rami undergo further subtle changes in length:width ratio during the molt to copepodid V. The exopod and baseoendopod remain fused in the adult female, but the exopod becomes separated from the baseoendopod in the male during the last molt from copepodid V to copepodid VI (adult). The female P5 EXP and BENP are fused in all other genera and species of the family; the same applies for the males of the species for which they have been described.The pair of fifth legs of the males of all genera and species for which the male is known are fused to the supporting somite. However, both P5 have been illustrated as being separate from the somite in the males of Z. typica and Z. montagnai; this requires confirmation.All the species of Zosime (except for Z. thistlei), and Heterozosime possess the full primitive armature complement of four setae on the female P5 EXP. Loss of one seta of the female P5 EXP—most probably the proximal outer element—occurred convergently in Z. thistlei (which is regarded as autapomorphic within Zosime), in two species of Peresime, Pe. pryorae and Pe. reducta (which is regarded here as a potential synapomorphy), and in the two species of Acritozosime. Further—potentially synapomorphic—reduction to two setae (most probably the proximal outer seta and one marginal element) on the female P5 EXP occurred in Pe. abyssalis and Pe. pryocta sp. nov. The condition of the female P5 EXP of P. browni Scott T., 1912 is uncertain. Scott T. [5] described the female P5 EXP with three marginal setae, but a small outer proximal bristle is present in his figure of the female P5 [5] (plate VIII, Figure 19). The outer proximal seta of the female P5 EXP of most species of Zosime with a tetrasetose ramus is displaced to the anterior surface of the ramus, resulting in three marginal setae and one proximal outer surface element. The four marginal setae on the female P5 EXP of Z. reyssi, Z. incrassata, Z bathybia, Z. carsteni, and Z. montagnai could be due to observational imprecisions, and the displacement of the proximal outer seta of the P5 EXP to a surface position on that ramus is a potential autapomorphy for Zosimeidae (see below).The tetrasetose male P5 EXP is present in most species of Zosime, and in the male of H. tenuis (the male of H. klemie remains undescribed). Secondary loss of one seta (most probably the proximal outer element) occurred in the male of two species of Zosime, Z. gisleni (the female P5 EXP possesses three marginal setae and one proximal outer surface seta) and Z. thistlei (the loss of one seta occurred also in the female), and in the male of A. sigsbeeiensis sp. nov. (the female P5 EXP is also trisetose). Further reduction to two setae on the male P5 EXP occurred in Pe. abyssalis (two setae are present also in the female of the species; the female of Pe. pryocta sp. nov. possess a bisetose P5 EXP, but the male of the species has not been described). Zosime gisleni seems to be unique in that the armature complement of the female and male P5 EXP is different, i.e., the female possesses three marginal setae and one proximal outer surface element, but the male possesses only three marginal setae. The proximal outer element of the male P5 EXP of Z. gisleni was most probably suppressed during the development from copepodid to the adult phase.The primitive tetrasetose baseoendopod of the female P5 is present in several species of Zosime (in two out of eight species of subordinate subgroup Aa (Z. anneae and Z. major), in all the species of subordinate subgroup Ab (Z. atlantica, Z. mediterranea, and Z. xiximi sp. nov.), in three out of four species of subordinate subgroup Ac (Z. eliasi, Z. tunnelli, Z. thistlei), in all the species of subordinate subgroup Bb (Z. paramajor, Z. erythraea, Z. comata, Z. bathyalis, and Z. gymnokosmosa), in two out of four species of the incrassata-group (Z. carsteni and Z. reyssi)), in P. browni, and in H. klemie sp. nov. The—secondary—loss of one seta resulting in a trisetose endopodal lobe occurred in Zosime (Z. gisleni and Z. typica (subordinate subgroup Aa), Z. bergensis (subordinate subgroup Ac), and Z. incrassata and Z. bathybia (incrassata-group)), in the four species of Peresime and in the two species of Acritozosime. Further—apomorphic—reduction to two setae occurred in Zosime (three species of subordinate subgroup Aa (Z. mexicana sp. nov., Z. montagni, and Z. pacifica), in all the species of subordinate subgroup Ba (Z. valida, Z. destituta, and Z. changi) for which is a potential synapomorphy), and in H. tenuis.The male P5 endopodal lobe bears the primitive armature complement of two setae in all genera and species of the family, except for Z. gymnokosmosa with one seta only. Kim and Lee [27] regarded the unisetose endopodal lobe of the male P5 of Z. gymnokosmosa as autapomorphic for the species.
- −
- Characters 86–88 (male P6). The ground pattern of the male P6 of Idyanthidae and Zosimeidae consists of three setae [11]. The innermost seta is very short in all the species within the family, but some differences were observed between the outer—basal—and the apical element. The outer—basal—and the apical—middle—element of Z. montagnai were described (they were not illustrated) as spiniform in Yeom et al. [2] (p. 9). Fiers [28] (p. 19) described the outer basal and the apical element of the male P6 of Z. pacifica as spinulose setae, but these elements look more like spiniform in his figure [28] (p. 20, Figure 4e). Pointner [13] (p. 2314) described the outer element of the male P6 of Z. eliasi as a plumose flexible seta, and the apical element as a bipinnate seta, but the latter was shown as a bipinnate spine in her figure [13] (p. 2313, Figure 20C). Due to these inconsistencies, these species were excluded from the present analysis. The primitive condition with setiform outer and apical elements on the male P6 is still present in Pe. abyssalis (the only species of that genus for which the male has been described), Z. thistlei, Z. carsteni, Z. valida, Z. gymnokosmosa, and Acritozosime sigsbeeiensis sp. nov. (the only species of that genus for which the male has been described). The most primitive condition amongst the above species is present in Pe. abyssalis, Z. thistlei, and Z. carsteni in which the outer and apical setiform elements are subequal in length, or the outer element slightly shorter than the apical seta. The apical seta underwent reduction in Z. valida and Z. gymnokosmosa. The outer seta of A. sigsbeeiensis underwent secondary reduction (it is half as long as the apical seta) and is regarded here as autapomorphic for the species. Heterozosime tenuis is unique within the family in the—potentially autapomorphic—transformation of the outer seta into a pinnate spine. The—apomorphic—transformation of the apical seta into a spine occurred convergently in some species of Zosime (Z. anneae, Z. xiximi sp. nov., and Z. changi), and in H. tenuis.
4.2. The Family Zosimeidae
- (i).
- (ii).
- loss of the lateral idyanthidimorph seta 2 of A2 ENP2 (see Seifried [11] (p. 89, Figure 26; p. 94, Figure 28))
- (iii).
- distal margin of the antennary endopod no beveled
- (iv).
- setae of the mandibular basis inserting on a bulge of inner border
- (v).
- mandibular endopod with one lateral and three apical setae [mandibular endopod one-segmented]
- (vi).
- mandibular exopod with three lateral and one distal seta [mandibular exopod one-segmented]
- (vii).
- (viii).
- exopod of the maxillule very short [with three setae]
- (ix).
- endites of the maxillary syncoxa inserted on distal half of syncoxa
- (x).
- endite of the (allo)basis of maxilla poorly developed
- (xi).
- maxillary endopod one-segmented with 5 setae
- (xii).
- syncoxa of maxilliped subquadrate, at most 1.5 times as long as wide [syncoxa with two setae]
- (xiii).
- maxilliped straight (stenopodial, without geniculation or flexure point between syncoxa and basis, and between basis and endopod)
- (xiv).
- endopod of the maxilliped one-segmented and directed outwardly; the idyanthidimorph first endopodal segment and its armature lost [endopod with two small outer and two geniculate distal setae]
- (xv).
- P1 ENP1 subquadrate; P1 ENP2 and ENP3 fused, with two inner setae and two terminal spines
- (xvi).
- outer basal seta of female and male P5 arising from long setophore
- (xvii).
- female P5 EXP and endopodal lobe separated by incision [endopodal lobe with two inner and two distal setae; exopod with four setae]
- (xviii).
- male P5 BENP very constricted [baseoendopods fused medially]
- (xix).
- male P5 endopodal lobe very small [with two small setae; exopod one-segmented with four setae (1,2,1)]
4.3. Key to the Genera of Zosimeidae
- 1a. P1 EXP three-segmented………………………………………………………………………………………………………………………………………………………………………………………………2
- 1b. P1 EXP two-segmented………………………………………………………………………………………………………………………………………………………………………………………………..4
- 2a. Female A1 six- to eight-segmented; P2 ENP three-segmented……………………………………………………………………………………………………………………………………………………3
- 2b. Female A1 five-segmented; P2 ENP two-segmented…………………………………………………………………………………………………………………………………………………Pseudozosime
- 3a. P3 ENP three-segmented………………………………………………………………………………………………………………………………………………………………………………………. Zosime
- 3b. P3–P4 ENP two-segmented…………………………………………………………………………………………………………………………………………………………………………………...Peresime
- 4a. Female A1 six-segmented; A2 EXP1 and EXP2 with one seta; A2 ENP2 with six distal elements; mandibular basis, EXP and ENP with 3,3,4 setae, respectively; maxillule with epipodal seta; syncoxa of maxilliped unarmed; P1 EXP2 with seven setae in all; P1 ENP1 with one seta, ENP2 with inner lateral seta and two or three distal elements; P2 EXP3 with two inner lateral setae; P2–P4 ENP three-segmented; female P5 EXP with three marginal elements and one outer surface seta; female P5 EXP and BENP fused…………………………………………………………………………………………………………………………………………………………………………………………………………….Heterozosime
- 4b. Female A1 seven-segmented; A2 EXP1 with two setae, EXP2 unarmed; A2 ENP2 with seven distal elements; mandibular basis, EXP and ENP with 2,4,5 setae, respectively; maxillule without epipodal seta; syncoxa of maxilliped armed; P1 EXP2 with six setae in all; P1 ENP1 unarmed, ENP2 without inner lateral seta and two distal elements; P2 EXP3 with one inner lateral seta; P2–P4 ENP two-segmented; female P5 EXP with three marginal elements (without outer surface seta); female P5 EXP and BENP separated………………………………………………………………………………………………………………………………………………………………………………………………………..Acritozosime
4.4. The Genus Zosime
4.5. Key to the Species of Zosime
- 1a. A2 EXP2 unarmed (except for Z carsteni with one seta), EXP3 without lateral seta (except for Z. carsteni with one lateral element); mandibular endopod with less than four setae (except for Z. carsteni with four setae); caudal rami short (length:width ratio < 1.6); outer spines of exopodal segments with homogeneous small spinules ………………………………….. 2 incrassata group
- 1b. A2 EXP2 always with one seta, EXP3 with one lateral seta and two or three distal elements (except for Z. atlantica and Z. xiximi sp. nov. without lateral seta); mandibular endopod with four setae; caudal rami elongate (length:width ratio > 1.6); at least one outer spine of exopodal segments with heterogeneous subdistal long spinules………………………………. 5 typica group
- 2a. Armature formula of A2 EXP 1,1,3 (EXP3 with one lateral and two distal elements); mandibular palp with trisetose basis, unisetose small, one-segmented exopod, and tetrasetose one-segmented endopod; female P5 EXP and BENP fused but with anterior suture between BENP and EXP, ENP with four setae………………………………………………………………….Z. carsteni
- 2b. Armature formula of A2 EXP 1,0,2–3 (EXP3 without lateral seta and with two or three distal elements); mandibular palp either with unisetose basis, exopod incorporated to basis and represented by one seta, and endopod with two setae (palp with less than four setae in all), or palp small, one-segmented with one seta; female P5 EXP and BENP fused or separated, ENP with three or four setae…………………………………………………………………………………………………………………………………………………………………………………………………….3
- 3a. Female A1 six-segmented; armature formula of A2 EXP 1,0,2; mandibular palp small, one-segmented, with one seta; P1 ENP2 without inner lateral seta and with two distal elements; P3 ENP3 with one inner lateral seta, and one inner distal, one distal, and one outer distal element; female P5 and BENP fused, ENP with four setae…………………………………………………………………………………………………………………………………………………………………………………………………………………..Z. reyssi
- 3b. Female A1 seven-segmented; armature formula of A2 EXP 1,0,2–3; mandibular palp with unisetose basis, exopod incorporated to basis and represented by one seta, endopod with two setae; P1 ENP2 with one inner lateral seta, and one inner distal, one distal, and one outer distal element; P3 ENP3 with two inner lateral setae, and one inner distal, one distal, and one outer distal element; female EXP and BENP separated, ENP with three setae ……………………………………………………………………………………………………………………………………………4
- 4a. A2 EXP3 with two distal setae……………………………………………………………………………………………………………………………………………………………………………Z. incrassata
- 4b. A2 EXP3 with three distal setae……………………………………………………………………………………………………………………………………………………………………………Z. bathybia
- 5a. Armature formula of P1 EXP 1,2,3 (with three outer spines)………………………………………………………………………………………………………………………………………6 subgroup A
- 5b. Armature formula of P1 EXP 1,2,2 (with two outer spines)……………………………………………………………………………………………………………………………………….20 subgroup B
- 6a. P1 ENP2 with four setae in all (one inner, one distal inner, one distal, and one outer distal seta)…………………………………………………………………………...7 subordinate subgroup Aa
- 6b. Armature complement of P1 ENP not like this…………………………………………………………………………………………………………………………………...14 subordinate subgroups Ab
- 7a. Middle apical seta of P4 ENP3 reduced……………………………………………………………………………………………………………………………………………………………………………..8
- 7b. Middle apical seta of P4 ENP3 well-developed…………………………………………………………………………………………………………………………………………………………………..10
- 8a. Female A1 six-segmented, ancestral segments II and III separated, proximal aesthetasc on segment 4, ancestral segments IV and V separated; maxillule with two epipodal setae………………………………………………………………………………………………………………………………………………………………………………………………………………..Z. pacifica
- 8b. Female A1 seven-segmented, ancestral segments II and III fused, proximal aesthetasc on segment 3, ancestral segments IV and V fused; maxillule with one epipodal seta…………………..9
- 9a. A2 EXP 3 with one lateral and three distal setae……………………………………………………………………………………………………………………………………………..Z. mexicana sp. nov.
- 9b. A2 EXP3 with one lateral and two distal setae………………………………………………………………………………………………………………………………………………………..Z. montagnai
- 10a. Female A1 eight-segmented; female P5 ENP with four setae………………………………………………………………………………………………………………………………………….Z. anneae
- 10b. Female A1 six- or seven-segmented; female P5 ENP with three or four setae………………………………………………………………………………………………………………………………11
- 11a. Female A1 seven-segmented; setal complement of female P5 ENP variable from three to four elements…………………………………………………………………………………..Z. paratypica
- 11b. Female A1 six-segmented………………………………………………………………………………………………………………………………………………………………………………………… 12
- 12a. Female P5 ENP with four setae……………………………………………………………………………………………………………………………………………………………………………..Z. major
- 12b. Female P5 ENP with three setae…………………………………………………………………………………………………………………………………………………………………………………..13
- 13a. Male P5 EXP with four setae in all (three marginal, 1 surface element)………………………………………………………………………………………………………………………………Z. typica
- 13b. Male P5 EXP with three setae in all (without surface element)………………………………………………………………………………………………………………………………………..Z. gisleni
- 14a. P1 ENP2 without inner lateral seta, with tree distal elements (inner distal, distal, and outer distal)……………………………………………………………………..15 subordinate subgroup Ab
- 14b. P1 ENP2 with inner lateral seta, and two distal elements………………………………………………………………………………………………………………………17 subordinate subgroup Ac
- 15a. Female A1 six-segmented; baseoendopods of both pairs of female P5 separated……………………………………………………………………………………………………………Z. mediterranea
- 15b. Female A1 seven- or eight-segmented; baseoendopods of both pairs of female P5 fused medially or separated………………………………………………………………………………………16
- 16a. Female A1 seven-segmented; A2 ENP2 with five elements distally; mandibular exopod with four setae; maxillule with two epipodal setae; baseoendopods of both pairs of female P5 separated ………………………………………………………………………………………………………………………………………………………………………………………………………...Z. atlantica
- 16b. Female A1 eight-segmented; A2 ENP2 with six elements distally; mandibular exopod with two setae; maxillule with one epipodal seta; baseoendopods of both pairs of female P5 fused medially ………………………………………………………………………………………………………………………………………………………………………………………………….Z. xiximi sp. nov.
- 17a. Female antennule seven-segmented, ancestral segments IV and V fused; A2 EXP3 with one lateral and three distal setae; A2 ENP1 unarmed; P1 ENP1 unarmed; female and male P5 EXP with three marginal setae (without surface outer element); baseoendopods of both pairs of female P5 fused medially……………………………………………………………………………Z. thistlei
- 17b. Segmentation of female antennule different, other characters not combined……………………………………………………………………………………………………………………………….18
- 18a. Female A1 eight-segmented; syncoxa of maxilliped with one seta……………………………………………………………………………………………………………………………………..Z. eliasi
- 18b. Female A1 six-segmented; syncoxa of maxilliped unarmed…………………………………………………………………………………………………………………………………………………..19
- 19a. Endopod of maxilliped with three elements; P2 ENP3 with four elements in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element); outer exopodal spines of P1 with heterogeneous subdistal long spinules; female P5 ENP with three setae…………………………………………………………………………………………………………..Z. bergensis
- 19b. Endopod of maxilliped with four elements; P2 ENP3 with three elements in all (without inner lateral armature); heterogeneous subdistal long spinules present only on the distalmost outer spine of P1 EXP; female P5 ENP with four setae……………………………………………………………………………………………………………………………………………………...Z. tunnelli
- 20a. P1 ENP2 without inner lateral seta, with three distal elements (inner distal, distal, and outer distal element)…………………………………………………………………………………………21
- 20b. P1 ENP2 with inner lateral seta and two distal elements………………………………………………………………………………………………………………………………………………………23
- 21a. P2–P4 EXP3 with two outer spines………………………………………………………………………………………………………………………………………………………………………...Z. changi
- 21b. P2–P4 EXP3 with three outer spines………………………………………………………………………………………………………………………………………………………………………………22
- 22a. Female A1 seven-segmented; maxillule with two epipodal setae; P4 ENP3 with four seta in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element)……………………………………………………………………………………………………………………………………………………………………………………………………………..Z. valida
- 22b. Female A1 six-segmented; maxillule with one epipodal seta; P4 ENP3 with three seta in all (without inner lateral armature)……………………………………………………………Z. destituta
- 23a. P2–P4 EXP3 with three outer spines……………………………………………………………………………………………………………………………………………………………………………....24
- 23b. P2–P4 EXP3 with two outer spines………………………………………………………………………………………………………………………………………………………………………………..25
- 24a. P1 EXP2 with one inner seta…………………………………………………………………………………………………………………………………………………………………………….Z. paramajor
- 24b. P1 EXP2 unarmed…………………………………………………………………………………………………………………………………………………………………………………………Z. erythraea
- 25a. Female A1 six-segmented; P2–P3 ENP3 with or without inner lateral seta but with three distal elements (inner distal, distal, and outer distal element); P4 ENP3 with two (one inner lateral and one distal seta) or three setae in all (without inner lateral seta, and with three distal elements)………………………………………………………………………………………………….26
- 25b. Female A1 eight-segmented; P2–P4 ENP3 with one inner lateral seta and two distal elements………………………………………………………………………………………….Z. gymnokosmosa
- 26a. P2–P3 ENP3 with four elements in all (one inner lateral seta, and one inner distal, one distal, and one outer distal element); P4 ENP3 with two elements in all (one inner lateral and one distal seta)………………………………………………………………………………………………………………………………………………………………………………………………………….Z. comata
- 26b. P2–P4 ENP3 with three elements in all (without inner lateral seta)………………………………………………………………………………………………………………………………...Z. bathyalis
4.6. The Genera Peresime and Pseudozosime
4.7. Key to the Species of Peresime
- 1a. Female A1 six-segmented; A2 ENP2 with five or six distal elements……………………………………………………………………………………………………………………………………………2
- 1b. Female A1 eight-segmented; A2 ENP2 with six or seven distal elements………………………………………………………………………………………………………………………………………3
- 2a. A2 EXP3 with four setae; A2 ENP2 with five distal elements; mandibular exopod with four setae; maxillule without epipodal seta; P1 ENP2 with three setae in all; female P5 EXP with three setae in all…………………………………………………………………………………………………………………………………………………………………………………………………..Pe reducta
- 2b. A2 EXP3 with three setae; A2 ENP2 with six distal elements; mandibular exopod with three setae; maxillule with one epipodal seta; P1 ENP2 with two setae in all; female P5 EXP with two setae in all………………………………………………………………………………………………………………………………………………………………………………………………….Pe. abyssalis
- 3a. A2 ENP2 with six distal setae; mandibular exopod with three setae; maxillule without epipodal seta; basis of maxilliped with one seta; female P5 EXP with three setae………………………………………………………………………………………………………………………………………………………………………………………………………………Pe. pryorae
- 3b. A2 ENP2 with seven distal setae; mandibular exopod with four setae; maxillule with one epipodal seta; basis of maxilliped unarmed; female P5 EXP with two setae……………………………………………………………………………………………………………………………………………………………………………………………………..Pe. pryocta sp. nov.
4.8. The Genus Heterozosime
4.9. Key to the Species of Heterozosime
- 1a. P1 ENP2 with four setae in all (one inner lateral and three distal elements); P2–P4 EXP3 with six elements in all (with two outer spines); medial apical seta on P3 ENP3 reduced; female P5 ENP with two setae………………………………………………………………………………………………………………………………………………………………………………………………..H. tenius
- 1b. P1 ENP2 with three setae in all (one inner lateral and two distal elements); P2–P4 EXP3 with seven elements in all (with three outer spines); medial apical seta on P3 ENP3 well-developed; female P5 ENP with four setae…………………………………………………………………………………………………………………………………………………………..H. klemie sp. nov.
4.10. The Genus Acritozosime
4.11. Key to the Species of Acritozosime
- 1a. A2 ENP1 without inner seta; maxilliped geniculate, syncoxa with two setae; P2–P3 EXP1 with one inner seta………………………………………………………………………………A. spinesco
- 1b. A2 ENP1 with inner seta; maxilliped non-geniculate, syncoxa with one seta; P2–P3 EXP1 unarmed……………………………………………………………………………A. sigsbeeiensis sp. nov.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kim, J.G.; Lee, J. Proposal of two new genera of the family Zosimeidae Seifried (Copepoda: Harpacticoida): Heterozosime gen. nov. and Acritozosime gen. nov. Eur. J. Taxon. 2021, 760, 32–60. [Google Scholar] [CrossRef]
- Yeom, J.; Rohal Lupher, M.; Lee, W. Four new species of Zosimeidae (Copepoda: Harpacticoida) from the southwestern Gulf of Mexico. Diversity 2022, 14, 198. [Google Scholar] [CrossRef]
- Dinet, A. Espèces nouvelles de copépodes harpacticoïdes (Crustacea) des sédiments profonds de la dorsale de Walvis. Arch. Zool. Expérimentale Générale 1974, 115, 549–577. [Google Scholar]
- Becker, K.-H.; Schriever, G. Eidonomie und Taxonomie abyssaler Harpacticoidea (Crustacea, Copepoda) Teil III. 13 Neue Tiefsee-Copepoda Harpacticoidea der Familien Canuellidae, Crviniidae, Tisbidae, Thalestridae, Diosaccidae und Ameiridae. “Meteor” Forschungsergebnisse 1979, Reihe D-Bi, 38–62. [Google Scholar]
- Scott, T. The Entomostraca of the Scottish National Antarctic Expedition, 1902–1904. Trans. Linn. Soc. Edinburgh. 1912, XLVIII, 521–599. [Google Scholar] [CrossRef]
- Gómez, S.; Rivera-Sánchez, K.I. The Deep-Water Benthic Harpacticoida (Copepoda) of the Americas. In Deep-Sea Pycnogonids and Crustaceans of the Americas; Hendrickx, M.E., Ed.; Springer: Cham, Switzerland, 2020; pp. 47–125. ISBN 978-3-030-58409-2. [Google Scholar]
- Koller, S.; George, K.H. Description of a new species of Zosime Boeck, 1872 (Copepoda: Harpacticoida: Zosimeidae) from the Great Meteor Seamount, representing one of the few eurybathic Harpacticoida among the distinct plateau and deep-sea assemblages. Meiofauna Mar. 2011, 19, 109–126. [Google Scholar]
- Kitahashi, T.; Kawamura, K.; Kojima, S.; Shimanaga, M. Assemblages gradually change from bathyal to hadal depth: A case study on harpacticoid copepods around the Kuril Trench (north-west Pacific Ocean). Deep. Res. Part I Oceanogr. Res. Pap. 2013, 74, 39–47. [Google Scholar] [CrossRef]
- Kitahashi, T.; Kawamura, K.; Kojima, S.; Shimanaga, M. Bathymetric Patterns of α and β diversity of harpacticoid copepods at the genus level around the Ryukyu Trench, and turnover diversity between trenches around Japan. Prog. Oceanogr. 2014, 123, 54–63. [Google Scholar] [CrossRef]
- Gómez, S.; Morales-Serna, F.N. Updated checklist of published and unpublished records of harpacticoid copepods (Crustacea: Copepoda: Harpacticoida) from Mexico. Proc. Biol. Soc. Washingt. 2014, 127, 99–121. [Google Scholar] [CrossRef]
- Seifried, S. Phylogeny of Harpacticoida (Copepoda): Revision of “Maxillipedasphalea” and Exanechentera; Cuvillier Verlag: Göttingen, Germany, 2003. [Google Scholar]
- Kim, J.G.; Jung, T.W.; Yoon, S.M. A new species of Zosime (Copepoda: Harpacticoida: Zosimeidae) from shallow subtidal habitats in Korea, with notes on the genus. Zootaxa 2016, 4136, 323–334. [Google Scholar] [CrossRef]
- Pointner, K. Description of two new species of Zosime (Copepoda: Harpacticoida: Zosimeidae), including remarks on its phylogeny and distribution on the Great Meteor Seamount Plateau (north-east Atlantic Ocean). J. Nat. Hist. 2017, 51, 2283–2330. [Google Scholar] [CrossRef]
- Yoder, M.; De Ley, I.T.; King, I.W.; Mundo-Ocampo, M.; Mann, J.; Blaxter, M.; Poiras, L.; De Ley, P. DESS: A versatile solution for preserving morphology and extractable DNA of nematodes. Nematology 2006, 8, 367–376. [Google Scholar] [CrossRef]
- Somerfield, P.J.; Warwick, R.M. Meiofauna in Marine Pollution Monitoring Programmes. A Laboratory Manual; Ministry of Agriculture, Fisheries and Food Directorate of Fisheries Research: Great Britain, UK, 1996.
- de Jonge, V.N.; Bouwman, L.A. A simple density separation technique for quantitative isolation of meiobenthos using the colloidal silica Ludox-TM. Mar. Biol. 1977, 42, 143–148. [Google Scholar] [CrossRef]
- Huys, R.; Boxshall, G.A. Copepod Evolution; The Ray Society: London, UK, 1991. [Google Scholar]
- Sewell, R.B.S. The littoral and semi-parasitic Cyclopoida, the Monstrilloida and Notodelphyoida. Sci. Reports John Murray Exped. Publ. Br. Museum Natural Hist. 1949, 9, 17–199. [Google Scholar]
- Lang, K. Marine Harpacticiden von der Campbell-Insel und einigen anderen südlichen Inseln. Acta Univ. Lund. New Ser. Avd. 2 Kungl. Fysiogr. Sällskapets Handl. 1934, 30, 1–56. [Google Scholar]
- Felsenstein, J. PHYLIP (Phylogeny Inference Package), Version 3.6; Department of Genome Sciences at University of Washington: Washington, DC, USA, 2005. Available online: http://evolution.genetics.washington.edu/phylip.html (accessed on 22 January 2023).
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogeny. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive tree of life (ITOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, W1, W293–W296. [Google Scholar] [CrossRef]
- ICZN (International Commission on Zoological Nomenclature). International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- Bodin, P. Copépodes Harpacticoïdes des etages bathyal et abyssal du Golfe de Gascogne. Mémoires du Muséum Natl. d´Histoire Nat. Sér. 1968, 55, 1–107. [Google Scholar]
- Monard, A. Les harpacticoïdes marins de la région d’Alger et de Castiglione. Bull. la Stn. d’Aquiculture Pêche Castiglione 1937, 1935, 9–93. [Google Scholar]
- Kornev, P.N.; Chertoprud, E.C. Copepod Crustaceans of the Order Harpacticoida of the White Sea: Morphology, Systematics, Ecology; Biology Faculty, M.S.U., Ed.; Tovarishchestvo Nauchnikh Izdanii KMK: Moscow, Russia, 2008. [Google Scholar]
- Kim, J.G.; Lee, J. New records of benthic harpacticoids from Korea: I. Three new species of the genus Zosime Boeck (Copepoda: Harpacticoida: Zosimeidae). Mar. Biol. Res. 2019, 15, 210–239. [Google Scholar] [CrossRef]
- Fiers, F. Three new harpacticoid copepods from the Santa Maria Basin off the Californian Pacific coast (Copepoda, Harpacticoida). Beaufortia 1991, 42, 13–47. [Google Scholar]
- Huys, R.; Clark, P.F. Case 3467 Zosimidae Seifried, 2003 (Crustacea, Copepoda, Harpacticoida): Proposed emendation of spelling to Zosimeidae to remove homonymy with Zosiminae Alcock, 1898 (Crustacea, Decapoda, Xanthidae). Bull. Zool. Nomencl. 2009, 66, 24–29. [Google Scholar] [CrossRef] [Green Version]
- Mu, F.H.; Somerfield, P.J.; Warwick, R.M.; Zhang, Z.N. Large-scale spatial patterns in the community structure of benthic harpacticoid copepods in the Bohai Sea, China. Raffles Bull. Zool. 2002, 50, 17–26. [Google Scholar]
- Ma, L.; Li, X. Benthic harpacticoid copepods of Jiaozhou Bay, Qingdao. Chinese J. Oceanol. Limnol. 2017, 35, 1127–1133. [Google Scholar] [CrossRef]
- Chertoprud, E.S.; Gómez, S.; Gheerardyn, H. Harpacticoida (Copepoda) fauna and the taxocene diversity of the South China Sea. Oceanology 2009, 49, 488–498. [Google Scholar] [CrossRef]
- Shimanaga, M.H.K.; Shirayama, Y. Temporal patterns in diversity and species composition of deep-sea benthic copepods in bathyal Sagami Bay, Central Japan. Mar. Biol. 2004, 144, 1097–1110. [Google Scholar]
- George, K.H.; Pointner, K.; Packmor, J. The benthic Copepoda (Crustacea) of Anaximenes Seamount (eastern Mediterranean Sea)—Community structure and species distribution. Prog. Oceanogr. 2018, 165, 299–316. [Google Scholar] [CrossRef]

































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Character | State | |
|---|---|---|
| 1 | Female antennule, number of segments | 9(0),8(1),7(2),6(3),5(4) |
| 2 | Female antennule, ancestral segments II and III | separated(0),fused(2) |
| 3 | Female antennule, ancestral segments II–IV | separated(0),fused(1) |
| 4 | Female antennule, ancestral segments III and IV | separated(0),fused(1) |
| 5 | Female antennule, ancestral segments IV and V | separated(0),fused(1) |
| 6 | A2 EXP, number of segments | 4(0),3(1),2(2),1(3) |
| 7 | A2 EXP1, number of setae | 2(0),1(1) |
| 8 | A2 EXP2, seta when EXP three-segmented | 1(0),0(1) |
| 9 | A2 EXP3, lateral seta when EXP three-segmented | 1(0),0(1) |
| 10 | A2 EXP3, number of distal setae when EXP three-segmented | 3(0),2(1) |
| 11 | A2 ENP1, inner seta | 1(0),0(1) |
| 12 | A2 ENP2, number of lateral elements | 4(0),3(1) |
| 13 | A2 ENP2, shape of seta 4 | long(0),short(1) |
| 14 | A2 ENP2, number of distal setae | 7(0),6(1) |
| 15 | Mandibular palp | three-segmented(0),one-segmented(1) |
| 16 | Mandibular basis, number of setae | 4(0),3(1),2(2),1(3),0(4) |
| 17 | Mandibular exopod, number of segments | 2(0),1(1),0(2) |
| 18 | Mandibular exopod, number of setae when EXP one-segmented | 4(0),3(1),2(2),1(3) |
| 19 | Mandibular endopod, number of setae | 10(0),9(1),8(2),5(3),4(4),2(5) |
| 20 | Maxillulary coxa, number of epipodal setae | 2(0),1(1),0(2) |
| 21 | Maxillary endopod, number of segments | 3(0),2(1),1(2) |
| 22 | Maxilliped, shape | subchelate(0),stenopodial geniculate(1),stenopodial non-geniculate(2) |
| 23 | Maxillipedal syncoxa, number of setae | 2(0),1(1),0(2) |
| 24 | Maxillipedal basis, number of setae | 1(0),0(1) |
| 25 | Maxillipedal ENP, number of segments | 2(0),1(1) |
| 26 | Maxillipedal ENP, number of elements when ENP one-segmented | 4(0),3(1),2(2) |
| 27 | Maxilliped ENP2, number of elements when ENP two-segmented | 4(0),2(1) |
| 28 | Caudal rami, l:w ratio | subquadrate(0),elongate(1) |
| 29 | P1 EXP, number of segments | 3(0),2(1) |
| 30 | P1 EXP2, inner seta when EXP three-segmented | present(0),absent(1) |
| 31 | P1 EXP3, number of outer spines when EXP three-segmented | 3(0),2(1) |
| 32 | P1 EXP2, number of inner setae when EXP two-segmented | 2(0),1(1) |
| 33 | P1 ENP, number of segments | 3(0),2(1) |
| 34 | P1 ENP1, inner seta | present(0),absent(1) |
| 35 | P1 ENP2, number of inner lateral setae when ENP two-segmented | 1(0),0(1) |
| 36 | P1 ENP2, number of distal setae when ENP two-segmented | 3(0),2(1),1(2) |
| 37 | P1 ENP3, number of inner lateral setae when ENP three-segmented | 2(0),0(1) |
| 38 | P2 EXP1, number of inner setae | 1(0),0(1) |
| 39 | P2 EXP3, number of inner setae | 2(0),1(1) |
| 40 | P2 EXP3, number of outer spines | 3(0),2(1) |
| 41 | P2 ENP, number of segments | 3(0),2(1) |
| 42 | P2 ENP1, number of inner setae | 1(0),0(1) |
| 43 | P2 ENP2, number of inner setae when ENP three-segmented | 2(0)1(1) |
| 44 | P2 ENP3, number of inner lateral setae when ENP three-segmented | 2(0),1(1),0(2) |
| 45 | P2 ENP3, number of distal setae when ENP three-segmented | 3(0),2(1) |
| 46 | P2 ENP2, number of inner lateral setae when ENP two-segmented | 2(0),0(1) |
| 47 | P2 ENP2, number of distal setae when ENP two-segmented | 3(0),2(1) |
| 48 | P3 EXP1, number of inner setae | 1(0),0(1) |
| 49 | P3 EXP3, number of inner setae | 3(0),2(1) |
| 50 | P3 EXP3, number of outer spines | 3(0),2(1) |
| 51 | P3 ENP, number of segments | 3(0),2(1) |
| 52 | P3 ENP1, number of inner setae | 1(0),0(1) |
| 53 | P3 ENP2, number of inner setae when ENP three-segmented | 2(0),1(1) |
| 54 | P3 ENP3, number of inner lateral setae when ENP three-segmented | 3(0),2(1),1(2),0(3) |
| 55 | P3 ENP3, number of distal setae when ENP three-segmented | 3(0),2(1) |
| 56 | P3 ENP2, number of inner lateral setae when ENP two-segmented | 2(0),1(1) |
| 57 | P3 ENP2, number of distal setae when ENP two-segmented | 3(0),2(1) |
| 58 | P4 EXP1, number of inner setae | 1(0),0(1) |
| 59 | P4 EXP3, number of inner setae | 3(0),2(1) |
| 60 | P4 EXP3, number of outer spines | 3(0),2(1) |
| 61 | P4 ENP, number of segments | 3(0),2(1) |
| 62 | P4 ENP2, number of inner setae when ENP three-segmented | 2(0),1(1) |
| 63 | P4 ENP3, number of inner lateral setae when ENP three-segmented | 2(0),1(1),0(2) |
| 64 | P4 ENP3, number of distal setae when ENP three-segmented | 3(0),2(1),1(2) |
| 65 | P4 ENP2, number of distal setae when ENP two-segmented | 2(0),1(1) |
| 66 | P1 EXP, ornamentation of outer pines | heterogeneous(0), homogeneous(1) |
| 67 | P2 EXP, ornamentation of outer spines | heterogeneous(0), homogeneous(1) |
| 68 | P3-P4 EXP, ornamentation of outer spines | heterogeneous(0), homogeneous(1) |
| 69 | P4 ENP3 middle apical seta, length when three apical setae | well-developed(0),short, reduced(1) |
| 70 | P5 EXP, female, number of setae | 5(0),4(1),3(2),2(3) |
| 71 | P5 EXP, female, surface seta when four setae | absent(0),present(1) |
| 72 | P5 ENP, female, number of setae | 4(0),3(1),2(2) |
| 73 | P5 BENP’s, female | separated(0),fused(1) |
| 74 | P5 EXP and BENP, female | separated(0),anterior suture(1),fused(2) |
| 75 | P2 ENP, male, number of segments | 3(0),2(1) |
| 76 | P2 ENP1, male, number of inner setae | 1(0),0(1) |
| 77 | P2 ENP2, male, number of setae when ENP three-segmented | 2(0),1(1) |
| 78 | P2 ENP, distal element, number of setae | 1(0),0(1) |
| 79 | P5, male | separated from somite(0),fused to somite(1) |
| 80 | P5 BENP’s, male | separated(0),fused(1) |
| 81 | P5 EXP and BENP, male | separated(0),fused(1) |
| 82 | P5 EXP, male, number of segments when EXP distinct | 2(0),1(1) |
| 83 | P5 EXP, male, number of setae when of last segment, when one-segmented or absorbed into baseoendopod | 4(0),3(1),2(2) |
| 84 | P5 EXP, male, surface seta when four setae | absent(0),present(1) |
| 85 | P5 ENP, male, number of setae | 2(0),1(1) |
| 86 | P6, male, outermost—basal—seta, shape | setiform(0),spiniform(1) |
| 87 | P6, male, apical—middle—seta, shape | setiform(0),spiniform(1) |
| 88 | P6, male, innermost seta, length | well-developed(0),short, reduced(1) |
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | I-0;I-1;III,I1,1 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 |
| ENP | 0-1;I,I,1 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I2,1 |
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | I-0;I-1;III,2,1 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 |
| ENP | 0-1;0,I1,2 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I1,1 |
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | I-0;I-1;III,I1,1 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 |
| ENP | 0-1;0,I1,1 | 0-1;0-1;0,I1,1 | 0-1;0,I1,2 | 0-1;0,1,1 |
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | I-0;III,2,2 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 | I-1;I-1;III,I1,2 |
| ENP | 0-1;0,I1,1 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I2,1 | 0-1;0-1;0,I2,1 |
| P1 | P2 | P3 | P4 | |
|---|---|---|---|---|
| EXP | I-0;III,I1,1 | I-0;I-1;II,I1,1 | I-0;I-1;II,I1,2 | I-1;I-1;II,I1,2 |
| ENP | 0-0,0,I1,0 | 0-0;0,I1,0♀ 0-0;0,Apo1,0♂ | 0-0;0,2,1 | 0-1;0,2,1 |
| Apo, apophysis | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gómez, S.; Yáñez-Rivera, B. On New Species of Three Genera of Zosimeidae Seifried, 2003 (Copepoda: Harpacticoida) from the Deep Sea of the Gulf of California and Gulf of Mexico, with Notes on the Phylogeny of the Family and on the Species Groups of Zosime Boeck, 1873. Diversity 2023, 15, 363. https://doi.org/10.3390/d15030363
Gómez S, Yáñez-Rivera B. On New Species of Three Genera of Zosimeidae Seifried, 2003 (Copepoda: Harpacticoida) from the Deep Sea of the Gulf of California and Gulf of Mexico, with Notes on the Phylogeny of the Family and on the Species Groups of Zosime Boeck, 1873. Diversity. 2023; 15(3):363. https://doi.org/10.3390/d15030363
Chicago/Turabian StyleGómez, Samuel, and Beatriz Yáñez-Rivera. 2023. "On New Species of Three Genera of Zosimeidae Seifried, 2003 (Copepoda: Harpacticoida) from the Deep Sea of the Gulf of California and Gulf of Mexico, with Notes on the Phylogeny of the Family and on the Species Groups of Zosime Boeck, 1873" Diversity 15, no. 3: 363. https://doi.org/10.3390/d15030363
APA StyleGómez, S., & Yáñez-Rivera, B. (2023). On New Species of Three Genera of Zosimeidae Seifried, 2003 (Copepoda: Harpacticoida) from the Deep Sea of the Gulf of California and Gulf of Mexico, with Notes on the Phylogeny of the Family and on the Species Groups of Zosime Boeck, 1873. Diversity, 15(3), 363. https://doi.org/10.3390/d15030363